Introduction
Head and neck squamous cell carcinoma (HNSCC) is an
aggressive life-threatening disease that constitutes 90% of head
and neck carcinoma (HNC). HNSCC represents 3.5% of all cancers, and
is one of the most common type of cancer worldwide despite
significant progress in both early diagnosis and therapy. HNC
originates from the mucosal epithelia of the upper aerodigestive
tract, including the tongue, lip, salivary glands, sinuses, oral
cavity, pharynx, larynx and thyroid (1,2).
Over the past few years, although advanced therapies have been
applied for the treatment of HNC, the 5-year survival rate has not
increased significantly (3,4).
Thus, there is an urgent need for the identification of new
molecules with antitumor activity and less therapeutic toxicity
than the currently used drugs and which are, at the same time,
effective against HNC. In this regard, the identification and
development of naturally occurring compounds may greatly contribute
to this request.
S-Adenosyl-L-methionine (AdoMet, also known as SAM)
is a widely naturally-occurring sulfonium compound that plays a
primary role in cellular metabolism, since it is involved in a
variety of important biochemical processes being indeed, the link
to three key metabolic pathways: transmethylation, polyamine
synthesis and transsulfuration (5-7).
Over the past few decades, a number of in vitro and in
vivo studies have demonstrated the involvement of AdoMet in
various cellular processes, including proliferation,
differentiation, cell cycle regulation and apoptosis, demonstrating
that the sulfonium compound exerts pleiotropic effects on signal
transduction in a variety of cell types and that AdoMet is able to
halt the progression of several human tumors (8-13).
The development of metastases is a multistep process
that requires active and specifically localized extracellular
proteolysis, as well as the activation of a series of physiological
and biochemical processes that govern the migration from the
primary tumor site, invasion through the basement membrane, the
entry of metastatic cells into blood vessels and finally,
localization to the second site (14). Despite significant progress
regarding potential therapeutic targets aimed at improving
survival, the median time to mortality for patients affected by
metastatic HNSCC is approximately 4 months (15). Therefore, the development of novel
strategies aimed at preventing the migration and extracellular
invasion of HNSCC is urgently required.
There is emerging evidence to document the
involvement of AdoMet in the regulation of genes responsible for
cell invasion and metastasis (16-19)
and several research groups have investigated in depth the
epigenetic regulation induced by AdoMet on the methylation status
of genes involved in invasion and metastases processes, including
the urokinase-type plasminogen activator (uPA) and matrix
metalloproteinases (MMPs) (16,19).
It has been demonstrated that the treatment of highly invasive
MDA-231 breast cancer cells and PC-3 prostate cancer cells with
AdoMet, significantly inhibits uPA and MMP2 expression, resulting
in the potent inhibition of tumor cell invasion in vitro and
tumor growth and metastasis in vivo (16,17).
Furthermore, Chik et al demonstrated that AdoMet synergizes
with the DNA methylation inhibitor, 5-aza-2-deoxycytidine, to
suppress uPA expression, thereby blocking MDA-MB-231 cell
invasiveness (18). Another study
demonstrated that in the highly invasive SW-620 colorectal cancer
cell line, treatment with the sulfonium compound induced the
inhibition of MMP2, and a decrease in membrane type 1 matrix
metalloproteinase mRNA levels together with the upregulation of the
tissue inhibitor of MMP2 (19). It
was recently demonstrated that in human LM-7 and MG-63 osteosarcoma
cells, AdoMet treatment led to a dose-dependent decrease in the
proliferation and invasiveness of the tumor cells by inhibiting the
expression of genes involved in the formation of metastasis,
angiogenesis and cellular invasion, including uPA, MMP2 and MMP9
(20). More recently, it was
reported that AdoMet was able to enhance the anti-metastatic effect
of gemcitabine in pancreatic cancer through the inhibition of the
JAK2/STAT3 pathway (21). In
addition, and in association with selenium compounds in human
cervical cancer HeLa cells, AdoMet was shown to inhibit cell
proliferation, migration and adhesion by affecting the ERK and AKT
signaling pathways (22).
In light of these data, the present study
investigated the effects of AdoMet on cell proliferation, migration
and invasion in HNSCC, and also aimed to elucidate the underlying
mechanisms. The findings of this study demonstrate that AdoMet
potently inhibits the migration and invasion of two different types
of HNSCC cells, oral Cal-33 and laryngeal JHU-SCC-011 cells through
the modulation of the AKT, β-catenin and SMAD signaling pathways.
Moreover, the synergistic effects of AdoMet and cisplatin on cell
migration are reported via the evaluation of wound recovery. Taken
together these results highlight AdoMet as a potential candidate
for the development of novel treatment strategies for patients with
HNSCC.
Materials and methods
Reagents
AdoMet was obtained from New England BioLabs, Inc.
and prepared as previously described (13). The Annexin V-fluorescein
isothiocyanate (V-FITC) Apoptosis Detection kit was purchased from
eBioscience; Thermo Fisher Scientific, Inc. Tissue culture dishes
were purchased from Corning Inc. Monoclonal antibodies (mAbs) to
p21 (#2947S), p53 (#2524S), β-actin (#3700S), β-catenin (#8480S),
AKT (#2966S), phospho-AKT (#2965S), cyclin E1 (#4129S), cyclin A2
(#4656S), cyclin D1 (#2978S), phospho-cdc25C (#4901S), vimentin
(#5741S), E-cadherin (#14472S), phospho-SMAD3 (#9520S), SMAD3
(#9523S) and polyclonal antibodies (polyAbs) to cyclin B1 (#4138S),
SMAD2 (#3102S), phospho-SMAD2 (#3104S), MMP2 (#4022S) were
purchased from Cell Signaling Technology, Inc.; uPA (AB2335602,
cat. no. 119) polyAb was obtained from American Diagnostica, Inc.,
while mAbs directed to N-cadherin (05-915) and MMP9 (AB6001) were
purchased from Merck Millipore. PolyAb to cdc25c (sc-13138) was
purchased from Santa Cruz Biotechnology, Inc. Goat anti-rabbit IgG
Alexa Fluor647 was obtained from Abcam. Horseradish peroxidase
(HRP)-conjugated goat anti-mouse (GtxMu-003-DHRPX) and
HRP-conjugated goat anti-rabbit (GtxRb-003-DHRPX) secondary
antibodies were obtained from ImmunoReagents Inc. All buffers and
solutions were prepared with ultra-high quality water. All reagents
were of the purest commercial grade.
Cells and cell culture
The HNC cell lines, Cal-33 (oral squamous cancer
cell line) and JHU-SCC-011 (laryngeal squamous cell carcinoma line)
were obtained from the American Type Culture Collection (ATCC). The
cells were cultured at 37˚C in a 5% CO2 humidified
atmosphere and grown in RPMI supplemented with 10% heat-inactivated
FBS, 100 U/ml penicillin, 100 µg/ml streptomycin and 1%
L-glutamine.
Flow cytometry for the analysis of
apoptosis
Annexin V-FITC was used in conjunction with the
vital dye prop-idium iodide (PI) as previously described
(13) to distinguish apoptotic
(Annexin V-FITC-positive, PI-positive) from necrotic (Annexin
V-FITC-negative, PI-positive) cells (23). The detection of viable cells, early
apoptotic cells, late apoptotic cells and necrotic cells was
performed using a BD Accuri™ C6 flow cytometer (BD Biosciences).
For each sample, 20,000 events were acquired. Analysis was carried
out by triplicate determination on at least 3 separate
experiments.
Preparation of cell lysates
Oral cancer cells Cal-33 and JHU-SCC-011 cells,
grown at 37˚C for 24 and 48 h with or without AdoMet treatment,
were collected by centrifugation, washed twice with ice-cold PBS
and the pellet was lysed using 100 µl RIPA Buffer. Following
incubation on ice for 30 min, the samples were centrifuged at
18,000 x g in an Eppendorf microcentrifuge for 30 min a 4˚C and the
supernatant was recovered. The protein concentration was determined
using the Bradford method (24)
and compared with the BSA standard curve.
Western blot analysis
Equal amounts of cell proteins were separated by
SDS-PAGE (separating gel, 10 or 12%; stacking gel, 5%) and
electrotransferred onto nitrocellulose membranes by Trans blot
turbo (Bio-Rad Laboratories, Inc.). The mass of protein loaded per
lane was 10-50 µg. Membrane were washed in TBST (10 mM Tris, pH
8.0, 150 mM NaCl, 0.05% Tween-20), and blocked with TBST
supplemented with 5% non-fat dry milk. Thereafter, the membranes
were incubated overnight with the different primary antibodies at
4˚C in TBST and 5% non-fat dry milk, washed and incubated for 1 h
at room temperature with HRP-conjugated secondary antibodies. All
primary antibodies were used at a dilution of 1:1,000, and all
secondary antibodies were used at a 1:5,000 dilution. Blots were
then developed using enhanced chemiluminescence detection reagents
ECL (Cyanagen,) and exposed to X-ray film. All films were scanned
using ImageJ software (National Institutes of Health).
Flow cytometric analysis of the cell
cycle
The Cal-33 and JHU-SCC-011 cells were seeded in
6-well plates at a density of 75x103 cells/well and
30x103 cells/well, respectively. The following day, the
cells were treated with 300 µM AdoMet. The Cal-33 and JHU-SCC-011
cells were recovered with trypsin-EDTA after 24 and 48 h,
respectively, washed in PBS and stained in a PI solution (50 µg/ml
PI, 0.1% sodium citrate, 25 µg/ml RNase A, 0.1% triton in PBS) for
1 h at 4˚C in the dark. Flow cytometric analysis was performed
using a BD Accuri™ C6 flow cytometer (BD Biosciences). To evaluate
cell cycle progression, PI fluorescence was collected as FL3-A
(linear scale) using ModFIT software (Verity Software House). For
each sample at least 20,000 events were analyzed in at least 3
different experiments giving a standard deviation (SD) <5%.
Fluorescence microscopy
To visualize the polymerized F-actin, cells
(~2x104/sample) were seeded on glass cover-slips and
cultured in growth medium in the absence (control) or presence of
300 µM AdoMet. After 24 and 48 h, the slides were washed with PBS,
fixed with 2.5% formaldehyde, permeabilized with 0.1% Triton X-100
for 10 min at 4˚C, and incubated with 0.1 µg/ml
rhodamine-conjugated phalloidin (Sigma-Aldrich; Merck KGaA) for 40
min at 23˚C (25). Nuclear
staining was performed with the 10 mM 4-6-diamidino-2-phenylindole
(DAPI) dye (#228549, Abcam) for 5 min at 23˚C. Finally, coverslips
were mounted using 20% (w/v) Mowiol, and visualized with an
Axiovert 200 M fluorescence inverted microscope connected to a
video camera (Carl Zeiss AG).
Invasion kinetic of cells monitored in
real-time
This assay was performed using E-16-well plates and
the xCELLigence Real-Time Cell Analysis (RTCA) technology as
previously described (26). This
technology measures impedance changes caused by the gradual
increase in electrode surface occupation by cells during the course
of time and provides a Cell Index value which is proportional to
the number of adherent cells. The bottom wells were coated with 20
µg/well Matrigel diluted in serum-free medium. Matrigel was allowed
to polymerize for 1 h at 37˚C prior to seeding the cells
(1x104 cells/well) suspended in serum-free medium
(control) or growth medium. Cells that cross the Matrigel adhere to
the bottom of plates causing impedance changes, which are
proportional to the number of invading cells. Matrigel invasion was
monitored in real-time for 24 h and impedance changes were recorded
and expressed as a cell index value. Slopes represent the change
rate of cell index generated in a 3 to 20-h time frame. The
experiments were performed 3 times in quadruplicate.
Migration process evaluated by
scratch-wound assay
The Cal-33 and JHU-SCC-011 cells were seeded in the
appropriate number in a 6-well culture plates until 100% confluence
was reached in 24 h and treated with 200 and 300 µM AdoMet for 24
and 48 h, respectively or with 300 µM AdoMet alone or in
combination with 0.18 or 0.36 µM cisplatin (cDDP) (Sigma-Aldrich;
Merck KGaA). Following treatment, in a sterile environment, a 200
µl pipette tip was used to manually press against the top of the
tissue culture plate and a vertical wound down was rapidly created
through the confluent cellular monolayer. Carefully, medium and
cell debris were aspirated away, replaced with 2 ml of serum-free
RPMI and initial images of the wounds were captured using a
microscope (Leica Microsystems GmbH) corresponding to time zero
(T0). Following 24 h (T1) of treatment, snapshot images were
captured to examine for wound closure. The wound areas of the
control and treated cells were quantified using ImageJ software
1.48v.
Statistical analysis
The results are expressed as the means of at least 3
independent experiments performed in quadruplicate, unless
otherwise indicated. Data are expressed as the means ± SD. The
means were compared using analysis of variance (ANOVA) plus the
Bonferroni correction. A P-value <0.05 was considered to
indicate a statistically significant difference.
Results
Effects of AdoMet on the apoptosis of
Cal-33 and JHU-SCC-011 cells
In a previous study by the authors, to the best of
our knowledge, the mechanisms underlying the anti-tumor effect of
AdoMet on HNSCC were reported for the first time, demonstrating
that AdoMet was able to induce apoptosis, involving a
caspase-dependent mechanism paralleled by an increased Bax/Bcl-2
ratio in Cal-33 and JHU-SCC-011 cells following 48 and 72 h of
treatment, respectively (13).
In order to evaluate the effects of AdoMet prior to
the induction of apoptotic cell-death, the Cal-33 and JHU-SCC-011
cells were treated with 300 µM AdoMet, and the apoptotic process
was evaluated after 24 and 48 h, respectively, by
fluorescence-activated cell sorting (FACS) analysis after double
labeling with Annexin V-FITC and PI. In the absence of any
treatment, very few apoptotic cells were found in both cell
populations (Fig. 1, control).
The Cal-33 cells treated with 300 µM AdoMet for 24
h, exhibited an approximately 10% greater number of apoptotic cells
compared to the control group (Fig.
1A), while the JHU-SCC-011 cells treated with 300 µM AdoMet for
48 h exhibited an approximately 3% greater number of apoptotic
cells compared to the control group (Fig. 1B). According to a previous study by
the authors, the two cell lines are character-ized by a different
susceptibility to AdoMet. Therefore, the longer exposure time (48
h) of the more resistant JHU-SCC-011 cells resulted in lower
apoptotic rates compared with those observed following a shorter
exposure time (24 h) of the more sensitive Cal-33 cells. These
results indicated that AdoMet at a 300 µM concentration and at 24 h
for the Cal-33 and 48 h for the JHU-SCC-011 cells, did not induce
any relevant apoptotic death in both cell lines.
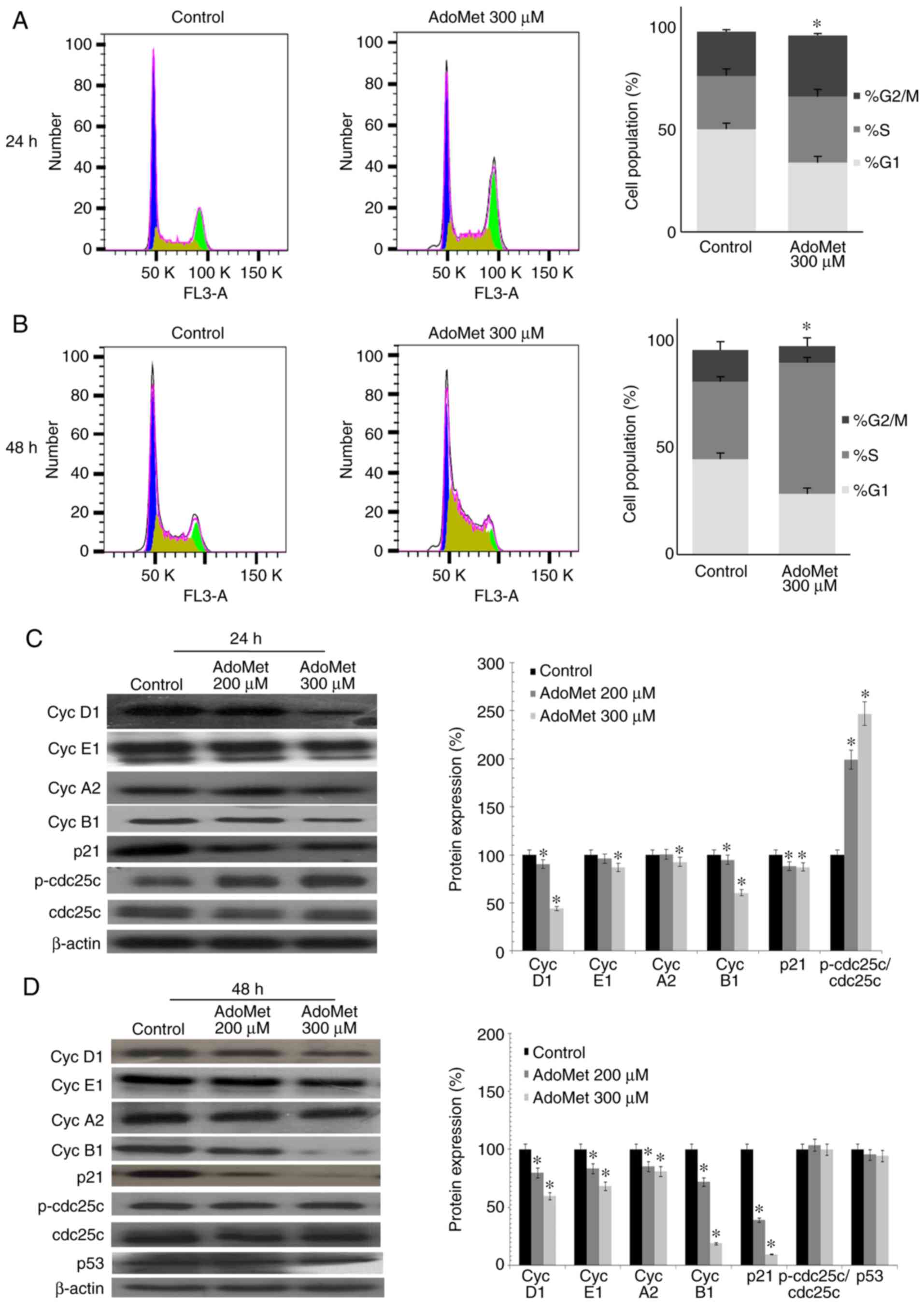
AdoMet promotes the cell cycle arrest of
Cal-33 and JHU-SCC-011 cells
In order to determine whether AdoMet treatment
promotes cell cycle arrest, cell cycle progression was analyzed by
flow cytometry in the Cal-33 and JHU-SCC-011 cells treated with 300
µM AdoMet for 24 and 48 h, respectively. As shown in Fig. 2A, AdoMet induced the marked
accumulation of Cal-33 cells in the S/G2M phase with an increase
from 47.6 to 61.9%, while the percentage of cells in the G1 phase
decreased from 50.2 to 33.9%. A different effect was observed with
the JHU-SCC-011 cells. Indeed, as shown in Fig. 2B, following AdoMet treatment, the
G1 and G2/M phase populations decreased from 44.6 to 28.4% and from
14.7 to 7.6%, respectively, with a concomitant significant increase
from 36.1 to 61% of cells in the S phase. These results demonstrate
that AdoMet is able to modulate cell cycle progression of HNSCC
cells.
To further investigate the effects of AdoMet on cell
cycle distribution, the expression level of several key cell cycle
regulators, such as cyclin D1, E1, A2 and B1 was examined by
western blot analysis. As shown in Fig. 2C and 2D, a dose-dependent decrease in the
expression of cyclin B1, E1 and D1 was observed in the Cal-33 and
JHU-SCC-011 cells, respectively following treatment with AdoMet
compared to the untreated cells. The level of cyclin A2 was shown
to decrease only with the 300 µM concentration of AdoMet in the
Cal-33 cells in (Fig. 2C), while a
dose-dependent decrease was observed in the expression of cyclin A2
in the JHU-SCC-011 cells. These findings suggest that AdoMet
attenuated cell proliferation via the downregulation of cyclin
expression. Subsequently, the expression levels of cdc25C,
phospho-cdc25C, p53 and the cyclin-dependent inhibitor, p21, were
examined. The results revealed a substantial dose-dependent
increase in the phospho-cdc25C/cdc25C ratio in the Cal-33 cells,
while no significant change was detected in the JHU-SCC-011 cells.
Notably, p21 expression was significantly decreased in both cell
lines following AdoMet treatment, while the expression of p53,
which is mutated in the Cal-33 cells, did not appear to be modified
in the JHU-SCC-011 cells, indicating that the AdoMet-induced cell
cycle arrest is a p53/p21-independent process.
Effects of AdoMet on the cytoskeletal
organization and migration of Cal-33 and JHU-SCC-011 cells
In order to investigate the early intracellular
effects occurring during AdoMet treatment, cytoskeletal
organization and the motility of Cal-33 and JHU-SCC-011 cells
exposed to 300 µM AdoMet for 24 and 48 h, respectively were
analyzed. As revealed by the rhodamine-phalloidin staining of
F-actin polymerization, in the absence of AdoMet, both the Cal-33
and JHU-SCC-011 cells exhibited a well-organized cytoskeleton with
condensed aggregates local ized below the membrane (Cal-33 cells)
or stress fibers parallel to the longitudinal axis of the cells
(JHU-SCC-011 cells). Cell exposure to AdoMet did not significantly
modify the cyto-skeletal organization of either the Cal-33 and
JHU-SCC-011 cells (Fig. 3A and B).
Conversely, AdoMet decreased the migration of the Cal-33 and
JHU-SCC-011 cells in a wound healing experiment monitored for 24 h.
In both cases, the wounds disappeared after 24 h in the absence of
AdoMet, as shown by the microscopy images of the same field
recorded at time 0 and after 24 h (Fig. 3C and D). Cell exposure to 200 and
300 µM AdoMet for 24 h led to an approximately 24 and 40%
reduction, respectively, in the Cal-33 wound closure (Fig. 3C and E). Of note, the spreading of
the JHU-SCC-011 cells was markedly reduced by AdoMet treatment for
48 h. Indeed, treatment with 200 and 300 µM AdoMet led to a 50 and
70% reduction in wound closure, respectively (Fig. 3D and F). These findings thus
indicate that AdoMet inhibits the migration of both HNSCC lines,
without affecting cytoskeletal organization.
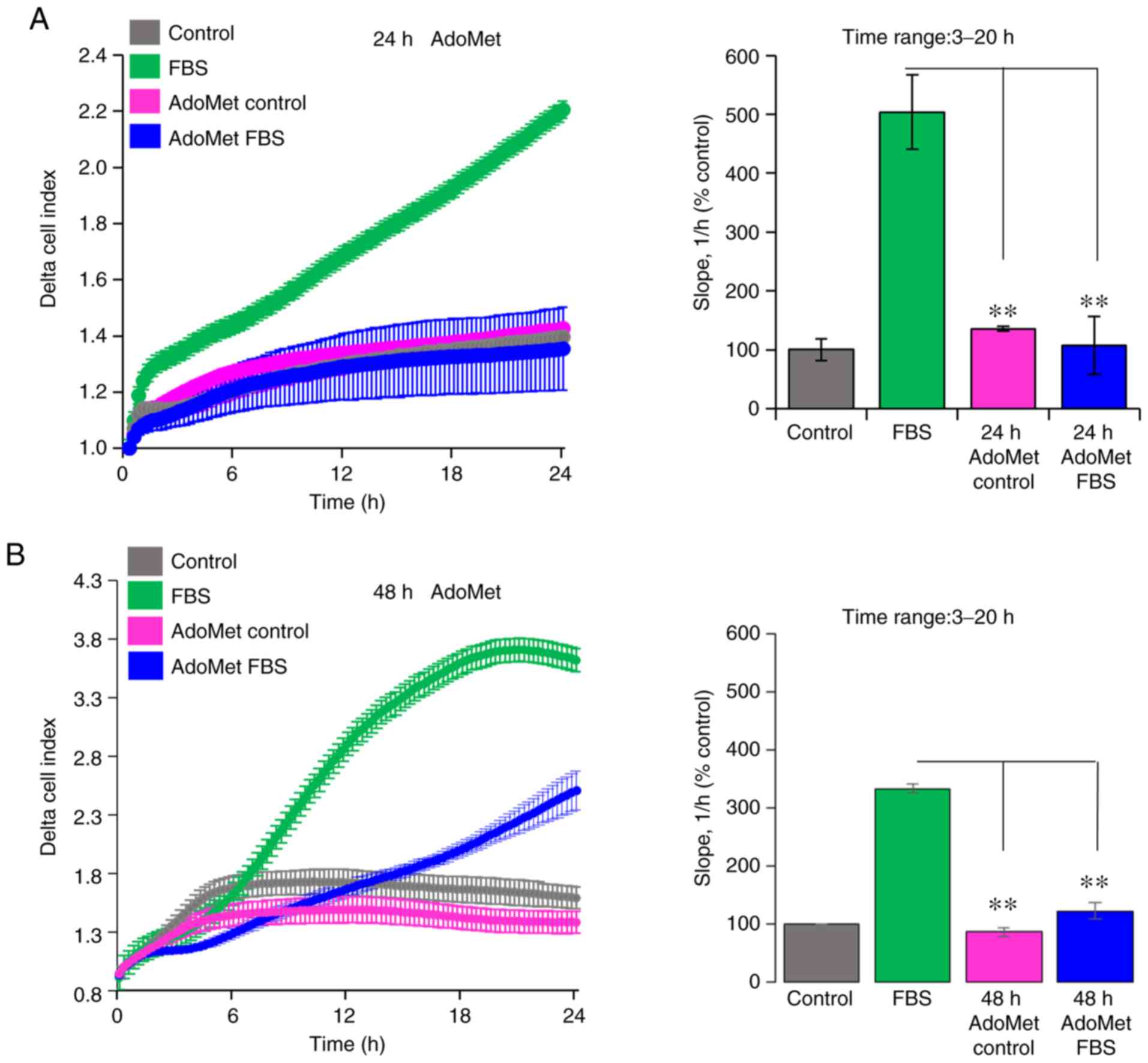
AdoMet affects the invasion of Cal-33 and
JHU-SCC-011 cells on Matrigel
Since cell motility is a prerequisite for the
acquisition of an invasive phenotype, the effects of AdoMet on the
ability of Cal-33 and JHU-SCC-011 cells to cross the Matrigel, a
reconstitute basal membrane using xCELLigence RTCA technology, were
examined (26). The Cal-33 and
JHU-SCC-011 cells treated with diluents or 300 µM AdoMet for 24 and
48 h, respectively, were seeded at the bottom of E-plates coated
with polymerized Matrigel. Matrigel invasion was monitored in
real-time for 24 h as the cell index changes due to the adhesion of
invading cells to microelectrodes. Although to a different extent,
both the Cal-33 (Fig. 4A) and
JHU-SCC-011 (Fig. 4B) cells were
able to cross the Matrigel, and AdoMet did not affect their basal
cell invasion, as compared to the untreated cells (Fig. 4). According to the cell migration
data, pre-exposure of the cells to 300 µM AdoMet led to a 73 and
63% inhibition of Cal-33 and JHU-SCC-011 cell invasive ability,
respectively, as indicated by the slopes representing the cell
index changes generated in the 3 to 20-h time frame (Fig. 4).
Taken together, these data highlight the inhibitory
effect of AdoMet on the migratory and invasive ability of HNC
cells, raising the possibility that AdoMet may be considered, not
only as a pro-apoptotic drug for the treatment of HNSCC, but also
as a good candidate for preventing the cell invasive ability.
AdoMet downregulates the levels of
proteins involved in migration and in invasion processes
The metastatic process consists of a series of
sequential, interrelated steps, including tumor cell detachment
from the primary tumor, increased motility and invasion,
proteolysis and resistance to apoptosis. To further examine the
effects of AdoMet treatment on the motility and invasiveness of
HNSCC cells, the main migration and invasion markers characterizing
the epithelial-mesenchymal transition (EMT) process, such as
E-cadherin and N-cadherin, belonging to a family of transmembrane
glycoproteins that mediate cell-cell adhesions, and vimentin, a
protein that plays a significant role in anchoring organelles in
the cytosol, were examined by western blot analysis (Fig. 5). The analysis revealed a marked
concentration-dependent decrease in the N-cadherin and vimentin
levels, and a concomitant increase in E-cadherin levels in the
Cal-33 and JHU-SCC-011 cells exposed to 200 and 300 µM AdoMet for
24 h, as compared with the control (untreated cells) (Fig. 5). In addition, according to the
cell invasion data, treatment with 200 and 300 µM AdoMet for 24 h
led to a clear-cut reduction in the levels of the uPA, MMP2 and
MMP9 proteolytic enzymes in both the Cal-33 and JHU-SCC-011 cell
lysates (Fig. 5). Moreover, to
further investigate the mechanisms underlying the inhibitory
effects of AdoMet on the invasive and migratory ability of Cal-33
and JHU-SCC-011 cells, several cancer-related signaling pathways
associated with invasion and migration were examined by western
blot analysis. The results revealed that the sulfonium compound
induced a decrease in the p-AKT/AKT, p-SMAD2/SMAD2 and
p-SMAD3/SMAD3 ratios, and a downregulation in the expression of
β-catenin (Fig. 5).
 | Figure 5Effect of AdoMet on proteins involved
in the migration and invasion processes and in the main cell growth
and survival pathways of HNSCC. (A) Cal-33 and (B) JHU-SCC-011
cells were treated or not (control), with 200 or 300 µM AdoMet for
24 and 48 h, respectively. The expression levels of E-cadherin,
N-cadherin, vimentin, MMP2, MMP9 and uPA along with phospho-AKT,
AKT, β-catenin and p-SMAD2, SMAD2, p-SMAD3, SMAD3 were detected by
western blot analysis using the total cell lysates. Densitometric
analyses results are also presented. Data are representative of 3
experiments performed with 3 different cellular preparations, are
expressed as the means + standard deviation and are reported as a
percentage of the protein expression of the untreated control
(100%). *P<0.05 vs. control. For the equal loading of
protein in the lanes, β-actin was used. HNSCC, head and neck
squamous cell carcinoma; AdoMet, S-adenosyl-L-methionine; MMP,
matrix metalloproteinase; uPA, urokinase-type plasminogen
activator; SMAD, small mothers against decapentaplegic. |
AdoMet synergistically enhances the
cDDP-induced inhibition of cell migration
In a previous study by the authors, the synergistic
effect of AdoMet in association with cDDP, an agent commonly used
in cancer therapy, in inhibiting Cal-33 cell proliferation and in
enhancing cell apoptosis, was demonstrated (13). It was found that the optimal
combination of the two drugs, highly synergistic with the CalcuSyn
calculation, corresponded to 200 µM AdoMet and 0.18 µM cDDP
following 72 h of treatment. In the present study, in order to
examine the synergistic effect induced by the AdoMet/cDDP
combination on the migration process, the wound closures in the
Cal-33 and JHU-SCC-011 cells treated with 300 µM AdoMet plus/minus
0.18 or 0.36 µM cDDP for 24 and 48 h, respectively, were examined.
As shown in Fig. 6, the wound
width with the combination treatments were significantly greater
compared to those of the control or to the single compound
treatments with wound closure percentage values of 23 and 31% in
the Cal-33 and JHU-SCC-011 cells, respectively compared to 100% of
the control. The obtained results indicated that, when used in
combination, AdoMet and cDPP were more effective in reducing cell
migration than individual agents confirming that AdoMet potentiates
the cytotoxic effects of cDDP on HNSCC.
Discussion
Although the majority of cancer-associated mortality
cases are caused by metastatic cancer rather than primary tumors,
this biology complex process remains the least understood feature
of cancer. The biochemical mechanisms and processes involved in
metastatic cancer have been designed, such as potential targets for
the prevention and inhibition of metastasis. HNSCC represents one
of the most common types of tumor worldwide and almost 60% of
patients with HNSCC develop metastases that limit their survival
(1-4). Adverse toxic side-effects of
chemotherapy during HNSCC treatment have shifted the focus towards
anticancer natural compounds as a valuable source of novel and less
toxic drugs that can be useful to block both tumor growth and
metastatic spread of cancer cells.
The naturally occurring sulfur-containing
nucleoside, AdoMet, is an important and ubiquitous biomolecule with
a variety of biological functions, and the majority of these have
been fully elucidated. AdoMet is one of the most frequently
metabolites involved in intermediary metabolism, since it is the
principal biological methyl donor in the cytosol of all mammalian
cells. AdoMet also plays a role in other processes, such as
transsulfuration reactions and polyamine synthesis (5-7).
Despite emerging data from the literature on the
anti-prolif-erative and anti-metastatic effects exerted by AdoMet
in a variety of cancer cells are accumulating (8-13),
currently, only limited data are available on the molecular
mechanisms underlying the anticancer effects of AdoMet on HNSCC
(13), and the role of AdoMet in
the invasion and migration of HNSCC has not been investigated to
date, at least to the best of our knowledge.
Firstly, the present study evaluated the effect of
AdoMet on cell viability. Cell cycle progression and its regulation
is the key event of cell proliferation. In cancer cells, a number
of cell cycle regulators, including p53, are mutated or
inactivated, leading to uncontrolled cell proliferation. Thus, the
ability of natural compounds with anticancer properties to reduce
malignant growth by controlling cell cycle events, may be
considered useful for cancer treatment and prevention.
In the current study, it was demonstrated that
AdoMet induced a marked cell cycle arrest at the G2/M phase in
Cal-33 cells, while it significantly increased the percentage of
JHU-SCC-011 cells in the S phase. G2/M arrest is largely mediated
through the phosphorylation of cdc25C, a p53-independent mechanism,
and the consequential maintenance of cyclin-dependent kinases
1/cyclin B in the phosphory-lated and inhibited state (27,28).
Of note, the present study found that AdoMet-induced G2/M phase
arrest in Cal-33 cells was accompanied by a notable increase in the
level of phosho-cdc25C protein. Moreover, it was found that AdoMet
treatment modulated the cell cycle by downregulating the levels of
cyclin D1, E1, A2, B1 and p21, while leaving the p53 levels
unmodified, suggesting that cell cycle arrest at the G2/M and S
phase in the Cal-33 and JHU-SCC-011 cells, respectively, is a
p53/p21-independent process.
DNA methylation represents one of the most
well-studied epigenetic processes in tumors (29). The ability of the sulfonium
compound to regulate genes responsible for cell invasion and
metastasis, has been well documented in human cancer and in
particular, AdoMet has been reported to induce the downregulation
of pro-metastatic genes, including uPA and MMPs (16,19).
Accordingly, the present study found that AdoMet treatment
decreased the uPA, MMP2 and MMP9 content in the Cal-33 and
JHU-SCC-011 cells. uPA is an extracellular serine protease
implicated in tumor invasion and metastasis processes either
directly by degrading extracellular matrix barriers or indirectly,
by inducing plasmin-dependent activation of latent MMPs (30). Similar to uPA, MMPs are involved in
extracellular matrix remodeling and degradation and play a role in
modulating all stages of carcinogenesis, from tumor initiation to
metastasis (31). The
AdoMet-induced decrease in the levels of uPA, MMP2 and MMP9 are in
good accordance with the reduced invasive ability on Matrigel of
both the Cal-33 and JHU-SCC-011 cells treated with the sulfonium
compound. It was found that AdoMet did not affect basal cell
invasion, as compared to the untreated cells. However, despite
different cell invasive abilities, both cell lines responded to
AdoMet treatment to a similar extent, as 300 µM AdoMet reduced the
invasion of Cal-33 and JHU-SCC-011 cells on Matrigel by 73 and 63%,
respectively. Although the present study did not examine the effect
of AdoMet on the invasive ability of HNSCC in animal models, to the
best of our knowledge, this is first study to report the inhibitory
effects of AdoMet on the invasive abilities of two HNSCC lines,
Cal-33 and JHU-SCC-011 cells. The experimental evidence obtained by
the in vitro experiments however, needs to be verified and
broadened by further in vivo studies in the future.
The present study also demonstrated that AdoMet
reduced the migration of both HNSCC cell lines. Notably, the
inhibitory effect of AdoMet on cell migration could not be due to
apoptosis for the following reasons: i) treatment of the Cal-33 and
JHU-SCC-011 cells with 300 µM AdoMet for 24 and 48 h, respectively,
did not trigger appreciable apoptotic cell death in both cell
lines; ii) the cytoskeleton machinery, which is required for cell
migratory ability, was not modified by AdoMet treatment. Moreover,
in line with the AdoMet-dependent decrease in cell motility and
invasiveness, it was found that AdoMet treatment led to an
appreciable decrease in the levels of N-cadherin and vimentin
mesenchymal makers, and to a concomitant increase in the levels of
the E-cadherin epithelial maker in HNSCC.
The characterization of the main growth and survival
pathways involved in HNSCC seems to be the same in spite of the
variety of risk factors and anatomical locations of origin. The
molecular signaling pathways that appear to be most consistently
modified are the PI3K/AKT and TGFβ/SMAD pathways (32). These pathways have been found to be
responsible for neoplastic transformation, as well as for cancer
invasion, a prerequisite for the development of metastatic disease.
It has been reported that the phospho-AKT levels are positively
with the prognosis of patients affected by oral squamous cell
carcinoma, whereas the E-cadherin levels are inversely associated
(33). In HNSCC, the activation of
PI3K/AKT increases MMP9 expression, degrades E-cadherin and
promotes cell invasion and migration (34). Moreover, the phosphorylation of AKT
causes the inactivation of phos-phorylated glycogen synthase kinase
3b, which is an important component of the canonical Wnt pathway,
one of the hallmarks for the initiation of the EMT, leading to the
accumulation of β-catenin in the nucleus and to the induction of
cell proliferation and migration (35-37).
TGF-β/SMAD signaling promotes EMT in late-stage
cancer, playing an important role in the wound healing process,
which requires cell migration (38,39).
Several studies have demonstrated synergistic effects between the
TGF-β/SMAD and Wnt/β-catenin signaling pathways in a number of
cellular functions, including wound healing (40-43).
The activation of SMAD2/3, AKT and β-catenin leads to the
upregulation of EMT-related transcription factors and mesenchymal
markers, and to the downregulation of epithelial makers (33,40-45).
In line with this experimental evidence, the present study reported
that AdoMet reduced the phospho-AKT levels, downregulated β-catenin
accumulation and modulated the levels of SMAD2 and SMAD3, as well
as the levels of their phosphorylated forms in the Cal-33 and
JHU-SCC-011 cells, impairing cell proliferation and spreading.
It has been amply reported that cyclin D1, in
addition to playing an important role in controlling the transition
G1/S phase during cell cycle progression, is also a proto-oncogene
involved in the regulation of cell migration and invasion (46). In analogy, p21, initially
recognized as a downstream effector of p53-dependent cell cycle
arrest induced by DNA damage, also functions as a 'two-faced'
regulator. Indeed, depending on the cell type, cellular
localization, the p53 status and the type and level of genotoxic
stress, p21 can acquire either onco-suppressive or onco-promoting
properties (47,48). Cyclin D1 and p21 are often
overexpressed in human cancers and their levels are associated with
a high tumor grade, a poor prognosis and increased metastasis in
several types of cancer (46-48).
Furthermore, it has been recently demonstrated that p21 acts as a
transcriptional co-regulator of SMAD that mediates TGFβ-induced
breast cancer cell migration and invasion (49). It has been also demonstrated that
cyclin D1 cooperates with p21 to regulate TGFβ-mediated breast
cancer cell migration and that p21/cyclin D1-depleted tumors
displayed less invasive features (50). According to this experimental
evidence, it is conceivable that the AdoMet-induced downregulation
of p21 and cyclin D1 in the Cal-33 and JHU-SCC-011 cells, may
represent a possible mechanism, which has not been reported to
date, through which the sulfonium compound inhibits HNSCC migration
through the TGF-β/SMAD signaling pathway.
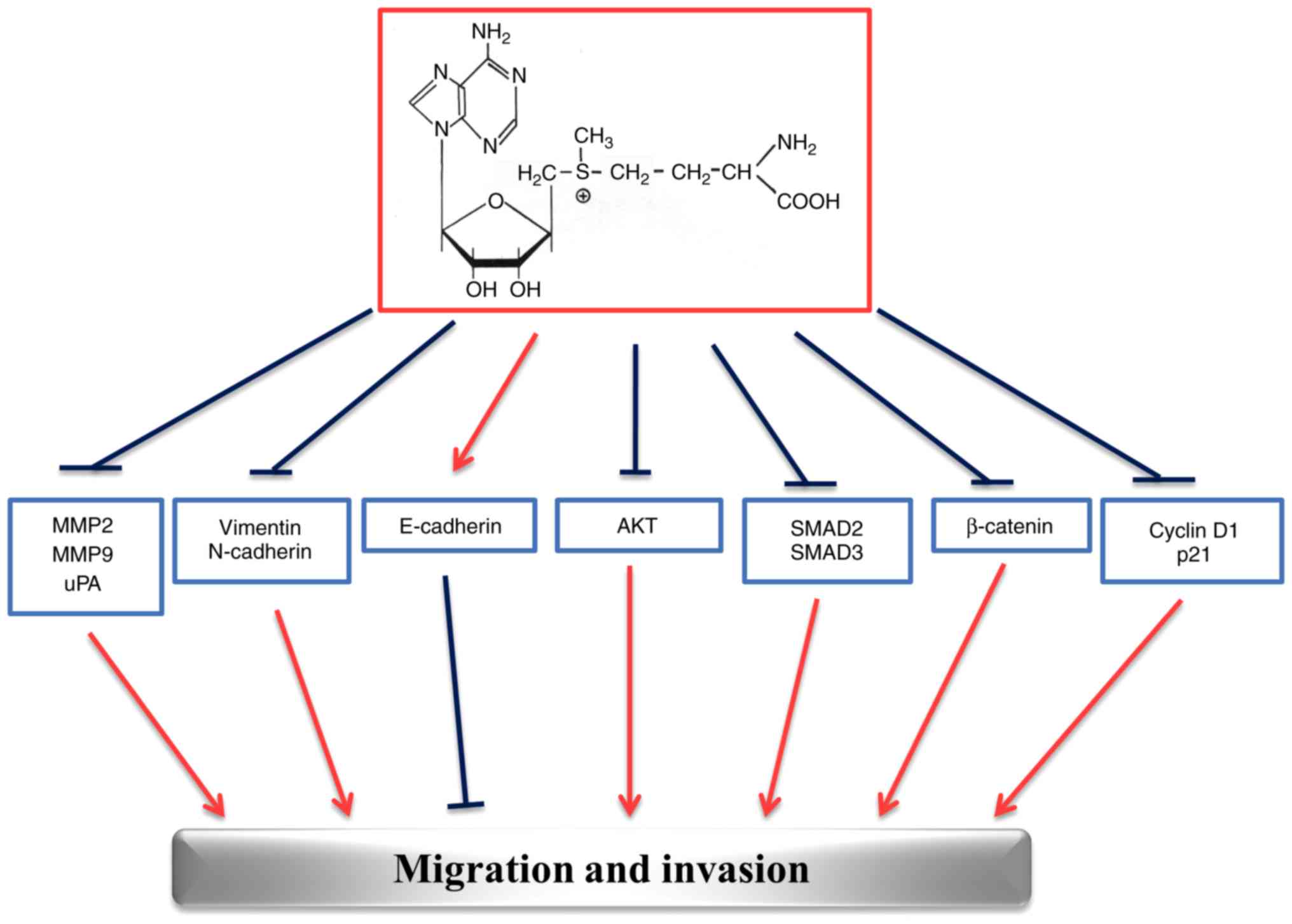
The multi-targeted effects exerted by AdoMet in
HNSCC are summarized in Fig. 7.
The ability of AdoMet to inhibit migration and invasion processes
at multiple levels, stimulates great interest for future
investigations and suggests that AdoMet may be an attractive
candidate for the development of drugs against HNSCC.
Finally, to emphasize the potential role of AdoMet
as an anticancer molecule, the present study analyzed the wound
closure effectiveness of the combination of sulfonium compound with
cDDP, a characteristic platinum complex already applied in HNSCC
alone or in synergy with other drugs for therapeutic purposes
(51). In a previous study on
Cal-33 cells, it was demonstrated that after 72 h, combined
AdoMet/cDPP treatment resulted in a potent synergistic
pro-apoptotic effect on Cal-33 cells; the use of AdoMet at the 200
mM concentration meant that the concentration of active cDDP was
able to be reduced up to 0.18 µM, the lowest concentration used in
combination with other natural compounds (13).
The present study demonstrates, for the first time,
to the best of our knowledge, that AdoMet and cDPP synergize to
prevent wound closure in Cal-33 and JHU-SCC-011 cells. Indeed, by a
wound healing assay, it was found that, after 24 h, the wounded
areas in the combination treatment groups were significantly larger
as compared to those of the control or single compound treatment
groups.
The mechanism of action of cDDP has been linked to
its ability to induce DNA damage via the formation of crosslinks
with the purine bases on DNA, interfering with DNA repair
mechanisms, and subsequently inducing the apoptosis of cancer
cells. Moreover, multiple signal transduction pathways are
activated by cDDP-induced genotoxic stress, through the activation
of MAPK pathways, particularly JNK and ERK (13,52).
These mechanisms may contribute to either apoptosis or
chemoresistance. Furthermore, it was recently demonstrated that
cDDP has the ability to interfere with the proliferation, migration
and invasion of nasopharyngeal carcinoma cells in vitro by
suppressing the Wnt/β-catenin/Endothelin-1 axis via the activation
of the B cell translocation gene 1 (52).
Taken together, these findings strongly support the
notion that AdoMet may be considered as an anticancer agent in
virtue of its ability to prevent the proliferation, migration and
invasion of HNSCC cells. Furthermore, the fact that AdoMet
effectively improves the cDDP-dependent inhibition of cell
migration, provides the basis for including this physiological
compound in the therapeutic management of metastatic HNSCC.
Moreover, AdoMet has been available as a dietary supplement in the
United States since 1999. Reviews of clinical studies to date have
indicated that, at pharmacological doses, AdoMet has a low
incidence of side-effects with an excellent record of tolerability.
Moreover, no toxic or anti-proliferative effects have been reported
in normal, non-tumorigenic cells (53-55).
Thus, it is conceivable that the concentrations of AdoMet that
would inhibit cancer cell proliferation, migration and invasion
processes, utilized in the present, as well as in other studies,
may be useful for further trials on patients.
Funding
The present study was partially supported by
Programme Valere 2019 (VANVITELLI per la Ricerca; Università della
Campania 'Luigi Vanvitelli').
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
MPo and LM conceived the study, designed the
experiments, analyzed the data and drafted the manuscript. GC and
MVC contributed to the design of the study and to the data analysis
and reviewed the manuscript. MPa and FV performed the experiments,
analyzed the data and prepared the figures; MM contributed to the
design and accomplishment of the experiments based on Matrigel
invasion and on cytoskeletal organization. All authors contributed
to the revision of the manuscript and read and approved the final
version of manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Abbreviations:
|
AdoMet
|
S-adenosyl-L-methionine
|
|
HNSCC
|
head and neck squamous cell
carcinoma
|
|
HNC
|
head and neck carcinoma
|
|
uPA
|
urokinase-type plasminogen
activator
|
|
MMP
|
matrix metalloproteinase
|
|
RTCA
|
real-time cell analysis
|
|
SMAD2
|
small mothers against
decapentaplegic-2
|
|
SMAD3
|
small mothers against
decapentaplegic-3
|
|
cDDP
|
cisplatin
|
|
TBST
|
Tris-buffered saline with Tween-20
|
|
PI
|
propidium iodide
|
|
HRP
|
horseradish peroxidase
|
|
Annexin V-FITC
|
Annexin V-fluorescein
isothiocyanate
|
|
TBS
|
Tris-buffered saline
|
|
FACS
|
fluorescence-activated cell
sorting
|
|
SD
|
standard deviation
|
|
EMT
|
epithelial-mesenchymal transition
|
References
|
1
|
Chin D, Boyle GM, Porceddu S, Theile DR,
Parsons PG and Coman WB: Head and neck cancer: Past, present and
future. Expert Rev Anticancer Ther. 6:1111–1118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marur S and Forastiere AA: Head and neck
squamous cell carcinoma: Update on epidemiology, diagnosis, and
treatment. Mayo Clin Proc. 91:386–396. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar M, Nanavati R, Modi TG and Dobariya
C: Oral cancer: Etiology and risk factors: A review. J Cancer Res
Ther. 12:458–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kulkarni P and Saxena U: Head and neck
cancers, the neglected malignancies: Present and future treatment
strategies. Expert Opin Ther Targets. 18:351–354. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu SC: S-Adenosylmethionine. Int J Biochem
Cell Biol. 32:391–395. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mato MJ, Cor rales FJ, Lu SC and Avila MA:
S-Adenosylmethionine: A control switch that regulates liver
function. FASEB J. 16:15–26. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fontecave M, Atta M and Mulliez E:
S-Adenosylmethionine: Nothing goes to waste. Trends Biochem Sci.
29:243–249. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ansorena E, García-Trevijano ER,
Martínez-Chantar ML, Huang ZZ, Chen L, Mato JM, Iraburu M, Lu SC
and Avila MA: S-Adenosylmethionine and methylthioadenosine are
anti-apoptotic in cultured rat hepatocytes but pro-apoptotic in
human hepatoma cells. Hepatol. 35:274–280. 2002. View Article : Google Scholar
|
|
9
|
Lu SC and Mato JM: S-Adenosylmethionine in
cell growth, apoptosis and liver cancer. J Gastroenterol Hepatol.
23(Suppl 1): S73–S77. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martínez-López N, Varela-Rey M, Ariz U,
Embade N, Vazquez-Chantada M, Fernandez-Ramos D, Gomez-Santos L, Lu
SC, Mato JM and Martinez-Chantar ML: S-Adenosylmethionine and
proliferation: New pathways, new targets. Biochem Soc Trans.
36:848–852. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ilisso CP, Sapio L, Delle Cave D, Illiano
M, Spina A, Cacciapuoti G, Naviglio S and Porcelli M:
S-Adenosylmethionine affects ERK1/2 and Stat3 pathways and induces
apoptosis in osteosarcoma cells. J Cell Physiol. 231:428–435. 2016.
View Article : Google Scholar
|
|
12
|
Cave DD, Desiderio V, Mosca L, Ilisso CP,
Mele L, Caraglia M, Cacciapuoti G and Porcelli M:
S-Adenosylmethionine-mediated apoptosis is potentiated by autophagy
inhibition induced by chloroquine in human breast cancer cells. J
Cell Physiol. 233:1370–1383. 2018. View Article : Google Scholar
|
|
13
|
Mosca L, Pagano M, Ilisso CP, Cave DD,
Desiderio V, Mele L, Caraglia M, Cacciapuoti G and Porcelli M:
AdoMet triggers apoptosis in head and neck squamous cancer by
inducing ER stress and potentiates cell sensitivity to cisplatin. J
Cell Physiol. 234:13277–13291. 2019. View Article : Google Scholar
|
|
14
|
Steeg PS: Tumour metastasis: Mechanistic
insights and clinical challenges. Nat Med. 12:895–904. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Braakhuis BJ, Senft A, de Bree R, de Vries
J, Ylstra B, Cloos J, Kuik DJ, Leemans CR and Brakenhoff RH:
Expression profiling and prediction of distant metastases in head
and neck squamous cell carcinoma. J Clin Pathol. 59:1254–1260.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pakneshan P, Szyf M, Farias-Eisner R and
Rabbani SA: Reversal of the hypomethylation status of urokinase
(uPA) promoter blocks breast cancer growth and metastasis. J Biol
Chem. 279:31735–31744. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shukeir N, Pakneshan P, Chen G, Szyf M and
Rabbani SA: Alteration of the methylation status of tumor-promoting
genes decreases prostate cancer cell invasiveness and tumorigenesis
in vitro and in vivo. Cancer Res. 66:9202–9210. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chik F, Machnes Z and Szyf M: Synergistic
anti-breast cancer effect of a combined treatment with the methyl
donor S-adenosylmethionine and the DNA methylation inhibitor
5-aza-2'-deoxycytidine. Carcinogenesis. 35:138–144. 2014.
View Article : Google Scholar
|
|
19
|
Hussain Z, Khan MI, Shahid M and Almajhdi
FN: S-adenosylmethionine, a methyl donor, up regulates tissue
inhibitor of metalloproteinase-2 in colorectal cancer. Genet Mol
Res. 12:1106–1118. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Parashar S, Cheishvili D, Arakelian A,
Hussain Z, Tanvir I, Khan HA, Szyf M and Rabbani SA:
S-Adenosylmethionine blocks osteosarcoma cells proliferation and
invasion in vitro and tumor metastasis in vivo: Therapeutic and
diagnostic clinical applications. Cancer Med. 4:732–744. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Bi T, Liu L, Gao Q, Shen G and Qin
L: S-Adenosylmethionine synergistically enhances the antitumor
effect of gemcitabine against pancreatic cancer through JAK2/STAT3
pathway. Naunyn Schmiedebergs Arch Pharmacol. 392:615–622. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun L, Zhang J, Yang Q, Si Y, Liu Y, Wang
Q, Han F and Huang Z: Synergistic Effects of SAM and selenium
compounds on proliferation, migration and adhesion of HeLa cells.
Anticancer Res. 37:4433–4441. 2017.PubMed/NCBI
|
|
23
|
Vermes I, Haanen C, Steffens-Nakken H and
Reutelingsperger C: A novel assay for apoptosis. Flow cytometric
detection of phosphatidylserine expression on early apoptotic cells
using fluorescein labelled Annexin V. J Immunol Methods. 184:39–51.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bradford MM: Rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fratangelo F, Camerlingo R, Carriero MV,
Pirozzi G, Palmieri G, Gentilcore G, Ragone C, Minopoli M, Ascierto
PA and Motti ML: Effect of ABT-888 on the apoptosis, motility and
invasiveness of BRAFi-resistant melanoma cells. Int J Oncol.
53:1149–1159. 2018.PubMed/NCBI
|
|
26
|
Ingangi V, Bifulco K, Yousif AM, Ragone C,
Motti ML, Rea D, Minopoli M, Botti G, Scognamiglio G, Fazioli F, et
al: The urokinase receptor-derived cyclic peptide [SRSRY]
suppresses neovascularization and intravasation of osteosarcoma and
chon-drosarcoma cells. Oncotarget. 7:54474–54487. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Malumbres M and Barbacid M: Mammalian
cyclin-dependent kinases. Trends Biochem Sci. 30:630–641. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gutierrez GJ, Tsuji T, Cross JV, Davis RJ,
Templeton DJ, Jiang W and Ronai ZA: JNK-mediated phosphorylation of
Cdc25C regulates cell cycle entry and G(2)/M DNA damage checkpoint.
J Biol Chem. 285:14217–14228. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kanwal R and Gupta S: Epigenetic
modifications in cancer. Clin Genet. 81:303–311. 2012. View Article : Google Scholar
|
|
30
|
Carriero MV and Stoppelli MP: The
urokinase-type plasminogen activator and the generation of
inhibitors of urokinase activity and signaling. Curr Pharm Des.
17:1944–1961. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kidacki M, Lehman HL, Warrick JI and
Stairs DB: Signaling pathways supporting tumor invasion in head and
neck squamous cell carcinoma. J Clin Exp Pathol. 5:2272014.
|
|
33
|
Silva BS, Yamamoto FP, Pontes FS, Cury SE,
Fonseca FP, Pontes HA and Pinto-Júnior DD: TWIST and p-Akt immune
expression in normal oral epithelium, oral dysplasia and in oral
squamous cell carcinoma. Med Oral Patol Oral Cir Bucal. 17:e29–e34.
2012. View Article : Google Scholar
|
|
34
|
Zuo JH, Zhu W, Li MY, Li XH, Yi H, Zeng
GQ, Wan XX, He QY, Li JH, Qu JQ, et al: Activation of EGFR promotes
squamous carcinoma SCC10A cell migration and invasion via inducing
EMT-like phenotype change and MMP-9-mediated degradation of
E-cadherin. J Cell Biochem. 112:2508–2517. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sheng S, Qiao M and Pardee AB: Metastasis
and AKT activation. J Cell Physiol. 218:451–454. 2009. View Article : Google Scholar
|
|
36
|
Marcucci F, Stassi G and De Maria R:
Epithelial-mesenchymal transition: A new target in anticancer drug
discovery. Nat Rev Drug Discov. 15:311–325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu W, Yang Z and Lu N: A new role for the
PI3K/Akt signaling pathway in the epithelial-mesenchymal
transition. Cell Adh Migr. 9:317–324. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Feng XH and Derynck R: Specificity and
versatility in TGF-β signaling through Smads. Annu Rev Cell Dev
Biol. 21:659–693. 2005. View Article : Google Scholar
|
|
39
|
Suriyamurthy S, Baker D, Ten Dijke P and
Iyengar PV: Epigenetic reprogramming of TGF-β signaling in breast
cancer. Cancers (Basel). 11. pp. E7262019, View Article : Google Scholar
|
|
40
|
Guo W, Flanagan J, Jasuja R, Kirkland J,
Jiang L and Bhasin S: The effects of myostatin on adipogenic
differentiation of human bone marrow-derived mesenchymal stem cells
are mediated through cross-communication between Smad3 and
Wnt/beta-catenin signaling pathway. J Biol Chem. 283:9136–9145.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Minoo P and Li C: Cross-talk between
transforming growth factor-beta and Wingless/Int pathways in lung
development and disease. Int J Biochem Cell Biol. 42:809–812. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu J, Wang Y, Pan Q, Su Y, Zhang Z, Han
J, Zhu X, Tang C and Hu D: Wnt/β-catenin pathway forms a negative
feedback loop during TGF-β1 induced human normal skin
fibroblast-to-myofi-broblast transition. J Dermatol Sci. 65:38–49.
2012. View Article : Google Scholar
|
|
43
|
Boldbaatar A, Lee S, Han S, Jeong AL, Ka
HI, Buyanravjikh S, Lee JH, Lim JS, Lee MS and Yang Y: Eupatolide
inhibits the TGF-β1-induced migration of breast cancer cells via
downregu-lation of SMAD3 phosphorylation and transcriptional
repression of ALK5. Oncol Lett. 14:6031–6039. 2017.PubMed/NCBI
|
|
44
|
Jin Y, Chen W, Yang H, Yan Z, Lai Z, Feng
J, Peng J and Lin J: Scutellaria barbata D. Don inhibits migration
and invasion of colorectal cancer cells via suppression of PI3K/AKT
and TGF-β/Smad signaling pathways. Exp Ther Med. 14:5527–5534.
2017.PubMed/NCBI
|
|
45
|
Cai W, Yu D, Fan J, Liang X, Jin H, Liu C,
Zhu M, Shen T, Zhang R, Hu W, et al: Quercetin inhibits
transforming growth factor β1-induced epithelial-mesenchymal
transition in human retinal pigment epithelial cells via the Smad
pathway. Drug Des Devel Ther. 12:4149–4161. 2018. View Article : Google Scholar :
|
|
46
|
Fustè NP, Castelblanco E, Felip I,
Santacana M, Fernández-Hernández R, Gatius S, Pedraza N, Pallarés
J, Cemeli T, Valls J, et al: Characterization of cytoplasmic cyclin
D1 as a marker of invasiveness in cancer. Oncotarget.
7:26979–26991. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Georgakilas AG, Martin OA and Bonner WM:
p21: A two-faced genome guardian. Trends Mol Med. 23:310–319. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jung YS, Qian Y and Chen X: Examination of
the expanding pathways for the regulation of p21 expression and
activity. Cell Signal. 22:1003–1012. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dai M, Al-Odaini AA, Arakelian A, Rabbani
SA, Ali S and Lebrun JJ: A novel function for p21Cip1 and
acetyltransferase p/CAF as critical transcriptional regulators of
TGFβ-mediated breast cancer cell migration and invasion. Breast
Cancer Res. 14:R1272012. View Article : Google Scholar
|
|
50
|
Dai M, Al-Odaini AA, Fils-Aimé N,
Villatoro MA, Guo J, Arakelian A, Rabbani SA, Ali S and Lebrun JJ:
Cyclin D1 cooperates with p21 to regulate TGFβ-mediated breast
cancer cell migration and tumor local invasion. Breast Cancer Res.
15:R492013. View Article : Google Scholar
|
|
51
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yin P, Song G and Jiang Z: Cisplatin
suppresses proliferation, migration and invasion of nasopharyngeal
carcinoma cells in vitro by repressing the
Wnt/β-catenin/Endothelin-1 axis via activating B cell translocation
gene 1. Cancer Chemother Pharmacol. 81:863–872. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Luo J, Li YN, Wang F, Zhang WM and Geng X:
S-adenosylmethionine inhibits the growth of cancer cells by
reversing the hypomethylation status of c-myc and H-ras in human
gastric cancer and colon cancer. Int J Biol Sci. 6:784–795. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lu SC and Mato JM: S-adenosylmethionine in
liver health, injury, and cancer. Physiol Rev. 92:1515–1542. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mahmood N, Cheishvili D, Arakelian A,
Tanvir I, Khan HA, Pépin AS, Szyf M and Rabbani SA: Methyl donor
S-adenosylmethionine (SAM) supplementation attenuates breast cancer
growth, invasion, and metastasis in vivo; therapeutic and
chemopreventive applications. Oncotarget. 9:5169–5183. 2018.
View Article : Google Scholar : PubMed/NCBI
|