1. Introduction
Glioma is the most common primary tumour of the
central nervous system, and its incidence accounts for ~80% of
primary brain malignancies. Glioma is a high-grade malignancy and
has a poor prognosis. Despite various high-intensity treatments,
including surgery combined with chemoradiation, the median survival
time of patients with glioblastoma (GBM) is only 12-15 months, and
only 3-5% of the patients survive for >3 years (1-3). In
recent years, genetic diagnosis and targeted therapy have been
attracting widespread attention as emerging research hotspots, and
studies have confirmed that the targeting of non-coding RNAs in
glioma may be more efficacious. Non-coding RNAs are regulatory
factors that participate in embryonic development, inflammatory
response, metabolism and chemotherapy resistance. Additionally,
tumour development, cell invasion, proliferation and apoptosis are
also closely associated with the expression and regulation of
non-coding RNAs (4-6).
Circular RNAs (circRNAs) are important members of
the non-coding RNA family, and they are single-stranded closed
circular RNA molecules without a 5′-end cap or a 3′-end poly(A)
tail, which are formed by covalent bonding. Due to their special
circular stable structure, circRNAs cannot be degraded by RNase R,
and they are highly evolutionarily conserved. When circRNAs were
first discovered in the 1970s, they were considered as 'noise'
resulting from incorrect splicing. However, with the rapid
development of high-throughput sequencing-based molecular
biotechnology in recent years, several circRNAs have been detected
in eukaryotic cells. As regards the gene expression of human cells,
circRNA molecules have more general expression characteristics
compared with their linear counterparts (7,8).
Moreover, circRNAs are abundant and stable. A large number of
studies have confirmed that circRNAs are differentially expressed
in various tumour cells (9,10).
Their regulatory effects on tumourigenesis, tumour development and
corresponding biological behaviours require further in-depth
research. The aim of the present review was to focus on the
research progress in this field, in order to provide a theoretical
and literature basis for further exploring the association between
the expression of circRNAs and gliomas. The findings of the study
may prove to be of value for the early diagnosis, pathological
grading, targeted therapy and prognostic evaluation of gliomas.
2. History of circRNAs
In the 1970s, Sanger et al discovered the
presence of circRNAs in RNA viruses (11). In 1979, Hsu and Coca-Prados first
observed, by means of electron microscopy, that RNA in the
cytoplasm of eukaryotic cells may exist in a circular form
(12). One year later, Arnberg
et al also observed the presence of circRNAs while studying
the components of yeast mitochondria (13). In 1993, Cocquerelle et al
reported that there were several exon-derived circRNAs in human
cell transcripts (14). During the
early years of circRNA discovery, circRNAs were considered
non-functional, lowly expressed RNA molecules resulting from
'mis-splicing' of exon transcripts. Due to this interpretation, the
depth and breadth of circRNA research has been inconsistent. Up
until the beginning of the 21st century, scientists had identified
no more than 10 types of circRNAs. However, in recent years, with
the rapid development of molecular biology technology and
bioinformatics analysis based on RNA sequencing (RNA-seq),
scientists have identified several exon-derived transcripts that
form circRNAs by non-linear reverse splicing or gene
rearrangements. These transcripts account for a large proportion of
the entire splicing transcript. In 2012, Salzman et al
discovered hundreds of circRNAs and established that they are
closely associated with human gene expression (15). Jeck and Sharpless identified
~25,000 circRNAs through RNA detection in human fibroblasts
(16). Memczak et al
compared the RNA-seq results with the human leukocyte database and
found 1,950 human circRNAs, 1,903 mouse circRNAs (81 circRNAs were
the same as human circRNAs), and 724 nematode circRNAs (17). Guo et al conducted deep
sequencing on 39 biological samples related to human cell lines and
found >7,000 circRNAs (18). In
2013, two major studies on circRNAs were published in Nature
(17,19). Since then, numerous related studies
have been published, and circRNAs have come to represent a new
direction in the field of non-coding RNA worldwide.
3. Basic characteristics of circRNAs
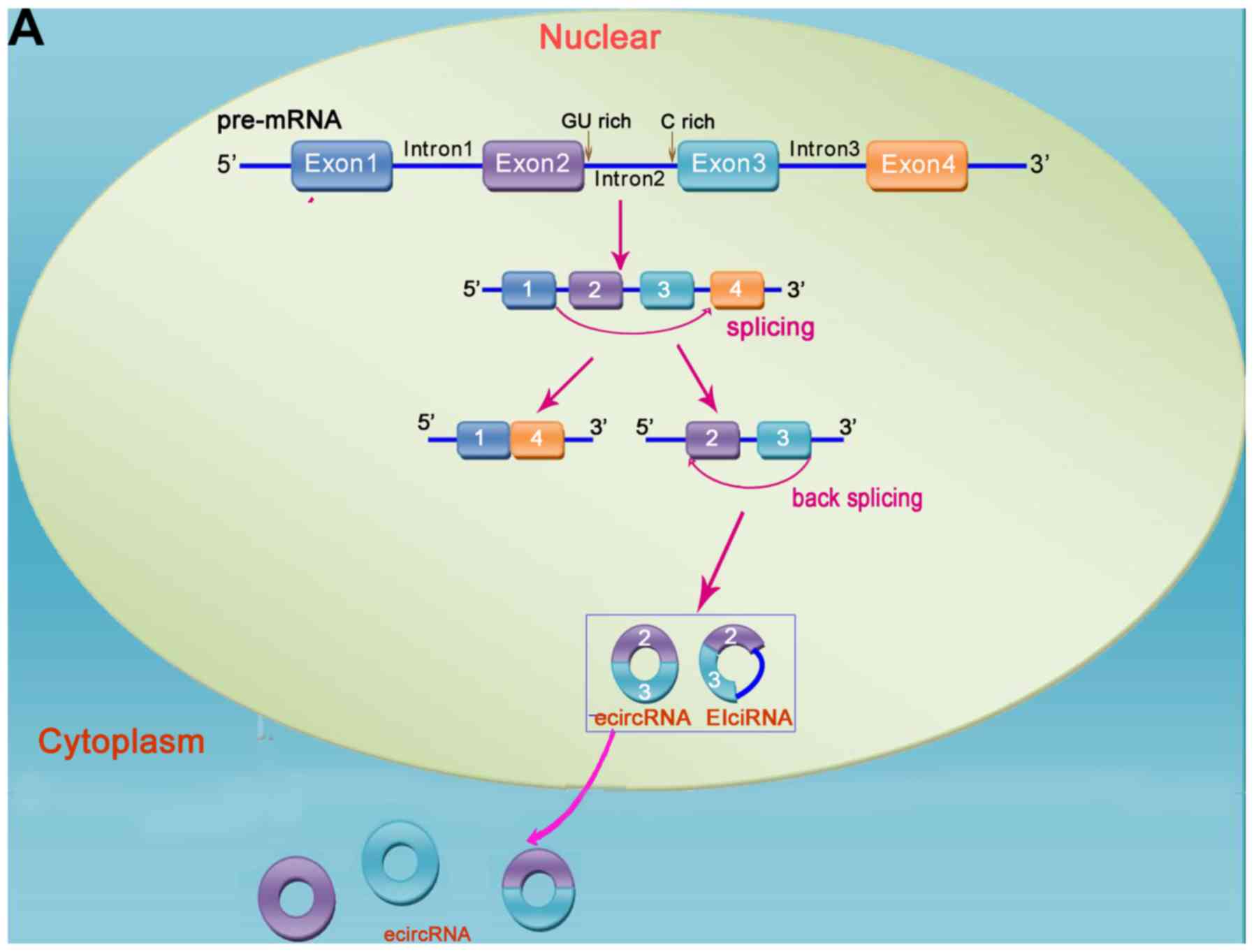
Novel and unique circRNAs
circRNAs are generated from variable splicing. The
majority are formed by the circularization of exons, and a few are
derived from introns (Fig. 1). The
majority of circRNAs are located in the cytoplasm of eukaryotic
cells, but a small proportion are located in the nucleus (mainly
intron-derived circRNAs). They are specific per tissue type,
disease type and chronological order; overall, they are highly
evolutionarily conserved, although there are also certain
evolutionary changes (20,21).
'Tailless' circRNAs
The conventional 5′-end cap and 3′-end poly(A) tail
structure in linear RNA molecules are absent in circular RNAs due
to their closed circular structure. As one of the key steps in
classical RNA detection methods (RNA extraction) the principle of
isolating RNA depends on the structure of the poly(A) tail. This
may be an important reason that the research on circRNA molecules
was overlooked prior to the rapid development of sequencing
technology.
Non-translated circRNAs
A number of circRNAs are produced by related genes
with the ability to encode proteins; however, to date, the vast
majority of circRNAs are still classified as new non-coding
RNAs.
Localization and stability of
circRNAs
The majority of circRNAs are located in the
cytoplasm and, occasionally, their expression level is >10 times
that of the corresponding linear isomers. The increased expression
may be explained by the fact that RNase R acts by recognizing the
end sites of linear RNA structures, and circRNAs have a special
'end-to-end' closed loop structure, which makes them highly
resistant to nucleases and degradation.
circRNAs act as competitive endogenous
RNAs (ceRNAs)
circRNAs are enriched in the cytoplasm and have the
same transcription sequence as their corresponding linear
counterparts. This characteristic also indirectly indicates that
circRNAs are likely to play biological roles by regulating the
corresponding linear RNAs. In fact, given the high abundance and
stability of circRNAs in specific tissues, they exhibit extremely
strong ceRNA activity when they interact with linear RNAs. It was
recently demonstrated that the newly discovered circRNA-CDR1as
(also referred to as ciRS-7) has >60 conserved binding sites for
miR-7. Experiments in zebrafish were later conducted, and their
results revealed that the expression of ciRS-7 may damage the
development of the midbrain. This effect was the same as that of
knocking out miR-7 (22).
4. The present and future of the biological
functions of cir-cRNAs
circRNAs are mainly located in the cytoplasm or
exosomes, and exhibit high stability and strong specificity in
terms of tissue, disease and timing. In recent years, with the
in-depth study of gene function and mechanisms, an increasing
number of studies have demonstrated that circRNAs have important
regulatory functions in the growth and development of organisms,
immune responses, inflammatory responses, and tumour development.
circRNAs are also becoming prominent in the field of disease
diagnostic markers in biomedical medicine. They have great
potential for clinical application, but the types of biological
functions and underlying mechanisms of action of circRNAs remain
unknown. To date, circRNAs have been found to have several
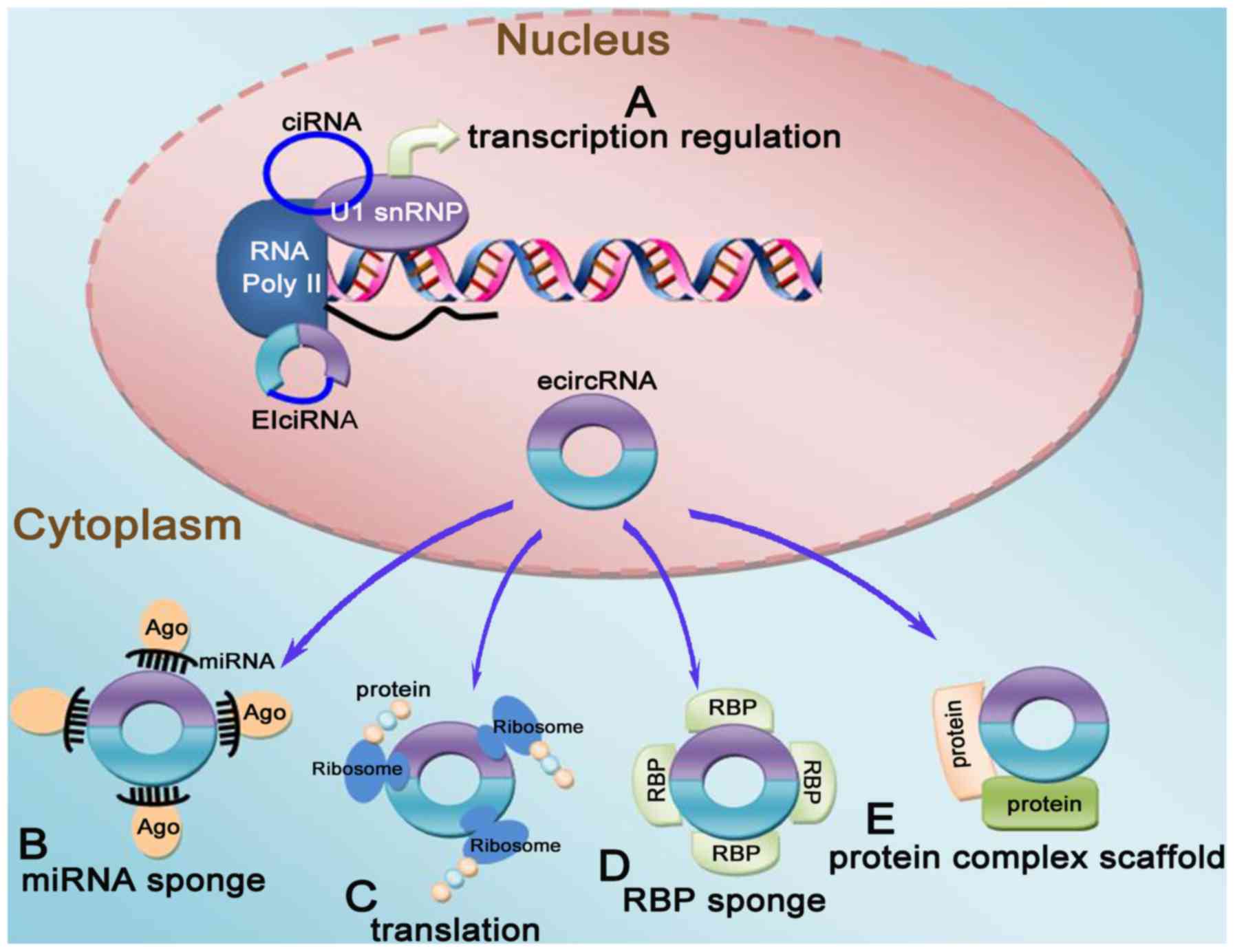
biological functions as follows (Fig.
2): i) miRNA sponging: circRNAs contain abundant miRNA-binding
sites, which are the structural basis for their molecular sponge
action, and inhibit the interaction between the 3′ untranslated
region of miRNAs and mRNAs, thereby indirectly affecting the
expression of downstream target genes and proteins of miRNAs.
However, other studies reported that most identified human and
mouse circRNAs rarely contain miRNA target sites, indicating that
these circRNAs may function in other manners that have yet to be
determined. Therefore, it was inferred that the main mode of
circRNA action may not be miRNA sponging, or that there may be
other unknown mechanisms (23).
ii) Binding of regulatory proteins: circRNAs can interact with
RNA-binding proteins (RBPs), which are regulated by mRNAs, and
change the splicing mode or regulate the stability of mRNAs. iii)
Regulation of gene transcription: circRNAs can interact with RNA
polymerase II, or exon-intron circRNA can first bind with a small
ribonucleoprotein and then with RNA polymerase II to play a role in
regulating gene transcription. iv) Potential encoding capabilities:
Although circRNAs were originally classified as non-coding genes,
certain circRNAs can be translated by ribosomes and encode
polypeptides based on the latest protein translation research.
Moreover, some circRNAs already have the ability to translate
proteins and then play roles corresponding to their related
proteins. These findings have opened up a new and fascinating
perspective in functional studies of circRNAs (24-26).
5. circRNAs in the central nervous
system
Compared with other tissues, circRNAs are more
diverse and abundant in the central nervous system. Rybak-Wolf
et al reported high-level dynamic expression of circRNAs in
mammalian brain tissue. The authors analysed the relevant data
based on RNA-seq in separate brain regions, and discovered 65,731
brain-specific circRNAs in primary neurons and isolated synaptic
samples (27). In addition, some
researchers have collected specific circRNAs in human and mouse
tissues and identified 89,137 brain-specific circRNAs in human
foetuses (28). In 2014, Westholm
et al analysed the structure and distribution
characteristics of circRNAs in the brain of Drosophila, and found
that the formation of circRNAs tends to surround the long intron
regions of brain tissue and to aggregate in ageing brain tissue
(29). In 2015, in a report
published by Professor You in the Journal of Nature Neuroscience,
his research team analysed the basic characteristics of central
nervous system circRNAs and confirmed that, based on animal
experiments, the expression of circRNAs in the central nervous
system is indeed higher compared with that in other systems, and
that the genes associated with synapses are more easily
circularized. Immediately after that, scientists used optimized
high-resolution fluorescence in situ hybridization (FISH) to
prove that circRNAs are indeed highly localized in the dendrites of
neurons (30). In the same year,
Rybak-Wolf et al reported the same conclusion in his studies
(27). By analysing the expression
of circRNAs in mammalian brain tissue, that research group
confirmed the abundance of circRNA molecules in brain tissue and
that the expression of circRNAs was positively associated with
developmental brain processes, particularly in synaptic structures.
They subsequently hypothesized that the expression of circRNAs may
differ among different anatomical structures of the brain, and
proceeded to analyse the expression of circRNAs in different
brain-related structures (striatum, prefrontal cortex, olfactory
cortex, cerebellum and hippocampal tissue). As predicted, the
expression profiles of circRNAs in different neural tissues were
indeed different (27). A report
published in Science in 2017 demonstrated that the loss of
mammalian circRNA loci directly led to miRNA imbalance, which
affected brain function (31).
In addition, the miRNA-mRNA-circRNA interaction
network may form the epigenetic basis of neurological diseases. For
example, miRNA-7 acts as a regulator of α-synuclein and ubiquitin
protein ligase A, which is a causative agent of chronic
neurodegenerative diseases, such as Alzheimer's disease,
Parkinson's disease, amyotrophic lateral sclerosis, multiple
sclerosis and multiple system atrophy. In addition, circRNAs also
play an important role in secondary brain damage following acute
central nervous system insults, such as stroke and neonatal
hypoxia-ischemia. Recent studies have also indicated a potential
link between depressive disorder or Moyamoya disease and circRNA
regulation (32,33). As regards why circRNAs are more
likely to be enriched in brain tissue, as mentioned earlier,
circRNAs are more likely to form near long introns; in the central
nervous system, important genes carry longer introns, which may be
the genetic basis for the formation and enrichment of circRNAs
(34). In addition, nerve-specific
RBPs are likely to be closely associated with the formation of
circRNAs. However, the mechanisms of action of some known RBPs,
such as splicing factor muscleblind, Quaking and
adenosine-to-inosine acting on RNA enzyme 1, which are associated
with circRNA formation, appear to contradict this hypothesis.
Therefore, scientists inferred that other unknown RBPs, which also
play important roles in promoting the formation and enrichment of
circRNAs, may be present in the central nervous system.
As regards the role of circRNAs in the nervous
system, an in vitro differentiation model was established to
systematically analyse the change trends in the expression of
circRNAs during the differentiation processes. The results
demonstrated that the expression of circRNAs suddenly increases
during the formation of synapses (33). A related study revealed that an
abrupt increase in circRNA expression levels occurs during the
transition from P10 to P30 in mice (30). Another study on circRNAs in the
human brain reported the same findings. Compared with the amount of
circRNAs during embryonic development, their levels increased
significantly in foetal and adult brains (27,28).
The synaptogenic function of circRNAs was also supported by their
host genes that are associated with synaptic function. On the other
hand, a number of brain-enriched circRNAs were found to be
associated with neurotransmitter function, neuron maturation and
synaptic activity. For example, circFoxo3 was shown to be highly
active in binding and deactivating cyclin-dependent kinase 2,
leading to disrupted cell cycle progression (35). In addition, circHomer 1 was proven
to modulate some of the structural changes at the synapse during
neuronal plasticity and development (30). Furthermore, it has been
demonstrated that 58% of cerebral circRNAs are developmentally
regulated, while only 2% of homologous linear isoforms have
exhibited this trend (36).
Interestingly, several developmentally regulated circRNAs also
exhibited sexual dimorphism in the brain and were observed to
target ageing-associated mRNAs (32,33).
6. circRNA expression in gliomas
In recent years, several non-coding RNAs associated
with the occurrence and development of gliomas have been
identified. The pattern and characteristics of circRNA expression
in gliomas have been attracting increasing attention in the field
of non-coding RNA research (37,38).
Glioma is a term referring to a large group of
central nervous system tumours, and its pathological classification
is variable, so the expression and regulation of circRNAs are also
different (39). Song et al
included 20 fresh-frozen de novo GBM samples, 19 normal
brain tissue samples and 7 samples of other oligodendrogliomas (3
WHO grade III and 4 WHO grade II) as the basis of library
preparation and sequencing (40).
The researchers found 572 highly expressed circRNAs in the 46
samples, 476 of which were differentially expressed between GBM and
normal brain tissue, and the expression of 468 circRNAs in normal
brain tissue was significantly upregulated compared with that in
GBM tissue. Even in normal brain tissue, the researchers observed
differential expression profiles of circRNAs among different
anatomical locations. The findings mentioned above provided an
important molecular and theoretical basis for the tissue- and
disease-specific expression patterns of circRNAs. Moreover, studies
on the correlation between the differential expression of circRNAs
and the pathological grade or prognosis of glioma also confirmed
the tissue specificity (41,42).
Wang et al analysed 33 pairs of matched
isocitrate dehydrogenase wild-type GBM and paracancerous tissues by
microarray (43). The results
demonstrated that, compared with paracancerous tissues, there were
254 upregulated circRNAs and 361 downregulated circRNAs
(>1.5-fold) in GBM tissues. In addition, 3 pairs of matched GBM
and para-cancerous tissues were also selected for microarray
analysis (lncRNAs, miRNAs and circRNAs), and 548 upregulated
circRNAs and lncRNAs (>1.5-fold) were identified. The expression
levels of circRNAs were further verified between 10 pairs of
matched gliomas and adjacent normal brain tissue and 2,709
differentially expressed circRNAs were identified. In total, 105
differentially expressed circRNAs were later discovered by
cross-matching the two abovementioned data sets (43). Xu et al downloaded 3 pairs
of RNA-seq-related data for gliomas and normal brain tissue from
the Gene Expression Omnibus database and analysed the expression of
circRNAs. A total of 5 different circRNA analysis tools were used
and 12 universally expressed circRNAs were identified (44).
In addition to the abovementioned studies based on
high-throughput screening technology, other studies have also
reported that circRNAs may be potential biomarkers for gliomas.
Barbagallo et al studied 56 paraffin-embedded GBM biopsy
samples and 7 normal brain tissue samples. The expression level of
circSMARCA5 was determined in the control group. The results
revealed that the expression of circ-SMARCA5 in GBM was
significantly lower compared with that in the control group, but
there was no significant difference in the linear mRNA level of the
host gene SMARCA5. Furthermore, the research team also analysed the
expression of miR-671 in 45 GBM samples and 5 GBM cell lines. The
results revealed the presence of the miR-671/CDR1as/miR-7 axis in
GBM, and that miR-671-5p overexpression significantly increased the
migration capacity of GBM cells, but exerted a less prominent
effect on cell proliferation (45). In addition, some researchers
analysed 31 glioma tumour samples and matched adjacent normal
tissues. Based on the obtained data and biological information
analysis, circCFH was found to be upregulated in WHO grade I-II and
III-IV glioma samples (46). Xie
also investigated the expression of hsa-circ-0012129 in 31 matched
glioma tumours and adjacent normal tissues, and the results
revealed that the expression of hsa-circ-0012129 was significantly
higher in glioma tissues compared with that in adjacent tissues
(47).
Bioinformatics predictive analysis and
biotin-labelled miR-442a pull-down assays have confirmed that the
blinding of miR-442a to circNT5E is significantly upregulated in
glioma samples. In a study by Wang et al, the interaction
between miR-442a and circNT5E was verified using RNA pull-down and
FISH techniques. The researchers had previously demonstrated that
circNT5E can bind to miR-442a and inhibit its function (48). In this case, the expression levels
of miR-442a and circNT5E were negatively correlated. This
represents the adsorption and inhibitory action of typical circRNAs
as miRNA sponges. However, in the earliest miRNA sponge models of
CDR1as and miR-7, the expression level of CDR1as was not affected
by miR-7, but was regulated by miR-671. The same research group
also observed an interaction between circNFIX and miR-34a-5p
through RNA co-immunoprecipitation, and confirmed that NOTCH1 is a
target of miR-34a-5p, i.e., when circNFIX is inhibited by siRNA,
the expression level of miR-34a-5p is upregulated, while NOTCH1 is
inhibited (44).
Thus far, although most studies have focused on the
differential expression of circRNAs in gliomas and some emerging
molecular circRNA species have also been discovered and identified,
the expression patterns of circRNAs in gliomas exhibit high
specificity and diversity among individuals. This indicates that
the expression pattern of circRNAs in glioma and normal brain
tissues requires a larger-sample in-depth study to verify.
7. circRNAs are involved in regulating the
life activities of glioma cells
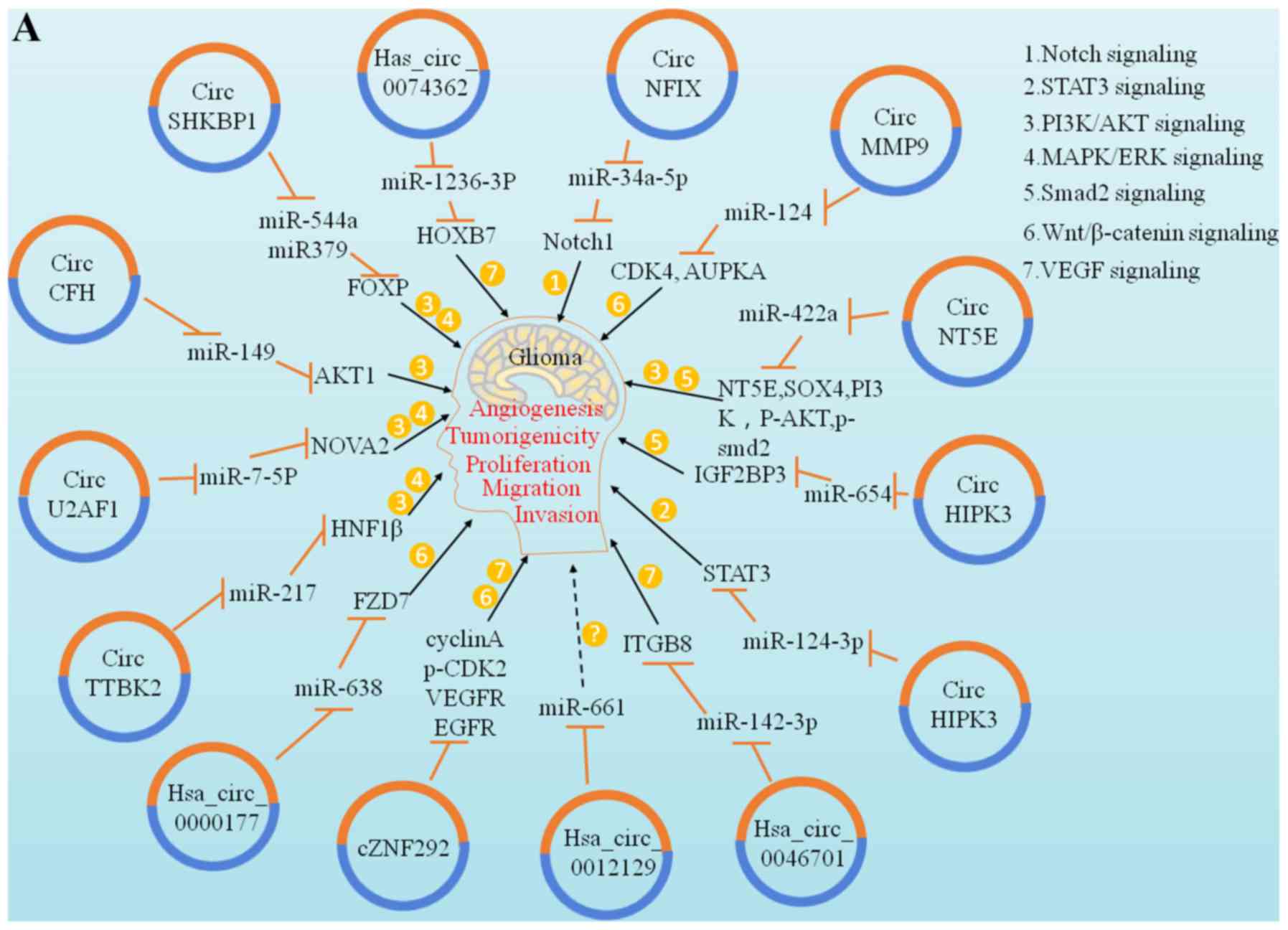
circRNAs and the glioma cell cycle
The ability of tumour cells to proliferate
indefinitely is the most important characteristic that
distinguishes them from normal cells. The overexpression of genes
and proliferation of tumour cells play important roles in the
regulation of the cell cycle. Numerous studies have demonstrated
that circRNAs may play the role of competitive endogenous RNAs
(ceRNAs) in tumour proliferation control. A study by Kumar et
al reported that has-circ-0046701 is over-expressed in glioma
tissues and cell lines. If has-circ-0046701 is knocked out, it can
significantly inhibit the proliferation and invasion of tumour
cells; further investigation revealed that it can promote the
expression of ITGβ8, a target of miR-142-3p, and regulate the
proliferation and invasion of glioma cells. Therefore, the
has-circ-0046701/miR-142-3p/ITGβ8 loop is considered to be involved
in regulating the occurrence and development of gliomas (49). The study of glioma cells by Liu
et al and Yang et al revealed that the cZNF292 gene
is associated with angiogenesis and that it is an important
carcinogenic circRNA that participates in the important process of
tumour cell lumen formation, whereas cZNF292 gene knockout can
inhibit cell proliferation and causes cell cycle arrest via the
Wnt/β-catenin signalling pathway (50,51)
(Fig. 3).
circRNAs and glioma cell apoptosis
Loss of apoptosis is another important
characteristic of tumour cells. It was recently reported that some
circRNAs are involved in regulating tumour cell apoptosis (52). The Bcl-2 protein family plays a key
role in the apoptotic pathway. The anti-apoptotic protein Bcl-2 is
located in the outer mitochondrial membrane and can inhibit the
secretion and release of pro-apoptotic factors, such as cytochrome
c. Bcl-2 is a target of miR-143. circRNAs also play an
important role in regulating the apoptotic pathway as miRNA
molecular sponges (53). A study
by Zhang et al revealed that the circular form of the snf2
histone linker PHD cyclohelicase (circ-SHPRH) can encode
SHPRH-146aa and protects SHPRH from degradation by ubiquitin
protease. Both are widely present and highly expressed in the
normal human brain. However, circ-SHPRH expression is significantly
reduced in GBM. Therefore, the low expression of SHPRH-146aa may
promote the malignant behaviour and tumourigenicity of tumour cells
in vivo (54). circ-SHPRH
generates 'UGA' stop codons to promote the translation of
SHPRH-146aa. It is hypothesized that circ-SHPRH and SHPRH-146aa are
tumour suppressors of human GBM and promote glioma cell apoptosis
(Fig. 3).
circRNAs and glioma cell invasion and
migration
Invasion and migration are hallmarks of malignant
tumours. Epithelial-to-mesenchymal transition (EMT) plays an
important role in regulating early invasion and migration in
several tumours. miRNAs are involved in the regulation of EMT
(55). circRNAs, acting as miRNA
sponges, may also participate in the regulation of EMT through
endogenous competitive mechanisms to affect tumour cell invasion
and migration. A study by Zheng et al demonstrated that
hepatocyte nuclear factor 1 (HNF1) is a direct target of miR-217
and is an important carcinogen at the glioma molecular level. In
addition, circ-TTBK2 is a circRNA gene that can bind to and
inactivate miR-217, thereby promoting the activation of the
phosphatidylinositol 3-kinase/protein kinase B-extracellular
signal-regulated kinase pathway through HNF-1β. Activation can
trigger the invasion of glioma cells, leading to a further increase
in tumour malignancy. When circ-TTBK2 is knocked out, miR-217
overexpression can inhibit tumour cell invasion and migration in
vivo and promote glioma cell apoptosis (56). Li et al observed that
has-circ-0007534 is highly expressed in glioma tissues and that its
knockdown can also inhibit the occurrence and development of
gliomas (57). ZIC5 has also been
found to be a target of miR-761 (57). Has-circ-0007534 can promote the
expression of ZIC5 by inhibiting miR-761 in glioma cells, thereby
markedly enhancing the proliferation and migration ability of
glioma cells. It has been reported that the expression level of
has-circ-0007534 is positively correlated with advanced glioma
grade (WHO III/IV) (57). In
addition, the degradation of has-circ-0007534 significantly
inhibited the proliferation and migration of glioma cells,
indicating that circRNAs may also play a role in the
differentiation of glioma cells (Fig.
3).
8. circRNAs promote the occurrence and
development of gliomas
circRNAs act as promoters in glioma
Song et al collected 46 samples of glioma
tissues and normal brain tissues for gene chip analysis. Among 572
measurable circRNAs, 476 exhibited a significantly different
expression between glioma and normal brain tissue. The expression
of circ_CDR1, circ_ATRNL1, circ_AKT3, circ_SPTAN1, circ_ZNF483,
circ_RIMS1, circ_FKBP8 and circ_UNC13C was low in glioma tissues,
while the expression of circ_COL1A2, circ_PTN, circ_VCAN, circ_SMO,
circ_PLOD2, circ_GLIS3 and circ_EPHB4 and circ_GLIP2 was high
compared with that in normal brain tissues (40).
Due to the strong specificity of the expression
pattern of circRNAs in gliomas, their regulatory effects on the
occurrence and development of gliomas have also attracted
significant attention. A series of circRNAs implicated in
carcinogenesis are involved in initiating the malignant progression
of glioma. For example, circNFIX is upregulated in GBM tissues and
cells. The inhibition of circNFIX can inhibit cell migration and
proliferation by downregulating the NOTCH1 pathway. This also
indicates that circNFIX may be involved in the occurrence and
development of gliomas (44). In
addition, circNT5E affects the proliferation, invasion and
migration ability of GBM cells in the interaction system by
stimulating miR-422a. This also provides further evidence of
circRNAs acting as molecular sponges (48).
It has been reported that the expression of
circ-TTBK2 is significantly increased in glioma tumour tissues and
corresponding cell lines, while the RNA expression of TTBK2,
corresponding to the linear structure, is not significantly
different. It may be hypothesized that the overexpression of
circ-TTBK2 plays a role in the proliferation, invasion and
migration of tumour cells. This also confirms that circ-TTBK2 plays
a key role in the progression of gliomas (56). A study by Chen and Duan
demonstrated that the inhibitory effect of miRNA on
hsa_circ_0000177 can significantly inhibit cell proliferation and
invasion, which affects the participation of the FZD7/Wnt7 pathway
in the progression of gliomas through sponge adsorption of miR-638
(58). In addition to the
abovementioned circRNAs, other studies have also reported that
hsa_circ_0046701 and circHIPK3 can promote the proliferation and
migration of glioma cells (59,60).
In addition to the role of circRNAs in promoting
cell proliferation and migration, circRNAs can also promote
angiogenesis. It was demonstrated that, compared with endothelial
cells exposed to astrocytes, the corresponding SHKBP1 mRNA of
circ-SHKBP1 is upregulated in U87 glioma-exposed endothelial cells
(GECs), and knocking out circ-SHKBP1 can significantly inhibit GEC
proliferation, migration and angiogenesis. circ-SHKBP1 affects the
proliferation of GECs through the miR-544a/FOXP1 and miR-379/FOXP2
pathways (61). In another study,
Yang et al reported that cZNF292 may be involved in the
angiogenesis of human gliomas and that cZNF292 circRNA inhibits
glioma angiogenesis via the Wnt/β-catenin pathway (51).
circRNAs as inhibitors in gliomas
The regulatory effect of several circRNAs on gliomas
is mediated via tumour suppression. A study by Zhang et al
demonstrated that circ-FBXW7 encodes FBXW7-185aa, a protein
consisting of 185 amino acids that can significantly inhibit tumour
cell processes in vivo and reduce the half-life of c-Myc by
antagonizing the stabilization effect of c-Myc induced by USP28. An
in situ GBM mouse model also demonstrated the
tumour-suppressive effect of FBXW7-185aa through IRES mutation. In
addition, circ-SHPRH can be translated into a 146-aa peptide called
SHPRH-146aa. Compared with paracancerous tissues, the expression of
SHPRH-146aa in tumour tissues was found to be significantly lower
(62). Other findings have
demonstrated that SHPRH-146aa overexpression can significantly
inhibit the growth of gliomas in xenograft mouse models; in
addition, it can protect the full-length SHPRH protein from
degradation by the ubiquitin proteasome system (54). In addition to the common tumour
suppressor genes mentioned above, recent studies have also reported
other tumour suppressor genes associated with gliomas. circ-SMARCA5
is downregulated in glioma cells, and circ-SMARCA5 overexpression
can increase SRSF3 expression and effectively control the excessive
proliferation of glioma cells (45). In addition, other researchers have
confirmed that circ-ITCH and hsa_circ_0001649 act as tumour
suppressors in gliomas, and they may prove beneficial for the
treatment of gliomas in the future (63,64).
9. Emergence of circRNAs in the clinical
treatment of glio-mas
Relevant research shows that circRNAs are widely
present in body fluids, such as plasma, cerebrospinal fluid and
breast milk, particularly in exosomes. The circular closed
structure of circRNAs protects them from degradation and confers
stability (65). Therefore, their
potential application for gene diagnosis and targeted therapy
appears to be a promising approach.
It was demonstrated that hsa_circ_0133159 and
circ_ZEB1 are significantly overexpressed in glioma tissues and
cell lines, and that they may be used as biomarkers in the
differential diagnosis and prognostic evaluation of gliomas
(66).
miR-24-3p is overexpressed in glioma cells and
promotes tumour cell proliferation, migration and invasion by
down-regulating bone morphogenetic protein 3 (67). Recent studies have revealed that
endothelial monocyte-activating polypeptide II (EMAP-II) can
inhibit angiogenesis and promote endothelial cell apoptosis, and
can directly promote tumour cell autophagy and apoptosis. The
binding of EMAP-II to temozolomide (TMZ) reduces the expression of
miR-24-3p, thereby enhancing the cytotoxic effect of TMZ on the
proliferation, migration and invasion of germline stem cells in
vitro and in vivo, further enhancing its therapeutic
efficacy. circRNAs play important roles in the regulation of this
process (68). Combined treatment
with miR-24-3p (-), EMAP-II and TMZ can reduce tumour malignancy
and increase the survival rate, whereas it has also been reported
that EMAP-II can promote tumour necrosis factor receptor 1
apoptosis, thereby exerting an antitumour effect (68). In addition, the permeability of the
blood-brain barrier is increased to further promote tumour cell
apoptosis.
Wesselhoeft et al invented a new method for
expressing proteins through engineered circRNA vectors. As circRNAs
are relatively more stable compared with normal linear RNA, protein
expression based on circRNAs can greatly increase production
efficiency. The results revealed that exogenous circRNAs can
produce high-quality and high-yield proteins, and indicated that
circRNAs are good and efficient RNA molecules that can be used for
protein expression in the future (69). Meganck et al constructed a
tissue-specific circRNA expression vector based on a recombinant
adeno-associated virus (AAV) vector. The results demonstrated that
the vector efficiently expressed and translated circRNAs in mouse
brain, eyes and heart (70). Based
on the research results mentioned above, in the near future,
protein-based expression systems will likely be used to design
circRNAs that target specific brain areas to treat glioma.
In addition, the molecular sponge effect of circRNAs
may also be used in the treatment of gliomas in the future. It was
recently reported that a circRNA with 5 miR-21-binding sites,
scRNA21, can significantly reduce the expression level of miR-21
and inhibit the proliferation of gastric cancer cell lines
(71). Although that study
addressed gastric cancer treatment, the concept of designing and
synthesizing circRNAs for specific miRNAs may hold promise in the
treatment of gliomas. miR-21 and several other miRNAs play
important roles in the development of gliomas. This also provides a
basis for gene therapy for gliomas, and scRNA21 may also be used to
interfere with these miRNAs in gliomas. AAV-based circRNA
expression vectors are a promising method and experimental basis
for this novel treatment concept (72).
TMZ is a first-line chemotherapy agent for glioma
treatment. According to tumour histopathology, immunohistochemistry
and molecular diagnosis, for patients with high-grade (WHO
classification) gliomas, particularly GBM, who undergo surgical
treatment, the routine use of TMZ after surgery is crucial for
preventing glioma recurrence and prolonging patient survival.
However, tumour resistance to chemotherapeutic drugs is an
important cause of treatment failure (73). Ding et al found that
exosomal circNFIX was upregulated in the serum of TMZ-resistant
patients and predicted a poor prognosis. In addition, TMZ-resistant
cell-derived circNFIX conferred TMZ resistance to recipient
sensitive cells by regulating cell migration, invasion and
apoptosis following TMZ exposure. Further analyses revealed that
following the knockdown of circNFIX, this enhanced TMZ sensitivity
in resistant glioma cells by upregulating miR-132. Therefore, in
terms of the circRNA molecular sponge effect, circNFIX can induce a
sponge effect of miR-132, thereby enhancing TMZ resistance in
glioma. (74,75).
Finally, the latest study by Yang et al in
2017 revealed that circ-FBXW7 has the ability to translate and
encode proteins. The 185-amino acid circ-FBXW7-185aa encoded by
circRNA-FBXW7 has a marked antitumour effect. At the animal and
clinical levels, after analysing the correlation between the
expression of circRNA-FBXW7 and overall survival, it was observed
that its expression level was positively associated with overall
survival (62). Therefore,
circRNAs have potential application prospects as biomarkers for
evaluating the prognosis of patients with glioma.
10. Final remarks
The role of RNAs in cellular activities is
multi-layered and multi-faceted. Despite being the most recently
discovered non-coding RNA molecules, circRNAs still follow the
basic rules of RNA. Current studies indicate that, in the human
genome, circRNAs are functional molecules in various tissues and
cells (10). They are
differentially expressed across different functional tissues and
cells, which indicates that circRNAs may regulate different
biological functions. circRNAs may play important roles in the
proliferation, apoptosis and invasion of glioma cells through
functions such as competitive inhibition, alternative splicing,
transcriptional regulation, protein coding, antiviral immune
responses and miRNA sponging; however, the specific mechanisms
remain elusive (76). The
continuous development of bioinformatics analysis technology may
further promote the exploration of circRNAs in the field of
glioma.
Although the popularity of circRNAs in the research
field is increasing, glioma is a highly heterogeneous disease that
has different pathological manifestations at different stages of
the disease and in different anatomic locations, in the same
patient or in different patients. Therefore, the association
between circRNAs and the clinical characteristics of gliomas is
complex and remains largely elusive, and the mechanisms underlying
glioma-specific circRNA expression patterns require further
investigation. Thus far, circRNA knockout remains challenging. As
the majority of circRNAs form from genes with coding ability,
knockout or downregulation of the corresponding circRNAs will
inevitably affect the host genes. In addition, widespread
low-abundance expression levels may also hinder protein translation
of circRNAs in gliomas and other human diseases.
Finally, high-throughput sequencing technology, also
referred to as 'next-generation' sequencing technology, removes
technical obstacles to reveal a large number of unknown circRNAs.
However, research on the function and mechanism of circRNAs is in
its early stages (77,78). In terms of glioma research at the
cellular and molecular biology level, the concept of studying a
certain gene, protein or pathway alone is outdated. In the future,
the key stages of glioma development must be further investigated
to identify specific circRNAs, in order to establish a complete
gene-protein interaction network and to determine their function
and underlying mechanism of action by comprehensively using both
in vitro and in vivo experiments and clinical trials.
In the foreseeable future, circRNAs are expected to become a focus
in the field of non-coding RNA research. Related studies will be
required to confirm that circRNAs play key roles in glioma and
other diseases, thereby providing new perspectives for the early
diagnosis, pathological grading, targeted therapy and prognosis of
this disease.
Finally, relevant websites on circRNA biological
information analysis and databases for researchers have been
organized (Tables I and II).
 | Table ISoftware for circRNA
identification. |
Table I
Software for circRNA
identification.
| Table IIDatabases for circRNA deposition. |
Table II
Databases for circRNA deposition.
Funding
The present study was financially supported by the
government-funded Provincial Clinical Medicine Talent Programs in
2017; the High-level Talents Scientific Research Launched Project
of Hebei University (grant no. 521000981301); the Science and
Technology Capacity Improvement Projects of Hebei University of
Chinese Medicine in 2019 (grant no. KTZ2019019); the Outstanding
Student Scientific Research Ability Improvement Projects of Hebei
University of Chinese Medicine in 2019 (grant no. YXZ2019002); the
Graduate Innovative Ability Training Projects of Hebei University
of Chinese Medicine in 2020 (grant no. XCXZZBS2020002); and the
Graduate Innovative Ability Training Projects of Hebei University
in 2020 (grant no. hbu2020bs003).
Availability of data and materials
The datasets used and analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and XL were major contributors to the writing and
revision of the manuscript. XG, LS, QL and FL collected the related
references and participated in the discussions. CF and HWwere
involved in the conception and design of the study. All authors
have read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
Acknowledgments
The authors would like to give special thanks to
Ruijuan Ren, the curator of Hebei University library, for her
guidance and assistance in the literature retrieval process of this
manuscript.
References
|
1
|
Pace A, Dirven L, Koekkoek JAF, Golla H,
Fleming J, Rudà R, Marosi C, Rhun EL, Grant R, Oliver K, et al:
European Association of Neuro-Oncology palliative care task force:
European Association for Neuro-Oncology (EANO) guidelines for
palliative care in adults with glioma. Lancet Oncol. 18:e330–e340.
2017. View Article : Google Scholar
|
|
2
|
Weller M, van den Bent M, Tonn JC, Stupp
R, Preusser M, Cohen-Jonathan-Moyal E, Henriksson R, Rhun EL,
Balana C, Chinot O, et al: European Association for Neuro-Oncology
(EANO) Task Force on Gliomas: European Association for
Neuro-Oncology (EANO) guideline on the diagnosis and treatment of
adult astrocytic and oligodendroglial gliomas. Lancet Oncol.
18:e315–e329. 2017. View Article : Google Scholar
|
|
3
|
Bush NA, Chang SM and Berger MS: Current
and future strategies for treatment of glioma. Neurosurg Rev.
40:1–14. 2017. View Article : Google Scholar
|
|
4
|
Calore F, Lovat F and Garofalo M:
Non-coding RNAs and cancer. Int J Mol Sci. 14:17085–17110. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shukla GC and Gupta S: Hallmarks of
cancer- focus on RNA metabolism and regulatory noncoding RNAs.
Cancer Lett. 420:208–209. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bach DH, Lee SK and Sood AK: Circular RNAs
in Cancer. Mol Ther Nucleic Acids. 16:118–129. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, Lu T, Wang Q, Liu J and Jiao W:
Circular RNAs: Crucial regulators in the human body (Review). Oncol
Rep. 40:3119–3135. 2018.PubMed/NCBI
|
|
8
|
Patop IL, Wüst S and Kadener S: Past,
present, and future of circRNAs. EMBO J. 38:e1008362019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Arnaiz E, Sole C, Manterola L,
Iparraguirre L, Otaegui D and Lawrie CH: circRNAs and cancer:
Biomarkers and master regulators. Semin Cancer Biol. 58:90–99.
2019. View Article : Google Scholar
|
|
10
|
Patop IL and Kadener S: circRNAs in
Cancer. Curr Opin Genet Dev. 48:121–127. 2018. View Article : Google Scholar :
|
|
11
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:3852–3856. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hsu MT and Coca-Prados M: Electron
microscopic evidence for the circular form of RNA in the cytoplasm
of eukaryotic cells. Nature. 280:339–340. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arnberg AC, Van Ommen GJ, Grivell LA, Van
Bruggen EF and Borst P: Some yeast mitochondrial RNAs are circular.
Cell. 19:313–319. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cocquerelle C, Mascrez B, Hétuin D and
Bailleul B: Mis-splicing yields circular RNA molecules. FASEB J.
7:155–160. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo JU, Agarwal V, Guo H and Bartel DP:
Expanded identification and characterization of mammalian circular
RNAs. Genome Biol. 15:4092014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hansen T: Biogenesis and function of
circRNAs. FEBS Open Bio. 9:392019.
|
|
22
|
Shen Y, Guo X and Wang W: Identification
and characterization of circular RNAs in zebrafish. FEBS Lett.
591:213–220. 2017. View Article : Google Scholar
|
|
23
|
Huang G, Li S, Yang N, Zou Y, Zheng D and
Xiao T: Recent progress in circular RNAs in human cancers. Cancer
Lett. 404:8–18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pamudurti NR, Bartok O, Jens M,
Ashwal-Fluss R, Stottmeister C, Ruhe L, Hanan M, Wyler E,
Perez-Hernandez D, Ramberger E, et al: Translation of circRNAs. Mol
Cell. 66:9–21.e7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu XQ, Gao YB, Zhao LZ, Cai YC, Wang HY,
Miao M, Gu LF and Zhang HX: Biogenesis, research methods, and
functions of circular RNAs. Yi Chuan. 41:469–485. 2019.In Chinese.
PubMed/NCBI
|
|
26
|
Harper KL, Mcdonnell E and Whitehouse A:
circRNAs: From anonymity to novel regulators of gene expression in
cancer (Review). Int J Oncol. 55:1183–1193. 2019.PubMed/NCBI
|
|
27
|
Rybak-Wolf A, Stottmeister C, Glažar P,
Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss
R, et al: Circular RNAs in the Mammalian Brain Are Highly Abundant,
Conserved, and Dynamically Expressed. Mol Cell. 58:870–885. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen BJ, Huang S and Janitz M: Changes in
circular RNA expression patterns during human foetal brain
development. Genomics. 111:753–758. 2019. View Article : Google Scholar
|
|
29
|
Westholm JO, Miura P, Olson S, Shenker S,
Joseph B, Sanfilippo P, Celniker SE, Graveley BR and Lai EC:
Genome-wide analysis of drosophila circular RNAs reveals their
structural and sequence properties and age-dependent neural
accumulation. Cell Rep. 9:1966–1980. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
You X, Vlatkovic I, Babic A, Will T,
Epstein I, Tushev G, Akbalik G, Wang M, Glock C, Quedenau C, et al:
Neural circular RNAs are derived from synaptic genes and regulated
by development and plasticity. Nat Neurosci. 18:603–610. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Piwecka M, Glažar P, Hernandez-Miranda LR,
Memczak S, Wolf SA, Rybak-Wolf A, Filipchyk A, Klironomos F, Cerda
Jara CA, Fenske P, et al: Loss of a mammalian circular RNA locus
causes miRNA deregulation and affects brain function. Science.
357:3572017. View Article : Google Scholar
|
|
32
|
Wang Q, Qu L, Chen X, Zhao YH and Luo Q:
Progress in Understanding the Relationship Between Circular RNAs
and Neurological Disorders. J Mol Neurosci. 65:546–556. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mehta SL, Dempsey RJ and Vemuganti R: Role
of circular RNAs in brain development and CNS diseases. Prog
Neurobiol. 186:1017462020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Du WW, Yang W, Chen Y, Wu ZK, Foster FS,
Yang Z, Li X and Yang BB: Foxo3 circular RNA promotes cardiac
senescence by modulating multiple factors associated with stress
and senescence responses. Eur Heart J. 38:1402–1412. 2017.
|
|
36
|
Mahmoudi E, Fitzsimmons C, Geaghan MP,
Shannon Weickert C, Atkins JR, Wang X and Cairns MJ: Circular RNA
biogenesis is decreased in postmortem cortical gray matter in
schizophrenia and may alter the bioavailability of associated
miRNA. Neuropsychopharmacology. 44:1043–1054. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rynkeviciene R, Simiene J, Strainiene E,
Stankevicius V, Usinskiene J, Miseikyte Kaubriene E, Meskinyte I,
Cicenas J and Suziedelis K: Non-Coding RNAs in Glioma. Cancers
(Basel). 11:172018. View Article : Google Scholar
|
|
38
|
Liu J, Zhao K, Huang N and Zhang N:
Circular RNAs and human glioma. Cancer Biol Med. 16:11–23. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 World Health Organization
Classification of Tumors of the Central Nervous System: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Song X, Zhang N, Han P, Moon BS, Lai RK,
Wang K and Lu W: Circular RNA profile in gliomas revealed by
identification tool UROBORUS. Nucleic Acids Res. 44:e872016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sun J, Li B, Shu C, Ma Q and Wang J:
Functions and clinical significance of circular RNAs in glioma. Mol
Cancer. 19:342020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhu J, Ye J, Zhang L, Xia L, Hu H, Jiang
H, Wan Z, Sheng F, Ma Y, Li W, et al: Differential Expression of
Circular RNAs in Glioblastoma Multiforme and Its Correlation with
Prognosis. Transl Oncol. 10:271–279. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang HX, Huang QL, Shen JY, Xu T, Hong F,
Gong ZY, Li F, Yan Y and Chen JX: Expression profile of circular
RNAs in IDH-wild type glioblastoma tissues. Clin Neurol Neurosurg.
171:168–173. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu H, Zhang Y, Qi L, Ding L, Jiang H and
Yu H: NFIX circular RNA promotes glioma progression by regulating
miR-34a-5p via notch signaling pathway. Front Mol Neurosci.
11:2252018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Barbagallo D, Caponnetto A, Cirnigliaro M,
Brex D, Barbagallo C, D'Angeli F, Morrone A, Caltabiano R,
Barbagallo GM, Ragusa M, et al: circSMARCA5 inhibits migration of
glioblastoma multiforme cells by regulating a molecular axis
involving splicing factors SRSF1/SRSF3/PTB. Int J Mol Sci.
19:4802018. View Article : Google Scholar :
|
|
46
|
Bian A, Wang Y, Liu J, Wang X, Liu D,
Jiang J, Ding L and Hui X: Circular RNA complement factor H (CFH)
promotes glioma progression by sponging mir-149 and regulating
AKT1. Med Sci Monit. 24:5704–5712. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xie G: Circular RNA hsa-circ-0012129
promotes cell proliferation and invasion in 30 cases of human
glioma and human glioma cell lines U373, A172, and SHG44, by
targeting MicroRNA-661 (miR-661). Med Sci Monit. 24:2497–2507.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang R, Zhang S, Chen X, Li N, Li J, Jia
R, Pan Y and Liang H: CircNT5E acts as a sponge of miR-422a to
promote glioblastoma tumorigenesis. Cancer Res. 78:4812–4825. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kumar V, Soni UK, Maurya VK, Singh K and
Jha RK: Integrin beta8 (ITGB8) activates VAV-RAC1 signaling via FAK
in the acquisition of endometrial epithelial cell receptivity for
blastocyst implantation. Sci Rep. 7:18852017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu Z, Hu G, Zhao Y, Xiao Z, Yan M and Ren
M: Silence of cZNF292 suppresses the growth, migration, and
invasion of human esophageal cancer Eca-109 cells via upregulating
miR-206. J Cell Biochem. 121:2354–2362. 2020. View Article : Google Scholar
|
|
51
|
Yang P, Qiu Z, Jiang Y, Dong L, Yang W, Gu
C, Li G and Zhu Y: Silencing of cZNF292 circular RNA suppresses
human glioma tube formation via the Wnt/beta-catenin signaling
pathway. Oncotarget. 7:63449–63455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bi W, Huang J, Nie C, Liu B, He G, Han J,
Pang R, Ding Z, Xu J and Zhang J: circRNA circRNA_102171 promotes
papillary thyroid cancer progression through modulating
CTNNBIP1-dependent activation of β-catenin pathway. J Exp Clin
Cancer Res. 37:2752018. View Article : Google Scholar
|
|
53
|
Ouyang YB and Giffard RG: MicroRNAs affect
BCL-2 family proteins in the setting of cerebral ischemia.
Neurochem Int. 77:2–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang M, Huang N, Yang X, Luo J, Yan S,
Xiao F, Chen W, Gao X, Zhao K, Zhou H, et al: A novel protein
encoded by the circular form of the SHPRH gene suppresses glioma
tumori-genesis. Oncogene. 37:1805–1814. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chaffer CL, San Juan BP, Lim E and
Weinberg RA: EMT, cell plasticity and metastasis. Cancer Metastasis
Rev. 35:645–654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zheng J, Liu X, Xue Y, Gong W, Ma J, Xi Z,
Que Z and Liu Y: TTBK2 circular RNA promotes glioma malignancy by
regulating miR-217/HNF1β/Derlin-1 pathway. J Hematol Oncol.
10:522017. View Article : Google Scholar
|
|
57
|
Li GF, Li L, Yao ZQ and Zhuang SJ:
Hsa_circ_ 0007534/miR-761/ZIC5 regulatory loop modulates the
proliferation and migration of glioma cells. Biochem Biophys Res
Commun. 499:765–771. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen Z and Duan X:
hsa_circ_0000177-miR-638-FZD7-Wnt Signaling Cascade Contributes to
the Malignant Behaviors in Glioma. DNA Cell Biol. 37:791–797. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li G, Yang H, Han K, Zhu D, Lun P and Zhao
Y: A novel circular RNA, hsa_circ_0046701, promotes carcinogenesis
by increasing the expression of miR-142-3p target ITGB8 in glioma.
Biochem Biophys Res Commun. 498:254–261. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jin P, Huang Y, Zhu P, Zou Y, Shao T and
Wang O: circRNA circHIPK3 serves as a prognostic marker to promote
glioma progression by regulating miR-654/IGF2BP3 signaling. Biochem
Biophys Res Commun. 503:1570–1574. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
He Q, Zhao L, Liu Y, Liu X, Zheng J, Yu H,
Cai H, Ma J, Liu L, Wang P, et al: circ-SHKBP1 Regulates the
Angiogenesis of U87 Glioma-Exposed Endothelial Cells through
miR-544a/FOXP1 and miR-379/FOXP2 Pathways. Mol Ther Nucleic Acids.
10:331–348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao
F, Huang N, Yang X, Zhao K, Zhou H, et al: Novel role of FBXW7
circular RNA in repressing glioma tumorigenesis. J Natl Cancer
Inst. 110:304–315. 2018. View Article : Google Scholar :
|
|
63
|
Li F, Ma K, Sun M and Shi S:
Identification of the tumor-suppressive function of circular RNA
ITCH in glioma cells through sponging miR-214 and promoting linear
ITCH expression. Am J Transl Res. 10:1373–1386. 2018.PubMed/NCBI
|
|
64
|
Wang Y, Sui X, Zhao H, Cong L, Li Y, Xin
T, Guo M and Hao W: Decreased circular RNA hsa_circ_0001649
predicts unfavorable prognosis in glioma and exerts oncogenic
properties in vitro and in vivo. Gene. 676:117–122. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Bai H, Lei K, Huang F, Jiang Z and Zhou X:
ExocircRNAs: A new paradigm for anticancer therapy. Mol Cancer.
18:562019. View Article : Google Scholar
|
|
66
|
Edwards LA, Li A, Berel D, Madany M, Kim
N-H, Liu M, Hymowitz M, Uy B, Jung R, Xu M, et al: ZEB1 regulates
glioma stemness through LIF repression. Sci Rep. 7:692017.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen J, Liu L, Liu Y, Liu X, Qu C, Meng F,
Ma J, Lin Y and Xue Y: Low-Dose Endothelial-Monocyte-Activating
Polypeptide-II Induced Autophagy by Down-Regulating miR-20a in U-87
and U-251 Glioma Cells. Front Cell Neurosci. 10:1282016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang J, Liu L, Xue Y, Ma Y, Liu X, Li Z,
Li Z and Liu Y: Endothelial Monocyte-Activating Polypeptide-II
Induces BNIP3-Mediated Mitophagy to Enhance Temozolomide
Cytotoxicity of Glioma Stem Cells via Down-Regulating miR-24-3p.
Front Mol Neurosci. 11:922018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wesselhoeft RA, Kowalski PS and Anderson
DG: Engineering circular RNA for potent and stable translation in
eukaryotic cells. Nat Commun. 9:26292018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Meganck RM, Borchardt EK, Castellanos
Rivera RM, Scalabrino ML, Wilusz JE, Marzluff WF and Asokan A:
Tissue-dependent expression and translation of circular RNAs with
recombinant AAV vectors in vivo. Mol Ther Nucleic Acids. 13:89–98.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liu X, Abraham JM, Cheng Y, Wang Z, Wang
Z, Zhang G, Ashktorab H, Smoot DT, Cole RN, Boronina TN, et al:
Synthetic circular RNA functions as a miR-21 sponge to suppress
gastric carcinoma cell proliferation. Mol Ther Nucleic Acids.
13:312–321. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Møller HG, Rasmussen AP, Andersen HH,
Johnsen KB, Henriksen M and Duroux M: A systematic review of
microRNA in glioblastoma multiforme: Micro-modulators in the
mesenchymal mode of migration and invasion. Mol Neurobiol.
47:131–144. 2013. View Article : Google Scholar :
|
|
73
|
Annovazzi L, Caldera V, Mellai M, Riganti
C, Battaglia L, Chirio D, Melcarne A and Schiffer D: The DNA
damage/repair cascade in glioblastoma cell lines after
chemotherapeutic agent treatment. Int J Oncol. 46:2299–2308. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ding C, Yi X, Wu X, Bu X, Wang D, Wu Z,
Zhang G, Gu J and Kang D: Exosome-mediated transfer of circRNA
CircNFIX enhances temozolomide resistance in glioma. Cancer Lett.
479:1–12. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Geng X, Jia Y, Zhang Y, Shi L, Li Q, Zang
A and Wang H: Circular RNA: Biogenesis, degradation, functions and
potential roles in mediating resistance to anticarcinogens.
Epigenomics. 12:267–283. 2020. View Article : Google Scholar
|
|
76
|
Hao Z, Hu S, Liu Z, Song W, Zhao Y and Li
M: Circular RNAs: Functions and Prospects in Glioma. J Mol
Neurosci. 67:72–81. 2019. View Article : Google Scholar
|
|
77
|
Chaabane M, Rouchka EC and Park JW:
Circular RNA Detection from High-throughput Sequencing. In:
Proceedings of the International Conference on Research in Adaptive
and Convergent Systems; September 20-23, 2017; Krakow, Poland. pp.
19–24. 2017
|
|
78
|
Fischer JW and Leung AKL: circRNAs: A
regulator of cellular stress. Crit Rev Biochem Mol Biol.
52:220–233. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang K, Singh D, Zeng Z, Coleman SJ, Huang
Y, Savich GL, He X, Mieczkowski P, Grimm SA, Perou CM, et al:
MapSplice: Accurate mapping of RNA-seq reads for splice junction
discovery. Nucleic Acids Res. 38:e1782010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Hoffmann S, Otto C, Doose G, Tanzer A,
Langenberger D, Christ S, Kunz M, Holdt LM, Teupser D, Hackermüller
J, et al: A multi-split mapping algorithm for circular RNA,
splicing, trans-splicing and fusion detection. Genome Biol.
15:R342014. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Szabo L, Morey R, Palpant NJ, Wang PL,
Afari N, Jiang C, Parast MM, Murry CE, Laurent LC and Salzman J:
Statistically based splicing detection reveals neural enrichment
and tissue-specific induction of circular RNA during human fetal
development. Genome Biol. 16:1262015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Cheng J, Metge F and Dieterich C: Specific
identification and quantification of circular RNAs from sequencing
data. Bioinformatics. 32:1094–1096. 2016. View Article : Google Scholar
|
|
83
|
You X and Conrad TO: Acfs: Accurate
circRNA identification and quantification from RNA-Seq data. Sci
Rep. 6:388202016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ye CY, Zhang X, Chu Q, Liu C, Yu Y, Jiang
W, Zhu QH, Fan L and Guo L: Full-length sequence assembly reveals
circular RNAs with diverse non-GT/AG splicing signals in rice. RNA
Biol. 14:1055–1063. 2017. View Article : Google Scholar :
|
|
85
|
Metge F, Czaja-Hasse LF, Reinhardt R and
Dieterich C: FUCHS-towards full circular RNA characterization using
RNAseq. PeerJ. 5:e29342017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Zheng Y, Ji P, Chen S, Hou L and Zhao F:
Reconstruction of full-length circular RNAs enables isoform-level
quantification. Genome Med. 11:22019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhang XO, Dong R, Zhang Y, Zhang JL, Luo
Z, Zhang J, Chen LL and Yang L: Diverse alternative back-splicing
and alternative splicing landscape of circular RNAs. Genome Res.
26:1277–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Gao Y, Zhang J and Zhao F: Circular RNA
identification based on multiple seed matching. Brief Bioinform.
19:803–810. 2018. View Article : Google Scholar
|
|
89
|
Feng J, Xiang Y, Xia S, Liu H, Wang J,
Ozguc FM, Lei L, Kong R, Diao L, He C, et al: CircView: A
visualization and exploration tool for circular RNAs. Brief
Bioinform. 19:1310–1316. 2018.
|
|
90
|
Gao Y, Wang H, Zhang H, Wang Y, Chen J and
Gu L: PRAPI: Post-transcriptional regulation analysis pipeline for
Iso-Seq. Bioinformatics. 34:1580–1582. 2018. View Article : Google Scholar
|
|
91
|
Feng J, Chen K, Dong X, Xu X, Jin Y, Zhang
X, Chen W, Han Y, Shao L, Gao Y, et al: Genome-wide identification
of cancer-specific alternative splicing in circRNA. Mol Cancer.
18:352019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ji P, Wu W, Chen S, Zheng Y, Zhou L, Zhang
J, Cheng H, Yan J, Zhang S, Yang P, et al: Expanded expression
landscape and prioritization of circular RNAs in mammals. Cell Rep.
26:3444–3460.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Glažar P, Papavasileiou P and Rajewsky N:
circBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar
|
|
94
|
Xia S, Feng J, Lei L, Hu J, Xia L, Wang J,
Xiang Y, Liu L, Zhong S, Han L, et al: Comprehensive
characterization of tissue-specific circular RNAs in the human and
mouse genomes. Brief Bioinform. 18:984–992. 2017.
|
|
95
|
Dudekula DB, Panda AC, Grammatikakis I, De
S, Abdelmohsen K and Gorospe M: CircInteractome: A web tool for
exploring circular RNAs and their interacting proteins and
microRNAs. RNA Biol. 13:34–42. 2016. View Article : Google Scholar :
|
|
96
|
Liu YC, Li JR, Sun CH, Andrews E, Chao RF,
Lin FM, Weng SL, Hsu SD, Huang CC, Cheng C, et al: CircNet: A
database of circular RNAs derived from transcriptome sequencing
data. Nucleic Acids Res. 44D:D209–D215. 2016. View Article : Google Scholar
|
|
97
|
Chen X, Han P, Zhou T, Guo X, Song X and
Li Y: circRNADb: A comprehensive database for human circular RNAs
with protein-coding annotations. Sci Rep. 6:349852016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Xia S, Feng J, Chen K, Ma Y, Gong J, Cai
F, Jin Y, Gao Y, Xia L, Chang H, et al: CSCD: A database for
cancer-specific circular RNAs. Nucleic Acids Res. 46D:D925–D929.
2018. View Article : Google Scholar
|
|
99
|
Fan C, Lei X, Fang Z, Jiang Q and Wu FX:
circR2Disease: A manually curated database for experimentally
supported circular RNAs associated with various diseases. Database
(Oxford). 2018. pp. bay0442018, View Article : Google Scholar
|
|
100
|
Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao
L, Zhang Y, Wu YM, Dhanasekaran SM, Engelke CG, Cao X, et al: The
landscape of circular RNA in cancer. Cell. 176:869–881.e13. 2019.
View Article : Google Scholar : PubMed/NCBI
|