Introduction
Colorectal cancer (CRC) is one of the most common
gastrointestinal malignancies, and the second most common cause of
cancer-associated mortality in the United States in 2016 (1). Growth factor receptors are
therapeutic targets in numerous malignancies, and dysregulation of
the fibroblast growth factor receptor (FGFR) serves an important
role in the genesis, progression and metastasis of colorectal
cancer (2). FGFR4 (a member of the
FGFR family) serves a pivotal role in the regulation of cell
proliferation, differentiation, migration and angiogenesis
(3-5); however, the mechanism underlying the
role of FGFR4 in colorectal cancer has not yet been elucidated.
It has previously been revealed that FGFR4 is highly
expressed in gastric cancer tissues, and patients with high FGFR4
expression exhibited a poor prognosis (6). BLU9931 is a FGFR4 inhibitor and
significantly inhibits colorectal cancer cell proliferation, while
increasing the rate of apoptosis (7). Moreover, it was reported that FGFR4
was a prognostic indicator of advanced-stage, high-grade serous
ovarian cancer, and knockdown of FGFR4 using siRNA significantly
inhibited the growth of ovarian tumors in vivo and in
vitro (8). FGF19 is a specific
agonist of FGFR4 (9). Notably,
treatment of hepatoma cell lines with recombinant FGF19 protein
resulted in an increase in cell proliferation and invasion;
however, this tendency was partially reversed when FGFR4 was
knocked down (10). This suggests
that FGFR4 may represent a key target for the treatment of
colorectal cancer.
Tumor recurrence and metastasis significantly impact
the survival of patients, and ~50% of patients with colorectal
cancer eventually succumb to the disease following local recurrence
and metastasis (11).
Epithelial-mesenchymal transition (EMT) facilitates tumor cells to
acquire invasive and metastatic potential (12). The loss of E-Cadherin protein
function is a crucial step in the EMT process (13). Zhao et al (14) demonstrated that FGFR4 knockdown in
hepatoma cells attenuated EMT progression by enhancing E-cadherin
expression. Abnormal activation and dysregulation of Wnt/β-catenin
pathway can result in tumorigenesis. The abnormal accumulation of
β-catenin in the nucleus is markedly associated with the occurrence
of EMT (15). Knockdown of
β-catenin promotes the expression of E-cadherin to reverse EMT
progression in prostate cancer cells (16). However, few studies have
investigated whether FGFR4 influences EMT progression via the
Wnt/β-catenin signaling pathway in colorectal cancer.
To explore the mechanism underlying the role of
FGFR4 in tumorigenesis, progression and EMT of colorectal cancer,
the expression of FGFR4 in colorectal cancer tissues and adjacent
normal colorectal tissues was examined by immunohistochemistry
(IHC) staining. The possible association between FGFR4 protein
expression and clinicopathological features and prognosis of
patients with CRC was evaluated. Furthermore, FGFR4 was silenced in
the colorectal cancer cell line SW620 by constructing a small
hairpin RNA (shRNA) lentivirus. A series of in vitro
functional assays were performed to investigate the effect of FGFR4
silencing on colorectal cancer cells. XAV-939 selectively inhibits
Wnt/β-catenin-mediated transcription by inhibiting tankyrase1/2,
and also promotes β-catenin degradation by stabilizing Axin and
inhibiting Wnt/β-catenin signaling (17,18).
Finally, the effect of FGFR4 on EMT progression was explored by
inhibiting the Wnt/β-catenin signaling pathway using the β-catenin
inhibitor XAV-939.
Materials and methods
Tissue microarray and
immunohistochemistry
Human CRC tissue microarray slides (cat. no.
HColA160Su02) were purchased from Shanghai Outdo Biotech Co., Ltd.
and included 100 CRC tissues and 60 adjacent normal tissues. All
patients had undergone surgery between July 2005 and December 2010
and were followed up by an outpatient visit and telephone call
every 6 months until September 2015. The present study was approved
by the Ethics Committee of Shanghai Outdo Biotech Co., Ltd. The
patients were recruited by several hospitals in Shanghai, and all
provided informed written consent for research. For
immunohistochemistry (IHC), FGFR4 staining was performed using a
polyclonal rabbit anti-FGFR4 antibody (ProteinTech Group, Inc.;
cat. no. 11098-1-AP; 1:50) as previously described (4). The stained tissue microarray was
scanned using a Panoramic Midi automatic slide scanner (3DHISTECH,
Hungary) Ltd. and captured byimaged using 3DHISTECH
PannoramicPanoramic Viewer 1.15.2. Semiquantitative scores of
cytoplasmic FGFR4 immunostaining (four-grade system) as follows: 0,
negative; 1, weak; 2, moderate; and 3, strong expression. High
expression was defined as when the immunostaining scores were 2 and
3, whereas low expression was when the scores were 0 and 1. The
tissue microarray was evaluated by two double-blinded independent
observers who did not know the clinical data and results.
Cell lines and cell culture
Human colorectal cancer cell line SW620 was
purchased from the American Type Culture Collection. Cells were
cultivated in Dulbecco's modified Eagle's medium (DMEM; Medicago,
Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml of penicillin and 0.1 mg/ml of
streptomycin (Caisson Laboratories, Inc.) at 37°C in a humidified
atmosphere containing 5% CO2.
Antibodies and reagents
Rabbit monoclonal anti-FGFR4 antibody (1:1,000; cat.
no. 8562), anti-Caspase3 (1:1,000; cat. no. 14220),
anti-active-β-catenin (1:1,000: cat. no. 19807), anti-Stat3
(1:1,000; cat. no. 12640), anti-CyclinD1 (1:1,000; cat. no. 55506),
anti-P27kip1 (1:1,000; cat. no. 3686) and anti-β-actin
(1:1,000; cat. no. 4970) antibody were all obtained from Cell
Signaling Technology, Inc. Rabbit polyclonal anti-FGFR4 (1:200;
cat. no. 11098-1-AP) antibody was purchased from ProteinTech Group,
Inc.. Secondary horse-radish peroxidase-conjugated antibodies were
goat anti-mouse (1:4,000; cat. no. 12-349) and goat anti-rabbit
(1:5,000; cat. no. AP132) from Sigma-Aldrich; Merck KGaA. XAV-939
(10 mM; cat. no. HY-15147) was a free sample, provided by Shanghai
Haoyuan Chemical Co., Ltd. and was diluted as follows: 10 mM XAV939
to 10 µM DMSO.
Lentivirus-mediated RNA interference
The pGLV3 lentiviral packaging plasmid carrying
green fluorescent protein (GFP) was purchased from Shanghai
GenePharma Co., Ltd. The company designed four short hairpin RNA
(shRNA; cat. nos. LV3-497, LV3-1679, LV3-1750 and LV3-2074)
sequences for FGFR4 silencing, in which the sequence of shRNA (cat.
no. LV3-1750) with the most pronounced effect of silencing FGFR4
was 5′-GCCGACACAAGAACATCATCA-3′. A non-silencing shRNA was used as
a control and its sequence was 5′-TTCTCCGAACGTGTCACGT-3′.
Colorectal cancer cells were seeded in 6-well plates at a density
of 3×105 cells per well. After 24 h, the prepared lentivirus was
added to transfect colorectal cancer cells according to the
manufacturer's instructions. Fluorescence microscopy was used to
analyze cell transfection efficiency. After 72 h, subsequent
experiments were performed.
Protein extraction and western
blotting
Whole-cell lysates were prepared by direct lysis
using RIPA buffer. Protein concentrations of the samples were
determined using the bicinchoninic acid protein assay (Pierce;
Thermo Fisher Scientific, Inc.). Protein samples (30 µg of
each protein) that were boiled for 5 min were separated on a 10%
SDS-polyacrylamide gel and transferred to a PVDF membrane. The
membrane was blocked with phosphate-buffered saline (PBS)
containing 0.05% Tween-20 and 5% non-fat dried milk for 1 h at room
temperature and incubated with the primary antibody at 4°C
overnight. The immunoblots were washed 3 times with PBS containing
0.05% Tween-20 and 1% non-fat dried milk. Then PVDF membranes were
incubated with secondary antibodies conjugated with horseradish
peroxidase against mouse IgG or rabbit IgG for 1 h at room
temperature. The level of a particular protein in each lysate was
detected using an ECL detection system (ImageQuant LAS 3000; GE
Healthcare).
Wound-healing and transwell invasion
assays
For wound-healing migration assays, colorectal
cancer cells were plated in 6-well plates at a density of
5×105 cells per well. The cells reached 100% confluence
overnight. A single scratch wound was created by dragging a 10
µl plastic pipette tip over a monolayer of cell surface.
Cells were cultured in DMEM containing 2% FBS at 37°C and 5%
CO2 for 72 h. The degree of closure of the wound within
72 h was calculated and expressed as a percentage of the difference
between 0 and 72 h. The percentage of wound closure was calculated
by measuring the healed area relative to the initial wound
area.
For invasion assays, 8-µm pore size Transwell
filter inserts (Corning Inc.) covered with Matrigel (200 ng/ml; BD
Biosciences) diluted by pre-cooled and serum-free DMEM medium in a
24-well plate, and incubated at 37°C for 30 min. Cells were
cultured in DMEM without fetal bovine serum overnight at 37°C.
After trypsinization, cells added in the upper chamber at a density
of 1×105 cells per insert. The lower chamber was filled
with DMEM containing 30% FBS. After incubation for 72 h at 37°C
with 5% CO2, the cells that invaded through a filter to
the lower surface of the membrane were fixed using methanol for 10
min at 20°C and stained with 0.2% crystal violet for 10 min at
20°C. Invaded cells were counted in five randomly selected fields
under a phase-contrast microscope (magnification, ×200).
Proliferation assay
Cells were seeded at a density of 2×103
cells per well in the 96-well plate (Corning Inc.) in 100 µl
of DMEM with 10% fetal bovine serum. After 1, 2, 3 and 4 days of
culture, the cells were incubated with 5 mg/ml MTT (10
µl/well) in a cell culture incubator at 37°C for 4 h and the
supernatant was removed. DMSO (100 µl/well) was added to
each well, and suspensions were agitated for 10 min to precipitate
a purple crystal. Absorbance was measured at 490 nm using a
microplate reader
Apoptosis detection
The rate of apoptosis in vitro was detected
using Annexin V-adenomatous polyposis coli (APC)/propridium iodide
(PI; Becton Dickinson and Company) staining. Cells were seeded at a
density of 3×105 cells per well in the 6-well plate
(Corning Inc.) in 2 ml of DMEM with 10% FBS. After 24 h cells were
harvested with EDTA-free trypsin. The cells were then incubated
with Annexin V-APC and PI in the dark at room temperature for 15
min at 37°C. Thereafter, all samples were analyzed by a FACSCalibur
(BD Biosciences) flow cytometer with CellQuest Pro software
(version 5.1; BD Biosciences). Only the AnnexinV+/PI− cells (early
apoptotic) were quantified in the final graph.
Cell-cycle analysis
Cells were harvested with EDTA-free trypsin, then
added the pre-cooled PBS to wash the cells twice. Cells were fixed
using 70% ethanol at 4°C overnight. Then the cells were resuspended
using 1 mg/ml PI and 10 mg/ml RNase A staining buffer for 15 min at
37°C in the dark. Samples were immediately analyzed using flow
cytometry (ACEA NovoCyte; ACEA Biosciences, Inc.) to separate
G0/G1, S, G2/M and hypodiploid
nuclei. All assays were carried out in triplicate.
Statistical analysis
Statistical analysis was carried out with SPSS 13.0
(SPSS, Inc.) and GraphPad Prism 5.0 (GraphPad Software, Inc.).
Staging was performed according to the American Joint Committee on
Cancer (AJCC) Tumor-Node-Metastasis (TNM) staging classification
for colorectal carcinoma (7th edition; 2010) (19). Results were presented as means ± SD
and three individual experiments were performed. Pearson's
χ2 test was used to analyze the association between
categorical variables. The 5-year survival rate was calculated
using the Life Tables method. Survival curves were plotted
according to the Kaplan-Meier method and compared using a log-rank
test. Multivariate Cox regression analysis was used to determine
the prognostic covariates of patients with CRC. Unpaired Student's
t test was used to compare data between two groups. One-way ANOVA
followed by Tukey's post-hoc test were applied to compare data
between three or more groups. P<0.05 was considered to indicate
a statistically significant difference.
Results
Association between clinicopathological
features and FGFR4 expression in colorectal cancer
The expression level of FGFR4 was evaluated in 100
colorectal cancer tissues and 60 adjacent normal colorectal via
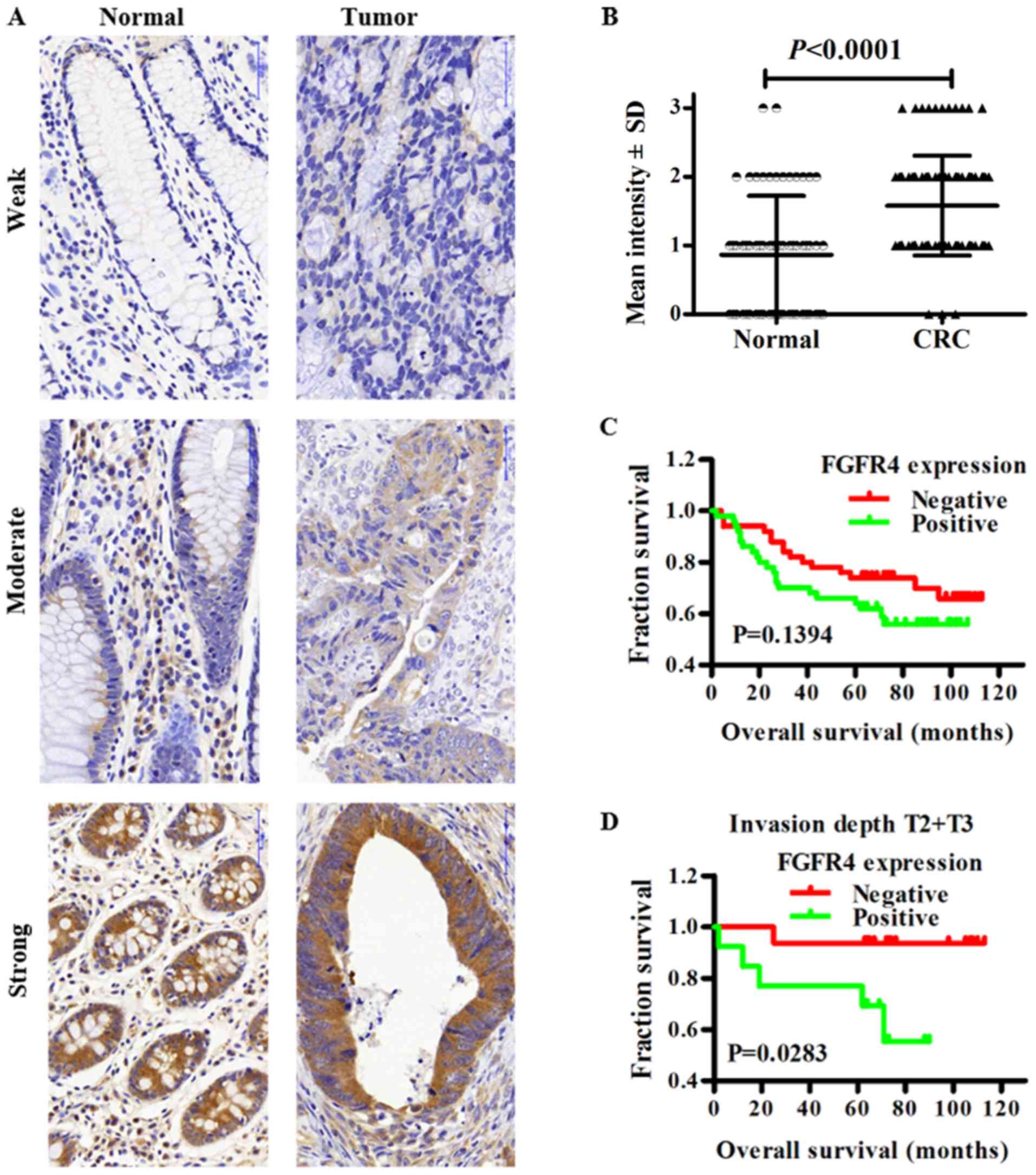
IHC. As exhibited in Fig. 1A,
FGFR4 was expressed in colorectal cancer tissues and normal colonic
tissues, and was primarily expressed in the cytoplasm. Notably,
~23.3% (14/60) of normal colorectal tissue showed positive staining
of FGFR4. However, 50.0% (50/100) of CRC cases showed positive
FGFR4 staining in tumor cells. The expression of FGFR4 in CRC was
significantly higher compared with in normal tissues (Fig. 1B; P<0.0001). Next, the
association between the expression of FGFR4 and the
clinicopathologic features of patients with CRC was evaluated.
Table I reveals that the
expression of FGFR4 in cancer cells was not significantly
associated with sex, age, lymph node metastasis, clinical stage,
invasion depth and vessel invasion.
 | Table IAssociation between FGFR4 expression
in CRC tissues and clinicopathological features. |
Table I
Association between FGFR4 expression
in CRC tissues and clinicopathological features.
| Clinicopathologic
features | FGFR4 expression
| P-value |
|---|
| High (n=47) | Low (n=48) |
|---|
| Sex | | | 0.7668 |
| Female | 26 | 28 | |
| Male | 21 | 20 | |
| Age, years | | | 0.3245 |
| <60 | 15 | 20 | |
| ≥60 | 32 | 28 | |
| Lymph node
metastasis | | | 0.7375 |
| N0 | 29 | 28 | |
| N1+N2 | 18 | 20 | |
| Clinical stage | | | 0.7375 |
| I+II | 29 | 28 | |
| III+IV | 18 | 20 | |
| Invasion depth | | | 0.7715 |
| T2+T3 | 17 | 16 | |
| T4 | 30 | 32 | |
| Vessel
invasion | | | 0.3177 |
| No | 40 | 44 | |
| Yes | 7 | 4 | |
Fig. 1C indicates
the Kaplan-Meier survival curve for patients with colorectal
cancer. Subjects with FGFR4 high or low expression did not exhibit
significant differences in overall survival (P=0.1394). However,
patients with FGFR4 high-expression had a lower 5-year survival
time compared with patients with FGFR4 low-expression (64 vs. 74%).
Subgroup analysis revealed the overall survival time of patients
with FGFR4 high-expression at stages Tumor (T)2 and T3 was
significantly poorer compared with those with FGFR4 low-expression
(P=0.0283). Univariate Cox regression analysis demonstrated that
the overall survival rate was significantly associated with lymph
node metastasis, clinical stage, invasion depth and vessel invasion
(Table II; P<0.05). All
statistically significant variables identified in the univariate
analysis were analyzed using multivariate Cox regression.
Multivariate Cox regression analysis revealed that vessel invasion
was an independent risk factor for overall survival time of
patients, whereas FGFR4 was not (Table II, P=0.004).
| Table IIUnivariate and multivariate Cox
regression analysis of the overall survival rate of patients with
CRC. |
Table II
Univariate and multivariate Cox
regression analysis of the overall survival rate of patients with
CRC.
| Parameter | Univariate
| Multivariate
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age, years | | | | | | |
| ≥60 | 1.745 | 0.814-3.741 | 0.152 | | | |
| Sex | | | | | | |
| Female | 1.392 | 0.710-2.727 | 0.335 | | | |
| FGFR4 high
expression Lymph node metastasis | 1.678 | 0.847-3.324 | 0.138 | 1.436 | 0.698-2.952 | 0.138 |
| Lymph node
metastasis N1+N2 | 2.351 | 1.282-4.999 | 0.007a | 1.65 | 0.757-3.600 | 0.208 |
| Invasion depth | | | | | | |
| Lymph node
metastasis T4 | 2.877 | 1.190-6.954 | 0.019a | 2.580 | 0.961-6.929 | 0.06 |
| Vessel
invasion | | | | | | |
| Lymph node
metastasis Yes | 3.393 | 1.699-8.470 | 0.001a | 3.734 | 1.526-9.138 | 0.004a |
| Clinical stage | | | | | | |
| Lymph node
metastasis III+IV | 2.351 | 1.282-4.999 | 0.007a | 1.65 | 0.757-3.600 | 0.208 |
Selection of a lentivirus vector
efficiently inhibiting FGFR4 expression in SW620 cells
To investigate the role of FGFR4 in the
tumorigenesis of colorectal cancer lentiviral vectors were
constructed containing four specific FGFR4 shRNAs (LV3-497,
LV3-1750, LV3-2074 and LV3-1670) to silence the expression of
FGFR4. Scrambled shRNA was used as a negative control. Fig. S1A depicts the construction of LV3
vectors. To determine the lentiviral transduction efficiency of
SW620 cells, green fluorescence protein (GFP) expression was
observed by fluorescence microscopy at 72 h after the transfection.
Lentivirus transfection in SW620 cells was demonstrated to be
efficient (Fig. S1B). Western
blot quantitative analysis indicated that FGFR4 protein expression
in the LV3-1750 group was lowest of the four FGFR4 shRNAs, compared
with NC group and mock group (Fig.
S1C). The above results indicated that LV3-1750 was the most
effective and specific in the silencing of FGFR4, so it was
selected for subsequent functional assays.
Effects of FGFR4 downregulation on
migration, invasion and proliferation of SW620 Cells
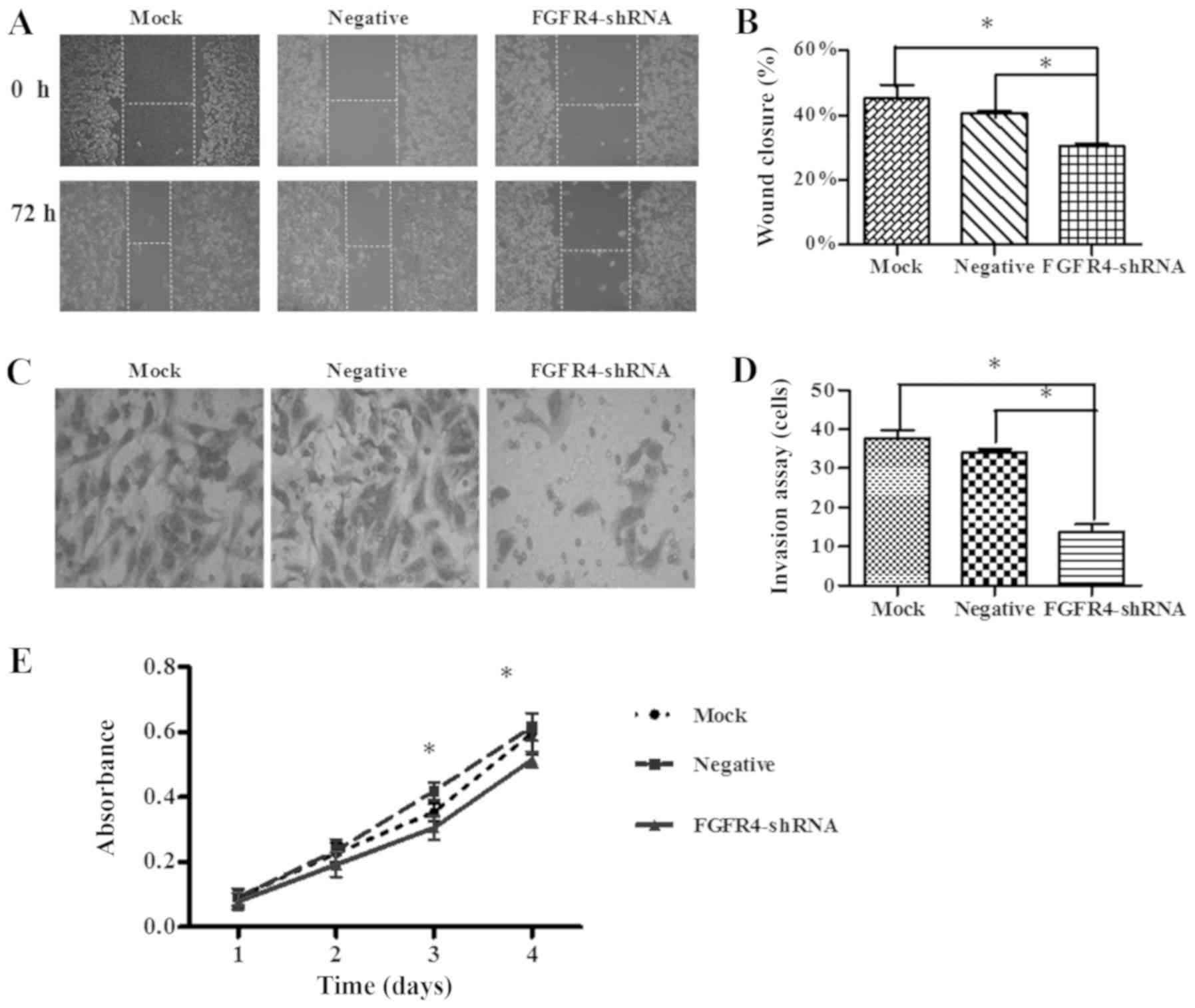
The wound heal assay revealed that SW620 cells
migrated into the wound scratch under a microscope (Fig. 2A). The wound closure was not
significantly different between the mock group and the negative
control group. However, following FGFR4-shRNA transfection, the
wound closure was significantly decreased (Fig. 2B; P<0.05). This indicates that
knockdown of FGFR4 is able to suppress the migration ability of
SW620 cells.
The Transwell invasion assay revealed that the
number of migrating cells in the mock group, negative group and the
FGFR4-shRNA group were 37.67±2.08, 34±1.00 and 13.67±2.08,
respectively. In the FGFR4-shRNA group, the number of invaded SW620
cells was significantly less compared with in the mock and NC
groups (Fig. 2C and D, P<0.05).
However, the numbers of migrated cells in the negative group and
the mock group were not significantly different.
An MTT assay was used to investigate the role of
FGFR4 in cell proliferation. As demonstrated in Fig. 2E, transfection with FGFR4-shRNA
significantly inhibited the proliferation ability of SW620 cells
when compared with the negative and mock group (P<0.05), at 3-
and 4-days post-transfection.
Effects of FGFR4 downregulation on cell
apoptosis and the expression of apoptotic-related proteins in SW620
cells
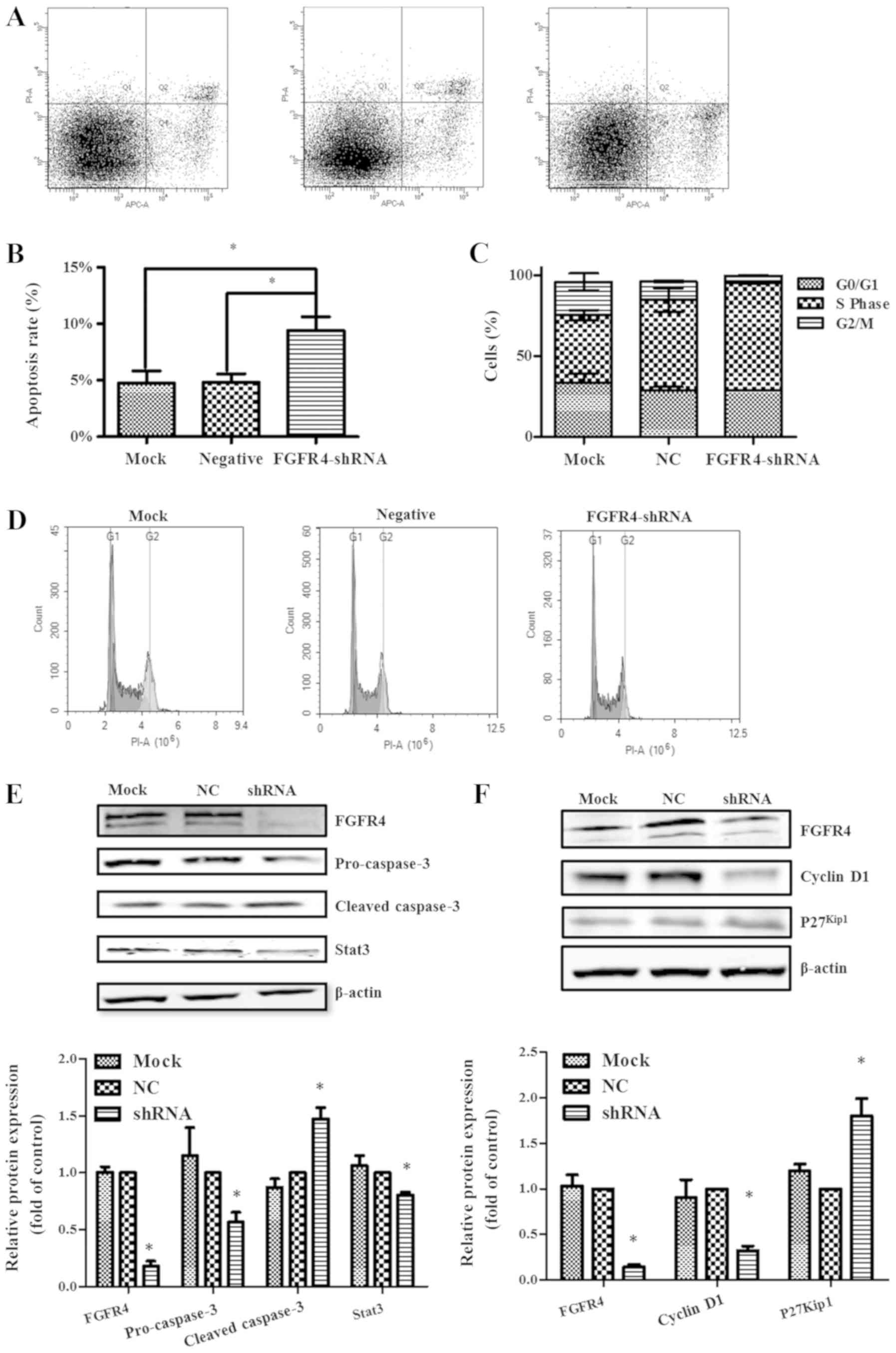
After 72 h of transfection of SW620 cells with FGFR4
shRNA, flow cytometry was used to assess the change in apoptotic
rate. In SW620 cells, the apoptosis rates in the FGFR4-shRNA, NC
and mock group were 9.40±1.21%, 4.83±0.74% and 4.77±1.05%,
respectively (Fig. 3A). As
indicated in Fig. 3B, knockdown of
FGFR4 significantly increased the apoptosis rate of SW620 cells
(P<0.05), but there was no significant difference between the
mock and NC group (Fig. 3B).
In addition, apoptosis-associated proteins Stat3 and
caspase3 were used to assess the apoptotic capacity of SW620 cells
with different treatments. In SW620 cells treated with FGFR4-shRNA,
pro-caspase3 and Stat3 expression decreased and cleaved-caspase3
expression increased compared with both the NC group and the mock
group (Fig. 3E).
Effects of FGFR4 downregulation on cell
cycle distribution and cycle-related proteins in SW620 cells
After 72 h of transfection of SW620 cells with FGFR4
shRNA, flow cytometry was used to assess the change in cell cycle
distribution (Fig. 3D). It was
revealed that knockdown of FGFR4 increased the number of S-phase
cells and decreased cells in the G2/M phase (Fig. 3C). The above results revealed that
knockdown of FGFR4 blocked SW620 cells in the S phase.
Subsequently, western blotting was used to detect cell cycle
related proteins. The expression of p27Kip1 in
FGFR4-shRNA group was significantly increased and the expression of
Cyclin D1 was significantly decreased, compared with both the NC
control group and mock group.
FGFR4 influences the EMT process via the
Wnt/β-catenin signaling pathway
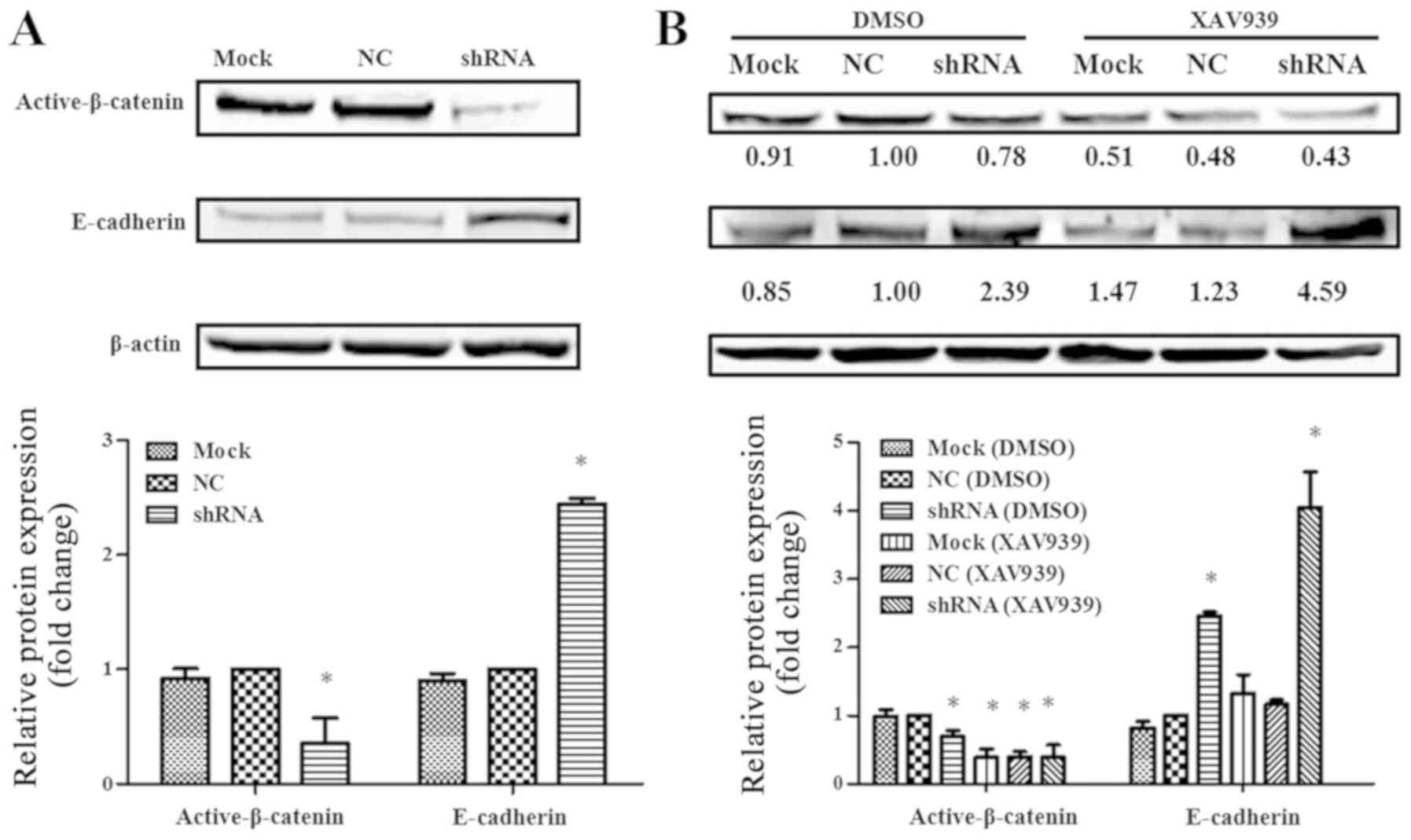
To explore whether the effect of FGFR4 on EMT
process through the Wnt/β-catenin signaling pathway, western
blotting was used to detect the levels of E-cadherin and active
β-catenin. As indicated in Fig.
4A, knockdown of FGFR4 in SW620 cells increased the level of
E-cadherin but decreased the level of active β-catenin. Following
XAV939 treatment, the levels of activated β-catenin in the mock, NC
and FGFR4-shRNA groups were decreased, and were most decreased in
FGFR4-shRNA group. Notably, E-cadherin levels were increased in the
mock, NC and FGFR4-shRNA groups, with the highest increase in the
FGFR4-shRNA group (Fig. 4B). Thus,
it was hypothesized that in colorectal cancer cells, activation of
FGFR4 increases the activity of β-catenin protein. Activated
β-catenin protein translocates to the nucleus and initiates the
expression of genes, followed by the suppression of E-Cadherin
expression.
Discussion
FGFRs belong to the family of receptor tyrosine
kinases (RTKs), all of which have an extracellular ligand-binding
domain, a transmembrane domain and a split intracellular tyrosine
kinase domain (4). The
ligand-binding domain of the receptor consists of three
immunoglobulin-like domains I, II and III. Immunoglobulin-like
domain III is the site to which FGF binds, and domain III
transcripts of FGFR1-3 undergo alternative splicing (20). Unlike FGFR1-3, FGFR4 does not
alternate splicing in this region (21). By contrast, FGFR4 mutations are
rare in tumor cells, but upregulation of FGFR4 is more common. To
investigate the role of FGFR4 in tumorigenesis, development and
metastasis, the expression level of FGFR4 in colorectal cancer
tissues was examined, then FGFR4 expression was silenced in SW620
colorectal cancer cells and a series of functional assays were
performed. Finally, the mechanism of action of FGFR4 in EMT was
explored.
In the present study, the expression of FGFR4 in CRC
was significantly higher compared with in normal tissues (Fig. 1B, P<0.0001). This is similar to
results reported in nasopharyngeal (22), liver (23), ovarian (8), melanoma (24) and breast cancer (25). Therefore, FGFR4 expression levels
may provide valuable clues for the diagnosis of colorectal cancer.
Table I reveals that the
expression level of FGFR4 was not significantly associated with
histopathological parameters. Combined with follow-up data,
patients with FGFR4 high-expression exhibited a lower 5-year
survival time than patients with FGFR4 low-expression (64 vs. 74%),
although the difference was not statistically significant. However,
Li et al (26) reported
that FGFR4 positivity was correlated with shortened disease free
survival (DFS) and overall survival (OS), and that high expression
of FGFR4 is an independent prognostic factor for colorectal cancer
(27). The differences between the
two sets of data may be caused by several factors. Primarily, in
the present study, the colorectal cancer tissue sample size was
relatively small compared to the aforementioned study (n=160 vs.
n=316, respectively); secondly, a tissue microarray was used for
immunohistochemistry, tissue samples on the microarray experimental
conditions are exactly the same, with excellent quality control;
and thirdly, the present study had a longer follow-up time.
Notably, subgroup analysis revealed that patients at T2 and T3 with
FGFR4-positive expression had less favorable overall survival times
(P=0.0283).
An article published previously revealed that FGFR4
protein expression is higher in HCT116 and SW620 cell lines
compared with LS47, DLD1 and SW116 (7). To investigate the role of FGFR4 in
the tumorigenesis of colorectal cancer, lentiviral vectors were
constructed containing four specific FGFR4 shRNAs (LV3-497,
LV3-1750, LV3-2074 and LV3-1670) to silence the expression of FGFR4
in SW620 and HCT116. However, it was revealed that the four
specific FGFR4 shRNAs were not able to silence the expression of
FGFR4 in HCT116 cells (data not shown). Therefore, the present
study aimed to evaluate the function of FGFR4 in the SW620. FGFR4
was silenced using shRNA and then flow cytometry was used to detect
the apoptotic rate of the cells. The apoptosis rate of SW620 cells
treated with shRNA was significantly increased. Western blot
analysis was used to detect the expression of apoptosis-associated
proteins. Consistent with the appeal, the cleaved-caspase3
expression was significantly increased in the FGFR4-shRNA group,
whereas the expression of Stat3 and pro-caspase3 was significantly
decreased, with no significant difference between the Mock and NC
groups. It was also demonstrated that down-regulation of FGFR4
blocked SW620 cells in S phase. Compared with the mock and NC
groups, CyclingD1 as a positive regulator of the cell cycle was
significantly downregulated in the FGFR-shRNA group. In the cell
cycle regulatory mechanism, p27 is one of the most critical
inhibitors, and inhibits the majority of CDK and cyclin complex
kinase activity (27). When FGFR4
was silenced, the expression of p27kip1 increased. MTT
assays showed that silencing of FGFR4 decreased the rate of
proliferation of SW620 cells. Shi et al (22) revealed that as the cell cycle
progresses, the expression of FGFR4 increased; the downregulation
of FGFR4 suppressed the proliferation of nasopharyngeal carcinoma
cells. Therefore, it was hypothesized that FGFR4 may affect the
proliferation of colorectal cancer cells by promoting apoptosis and
inhibiting the cell cycle.
Tumor recurrence and metastasis adversely affect the
prognosis of patients. In the current study, FGFR4 was revealed to
serve an important role in the migration and invasion abilities of
colorectal cancer cells. Wound scratch assays and transwell
migration assays revealed that silencing FGFR4 significantly
reduced the ability of SW620 cells to migrate and invade. EMT is a
biological process by which epithelial cell-derived malignant cells
adopt the ability to migrate and invade (12) EMT causes epithelial cells to lose
the phenotype of epithelial cells and affects the expression of
cell adhesion proteins such as E-Cadherin. Our previous study
demonstrated that FGFR4-specific inhibitor Blu9931 induced the high
expression of E-Cadherin in colorectal cancer cells (7). One study demonstrated that the
FGFR4/glycogen synthase kinase (GSK3β)/β-catenin axis may serve a
key role in FGF19-induced EMT in HCC cells (14). Peláez-García et al (2) reported that FGFR4 silencing promoted
the expression of E-cadherin on the surface of colorectal cancer
cells via immunofluorescence analysis. FGFR4 may promote the
migration and invasion of colorectal cancer cells via the EMT
process.
However, the mechanism by which FGFR4 influences the
EMT process is yet to be elucidated. The Wnt/β-catenin pathway and
EMT process are closely associated (15,16).
Disruption of the APC/Axin/GSK-3β complex reduces β-catenin
degradation followed by activation of β-catenin into the nucleus,
which is a key step in activating Wnt signaling (28). In the current study, after cells
were treated with XAV939 (an inhibitor of β-catenin), E-Cadherin
protein expression significantly increased compared with the no
treatment FGFR4-shRNA group, while the active-β-catenin expression
was significantly reduced. In colorectal cancer cells, activation
of FGFR4 increases the activity of β-catenin protein. FGFR4 may
influence the expression of E-Cadherin via the regulation of
β-catenin. So, it is hypothesized that FGFR4 promotes EMT
progression via activating the Wnt/β-catenin pathway. A similar
result was observed in liver cancer cells; after FGF19 agonized
FGFR4, FGFR4 further activated the Wnt/β-catenin pathway and
promoted EMT progression (14).
The present study also had certain limitations. In
the cell cycle regulatory mechanism, p27 is one of the most
critical inhibitors. Other checkpoint proteins such as cyclin B,
cyclin E, p21 and p53 should be detected. In the current study,
XAV939 was used as an inhibitor of β-catenin to study the
Wnt/β-catenin pathway. The addition of agonists to the
Wnt/β-catenin signaling pathway should also be conducted to further
investigate the Wnt pathway. Detection of other associated proteins
influencing the Wnt/β-catenin pathway, such as N-Cadherin would
lend support to the results of the current study. In a subsequent
study, agonists of the Wnt/β-catenin signaling pathway should be
investigated, along with the expression of other checkpoint
proteins of the cell cycle. To further investigate the role of
FGFR4 in colorectal cancer, tumor growth analysis in nude mice
should be performed.
In conclusion, in the present study it was
demonstrated that FGFR4 is upregulated in human colorectal cancer
tissues and appears to be associated with the prognosis of patients
with colorectal cancer. It was also observed that silencing FGFR4
inhibited SW620 cell proliferation, arrested cell cycle and
promoted apoptosis. In addition, the reversal of EMT progression
following FGFR4 silencing may be mediated via the Wnt/β-catenin
pathway. Therefore, the results indicate that FGFR4 may be an
effective target for the treatment of patients with colorectal
cancer, and may also represent a useful marker.
Supplementary Data
Abbreviations:
|
CRC
|
colorectal cancer
|
|
FGFR4
|
fibroblast growth factor receptor
4
|
|
EMT
|
epithelial-mesenchymal transition
|
Acknowledgements
Not applicable.
Funding
The present study was supported by the Department of
Gastrointestinal Surgery and Institute of Clinical Medicine, The
First Affiliated Hospital, Zhengzhou University (grant no.
SBGJ2018010) and National Natural Science Foundation of China
(grant no. 81201955).
Availability of data and materials
The datasets generated and analyzed during this
study are presented in the Figures and Additional Files.
Authors' contributions
YWY and DBJ conceived and designed the study. DBJ,
JL, FW, CH and XRW collected the data. YWY, DBJ, JJL and JL
analyzed the data. DBJ, XRW, CH and JL performed the experiments.
YWY and CH provided the resources and supervised the study. DBJ and
JL wrote the original draft. YWY, FW, CH, XRW and JJL reviewed and
edited the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of the Shanghai Outdo Biotech Co., Ltd. (Shanghai,
China). The patients were recruited by several hospitals in
Shanghai, and all provided written informed consent for research.
The study was undertaken in accordance with the ethical standards
of the World Medical Association Declaration of Helsinki.
Patient consent for publication
Informed written consent to publish was provided by
each patient that was recruited.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peláez-García A, Barderas R, Torres S,
Hernández-Varas P, Teixidó J, Bonilla F, de Herreros AG and Casal
JI: FGFR4 role in epithelial-mesenchymal transition and its
therapeutic value in colorectal cancer. PLoS One. 8:e636952013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Turner N and Grose R: Fibroblast growth
factor signalling: From development to cancer. Nat Rev Cancer.
10:116–129. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eswarakumar VP, Lax I and Schlessinger J:
Cellular signaling by fibroblast growth factor receptors. Cytokine
Growth Factor Rev. 16:139–149. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang J, Stockton DW and Ittmann M: The
fibroblast growth factor receptor-4 Arg388 allele is associated
with prostate cancer initiation and progression. Clin Cancer Res.
10:6169–6178. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ye YW, Zhou Y, Yuan L, Wang CM, Du CY,
Zhou XY, Zheng BQ, Cao X, Sun MH, Fu H, et al: Fibroblast growth
factor receptor 4 regulates proliferation and antiapoptosis during
gastric cancer progression. Cancer. 117:5304–5313. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang D, Li J, Li J, Wang M, Han C, Wang
X, Zhao C and Ye Y: Combination of FGFR4 inhibitor Blu9931 and
5-fluorouracil effects on the biological characteristics of
colorectal cancer cells. Int J Oncol. 51:1611–1620. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zaid TM, Yeung TL, Thompson MS, Leung CS,
Harding T, Co NN, Schmandt RS, Kwan SY, Rodriguez-Aguay C,
Lopez-Berestein G, et al: Identification of FGFR4 as a potential
therapeutic target for advanced-stage, high-grade serous ovarian
cancer. Clin Cancer Res. 19:809–820. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sawey ET, Chanrion M, Cai C, Wu G, Zhang
J, Zender L, Zhao A, Busuttil RW, Yee H, Stein L, et al:
Identification of a therapeutic strategy targeting amplified FGF19
in liver cancer by Oncogenomic screening. Cancer Cell. 19:347–358.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miura S, Mitsuhashi N, Shimizu H, Kimura
F, Yoshidome H, Otsuka M, Kato A, Shida T, Okamura D and Miyazaki
M: Fibroblast growth factor 19 expression correlates with tumor
progression and poorer prognosis of hepatocellular carcinoma. BMC
Cancer. 12:562012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Punt CJ and Tol J: More is less -
combining targeted therapies in metastatic colorectal cancer. Nat
Rev Clin Oncol. 6:731–733. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Paz H, Pathak N and Yang J: Invading one
step at a time: The role of invadopodia in tumor metastasis.
Oncogene. 33:4193–4202. 2014. View Article : Google Scholar :
|
|
13
|
Pez F, Lopez A, Kim M, Wands JR, Caron de
Fromentel C and Merle P: Wnt signaling and hepatocarcinogenesis:
Molecular targets for the development of innovative anticancer
drugs. J Hepatol. 59:1107–1117. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao H, Lv F, Liang G, Huang X, Wu G,
Zhang W, Yu L, Shi L and Teng Y: FGF19 promotes
epithelial-mesenchymal transition in hepatocellular carcinoma cells
by modulating the GSK3β/ β-catenin signaling cascade via FGFR4
activation. Oncotarget. 7:13575–13586. 2016.
|
|
15
|
Zhao JH, Luo Y, Jiang YG, He DL and Wu CT:
Knockdown of β-Catenin through shRNA cause a reversal of EMT and
meta-static phenotypes induced by HIF-1α. Cancer Invest.
29:377–382. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schlessinger J: Cell signaling by receptor
tyrosine kinases. Cell. 103:211–225. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang SM, Mishina YM, Liu S, Cheung A,
Stegmeier F, Michaud GA, Charlat O, Wiellette E, Zhang Y, Wiessner
S, et al: Tankyrase inhibition stabilizes axin and antagonizes Wnt
signalling. Nature. 461:614–620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Waaler J, Machon O, Tumova L, Dinh H,
Korinek V, Wilson SR, Paulsen JE, Pedersen NM, Eide TJ, Machonova
O, et al: A novel tankyrase inhibitor decreases canonical Wnt
signaling in colon carcinoma cells and reduces tumor growth in
conditional APC mutant mice. Cancer Res. 72:2822–2832. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Edge SB, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: AJCC cancer staging manual. 7th edition.
Springer; New York, NY: 2010
|
|
20
|
Zhang X, Ibrahimi OA, Olsen SK, Umemori H,
Mohammadi M and Ornitz DM: Receptor specificity of the fibroblast
growth factor family. The complete mammalian FGF family. J Biol
Chem. 281:15694–15700. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Haugsten EM, Wiedlocha A, Olsnes S and
Wesche J: Roles of fibroblast growth factor receptors in
carcinogenesis. Mol Cancer Res. 8:1439–1452. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi S, Li X, You B, Shan Y, Cao X and You
Y: High Expression of FGFR4 Enhances Tumor Growth and Metastasis in
Nasopharyngeal Carcinoma. J Cancer. 6:1245–1254. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ho HK, Pok S, Streit S, Ruhe JE, Hart S,
Lim KS, Loo HL, Aung MO, Lim SG and Ullrich A: Fibroblast growth
factor receptor 4 regulates proliferation, anti-apoptosis and
alpha-fetoprotein secretion during hepatocellular carcinoma
progression and represents a potential target for therapeutic
intervention. J Hepatol. 50:118–127. 2009. View Article : Google Scholar
|
|
24
|
Streit S, Mestel DS, Schmidt M, Ullrich A
and Berking C: FGFR4 Arg388 allele correlates with tumour thickness
and FGFR4 protein expression with survival of melanoma patients. Br
J Cancer. 94:1879–1886. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meijer D, Sieuwerts AM, Look MP, van
Agthoven T, Foekens JA and Dorssers LC: Fibroblast growth factor
receptor 4 predicts failure on tamoxifen therapy in patients with
recurrent breast cancer. Endocr Relat Cancer. 15:101–111. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li CS, Zhang SX, Liu HJ, et al: Fibroblast
growth factor receptor 4 as a potential prognostic and therapeutic
marker in colorectal cancer. Biomarkers. 19:81–85. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Conradie R, Bruggeman FJ, Ciliberto A,
Csikász-Nagy A, Novák B, Westerhoff HV and Snoep JL: Restriction
point control of the mammalian cell cycle via the cyclin E/Cdk2:p27
complex. FEBS J. 277:357–367. 2010. View Article : Google Scholar
|
|
28
|
MacDonald BT, Tamai K and He X:
Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev
Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|