1. The role of taurine in inflammation
Taurine (2-aminoethanesulfonic acid) is a
non-essential amino acid that is abundant in all mammalian tissues.
Taurine is essential for cell growth of renal, neural, and cardiac
cells, preventing death procedures (1,2).
Taurine plays a significant role in homeostasis because it is
involved in the regulation of the following processes: cell volume
regulation, osmo-regulation, protein phosphorylation, membrane
stability, bile acid conjugation, neuromodulation, maintenance of
calcium concentration, and detoxification of xenobiotics (3). The anti-oxidant and anti-inflammatory
properties of taurine constitute the main mechanisms that account
for its cytopro-tection (3,4).
Taurine accumulates in phagocytes (both neutrophils
and macrophages) as well as in inflammatory lesions, illustrating
its potential significance in innate immunity (5). It has been reported that taurine
concentration can reach 50-70% in neutrophilic granulocytes,
lymphocytes and monocytes (5-7). The
contribution of taurine to the immune surveillance relies on the
anti-oxidant properties of taurine (8) and its membrane-stabilizing capacity
(9). For example, experimental
evidence has highlighted that taurine mainly exerts its
anti-oxidant activity through inhibition of sodium arsenite-induced
apoptosis in neutrophils (10).
Taurine has a protective role, sustaining the phagocytic ability of
neutrophils independently the stimulus including age (11) or hyperlipidemia (12). Besides, some reports have
highlighted that taurine exerts its beneficial effect on
leukocytes, via alleviating the oxidative stress (13,14).
The pleiotropic nature of taurine is not tightly associated with
the anti-oxidant properties, but it is also related to its
membrane-stabilizing capacity in lymphocytes (15). In this direction, the anti-oxidant
nature of taurine has been shown to account for preserving the
viability of human lymphocyte-derived cultured lymphoblastoid
cells, protecting against oxidant-induced damage caused by ferrous
sulfate and ascorbate (15).
Taurine is regarded as a promising agent against
numerous types of inflammatory injury (inflammatory bowel disease,
pancreatitis, and gastric mucosal injury), due to its
immunoregulatory importance(16,17).
Taurine is effective against various acute inflammation related
diseases, including spinal cord injury (18), hepatic ischemia-reperfusion
(19), lung injury (20,21),
ischemic stroke (22), and
trinitrobenzene sulfonic acid (TNBS)-induced colitis in the rat
(23), lipopoly-saccharide
(LPS)-induced acute lung injury in sheep (20) and dextran sulfate sodium
(DSS)-induced colitis (24,25).
In these conditions, the anti-inflammatory action of taurine is
usually attributed to its antioxidant effect, which is manifested
by inhibiting lipid peroxidation (LPO) (16). The anti-inflammatory effect of
taurine has also been attributed to the reduced secretion of
interleukins (ILs) (such as IL-8), as shown by experiments in
Caco-2 cells, without any participation of polymorphonuclear
leukocytes (24). Accordingly,
there is a growing body of preclinical data that demonstrates the
anti-inflammatory effects of taurine in both neural and systemic
inflammation including cardiovascular disease (26), traumatic brain injury (27), liver/gallbladder disease (28), lung injury (29), diabetes (30), cataract (31). As a result, taurine fulfills the
necessary criteria to participate in the regulatory network of an
immune response.
Moreover, the immune-regulatory effect of taurine
has been validated through studies examining the consequences of a
taurine deficiency. When taurine elimination arose in cats, the
immune landscape was reorganized. In particular, taurine-deficient
cats presented significant leukopenia, decreased respiratory burst
in neutrophils, and depletion of cells from B cell areas in lymph
nodes and splenic follicle centers (32). Apart from leukopenia, taurine
deficiency proved to cause functional defects of the neutrophils
and decreased phagocytosis of microorganisms such as Staphylococcus
epidermis (5,33). Conversely, taurine was reported to
mediate its ameliorative effect on age-related decline in the
proliferative ability of lymphocytes through increasing calcium
levels. In particular, the effect of taurine was more potent on
T-cells, that were more susceptible to age-related decline in
proliferation than B lymphocytes (32). The effect of taurine on lymphocyte
function was substantiated through its chaperoning role concerning
MHC class II antigen expression (33). The aforementioned data were
evaluated given that T-cell proliferation is mediated independently
of the age-related alteration in taurine transport (34).
Besides, it is important to be noted, taurine
biosynthesis has been outlined to be divided into the oxidation of
cysteine to cysteine sulfinic acid followed by the decarboxylation
to hypotaurine, with the subsequent oxidation to taurine (35). In this sense, the significance of
taurine has been proven in the immune system through the
elimination of cysteine sulfinic acid decarboxylase (CSAD), which
is crucial for the conversion of cysteine sulfinic acid to
hypotaurine (35). In particular,
it has been observed that taurine concentration was significantly
higher in the splenocytes and macrophages from CSAD knock-out (KO)
mice compared to those encountered in the liver and plasma from
CSAD KO mice (7,36), implying its significance in the
immune system.
Beneficial effect of taurine in various
cancers (in vitro and in vivo)
Cancer is a direct consequence of gene mutations
that arise in a multistep process, enabling cancer cells to possess
a sustained replicative potential (16). Vogelstein declared that 'The
revolution in cancer research can be summed up in a single
sentence: Cancer is, in essence, a genetic disease' (37). Tumorigenesis comprises a series of
events, in which excessive reactive oxygen species (ROS) formation
is the determinant force for cancer development (38). In line with this, many
anti-oxidants including methionine, cysteine, and taurine have been
identified to display strong potential of minimizing oxidative
injury in cancer (32,39). On the contrary, cancer cells have
been reported to constitutively express low ROS levels and high
antioxidant responses during tumor progression (40). This specific vulnerability of
various tumor cells is termed 'non-oncogene dependency' (38). In keeping with this observation,
small molecular weight pro-oxidant drugs have been shown to cause
an oxidative burst in cancer cells harboring low oxidative status,
with the ultimate aim of eradicating them (41).
Since taurine exerts a strong anti-inflammatory
action, the functional significance of taurine has been presented
in orchestrating the landscape of tumor cells. Meeting this
objective, many research groups have illustrated the anti-cancer
impact of taurine, providing insights into its molecular
mechanisms. Taurine functions as a redox-directed agent to
specifically target tumor cells, raising the possibilities to
achieve drug selectivity without off-target toxicity. Several cases
have proved that taurine displays strong growth-inhibitory effect
on multiple cancer types including colon cancer (42,43),
lung cancer (44), hepatocarcinoma
(30), pancreatic cancer (45), glioma (46), melanoma (47) breast cancer (48-51),
nasopharyngeal carcinoma (NPC) (52), prostate cancer (53,54)
and ovarian cancer (55,56).
Regarding the molecular mechanisms underlying the
anti-cancer effect of taurine, it has been proposed that the effect
of taurine on tumor cells can be either cytostatic (i.e., cell
growth suppression) or cytotoxic (i.e., direct toxic effect). The
anti-cancer effect of taurine is mainly mediated through multiple
molecular mechanisms. Firstly, taurine exerts a growth-inhibitory
effect through its antioxidant nature (57). In most cases, the main molecular
mechanism underlying the anti-cancer effect of taurine relies on
modulating multiple signaling cascades (42,50,51,58-60),
through its anti-oxidant capacity (61-65).
For example, taurine has been reported to protect cells from
oxidant-induced injury by neutralizing insults derived from strong
oxidant and cytotoxic agents (66). As a further example, taurine has
been proposed as an effective antioxidant, preventing the
accumulation of ROS in tumor cells, thereby compromising cancer
progression (67). Secondly,
taurine ameliorates the efficacy of chemotherapeutic drugs,
minimizing their toxicity (68,69).
It has been pointed out that taurine supplementation overcomes
chemotherapy-induced complications, probably owing to its
antioxidant capacity (70-75). In particular, taurine shows strong
potential to attenuate toxic side effects of classic
chemotherapeutic drugs [doxorubicin (DOX), 5-fluorouracil (5-FU),
cisplatin, tamoxifen (TAM)], thereby enhancing their therapeutic
efficacy (61,74,76-78).
In this sense, taurine is crucial to expand the therapeutic window
of selected anti-tumor drugs, thereby optimizing the therapeutic
efficacy of drugs. Thirdly, taurine plays a significant role in the
immune rejection of cancer cells by enhancing immune surveillance
(31). Fourth, taurine imparts its
preventive action on cancer cells through the induction of
apoptosis (42). In support,
studies have shown that taurine triggers apoptosis in colon cancer
(42), breast cancer (50), and hepatocarcinoma (60). For example, the apoptotic effect of
taurine is accomplished by up-regulating the expression of the p53
transcription factor, while down-regulating the expression of
anti-apoptotic proteins such as B-cell lymphoma 2 (BCL-2) (42). In another case, taurine has been
proved to display its anti-neoplastic activity through the
induction of apoptosis in NPC (52). The mechanism underlying the
apoptotic effect of taurine is based on stimulating endoplasmic
reticulum stress and inactivating the protein kinase B (Akt)
signaling pathway (52). Besides,
tumors are characterized by a permissive microenvironment that
favors the induction of neo-angiogenesis for maintaining the supply
of oxygen and nutrients (16). In
this context, the anti-cancer activity of taurine has been
illustrated to be elicited through the inhibition of tumor
neovascularization and the induction of cytotoxicity on endothelial
cells. For example, taurine has been proved to downregulate matrix
metalloproteinase 2 (MMP-2), and to upregulate of
N-acetylgalactosaminytransferase, thereby preventing the increased
invasiveness of cancer cells from primary site through bloodstream
to other sites, in response to ionizing radiation (79). According to those viewpoints, a
comprehensive in-depth analysis regarding the molecular mechansims
of taurine underlying its therapeutic efficacy against distinct
cancer types was outlined.
In breast cancer, epidemiological studies have
suggested that an anti-oxidant enriched diet may be crucial to
reducing the emergence of breast cancer (80). In this context, the group of
Garmire used blood-based-metabolomics in conjunction with
RNA-Seq-based on The Cancer Genome Atlas (TCGA) breast cancer data
and highlighted that the taurine metabolic pathway is an important
regulatory pathway among eight others, enabling the diagnosis of
breast cancer occurrence in a personalized manner (81). Researchers have also used
high-resolution magic angle spinning magnetic resonance
spectroscopy (HR-MAS MRS) coupled with the relative principal
component analysis, in biological samples of breast cancer
patients, proving that small values for taurine were detected in
breast cancer patients with metastasis compared to healthy patients
(82). Similarly, four groups of
female patients were recruited and were divided as follows: i) 50
diagnosed patients with breast cancer subjected to surgery; ii) 10
female patients with benign breast cancer signs; iii) 5 females
equipped with high predisposition to breast cancer, due to their
family history; and iv) 20 healthy women who were used as control
samples to evaluate the diagnostic importance of taurine in
Egyptian patients with breast cancer (83). Following the evaluation of female
patients with breast cancer in various stages, taurine levels
appeared to be reduced in the serum of patients with a high risk of
breast cancer, providing a clue for the predisposition of women to
breast cancer or the early diagnosis of females with early
malignant lesions due to taurine detection (83).
In particular, the prognostic significance of
taurine was confirmed in the serum of patients with breast cancer,
because serum taurine levels were reduced in the breast cancer
group and were tightly linked to tumor angiogenesis, as evidenced
by reduced expression levels of angiogenesis markers [vascular
endothelial growth factor (VEGF), CD31] and apoptotic markers
[tumor necrosis factor-α (TNF-α), caspase-3] (83). Interestingly, females with positive
family history and women with benign breast lesions presented
taurine levels ranging from 40 to 57 µmol/l and from 18 to
31 µmol/l, respectively. In contrast, healthy women
presented taurine range from 46 to 70 µmol/l. It was
highlighted that minimal taurine value was found in women with high
susceptibility to breast cancer, proposing that minimal taurine
value of high-risk group did not exceed the lower limit recorded in
control healthy group (83).
Apart from the diagnostic and prognostic importance
of taurine, a wide range of tumor cell lines and mouse models
harboring mammary carcinogenesis have been employed to examine the
cytotoxic effect of taurine on breast cancer. Initial experiments
proved the beneficial impact of taurine on nude mice bearing breast
cancer xenografts (50). The
underlying molecular mechanism of taurine was based on inducing the
mitochondrial cell death pathway, as shown by increased expression
levels of p53-upregulated modulator of apoptosis (PUMA),
irrespective of the p53 genetic profile (50). In 2,4-dimethoxybenzaldehyde
(DMBA)-induced mammary carcinogenesis, the therapeutic impact of
taurine emerged by inhibiting the migration of breast cancer cells
through its strong antioxidant efficacy (59). In particular, the anti-oxidant
effect of taurine relied on its capacity to hinder mitochondrial
LPO and to normalize citric acid cycle enzyme expression, thus
augmenting electron transport chain complexes and delaying electron
cleavage responsible for the accumulation of ROS (59). In this context, taurine was proved
to exert a strong anti-neoplastic effect on rats harboring mammary
carcinogenesis, through its interference with energy metabolism of
rats not only by reducing breast cancer incidence, but also
forestalling breast cancer progression (84). The metabolic pattern of
taurine-treated tumor-bearing rats was distinguished from that of
tumor-bearing rats without taurine treatment, as shown by
experiments in the model of DMBA-induced mammary carcinogenesis
(84). In particular,
taurine-supplemented tumor-bearing rats presented remarkable
differences in 23 metabolites which participated in metabolic
pathways of the urea cycle, Krebs cycle, protein synthesis,
aspartic acid metabolism, alanine metabolism, ammonia circulation,
and the malic acid-aspartic acid shuttle, compared to normal
matched group (84).
Interestingly, the plasma concentrations of fumarate, malate,
citrate, α-ketoglutarate, and pyruvate were detected to be lower in
the taurine-supplemented breast cancer group relative to those
derived from normal matched group (84). As a result, the antitumor activity
of taurine was partially ascribed to the inhibition of aerobic
glycolysis and the downregulation of enzymes involved in Krebs
cycle (84). The beneficial impact
of taurine was attributed to the interference with energy
metabolism of breast cancer cells.
Taurine has also exhibited its favorable effect
against mammary carcinogenesis through its regulatory effect on the
extracellular matrix (ECM), thus attenuating breast cancer
recurrence. The therapeutic efficacy of taurine was ascertained in
either estrogen receptor-dependent breast cancer cells (MCF-7) or
estrogen receptor-independent breast cancer cells (MDA-MB-231).
Indeed, taurine decreased the expression levels of matrix
metalloproteinase 9 (MMP-9) and VEGF which are crucial proteins for
the degradation of the ECM (85)
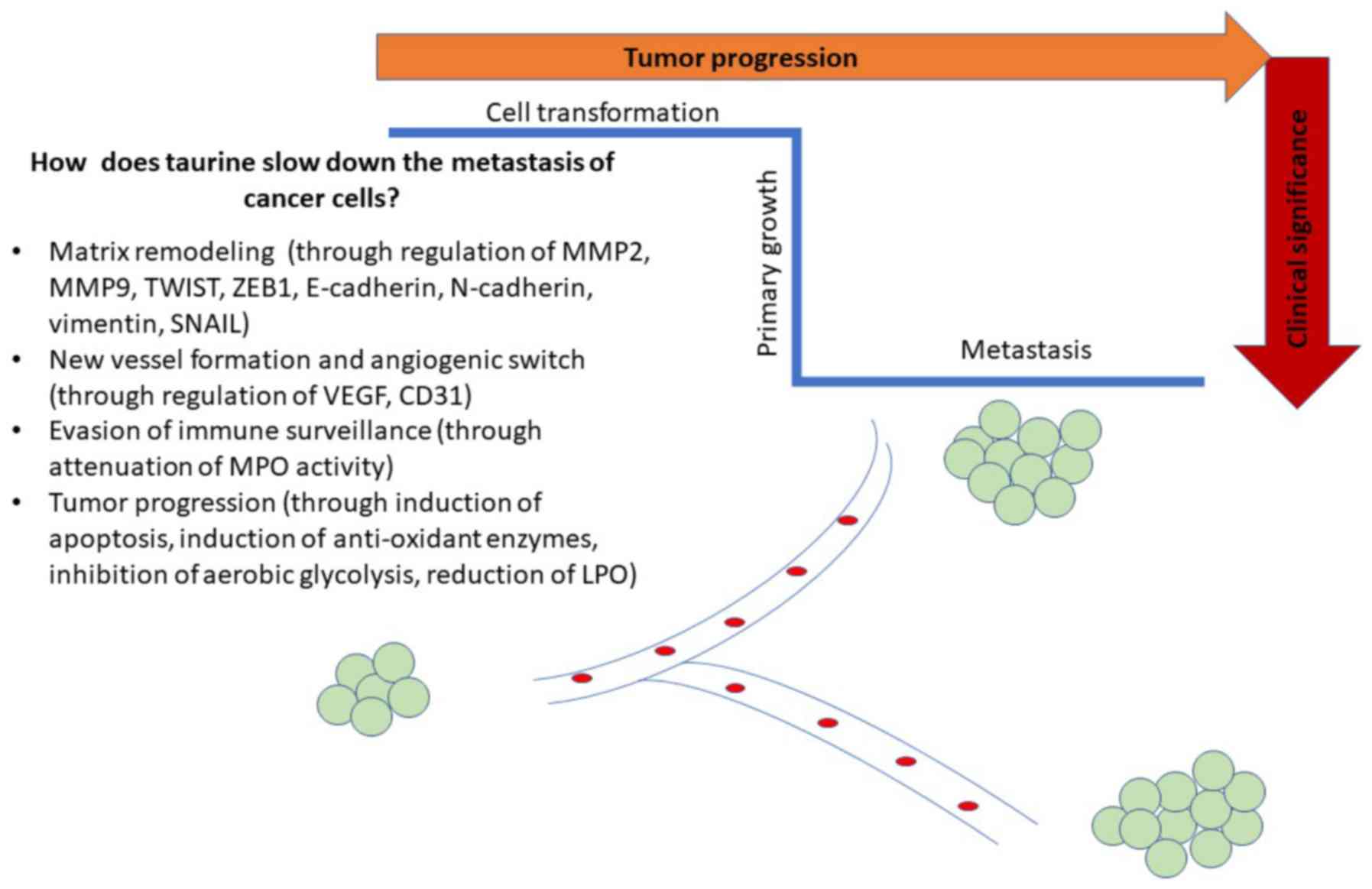
and angiogenesis (51) (Fig. 1). In that sense, taurine
compromised metastasis in both breast cancer cell lines,
independently of the presence of estrogen receptor. Besides, it
should be noted that estrogen exerted its significant effect on
taurine uptake, through increased expression of TauT transporter in
MCF-7 cells (86). It has been
reported that Na+-dependent uptake of taurine through
TauT transporter was activated by 17β-estradiol and p53
transcription factor, as shown by experiments in MCF-7 cells
(87). As a result, taurine
appears to be an attractive therapeutic agent because it can slow
down the metastasis of breast cancer at aggressive stages,
independent of the presence of estrogen.
Apart from the anti-oxidant and anti-angiogenic
effect of taurine, it has been shown that taurine displays a strong
chemo-preventive effect on breast cancer (88). In support of this, taurine has been
proved to alleviate methotrexate (MTX)-induced oxidative injury, by
modulating immune response (89,90)
and by attenuating toxic side effects on renal cells, due to TAM
administration in breast cancer cells (89,90).
The results were consistent with data derived from a clinical
setting where cancer patients have shown alterations in plasma
amino acids including taurine relative to their matched controls
(91,92).
In colon cancer, the main anti-tumor mechanism of
taurine is based on upregulating apoptosis at both the
transcriptional and translational levels (42,58).
For example, Zhang et al (42) have supported that taurine induced
the transcription and translation of the PUMA gene in HT-29
colorectal cancer (CRC) cells. Focusing on the molecular mechanisms
of taurine in more depth, taurine suppressed p53−/−
tumor cells more efficiently than p53+/+ tumor cells,
indicating that the apoptosis-stimulatory action of taurine is the
consequence of not only mitochondrial apoptotic pathway but also of
multiple signaling pathways in colon cancer cells. In support, Liu
et al (43) have shown that
the mammalian sterile 20-like kinase 1-c-Jun N-terminal kinase
(MST1-JNK) signaling pathway was essential for taurine-induced
apoptosis in colon cancer cells (Caco-2 and SW620 cells). Following
the treatment of colon cancer cells with taurine, the JNK signaling
cascade was activated, either by transmitting direct signals to the
MST1 target gene or by controlling the action of MST1 target via a
feedback mechanism, with the ultimate aim of inducing apoptosis
(43). Importantly, the
growth-inhibitory effect of taurine was also proved either in
colitis-model induced by TNBS (16) or in another colitis-inducible model
caused by DSS (24). In
particular, taurine appeared to alleviate clinical symptoms of
colitis through its inhibitory action on diarrhea/bleeding,
normalizing colon length, restoring histopathological alterations,
and compromising the activity of myeloperoxidase (MPO) (24). In addition, the beneficial effect
of taurine was ascertained in conditions where the MPO enzyme was
absent. In that direction, it was shown that taurine protected
human intestinal epithelial Caco-2 cells (MPO deficient) from
oxidative damage, after their coculture with human macrophage-like
THP-1 cells (93). Paradoxically,
those research findings were incompatible with clinical data that
supported the increment of taurine in colon cancer patients
compared to healthy patients (94).
In prostate cancer, taurine has come to the
forefront of research through its interference with the metastasis
of tumor cells. Taurine seems to reduce the migratory potential of
androgen-dependent human prostate cancer cells, though targeting
matrix metalloproteinases (MMPs), which are considered crucial
enzymes for the degradation of ECM. For example, the increased
invasion of androgen-sensitive human prostate adenocarcinoma LNCaP
cells and of androgen-dependent human prostate adenocarcinoma PC-3
cells was attenuated at 48 h and 8 h, following treatment of cells
with taurine (53). In particular,
it has been shown that taurine (125-1,000 µM) reduced the
values of MMP-9 and stimulated the expression of epithelial markers
such as E-cadherin and tight junction components, in a
dose-dependent manner in prostate cancer cells (53). Notably, the increased expression of
epithelial markers was accompanied by a marked reduction of
mesenchymal genes such as N-cadherin, twist family BHLH
transcription factor 1 (TWIST1), zinc finger E-box-binding homeobox
1 (ZEB1), SNAIL, and vimentin in LNCaP cells in response to taurine
treatment (53). In this way,
taurine was proved to be a promising therapeutic tool, restricting
not only the migratory properties of androgen-dependent human
prostate cancer cells but also reducing the recurrence of cancer
with stem-like characteristics, thereby circumventing the
possibility of tumor chemoresistance (95).
Besides, the proliferation of androgen-dependent
human prostate cancer cells (PC-3 cells) has been supported to be
hindered through the action of taurine haloamines (either
N-arachidonoyl taurine or N-oleoyl taurine), that arise through
their conjugation with fatty acids, thereby raising the possibility
of using taurine haloamines as favorable therapeutic agents
(54). Notably, there are two
signaling pathways, that account for the distribution of N-acyl
taurines in human prostate adenocarcinoma. In particular, fatty
acid amide hydrolase (FAAH) mediates the hydrolysis of N-acyl
taurines (N-arachidonoyl taurine, N-oleoyl taurine) which are
subjected to further catabolism (96). The FAAH has been shown to hydrolyze
both N-arachidonoyl taurine and N-oleoyl taurine (97). Interestingly, the silencing of the
FAAH enzyme can culminate in the concentration of N-acyl taurines
in the liver, kidney, and the central nervous system, reaching
micromolar levels (98).
Likewise, taurine has been suggested as a diagnostic
marker in bladder cancer, given that taurine levels were elevated
in the endometrial wall of bladder cancer patients (56). In the urine of bladder cancer
patients, the concentration of taurine seemed to be significantly
elevated, as its value was below the sensitivity limit of 400 MHz
in control cases (56).
Additionally to the functional significance of taurine in the
diagnosis of bladder cancer, it has been suggested that taurine was
effective in forestalling the proliferation of cervical cancer (CC)
SiHa cells, through induction of apoptosis. The underlying
molecular mechanism of taurine was based on upregulation of MST1
signaling pathway signaling pathway, leading to increased p53
nuclear transcriptional translocation (55).
Furthermore, taurine has conferred protection
against liver injury through its anti-oxidant properties (99). Several examples have demonstrated
that taurine ameliorates the cytotoxicity mediated by various
chemical compounds such as hydrazine, 1,4-naphthoquinone, and
carbon tetrachloride (100) and
by xenobiotics (101-103). In the case of arsenic-induced
cytotoxicity, taurine has been shown to protect damaged
hepatocytes, mainly by quenching free radicals and by detoxifying
toxic metabolites (104). Taurine
has also been illustrated to exert its cytoprotective effect
against liver injury, either by interfering with LPO/protein
oxidation or by reducing the accumulated hydrogen peroxide
(H2O2)/hydroxyl radicals (•OH) or by binding
to ferrum (Fe2+) like a chelator (105). Apart from its anti-oxidant
activity, taurine has been illustrated to fortify hepatocytes
against damage, by preventing osmolytic disturbance through ion
overloading in the mitochondrial matrix (15).
Since taurine exerts beneficial effect on acute
liver injury, it is plausible that taurine might be effective in
abnormal cases of chronic liver injury such as hepatocarcinoma.
Chronic liver injury exerts selective pressure on specific targets
in the microenvironment, driving the neoplastic transformation of
hepatic cells (106).
Hepatocellular cancer (HCC) development can be stimulated in an
inducible manner by diethylnitrosa-mine (DEN). The molecular
mechanism of DEN is based on triggering irreversible hepatocellular
necrosis coupled with compensatory proliferation (107). The DEN-mediated hepatic damage
becomes apparent through increased oxidative stress in hepatocytes
and it is followed by radical-based hepatic metabolic disturbance
(108). It is important to refer
to cytochrome P450 system (CYPs), especially CYP2E1 (109) that bio transforms DEN carcinogen
to the enhanced generation of ROS (107,108), by causing structural alterations
through the formation of alkylated DNA adducts in hepatocytes
(107). In this way, there is an
aberrant regulation of redox homeostasis and stress adaptation in
hepatocytes after DEN administration. In this context, taurine has
been proved to help inhibit oxidative stress-related hepatic injury
in DEN-treated rats. In particular, a single dose of taurine was
shown to reverse the action of DEN carcinogen (200 mg/kg), by
ameliorating the oxidative stress related-hepatic injury in mice
(92,109) and rats (108). In molecular setting, taurine
appeared to protect rat hepatocytes from DEN challenge, by
normalizing the values of disturbed enzymes such as serum alanine
aminotransferase (ALT), aspartate aminotransferase (AST), lactate
dehydrogenase (LDH), γ glutamyl transferase (GGT) activities
(110) or by interfering with LPO
(111). The main underlying
mechanism of taurine was based on reducing oxidant responses
including malondialdehyde (MDA), protein carbonyl (PC), and
nitrotyrosine levels. However, the activity of anti-oxidant enzymes
such as glutathione (GSH), glutathione peroxidase (GPx) levels,
super-oxide dismutase (SOD), and GSH transferase (GST) remained
unchanged in DEN-challenged rats following taurine administration
(110). Similarly, the anti-oxidant nature of taurine was
confirmed in other settings of oxidative damage mediated liver
injury. For instance, subcutaneous administration of taurine (2.5%
w/w) was proved to improve histopathological findings within the
time-window of 2 months in the liver of rats that had previously
been subjected to subcutaneous injection of galactose 300 mg/kg for
5 days per week (112). Taurine
ameliorated serum ALT, AST activities without any effects on the
anti-oxidant responses such as SOD, GPx in rats harboring
galactose-induced liver damage (112). Another example supporting the
advantageous effect of taurine was observed in ethanol-induced
hepatic dysfunction, due to its anti-oxidant nature, whereas rats
treated with β-alanine (taurine transporter inhibitor) presented
high susceptibility to ethanol-mediated liver damage (113).
Apart from its anti-oxidant activity, taurine
taurine has been suggested as a promising effective agent against
liver injury, by ameliorating membrane disintegration (114), inflammation (115), and calcium distribution (116). An interesting example supporting
the anti-inflammatory effect of taurine was observed in the model
of LPS-induced liver injury. Taurine conferred protection to
hepatic cells from LPS-induced liver injury due to its
anti-inflammatory nature, by reducing the secretion of
pro-inflammatory mediators (including TNF-α and IL-6) and by
elevating anti-oxidant responses [heme oxygenase-1 (OH-1), SOD]
(115). In another case, taurine
was documented to suppress the progression of alcoholic liver
disease (ALD) in male Wistar rats, by preventing the transmission
of signals through the LPS signaling pathway, in turn preventing
the possible activation of Kupffer cells. In particular, the
administration of taurine was reported to downregulate TNF-α, ILs
(IL-1β, IL-6), lipopolysaccharide-binding protein (LBP), cluster of
differentiation 14 (CD14), and nuclear factor-κB (NF-κB) (117). The underlying mechanism of action
of taurine was based on blocking the LPS-induced increase of
calcium (Ca2+) in Kupffer cells, taking into
consideration that intracellular calcium (Ca2+) plays an
important role in LPS-stimulated cytokine production, during
activation of Kupffer cells (116). In line with above, taurine
hindered the phagocytosis mediated by Kupffer cells and reduced
eicosanoid/TNF-α formation (118), thereby providing cytoprotection
against damaged hepatocytes, due to its inhibitory action on the
infiltration of immune cells in the liver and due to its
osmo-regulatory properties.
Hence, researchers have examined the possible
synergistic effect of taurine with curcumin in vitro as well
as in vivo conditions and they have supported that a
treatment scheme consisting of taurine combined with curcumin could
boost immune cell populations, culminating the therapeutic efficacy
of curcumin in hepatocarcinoma. In particular, that combination
treatment scheme was proposed to activate CD4+ T-helper
cells and to recruit CD8+ T-cytotoxic cells in cultured
human hepatoma (Huh-7) cells (119). Also, the treatment scheme of
taurine combined with curcumin was able to eliminate potential
malignant changes and to normalize IL-2, interferon-γ (IFN-γ),
α-fetoprotein (AFP) and α-L-fucosidase (AFU) levels in
DEN-stimulated model of hepatocarcinogenesis (120).
In addition, taurine has been reported to inhibit
the proliferation of murine melanoma B16F10 cells through the
mitochondrial apoptotic pathway (121). The therapeutic effect of taurine
was also shown in melanoma (B16F10 cells), through its anti-oxidant
properties (47,121). Interestingly, the beneficial
effect of taurine was proved to be more pronounced in metastatic
melanoma, which is usually treated with IL-2 immunotherapy, through
the clonal expansion of lymphocytes (122). Besides, this type of IL-2
immunotherapy is effective against melanoma, but its response rates
are hampered by vascular leak and lymphopenia (122). When taurine was conjugated to
IL-2, taurine increased the efficiency of immunotherapy in a B16
melanoma pulmonary metastases model, by mitigating toxic
side-effects of IL-2 itself (122). Interestingly, taurine exerted its
protective mode on reducing the tumor burden and attenuated IL-2
toxic symptoms such as vascular leak syndrome and lymphopenia, in a
model of metastatic melanoma (122). The results enabled the
dose-escalation use of taurine, extending treatment scheme without
causing any clinical sign of autoimmunity (122). In that sense, taurine maximized
the anti-tumor effect of IL-2 immunotherapy in an in vivo
metastatic melanoma model (123).
The results became more understandable since IL-2 is importantly
involved in activated-induced cell death (AICD). Notably, T cells
are accumulated in order to defense tumor cells and they are
eliminated due to AICD, under the rules of self tolerance (124). Additional research findings
proved that the mechanism underlying the cytoprotective mode of
taurine was attributed to the partial down-regulation of
FasL-mediated apoptotic pathway in IL-2 sensitized Jurkat T cells,
but not freshly isolated T cells through interference with NF-κB
transcriptional activation (6).
Accordingly, it is important to be noted that taurolidine (a
taurine derivative) has been reported to prevent the possibility of
disease relapse in mice bearing B16F10 melanoma cells, that were
assigned to two different types of surgery (laparotomy or
laparoscopy). The implementation of taurolidine gained significant
traction due to its effect on recovering natural killer (NK) and
lymphokine-activated killer (LAK) cell function, enhancing the
functional properties of immune cells. As a result, taurolidine
abrogated the effects of surgical trauma on primary and metastatic
tumor growth without any interference with host anti-tumor
surveillance mechanism, suggesting its potential significance in
the management of tumor-bearing patients undergoing resection
(124).
Furthermore, many skin tumors have shown increased
susceptibility to glucocorticoids (GC), inducing robust responses
but eventually acquired resistance and relapsed. According to the
above viewpoints, Logotheti et al indicated that
N-Bromotaurine (TauBr) might be suggested as a new therapeutic
agent in the treatment of skin cancer cells that were GC
unresponsive due to GC receptor (GR) impairment (125). It was proved that the therapeutic
efficacy of TauBr arose through its synergism with cisplatin,
exerting a growth-inhibitory effect on GC-resistant cells and
thereby pointing out its GC-mimicking therapeutic effect (125). The results strengthened the
potential therapeutic use of TauBr in other epithelial cancer
types. Accordingly, taurine haloamines have exerted their action,
showing good efficacy, tolerance, and insignificant toxic effects
on patients who were refractory to conventional GC-based
anti-inflammatory therapies (4,126).
Taken together, there are several indicative clues
on the therapeutic effect of taurine but there is diversity among
results derived from individual cell lines of various malignancies.
There is a lack of a comprehensive and comparative view across
several cell lines of different malignancies. Further studies are
needed to address this challenge and to shed light on the actual
anti-cancer action of taurine.
Taurine attenuates the drug-mediated side
effects
It is well-established that the administration of
adjuvant chemotherapeutic drugs can lead to 5-year survival rates
up to 70% for patients with non-metastatic disease. Even though
this success is accomplished through supplementation of specific
chemotherapeutic drugs, various combinations of approved
chemotherapeutic agents (i.e., DOX, cisplatin, etoposide and
ifosfamide) do not further increase patients survival over 10 years
(127,128). The mechanism underlying the
efficacy of prescribed chemotherapeutic agents is non-specific,
thus offering a window of off-target toxicity (127,128). The function of multiple
chemotherapeutic agents is related to multiple common unbearable
complications including cardiotoxicity, nephrotoxicity, hearing
loss, and the development of secondary malignancies (127,128). It should be highlighted that
toxicity also poses a significant challenge to the successful
combination of existing therapeutic options, by exploiting the
therapeutic index exerted by individual molecularly targeted drugs.
Considerable attention should be directed to the extent of
overlapping emerging toxicities derived from a possible combination
therapeutic scheme, to maximize the therapeutic efficacy elicited
by distinct drugs.
Beyond chemotherapy-related toxicity, the cancer
recurrence is a topic of huge interest given that there are no
available drugs that can overcome the resistance mechanisms of
classic chemotherapeutic drugs. In this frame, a couple of studies
have suggested that taurine is regarded as a promising agent to
alleviate side-effects of several chemotherapeutic drugs and to
ameliorate therapeutic outcomes, bypassing some challenges of drug
resistance. Reinforcing this suggestion, the rational combination
scheme of taurine with either chemotherapeutic drug has seemed to
optimize the efficacy of existing standard treatment, by improving
patient outcome and minimizing resistance conferred by the standard
therapy. The appropriate time of the combination scheme is
important to afford benefit to patient treatment. The benefits of
combined drugs become apparent, exerting their action against
different signaling pathways, their culminated efficacy and their
reduced toxicity profiles. To select the appropriate combination of
taurine with other agents, we should take into consideration that
some cancer types display drug resistance due to redox disturbance,
i.e., the disequilibrium between ROS and redox-sensitive survival
proteins.
In 1992, it was reported that taurine content was
eradicated after chemotherapy (71), whereas the expression levels of its
precursor molecules remained constant (71). It has been highlighted that the
pleiotropic nature of taurine enables the increased absorption of
chemotherapeutic drugs (129),
the alleviation of stress-induced insults (130), the reduction of radiation-induced
injury (131) and the attenuation
of inflammatory injury (132).
Indeed, taurine holds great promise in some oxidative stress
conditions mediated by ammonia (133) or acetaminophen (134) or gentamicin (135), without displaying any adverse
effect. In addition, taurine represents an invaluable tool to deal
with drug-induced myelosuppression or immunosuppression, which
accounts for reducing the efficiency of either chemotherapeutic
agent and for increasing the possibility of infections in patients
with immunocompromised system (136). Considering the potential of
taurine to alleviate the oxidative or inflammatory injury caused by
other drugs, it is plausible that taurine enhances the
tumor-inhibiting ability of chemotherapeutic agents (14), without off-target toxicity. In
support of this notion, it has been proposed that taurine is
capable of minimizing the injury triggered by classic
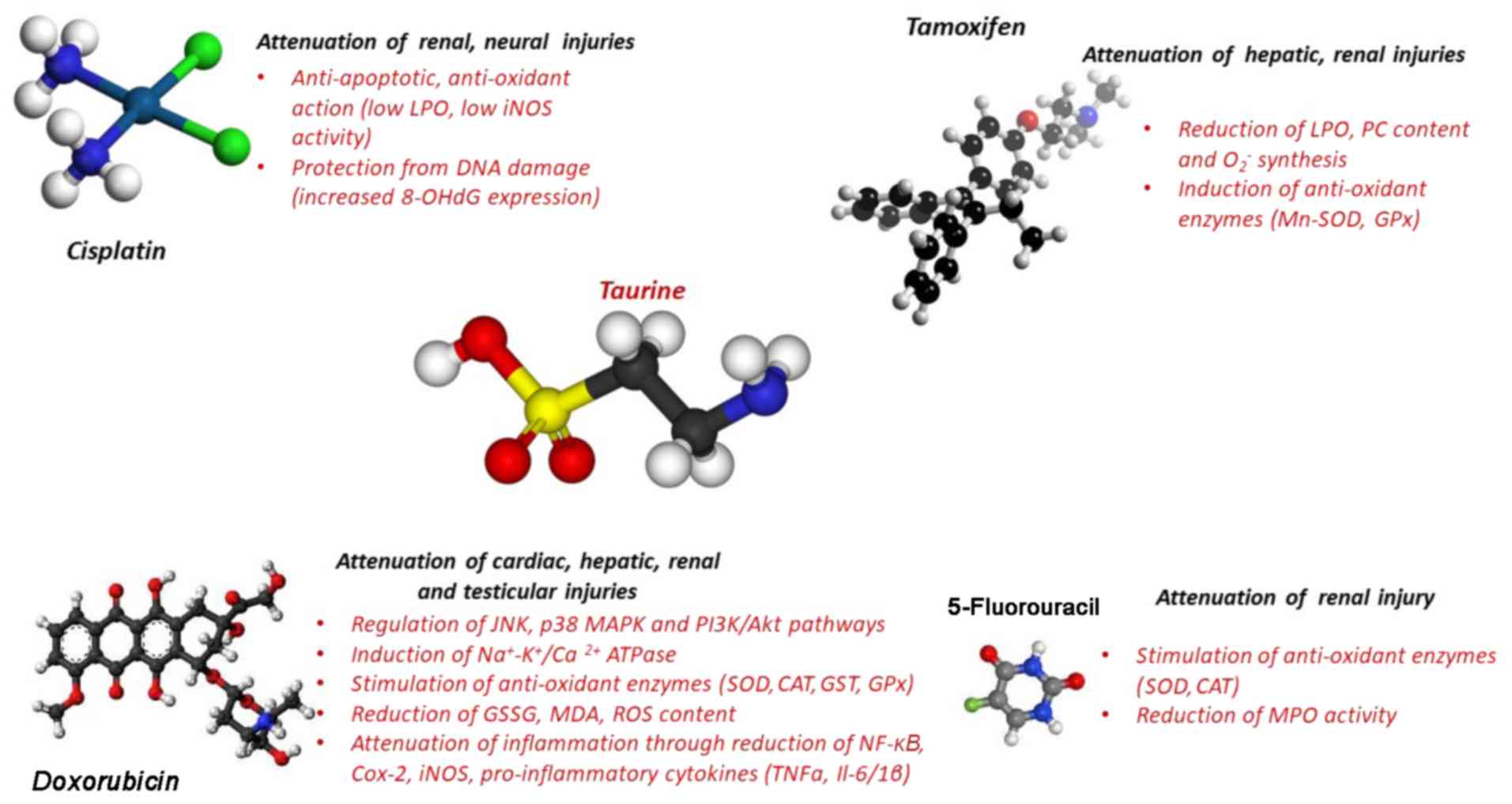
chemotherapeutic drugs such as as DOX (137), 5-fluorouracil (5-FU) (76), TAM (138), and cisplatin (139) (Fig.
2). DOX is one of the most widely used anthracycline drugs
against leukemia and sarcoma. DOX is regarded as a very effective
chemotherapeutic drug (78), due
to its capacity to prevent the replication of cancer cells as a
topoisomerase II inhibitor (140,141). As DOX forestalls topoisomerase II
action, it forms a high oxidative and pro-inflammatory environment,
thus causing DNA damage in cancer cells (140,142). Following DOX administration, the
DNA damage becomes apparent through DNA intercalation, which is
evidenced through bonds between nitrogen bases of the complementary
strands, thus causing disturbed DNA replication and transcription
(143). Despite the beneficial
impact of DOX against cancer, its use in the therapeutic arena is
impeded due to its toxic side effects including cardiomyopathic
failure and nephrotoxicity in patients (144-146).
Towards addressing the challenge of DOX mediated
toxicity (either nephrotoxicity or hepatotoxicity or
cardiotoxicity), researchers have proposed that taurine exerts
synergistic therapeutic effect with DOX to culminate its
therapeutic efficacy without off-target toxicity. Indeed,
DOX-induced cardiac and testicular injuries were attenuated owing
to the protection conferred by the anti-oxidant potential of
taurine (78). The protective
effect of taurine was also analyzed against DOX-induced testicular
oxidative complications (77). In
particular, 8-week old male rats were treated with either DOX alone
or taurine alone or taurine plus DOX within 28 days and it was
shown that taurine abrogated the DOX-induced testicular
side-effects, by reducing oxidative stress (reduced GSH, increased
GSSG and elevated MDA levels), by increasing activity of
antioxidant enzymes including SOD, catalase (CAT), glutathione
S-transferase (GST), GPx as well as membrane-bound enzymes such as
Na+-K+ and Ca2+ ATPases (77). The ameliorative effect of taurine
against DOX-induced testicular abnormalities relied on activation
of c-Jun N-terminal kinase (JNK), p38 mitogen-activated protein
kinase (MAPK) pathways, and p53 transcription factor (77). Similarly, taurine was documented to
provide marked protection against DOX-induced testicular damage,
due to its anti-apoptotic role (78). Furthermore, the concurrent
treatment scheme composed of DOX and taurine appeared to be
effective in neutralizing cytotoxicity in murine melanoma B16F10
cells, mainly through the taurine's ROS scavengering capacity
(143). The protective effect of
taurine against DOX-induced cytotoxicity was attributed to cell
cycle regulation and reduction of ROS production (143). Paradoxically, taurine employed
its anti-oxidant nature to bypass the barriers posed by DOX-induced
oxidant environment, thereby leading to DOX-mediated
hepatocarcinoma cells to apoptosis (147). It was highlighted that taurine
afforded protection against DOX-induced hepatoxicity through
elevating SOD activity and GSH content in the livers of DOX
intoxicated rats (147).
Taurine also acted as a renoprotective agent
against DOX-induced acute kidney injury (AKI), by inhibiting
apoptosis and inflammation. The ameliorative effect of taurine was
evidenced against renal-induced oxidative injury of eight-week-old
male Balb/c mice, that had previously been challenged with the DOX
(15 mg/kg body weight) for 24 h and then subjected to taurine
treatment (50 mg/kg or 100 mg/kg body weight) for 5 days (148). In particular, taurine
down-regulated the renal expression of apoptosis-related proteins
(p53, phospho-p53, caspase-9, and caspase-3) and renal expression
of inflammation-related mRNAs such as nuclear factor-κB (NF κB),
cyclo-oxygenase 2 (COX-2), and inducible nitric oxide synthase
(iNOS) (148). In another study,
taurine reduced the expression levels of pro-inflammatory cytokines
(TNF-α, IL-1β, IL-6, and COX-2) (149), that accumulated in renal tissues
of DOX-challenged animal models, proving its anti-inflammatory
action (150).
When taurine was administered to DOX-intoxicated
rats, taurine directed cardiac cells to defend against DOX-related
oxidative damage, thereby recovering them from the cell death
pathways. The protective mode of taurine against DOX-induced
cardiac oxidative stress was under the control of distinct
signaling cascades (78). In
particular, taurine ameliorated DOX-mediated injury, via the
inhibition of the p53 transcription factor, JNK, MAPK dependent
pathway and via the upregulation of phosphoinositide
3-kinase/protein kinase B (PI3K/Akt) pathway in cardiac cells
(78).
Another chemotherapeutic strategy is 5-FU, which is
directed to gastrointestinal solid tumors. The underlying mechanism
of 5-FU is based on causing oxidative injury, as shown by elevated
creatinine, blood urea nitrogen, and MDA content. However, the
protective effect of 5-FU is limited due to severe toxic effects,
including cardiac, renal, hepatic, diarrhea, myelosuppression,
dermatitis, and reproductive system anomalies that arise (151-156). The action of 5-FU is
non-specific, exerting its action not only in cancer cells but also
in normal healthy cells, thereby leading to genomic instability and
the accumulation of different toxic metabolites. When 5-FU is
absorbed in renal cells, nephrotoxicity emerges due to reduced
activity of either CAT or SOD and because of increased apoptosis
(157). In this context, it has
been shown that taurine alleviated FU-mediated side effects and in
turn, increased 5-FU therapeutic efficacy (58,76).
The ameliorative effect of taurine on FU-mediated complications
became obvious through counteracting FU-induced histological
changes such as distortion of normal cellular architecture,
infiltration of inflammatory cells, and loss of cellular integrity
(76). The underlying mechanism of
taurine was based on reversing the increased MPO activity to
eradicate FU-mediated abnormalities (76). Similarly, taurine proved to be
effective in reversing sulfasalazine-mediated effects, owing to its
anti-oxidant nature. In particular, taurine was presented as a
recommended option against Crohn's disease, through its inhibitory
action on LPO, and GSH status in both hepatic and renal cells
(158).
TAM is another therapeutic option against various
cancer types. The beneficial effects of TAM have shown to be
hampered by side effects that arise in the liver (73), kidney (72), and breast (159), thereby preventing its clinical
efficacy. Apoptosis, overproduction of toxic metabolites as well as
elevated LPO are the main routes by which TAM displays its toxicity
(160). Some studies have
reported that taurine exerted protective action in vivo
against TAM-induced hepatotoxicity (73) or nephrotoxicity (72). Taurine appeared to be effective in
reducing LPO, PC content, and O2− synthesis,
thereby ensuring normal redox homeostasis and maintaining the
integrity of hepatic cells in TAM-treated animal models (73). Taurine seemed to be indispensable
in restoring mitochondrial electron transport chain function in
mouse liver mitochondria of TAM-treated animals, either by its
ROS-scavenging capacity or by increasing activities of anti-oxidant
molecules such as mitochondrial manganese-dependent SOD (Mn-SOD)
and GPx (74), taking into
consideration that taurine itself was devoid of apparent
mitochondrial toxicity (74). As a
result, taurine afforded protection to hepatic cells of TAM-treated
animal models, either by reversing the decline of antioxidants or
by the direct free radical-scavenging activity of taurine (74). Besides, it is important to mention
that taurine proved to abrogate TAM-induced mitochondrial oxidative
damage, mainly through its anti-oxidant action (138) and its potential to induce
apoptosis in hepatic stellate cells (161).
In parallel, taurine appeared to confer protection
to cells from the toxic effects caused by the concurrent
administration of MTX and TAM. When MTX (10 mg/kg) and TAM (50
mg/kg) were intraperitoneally administered in Swiss albino mice,
after the pretreatment of mice with taurine (100 mg/kg) for nine
days, it was proved that pretreatment of mice with taurine seemed
to attenuate genotoxicity, through the synergism of two
chemotherapeutic drugs (162).
The underlying mechanism of taurine was based on increasing the
reduced GSH content and hindering chromosomal aberrations in both
somatic and germ cells. In that sense, it was proposed that taurine
provided therapeutic effectiveness not only alleviating toxic side
effects but also preventing the incidence of tumor recurrence
following chemotherapy (162).
Cisplatin (CDDP) is another classical
chemotherapeutic agent that is commonly prescribed in treatment for
a wide range of solid tumors including testicular and cervical
carcinoma, because of its efficacy and low cost (163,164). However, its clinical
effectiveness is hindered due to its toxic side effects in hepatic
and renal cells (165).
Interestingly, it has been reported that cisplatin accounts for
renal dysfunction in a significant proportion of cancer patients
(25-35%) (166). Following
cisplatin administration, the patients developed apparent tubular
injury at the proximal tubular level due to the induction of
inflammation, oxidative stress, apoptosis, and hypoxia (167-169). The main mechanisms of
cisplatin-mediated nephrotoxicity were based on increasing ROS
formation, DNA oxidation, and TNF-α secretion through increased
NF-κB transactivation (170). An
interesting example was shown in ovarian cancer women with advanced
disease who acquired resistance to cisplatin and relapsed, as shown
by their shorter disease-free intervals (171). In that frame, taurine seemed to
inhibit ovarian cancer cell proliferation, by enhancing the
therapeutic efficacy of cisplatin and by alleviating
cisplatin-mediated side effects (172,173). The induction of mitochondrial
apoptotic cell death was the main underlying mechanism by which
taurine exerted its advantageous action in cisplatin-treated human
CC (174). In another study, the
beneficial effect of taurine was demonstrated to be based on
ameliorating oxidative DNA damage signals, through inhibition of
p53 nuclear transcriptional translocation and elevation of
anti-oxidant responses, thereby culminating in the therapeutic
efficacy of cisplatin (175).
However, it is worth mentioning that the cisplatin resistance of CC
A2780 cells, was manifested through osmotic disequilibrium due to
an increased taurine uptake from cells (176).
Additionally, researchers have provided deep
insight into the ways by which taurine mediated its protective
action against nitrative stress that is usually encountered as
renal injury in cisplatin-treated animal models (177). In one interesting case, a single
intraperitoneal injection of cisplatin (15 mg/kg, or 25 mg/kg) in
male Wistar rats deteriorated kidney function for 7 days and
taurine (5% w/v) was administered in drinking water of rats four
days before the injection of cisplatin (175). The precise mechanism underlying
the cisplatin-mediated nephrotoxicity was the oxidative stress and
taurine protected renal cells against cisplatin-induced
nephrotoxicity, through its anti-inflammatory capacity, its
potential to boost anti-oxidant responses, its anti-apoptotic
action and its ability to relieve from DNA damage insults such as
8-hydroxy-2-deoxyguanosine (8-OHdG) expression (175). Following treatment with taurine,
the expression levels of citrulline, iNOS, and 8-nitroguanidine
were decreased in cisplatin administered animal models (139). Besides, it is important to note
that taurine transporter function was proved to be dysfunctional in
disturbed renal conditions mediated by cisplatin (172). In that sense, the favorable
effect of taurine against cisplatin-induced acute nephrotoxicity
was illustrated to be consistent with the deficiency of taurine
transporter (TauT) in renal cells, following administration of
cisplatin (178).
The toxic effects of cisplatin are not only
directed to renal cells but also expand to neural cells. To prove
the advantageous effect of taurine against cisplatin-induced neural
injury, researchers intraperitoneally injected 10 mg/kg of
cisplatin in rats for 13 days and they observed various
histological changes including a marked decrease in the total
traveled distance, average speed, total mobile time, total mobile
episode, number of crossing and absolute turn angle, leading to
neurological defects (179). The
administration of 100 or 200 mg/kg taurine for 13 consecutive days
before cisplatin injection was reported to be amazingly effective
in improving neurological abnormalities of rats (179). Taurine treatment caused a marked
improvement in brain anti-oxidant status, which became apparent
through elevated acetylcholinesterase activity, decreased oxidative
stress indices [low nitric oxide (NO), and LPO levels], increased
survival of neural cells in the cerebral cortices, and in the
hippocampus (179). Moreover,
taurine eliminated the dendritic arborization and mean diameter of
the somata of pyramidal neurons in the cisplatin treated rats,
implying that taurine afforded protection against cisplatin-induced
neurotoxicity (179).
Additionally, it has been reported that the
challenge of either cisplatin or paclitaxel (PTX) chemoresistance
in both ovarian cancer cells (A2780 and OAW42) was bypassed through
the action of taurine which impeded cancer stem cell population.
Taurine treatment is a powerful way to enable ovarian cancer cells
to respond to the therapeutic efficacy of classic chemotherapeutic
drugs (180). Also, nuclear
magnetic resonance (NMR) spectroscopy supported that the long term
administration of metformin accounted for the upregulation of
taurine in ovarian cancer cells, that had previously displayed
strong insensitivity to either cisplatin or PTX. Therefore, taurine
was considered as the underlying factor that inhibited cancer stem
cell population.
Ifosfamide is a chemotherapeutic agent, which can
lead to proximal renal tubular injury that mimics Fanconi syndrome.
Fanconi syndrome is considered a disease of the proximal renal
tubules of the kidney in which glucose, amino acids, uric acid,
phosphate, and bicarbonate are passed into the urine instead of
being reabsorbed. The study by Badary (181) highlighted that ifosfamide
injections in animal models rendered them to display all the
characteristics of Fanconi syndrome such as wasting of glucose,
electrolytes, and organic acids, along with increased serum
creatinine and urea, and diminished the creatinine clearance rate.
Taurine markedly attenuated some signs of renal dysfunction induced
by ifosfamide, through various mechanisms: diminished creatinine,
urea and albumin serum levels due to elevated creatinine clearance
rate and a marked decline in total and fractional excretion of
Na+, K+, PO4−3 and
glucose (181). However, taurine
did not alter the efficacy of ifosfamide in mice with
Ehrlich-Lettre ascites carcinoma (EAC) cells (182).
In the meantime, the ani-neoplastic effect of
taurine has arisen in great interest, due to its capacity to
orchestrate the inflammatory milieu. It is well established that
chemotherapeutic drugs exert their effect not only on cancer cells
but also on the strongly proliferated bone marrow hematopoietic
cells (183,184). Taurine has been presented as a
promising agent to circumvent chemotherapy-induced side effects,
due to its known immune-regulatory properties. Some researchers
believe that taurine is an effective immune adjuvant, which can
play a role in chemotherapy drugs, and has multi-directional
advantages (185). For example,
the beneficial action of taurine has been proved to be helpful in
attenuating the side effects of chemotherapy, thus potentiating the
immune function of mouse T-cell lymphoma. After quantification of
pro-inflammatory mediators IL-4, IL-12 and IFN-γ, it was observed
that there was a greater decline in the
taurine/chemotherapy-treated group of mice compared to the
chemotherapy group (186). In
addition, taurine emerged as the promising agent that bypassed the
toxic injuries derived from the combination of gemcitabine and
cisplatin, thereby maximizing the efficacy of chemotherapeutic
drugs (187). The therapeutic
efficacy of taurine was presented very strongly against peripheral
T-cell lymphoma, given that the tumor inhibition rate was
remarkably higher in the group treated with chemotherapy drugs and
taurine compared to chemotherapy group alone (187). Taurine exerted its ameliorative
action, by normalizing Th1/Th2 cytokine levels in both spleen and
thymus (186).
Of note, the Lewis lung carcinoma-bearing mice
presented accelerated tumor regression following taurine treatment
(40, 80, and 160 mg/kg) combined with cyclophosphamide, compared to
the chemotherapeutic group alone. Interestingly, all the doses of
taurine treatment increased the classic parameters of the immune
system (lymphocytes, macrophages, neutrophils), as demonstrated by
elevated bone marrow nucleated cells, augmented white blood cells,
increased spleen index as well as elevated thymus index (14). Alleviation of myelosuppression and
elevation of the phagocytic activity of peritoneal macrophages were
the main mechanisms behind the immunoregulatory role of taurine
against cyclophosphamide-induced damage. In that sense, taurine
reinforced cellular immune function and attenuated the
immunosuppression of cyclophosphamide (14). Accordingly, recent findings proved
that taurine up-regulated T cell responses in the thymus of
immunosuppressive mice, that had previously been injected with
dexamethasone (Dex) for 7 days. In particular, long-term taurine
supplementation (at a dose of 200 mg/kg for 30 days) was presented
to be remarkably effective in the development of T lymphocyte
subpopulations. Interestingly, taurine significantly increased the
number of CD4− CD8− double-negative (DN),
CD4+ CD8+ double-positive (DP),
CD4+ single-positive (CD4+) and
CD8+ SP (CD8+) cells in Dex-treated mice
compared with the control group. Furthermore, the
CD4+/CD8+ cell ratio did not display any
difference between thymus of Dex-induced immunosuppressive mice,
without or with the administration of taurine (136). From a clinical perspective, it
was highlighted that taurine attenuated the immune-suppressing
adverse effects of cyclophosphamide therapy by boosting the
phagocytic capacity of macrophage and neutrophil cells to dampen
inflammatory responses (14).
Similarly, young adults with acute lymphoblastic leukemia (ALL)
were characterized by lower incidence of febrile episodes,
neutropenia, and infectious complications following taurine
treatment compared to the placebo group that had received one of
the classic chemotherapeutic strategies (188). During taurine treatment, the
numbers of leukocyte populations were elevated, explaining why the
overall episodes were lower in ALL patients (188). In that way, taurine exhibits
immune-regulatory properties to ameliorate the unbearable
complications present in ALL. In the same context, it was reported
that chemotherapy mediated adverse effects (nausea, vomiting) were
attenuated through taurine supplementation in patients bearing ALL
and receiving one chemotherapeutic scheme (70).
2. Formation of taurine haloamines
In the regions of inflammatory or infected tissues,
neutrophils are recruited and they are regarded as the first-line
defense by eradicating the invading microorganisms through the
production of either oxidants or microbicidal proteins (189-191). When neutrophils engulf invading
microbes, superoxide anion (O2−) formation is
increased at the expense of adenosine triphosphate (ATP) synthesis
due to the action of nicotinamide adenine dinucleotide phosphate
(NADPH) oxidase. This occurs since the dysfunctional respiratory
chain accumulates electron donors, thus leading the transfer of
electrons from NADPH oxidase to oxygen, leading to an oxidative
burst. Then, O2− undergo a dismutation
reaction, converting them to accumulated
H2O2. In activated neutrophils, MPO enzyme
uses H2O2 to react with halides
(chloride/Cl− 2 or bromide/Br–), producing
hypohalous acids (HOCl or HOBr) which are very toxic oxidants,
impairing cell homeostasis (190,192,193). It is important to mention that
hypochlorous (HOCl) and hypobromous (HOBr) acids are highly
reactive but unstable oxidants with strong microbicidal and
cytotoxic activities (5,194).
In activated neutrophils, taurine fulfills its
cytoprotec-tive and antioxidant properties through the reaction of
taurine with HOBr or HOCl, contributing to the formation of taurine
haloamines including N-Chlorotaurine (TauCl) or N-Bromotaurine
(TauBr), respectively (4). It is
important to note that hypohalous acids arise from the
neutrophil-myelo-peroxidase (MPO) or eosinophil peroxidase (EPO)
halide system of metabolism during inflammation (193,195). In this way, taurine serves its
primary role to protect neutrophils from their self-destruction
caused by the hypohalous acid-mediated oxidative injury under
inflammatory conditions (4). Also,
taurine protects the surrounding cells from the inflammatory and
oxidative damage, through the generation of taurine
halo-amines.
It is commonly accepted that taurine haloamines are
long-lived oxidants that are less toxic than hypohalous acids and
confer protection against oxidative stress in inflammatory sites.
Due to the antimicrobial and antiseptic properties of taurine
haloamines, they seem to be invaluable in the treatment of local
mucosal and skin infections (196). Between taurine haloamines, TauBr
has stronger microbi-cidal activity and is more potent
membrane-permeable than TauCl (197). In contrast, TauCl is thought to
be more stable than TauBr, explaining its use as a local curative
agent in a wide spectrum of infections (126). Interestingly, TauCl is considered
to be a charged molecule, with low permeability capacity that
renders impossible the inactivation of the highly sensitive thiol
enzyme glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (198-200).
It should be highlighted that taurine haloamines
are only produced in O2−-generating
neutrophils. Interestingly, neutrophils derived from chronic
granulomatous disease (CGD) patients are unable to produce TauCl,
because they cannot genetically produce O2−
(201). The O −2
producing neutrophils are equipped with NADPH oxidase to ensure the
formation of taurine haloamines in conditions of oxidative burst
(201). In this sense, taurine
haloamines serve as important modulators of the immune system,
down-regulating pro-inflammatory cytokine production, ensuring the
compromised immune response. that influences the synthesis of
cytokines. However, taurine haloamines are not used in clinical
practice, because of their rapid degradation in blood. The
beneficial effects of taurine haloamines will be leveraged only if
the barrier will be circumvented. Therefore, stable TauBr compounds
such as N-monobromo-2,2-dimethyltaurine (Br-612),
N-dibromo-2,2-dimethyltaurine (Br-422) and Bromamine T (BAT) were
devised in an attempt to identify the anti-microbial and
anti-inflammatory properties of taurine analogs that were stable
(202).
3. Anti-microbial properties of taurine
haloamines
The loss of virulence and lag of bacterial regrowth
has been ascribed to the oxidizing effect of either TauCl or TauBr,
providing 'chlorine covers' or 'bromine covers' (creating either
covalent N-Cl or N-Br bonds) on the surface of target proteins
(203,204). When either TauCl or TauBr is
introduced into the cytosol, the chlorination or bromination is
followed and the oxidation of intracellular proteins is necessary
for complete eradication of pathogens (126). In particular, either TauCl or
TauBr has been shown to exert its action, mediating the chlorine or
the bromine transfer to the amino groups on proteins of microbial
membranes without the involvement of catalysts, suggesting that the
lone pair of electrons on the nitrogen atom of amino groups of
bacterial proteins associates with the chlorine atom of TauCl or
with the bromine atom of TauBr as an electrophilic chemical
reaction. It is important to note that the extent of chlorine
transfer reaction elicited by TauCl or the extent of bromine
movement mediated by TauBr depends on the type of microorganism
species, incubation time, pH, and the temperature (204).
In response to microbial and parasite infections,
neutrophils and eosinophils secrete abundant amounts of TauCl and
TauBr. Taurine haloamines are considered strong microbicidal
agents, eradicating a wide variety of Gram-positive or
Gram-negative bacteria, fungi, viruses, and protozoa (4), while taurine haloamines do not exert
any cytotoxicity to host tissues (4). For example, TauCl and TauBr have been
shown to kill the schistosomula of Schistosoma mansoni (205). Schistosomula can be eradicated by
1 mM TauCl or by 100 µM TauBr to nearly 40% (205).
As a general note, TauBr is an effective
therapeutic agent against chronic sinusitis, otitis media, acne
vulgaris, and periodontal diseases (126,203,206-212). Interestingly, TauBr killed a
specific type of skin bacteria (Propionibacterium acnes) and it was
used as a classic therapeutic agent in patients who developed
resistance to standard-of-care treatment (clindamycin). In the
clinical setting, the majority of patients displayed remission of
acne vulgaris symptoms at a rate of 65% after long-term treatment
with TauBr, without side effects (207,213). TauBr is supposed to exert greater
microbicidal effect than TauCl at very low concentrations (<10
µM) and neutral pH.
Even though TauCl was initially considered as a
molecule without bactericidal capabilities, TauCl is a potent
bactericidal compound, downregulating the extravagant bactericidal
potential that could be detrimental to the host. In this sense,
taurine chlorination confers the advantage of compromising
HOCl-induced tissue damage while sustaining anti-microbial
properties. The bactericidal potential of TauCl is ascribed to the
oxidation of sulfhydryl groups which are in the bacterial cell
membrane. TauCl exerts its action, neutralizing both gram-positive
(Staphylococcus aureus and Staphylococcus
epidermidis) and gram-negative bacteria (Escherichia
coli, Pseudomonas aeruginosa, and Proteus
mirabilis) when it was administered at the following
concentration range (12.5-50 µM) (209). It has also been shown that
taurine imparted its preventive action against Candida spp.,
Aspergillus spp., Fusarium moniliforme and
Polytrichum commune (126), and inactivate viruses including
type 1 and 2 human herpes simplex virus (HSV), adenovirus, human
immunodeficiency virus (HIV)-1, and influenza viruses (126). Interestingly, the killing
capacity of TauCl against Shiga toxin of enterohemor-rhagic E.
coli, its molecular mechanism relied on oxidizing the thiols
and aromatic amino acids of the bacterial proteins (214). Accordingly, the lipophilic nature
of NH2Cl was incorporated into the hydrophobic bacterial
cell membranes, achieving phagocytosis of E. coli (190).
4. Anti-inflammatory and anti-oxidant
properties of taurine haloamines
At inflammatory sites, toxic hypohalous acids are
neutralized by taurine, generating taurine haloamines (TauCl or
TauBr) (215). TauCl and TauBr
are products of either MPO or EPO halide system and they serve as
modulators of the immune system (215). Following the activation of
neutrophils or eosinophils, the release of taurine haloamines is
accelerated to confer important protection to many nearby cells in
several respects from inflammatory injury (216) and to attenuate oxidative stress
(13,217-219). Initially, taurine haloamines have
been identified to confer protection to neutrophils from toxic
hypohalous acids (hypochlorous or hypobromous), which are
detoxified with the presence of taurine (4). Secondly, taurine haloamines have been
shown to exert strong microbicidal properties, neutralizing either
bacteria or fungi or viruses (126). Thirdly, taurine haloamines have
been illustrated to display strong anti-inflammatory properties
that are primarily related to the reduction of various
pro-inflammatory mediators such as TNF-α, ILs (IL-1β, IL-6, IL-8,
IL-12), NO, prostaglandin E2 (PGE2) and
chemokines in both rodent and human leukocytes (197,220-223). In particular, TauCl was
demonstrated to exert a strong anti-inflammatory activity in many
cell types (5,212,224) whereas TauBr was proved to
suppress the synthesis of pro-inflammatory cytokines (TNF-α, IL-6,
IL-10, IL-12p40) and NO in macrophages (220,225). In that sense, taurine haloamines
inhibited inflammatory cell trafficking at injured sites and
probably blocked the incidence of chronic inflammation (4). Importantly, it was proposed that the
anti-inflammatory action of haloamines relied on the induction of
heme-oxygenase-1 (HO-1) in a dose-dependent manner (215,225-227). The aforementioned results were
evaluated since HO-1 exerts a potent anti-oxidant and
anti-inflammatory action through degradation of heme to bilirubin,
free iron, and carbon monoxide (CO) (228). When HO-1 enzyme is upregulated,
CO production is elevated, subsequently enabling cells to be
functional and safe against oxidative injury caused by
overproduction of O2− and NO, though
inhibition of either NADPH oxidase or iNOS enzyme (229). Taurine haloamines have been shown
to play a crucial role in averting the conversion of acute into
chronic inflammation, thus impairing the possibility of chronic
inflammatory diseases. Taurine haloamines have been reported to
protect cells from inflammation-derived oxidative stress, through
elimination of toxic •OH and additional ROS formation, thereby
reducing the cytochrome catalyzed electron transfer to oxygen and
ensuring cellular homeostasis. Besides, it is important to note
that the anti-oxidant potential of taurine haloamines has been
highlighted to be accomplished in three different ways. One
possible mechanism was manifested through the conjugation reaction
of taurine with mitochondrial tRNA. In particular, Schaffer et
al (13), and Jong et
al (230) supported that
taurine inhibited O2− generation and is
required for normal respiratory chain activity as well as the
appropriate synthesis of ATP through the formation of mitochondrial
taurine-conjugated tRNAs. Alternatively, taurine haloamines
appeared to reverse the redox inequilibrium, by increasing the
expression of many antioxidant enzymes, such as HO-1, SOD, and GPx,
peroxyredoxin-1 (Prx-1), thioredoxin-1 (Trx-1), and CAT (4).
In inflammatory-associated conditions, the
therapeutic potential of taurine haloamines has been highlighted in
both in vitro and in vivo settings. The research
group of Marcinkiewicz has provided convincing evidence that
taurine haloamines blocked the synthesis of COX-derived eicosanoid
such as PGE2 in LPS/IFN-γ stimulated macrophages
(LPS/IFN-γ J774A.2 mfs) via enhancing HO-1 enzyme expression
without altering COX-2 expression. Besides, the inhibitory action
of taurine haloamines against PGE2 accumulation was
confirmed in HO-1 deficient environment (227). In contract, taurine did not exert
any significant impact on PGE2 levels in stimulated
macrophages (227). One potential
underlying hypothesis was that taurine haloamines induced HO-1
enzyme at inflammatory sites to confer protection to neighboring
non-activated cells against oxidative stress (227). The beneficial impact of taurine
haloamines was also shown in vivo, using DSS-induced
experimental colitis model. The colon cancer regression was
observed after the reaction of exogenously administered taurine
with endogenous hypohalous acids at inflammatory sites (24). The anti-inflammatory capacity of
taurine haloamines was probably based on their capacity to hinder
phagocyte function and impair respiratory burst (24).
In rheumatoid arthritis (RA), taurine haloamines
have been shown to inhibit the protein expression of IL-6 and
PGE2 with similar potency. Even though both taurine
haloamines are considered powerful regulators of inflammation,
TauCl has been shown to exert more predominant anti-inflammatory
effects compared to those elicited by TauBr. In particular, TauCl
inhibited IL-8 and VEGF synthesis secreted by fibroblast-like cells
(FLS) from patients with RA whereas TauBr did not affect the levels
of IL-8 and VEGF. Besides, neither agent exerted a great impact on
regulating NO generation and iNOS protein expression (221).
The anti-inflammatory capacity of TauCl has been
highlighted in all activated types of leukocytes in vitro
(231,232) and animal models of both acute and
chronic inflammatory diseases (233-235). In 1996, Quinn et al
(236) supported that TauCl
suppressed PGE2 expression, mediating post-translational
effects on COX-2 mRNA in RAW 264.7 macrophages exposed to LPS or
IFN-γ. Then, the anti-inflammatory activity of TauCl was proved in
macrophages in response to an inflammatory stimulus. Importantly,
it was documented that the anti-inflammatory ability of TauCl was
tightly linked to increased HO-1 activity in macrophages (LPS/IFN-γ
J774A.2 mfs), suggesting that TauCl was a strong inducer of HO-1,
without any effects on COX-2 protein expression (227). Regarding the molecular mechanisms
involved, TauCl used different ways to tame inflammation by
targeting gene expression of proinflammatory cytokines, cell
adhesion molecules, and pro-inflammatory mediators such as COX-2 or
iNOS in a cell type-dependent manner (224). In particular, other research
findings supported that TauCl hampered the synthesis of
pro-inflammatory mediators such as NO, TNF-α, ILs (IL-6/8), PGs in
RAW 264.7 macrophages of murine origin in exposure to LPS or IFN-γ,
illustrating its important regulatory effect on macrophage function
(237-240). In these cases, the suppression of
pro-inflammatory genes was consistent with inhibition of NO
production in stimulated macrophages following TauCl treatment
(237,240). Notably, the anti-oxidant activity
of TauCl (a detoxified form of HOCl) was ascribed to its preventive
action against the catalytic activity of iNOS directly by targeting
the enzyme rather than by interfering with the interaction of
cofactors with iNOS (240). Many
antioxidant proteins including OH-1, Gpx, Prx 1 and CAT were
reported to be increased, upon exposure of macrophages to TauCl
(241). Similarly, it was
reported that TauCl reduced the expression of
O2−, ILs (IL-6/8) in human polymorphonuclear
leukocytes (223,242). In another study, TauCl interfered
with indoleamine-2,3 dioxygenase (IDO) activation, contributing to
low expression levels of IFN-γ (243). As a result, the anti-inflammatory
properties of TauCl were reported to be tightly associated with the
inhibition of many pro-inflammatory mediators, such as
O2−, NO, TNF-α, IL-1β, -2, -6, -8, and -10,
PGE2, macrophage inflammatory protein-2 (MIP-2),
monocyte chemoattractant protein-1 and -2 (MCP-1/2), and MMPs
(4).
Regarding the underlying molecular mechanism of
TauCl in more depth, TauCl appeared to coordinate the synthesis of
pro-inflammatory mediators through the regulation of NF-κB
transcriptional transactivation (239,244,245). Beyond identifying NF-κB as the
master transcription factor, the landscape remained obscure as to
which signaling pathways were activated to regulate the activation
of the NF-κB transcription factor, in various cell types under
inflammatory conditions following TauCl stimulation. The research
pertinent to the anti-inflammatory action of TauCl was focused on
the regulation of MAPK, which are composed of JNK, p38, and
extracellular signal-regulated kinase (ERK), accounting for the
activation of the NF-κB transcription factor (246,247). On one side, it was mentioned that
TauCl at 1 mM (not taurine) suppressed LPS-mediated NO production
in a dose-dependent manner, inhibiting ERK phosphorylation and
retaining p38 activity in RAW 264.7 macrophages. Elucidating the
inhibitory effect of TauCl on the ERK signaling pathway,
researchers proved that the inhibition of Ras activation was the
main principle of TauCl activity (248). The attenuation of LPS-induced
inflammation relied on the downregulation of ERK and its downstream
NF-κB activation, considering that inhibition of Ras small GTPase
was the most profound cause behind the anti-inflammatory action of
TauCl, without affecting the activity of activator protein (AP)-1
(248). Nonetheless, ERK
activation was not observed in resting RAW 264.7 macrophages after
treatment with TauCl alone (248), but ERK signaling pathway was
affected in human vein endothelial cells in response to TauCl
(198). In Jurkat T cells, it was
proposed that TauCl did not exert any effect on ERK phosphorylation
(224). In the same frame, the
capacity of TauCl to induce HO-1 was reported to be modulated only
using p38 MAPK inhibitor (not ERK inhibitor) in J774.2 macrophages
(227). Consistent with the
above, it is plausible to consider that the effect of TauCl has
been employed in a cell-type dependent manner since some reports
support that both ERK and p38 are required for interfering
LPS-mediated NO production (249,250) and others have claimed that only
p38 activation is linked to LPS-mediated NO synthesis (251).
After a thorough scrutinization of research
reports, it was illustrated that TauCl of various concentrations
hindered NF-κB activation in distinct cell types of myeloid or
lymphocytic or mesenchymal origin (224,239,244). NF-κB activation was the main
causative mechanism by which TauCl caused the decline of
pro-inflammatory cytokines in both macrophages and leukocytes.
TauCl seemed to impart its anti-inflammatory action, hindering the
NF-κB transcription that is a cornerstone for the synthesis of
pro-inflammatory cytokines, either mediating oxidation of IκB-α in
methionine residue at position 45 (244) or decreasing phosphorylation of
IkB-α in serine residue at position 32 (239). The oxidation of IkB-α at
methionine 45 was the main mechanism of neutral-izing NF-κB
activation mediated by TauCl in Jurkat T cells activated by TNF-α
(244). Conversely, the
anti-inflammatory action of TauCl was evidenced through suppressing
the IkB-α phosphorylation of serine 32 in the activated NR8383
macrophage cells stimulated by LPS and IFN-γ (239). Similarly, TauCl appeared to
inhibit IL-1β-derived NF-κB DNA binding activity in fibroblast-like
synoviocyte cells (FLS) derived from RA patients (252).
Apart from the effect of TauCl on innate immunity,
many research reports have provided convincing evidence that TauCl
had a strong anti-arthritic effect, as shown in various
experimental animal models and samples isolated from RA patients
(233,253,254). Interestingly, TauCl seemed to be
remarkably effective not only in macrophages but also in
mesenchymal cells of inflammatory-associated disorders. For
instance, TauCl was demonstrated to mediate its preventive action
on pro-inflammatory mediators, by inhibiting the expression levels
of TNF-α, ILs (IL-6/8) in distinct adipose tissue samples of RA
patients [articular adipose tissue (AAT), subcutaneous adipose
tissue (ScAT)] as well as in samples of rats derived from
adjuvant-induced arthritis (253,255). There was also a marked reduction
in the production of pro-inflammatory cytokines (IL-6, IL-8, and
PGE2) secreted by FLS, that originated from the joints
of RA patients, following TauCl treatment (252,256,257). Behind the mechanism underlying
the ameliorative effect of TauCl against arthritis, Kontny et
al (256) proved that TauCl
was a specific and potent inhibitor of COX-2 protein expression in
fibroblast like synoviocyte cells of rheumatoid arthritis patients
(RA FLS) after IL 1β stimulation. In that sense, it was illustrated
that the cell viability of fibroblast like synoviocyte cells of RA
patients was reduced, following concurrent treatment with
platelet-derived growth factor (PDGF) and TauCl. The
anti-proliferative effect of TauCl was also demonstrated in
fibroblasts which had been stimulated with either basic fibroblast
growth factor (bFGF) or TNF-α. Following treatment with TauCl, the
inhibition of FLS proliferation was attributed to the increased
nuclear accumulation of the p53 transcription factor, causing the
cell cycle arrest (256,258). In addition, the attenuating
effect of TauCl against impaired FLS function of RA patients was
ascribed to reduced MMP synthesis (226,259). As a result, TauCl could
ameliorate RA-associated symptoms through its blocking effect on
inflammatory injury.
It is important to note that there are
controversial data on the impact of TauCl on collagen-induced
arthritis (CIA) course. Some researchers have reported that TauCl
hinders the development of CIA whereas others have proved that
TauCl alleviates the severity of symptoms (233,260). CIA can be applied to genetically
susceptible (DBA 1/J) mice after their immunization with native
type II collagen in adjuvant, to delineate the pathogenesis and the
signaling pathways involved in RA (4). On one hand, the onset of CIA has been
illustrated to be slowed down in mice that had received TauCl
before or after collagen injections, thereby reducing the
possibilities for arthritis emergence in these mice (233). The development of arthritis was
attenuated in the DBA1/J mice with CIA, following TauCl treatment,
independently whether TauCl therapy was applied early (after
primary immunization) or late (after booster immunization) during
the CIA course (233). Focusing
on the underlying mechanism of TauCl, it has been substantiated
that TauCl inhibited collagenase activity action, thereby delaying
the incidence of CIA arthritis (261). On the other hand, systemic
application of TauCl has proved to alleviate severe unbearable
complications, that were presented in the DBA1/J mice with CIA,
such as paw swelling, arthritic scores, cartilage damage, synovial
inflammation and bone erosion (260). In particular, TauCl mediated its
action through interfering with lymphocyte proliferation and
osteoclast formation, thereby leading to a strong remission of
synovial inflammation, amelioration of cartilage damage and bone
erosion through inhibition of NF-κB activation (260). Following TauCl treatment, the
synthesis of pro-inflammatory mediators (TNF-α, IL-1, and IL-6) was
also reduced, thus compromising bone destruction (4). Similarly, the therapeutic benefits of
TauCl have arisen in septic arthritis mediated by Staphylococcus
aureus, when TauCl was locally administered after a single dose of
Staphylococcus aureus (254).
TauCl has been reported to exert its protective action by delaying
the development of arthritis and ameliorating bone erosion and
cartilage damage (233,253,254). It should be noted that no
advantageous effects of TauCl were observed when bacteria and TauCl
were systemically administered (254).
In this regard, researchers have provided
compelling evidence that TauCl suppressed inflammation-mediated
bone destruction, through its capacity to interfere with osteoclast
formation. To determine the mechanism which was involved in the
inhibitory effect of TauCl on osteoclast formation, bones of mice
with CIA, and the receptor activator of NF-κB ligand
(RANKL)-stimulated bone marrow-derived monocyte/macrophage
precursor cells (BMMs) were used. The results proved that the
nuclear factor of activated T cells 1 (NFATc1) was the main
osteoclast-specific transcription factor that was negatively
affected by TauCl. As a result, reduced tartrate-resistant acid
phosphatase (TRAP), cathepsin K activity, calcitonin receptor, and
the impaired formation of multi-nucleated osteoclasts were observed
following treatment with TauCl (260). Those findings proposed that TauCl
might be leveraged as a novel therapeutic strategy against bone
diseases, which are characterized by excessive bone resorption.
Beyond the anti-arthritis mode of TauCl, the
beneficial effect of TauCl has become clear in mesenchymal cells of
RA, ultimately altering arachidonic acid metabolism that is
commonly associated with an inflammatory response. An interesting
research report by Kim et al (226) provided insight into the favorable
impact of TauCl on RA, by using FLS isolated from RA patients
following stimulation with adiponectin or IL-1β, and their
treatment with TauCl. In both modes of stimulation, it was proved
that TauCl was a promising inhibitor of MMPs (226). In the case of adiponectin
stimulation of synoviocytes, TauCl exerted a greater inhibitory
effect than that mediated by IL-1β stimulation, as evidenced by
higher nuclear shutting of NF-κB transcription factor (226).
Even though TauCl has been reported to hinder the
proliferation of TNF-α stimulated neutrophils by inducing
mitochondrial apoptosis (262),
TauCl has been shown to confer protection to phagocytic cells from
cell death caused by the overproduction of
O2− and H2O2 in an
inflammatory milieu. Specifically, TauCl has been shown to reduce
the proliferation in mitogen-stimulated human peripheral blood
lymphocytes (223), in
IL-3-dependent murine hema-topoietic prolymphocytic B cells
(263), in human skin fibroblasts
(264) and tumor cells (B
lymphoma osteosarcoma cell lines) (265,266). Growth-inhibitory effect of TauCl
was presented in distinct cell types in response to various
stimuli, confirming that there are conserved mechanisms that
warrant further investigation.
Thus, TauCl has emerged as a new therapeutic
option, following spinal cord injury. The anti-inflammatory action
of TauCl was linked to its anti-oxidant activity, as manifested by
the reduction of ROS and other inflammatory mediators including
TNF-α (267).
5. Therapeutic perspectives and clinical
studies
Even though advances have been reported in the
field of therapeutics, both cancer type-dependent drug responses
and chemo-resistant therapies necessitate their enrichment with
novel pharmaceutical agents. The role of pharmacology in medicine
is to investigate new therapeutic drugs by using appropriate models
to acquire a greater understanding of the molecular interactions
that determine the outcome of cancer cells. Several in vitro
and in vivo methods have been performed for the pre-clinical
assessment of anti-inflammatory drugs such as taurolidine or TauBr
or TauCl and some clinical case reports have been employed.
Initially, elimination of tissue damage has been
attributed to the anti-inflammatory, not the anti-bacterial
potential of taurine (4). So,
researchers examined the therapeutic effect of taurolidine as well
as TauBr and TauCl, which arise through the reaction of taurine
with hypohalous acids.
Taurolidine is a derivative of taurine and its
common use is directed to deal with various infections. From a
structural perspective, taurolidine is a
bis-(1,1-dioxyperhydro-1,2,4-thi-adiazinyl-4) methane, comprising
of two taurolidine rings derived from taurine and three molecules
of formaldehyde, by forming a two-ringed structure bridged by a
methylene group (268). The first
clinical report supported that taurolidine (taurine derivative)
could be used in the prevention of severe surgical infections
(sepsis, peritonitis and pancreatitis), given that taurolidine
harbors strong bactericidal activity against antibiotic-resistant
bacteria (269-271). Taurolidine exerts anti-endotoxin,
anti-bacterial, and anti-adherence properties. Taurolidine is now
included in a new catheter lock solution to hinder catheter-related
infections (272,273). It has been pointed out that
taurolidine may have anti-bacterial action which is independent of
the resultant taurine metabolites. The anti-bacterial activity of
taurine has also been ascribed to its anti-inflammatory properties,
as manifested by inhibition of IL-1 and TNF-α (274).
Interestingly, intravenous administration of 2%
taurolidine was given to a gastric cancer patient with liver
metastasis for 39 cycles, each of which lasted 7 days of treatment
per month (300 mg/kg body weight per day). Taurolidine therapy was
devoid of any toxicity and rendered cancer cells stable, without
enabling them to colonize to other sites. Despite the encouraging
results by taurolidine administration following chemotherapy, the
gastric cancer patient died due to myocardial infarction (275). Similarly, two glioblastoma
patients achieved tumor remission due to taurolidine application,
within 4 months but then, they succumbed to the aggressiveness of
tumor cells (276).
In order to use the beneficial effect of taurine in
a clinical setting, certain obstacles should be addressed. Taurine
is poorly absorbed in the gastrointestinal tract and so it is
plausible that taurine is metabolically degraded in both the gut
and the liver. Taurine is also characterized by unfavorable
pharmacokinetics, a very strong hydrophilic nature, lipophobic
character, and fast rate of extraction through urine, which
potentially explains the difficulties to use taurine as a
therapeutic agent. For this reason, further investigations prompted
to understand the anti-inflammatory properties of taurine
derivatives in a clinical setting.
Experiments have shown that TauCl exerts good
tolerability in human tissues, as it is used in the treatment of
infections of the eye, skin, outer ear canal, nasal and paranasal
sinuses, and oral cavity. For example, the therapeutic efficacy of
TauCl has been proven at phase II clinical studies for the
treatment of external otitis, crural ulcerations, and
keratoconjunctivitis (126,210,277). Similarly, it has been proved that
TauBr seems to be the most suitable topical agent for the treatment
of biofilm-associated infections such as chronic sinusitis, otitis
media, acne vulgaris, and periodontal diseases (4). The therapeutic efficacy of TauBr has
especially been observed in the treatment of biofilm-associated
infections (278). In another
study, forty patients with mild to moderate inflammatory facial
acne vulgaris were randomly enrolled and received either TauBr (3.5
mM) or classical antibiotic option (clindamycin-1%) for 6 weeks,
twice a day. The results were very encouraging concerning the
promising therapeutic action of TauBr because the symptoms of
patients with acne vulgaris were recovered in 80% of patients, with
acne lesions being reduced from 65 to 68%. Interestingly, TauBr
appeared to ameliorate inflammatory acne vulgaris lesions even in
patients with antibiotic resistance (207). In addition, multiple sclerosis
(MS) patient with herpes zoster skin presented attenuation
of clinical symptoms when 0.8% TauCl was administered for four
days, followed by concurrent treatment with 0.8% TauCl and 1.0%
TauBr in the next three days. Interestingly, the treatment scheme
composed of taurine haloamines was able to bypass the resistance
developed in a MS patient to valacyclovir (279). Recently, it was shown that
chronic multi-bacterial biofilm scalp infection was treated with
the combined topical application of the active halogen compounds
TauCl, TauBr and BAT (280). This
is the reason why taurine and taurine haloamines can be considered
potential drugs in infectious and chronic inflammatory diseases.
Certainly, the use of taurine haloamines as anti-inflammatory
agents entails some risks that remain unknown.
In conclusion, both in vitro and in
vivo studies as well as clinical trials will encourage us to
consider taurine and taurine haloamines as potential drugs in human
medicine, including infectious and chronic inflammatory disease.
However, taurine and taurine haloamines warrant further
investigation to examine their therapeutic efficacy in a clinical
setting. Along with optimizing the drugs brought to the clinic,
patient selection is a prerequisite for clinical trials. This
refined patient matching can be achieved by our growing
understanding of the interaction between signaling molecules
involved in either inflammation or cancer. Undoubtedly, more
studies are urgently needed to delineate mechanisms of action
elicited by either agent as well as their pharmacodynamic and
pharmacokinetic properties. Experimental models are need based on
pharmacological principles to predict the intended therapeutic
response of taurolidine, TauBr or TauCl.
6. Significance of lncRNA TUG1 lncRNA in
cancer
Long non-coding RNAs (lncRNAs) are outlined as
noncoding transcripts with a length of more than 200 nucleotides,
and they were originally discovered through the large-scale
sequencing of mouse cDNA libraries (281). As a general note, lncRNAs deserve
particular attention due to their involvement in many physiologic
processes and their reported abnormal expression in pathologic
circumstances, including cancer (282,283). Since the impaired expression is
tightly related to human malignant tumor formation (284), delineating how lncRNAs control
gene expression is the major focus for cancer research (285). Interrogating the function of
tumor-associated lncRNAs and elucidating their subsequent clinical
impact comes with a surge of excitement. LncRNAs are important
regulators in cancer progression through their participation in
cancer proliferation, cancer invasion, replicative senescence,
resistance to radiation and drugs, and reprogrammed energy
metabolism (286,287). At molecular setting,
tumor-associated lncRNAs can serve either (I) as decoys to direct
transcription factors in a spatial-temporal manner (288) or (II) as carriers to transmit
regulatory signals for transcription among cells (289) or (III) as scaffolds to aggregate
a variety of RNA-associated proteins in transcriptional complexes
(290) or (IV) as competitive
endogenous RNAs to interact with functional microRNAs to cause
their silencing (291) or (V) as
guide molecules to recruit chromatin-modifying enzymes, conferring
epigenetic regulation of target genes (292-294). LncRNAs are also critical for
regulating cellular biological processes, through their binding to
kinase proteins, thereby causing the respective conformational
changes (295). Tumor-associated
lncRNAs have revolutionized therapeutics in cancer research,
enabling an outpour of studies documenting the startling
contribution of lncRNAs to either cancer progression or
remission.
In this context, taurine upregulated gene 1 (TUG1)
is a lncRNA, that has been identified due to its upregulation
during retinal development in response to taurine treatment
(296). Accumulating evidence has
supported the overexpression of lncRNA TUG1 in different disease
contexts, including MS (297),
diabetes mellitus (DM) (298),
chronic kidney disease (CKD) (299), and chronic obstructive pulmonary
disease (COPD) (300). In the
diabetic nephropathy and CKD, one potential mechanism underlying
the action of lncRNA TUG1 is based on increasing the transcription
of peroxisome proliferator-activated receptor-γ coactivator 1α
(PGC-1), thus enabling the improved mitochondrial bioenergetics
(299,301). Similarly, lncRNA TUG1 has been
illustrated to mediate its ameliorative effect on pancreatic
insulin secretion and pancreatic islet dysfunction (302).
Besides, the lncRNA TUG1 has been regarded as an
eminent lncRNA in cancer progression (303) among multiple cancer-associated
lncRNAs (304,305). Many research studies have
provided a wealth of information on the significance of TUG1 lncRNA
in many cancer types, dictating its contribution to the tumor
progression as well as its prognostic value for unfavorable
survival of cancer patients (Fig.
3). The lncRNA TUG1 is presented to be dysregulated in a
variety of malignancies, indicating its significance in
orchestrating tumor landscape. An apparent conundrum is that lncRNA
TUG1 acts either in promoting cancer cell proliferation or
abrogating cancer progression. In particular, the expression of
multiple target genes can be controlled, either by activation or
suppression, through the action of lncRNA TUG1 in a cell-type
dependent manner, determining the end transcriptional result of
target genes (306). From one
perspective, accumulating evidence has proved that lncRNA TUG1
functions as an oncogene (303,307), predicting poor prognosis for
melanoma (308), bladder cancer
(309), sarcoma (310), hepatocarcinoma (311), and colon cancer (312). For example, lncRNA TUG1 has been
reported to be increased either in osteosarcoma due to its
modulation of diverse transcription variants (310) or in hepatocellular carcinoma due
to epigenetic suppression of Kruppel like factor 2 (KLF2) protein
(311), implying the significant
gene-regulatory activity of lncRNA TUG1 in cancer. Even though most
of the studies have illustrated the overexpression of this lncRNA
in cancer tissues as opposed to noncancerous counterparts, few
studies have supported the opposite trend. For example, it has been
reported that lncRNA exerts a tumor-suppressor role, being
downregulated in glioma (313)
and lung cancer [non-small cell lung cancer (NSCLC)] (314). It should be pointed out that the
tumor-suppressor role of lncRNA TUG1 is consistent with the
evidence supporting that promoter of lncRNA TUG1 contains conserved
p53 binding sites (314).
At the molecular setting, aberrant signal
transduction by lncRNA TUG1 has been mainly shown to be mediated
through its interaction either with Polycomb repressive complex 2
(PRC2) or with miRNAs (315)
(Fig. 4). There is convincing
evidence supporting that the regulatory function of lncRNA TUG1
could be elicited, through its binding to PRC2 complex, thus
reorganizing the transcriptional landscape in target genes through
chromatin remodeling (316). When
the lncRNA TUG1 functioned as a guide molecule to recruit the
chromatin-modifying complex to target genes, subsequent epigenetic
alterations such as changes in DNA methylation patterns of histone
modifications were followed, thus abrogating target gene
expression. For that purpose, researchers considered that the
lncRNA TUG1 was the main determinant behind chromatin orientation
and they tried to understand how lncRNA TUG1 spatially organized
the chromatin, affecting the expression of target genes involved in
tumorigenesis. Indeed, the association of lncRNA TUG1 with PRC2
complex caused the inhibition of specific genes through
methyltransferase activity conferred by the PRC2 complex, as it has
been observed in 20% of lncRNAs (317). Interestingly, it was proven that
the lncRNA TUG1 interacted with enhancer of zeste homolog 2 (EZH2)
enzyme, epigenetically causing the reduced expression levels of
tumor suppressor genes, through trimethylation of histone 3 at
lysine 27 (H3K27me3) in target genes (318), as shown in human NSCLC (319), gastric cancer (320) and hepatocellular carcinoma
(321). For example, it was
illustrated that overexpressed lncRNA TUG1 expression levels
epigenetically modified homeobox B7 (HOXB7) expression, through its
association with polycomb repressive complex 2 (PRC2), contributing
to remission of NSCLC (319).
Furthermore, Yang et al (322) showed that the association of
lncRNA TUG1 with the PRC2 complex was crucial to coordinate the
gene expression of transcriptional units in the three-dimensional
space. In response to growth signals, it was pointed out that
growth-regulatory genes could be shuttled between polycomb bodies
and interchromatin granules, according to the recruitment of PRC2
chromatin modifying complex to lncRNA TUG1 (322).
On the contrary, numerous related studies have
demonstrated that the regulatory network between lncRNA and
microRNAs, determined the tumor progression either positively or
negatively. Indeed, the lncRNA TUG1 exerted its action as a miRNA
sponge consistent with the known function of lncRNAs as competing
endogenous RNAs (ceRNAs), to antagonize the function of miRNAs
(323). The classical 'sponge'
function of lncRNAs is identified as the main mechanism accounting
for posttranscriptional regulation of target genes. LncRNA binds to
miRNA, competitively inhibiting the binding of the miRNA to its
target mRNA, thus stimulating the expression of the downstream
target mRNA (324). Moreover,
miRNA functions as an inhibitor of target mRNA, counteracting
target gene expression through its base-pairing with the
3-untranslated region (3-UTR) of target mRNA (325). In this direction, comprehensive
detailed research progress has been performed regarding the
inhibitory functions of lncRNA TUG1 against miRNAs. The lncRNA TUG1
was proved to be recruited at specific sites of following miRNAs:
miR-212-3p, miR-132-3p, miR-145, miR-26a, miR-9, miR-34a-5p,
miR-382, miR-300, miR-335-5p, miR-144, miR-138-5p, miR-219,
miR-142, miR-153, miR-299, miR-600, and miR-129-5p, thus inhibiting
the expression of aforementioned miRNAs and affecting cancer
progression in a cancer type-dependent manner (326-330). In addition, many studies shed
light on the involvement of lncRNA TUG1 in increased metastasis of
distinct tumor types, either directly targeting mesenchymal genes
or indirectly through its interaction with multiple miRNAs. Indeed,
lncRNA TUG1 interacted with the following miRNAs (miR-144, miR-145,
miR-26a, miR-9-5p, miR-34a-5p, miR-229, and miR-300), thus leading
to radioresistance, carcinogenesis, invasion, angiogenesis, and
blood-tumor barrier permeability (313,331-336). A characteristic example
demonstrated that lncRNA TUG1 directed glioma stem cells to the
uncontrolled growth, by impeding the degradation of stemness genes
by retaining miR-145 and hindering the expression of neural
differentiation-associated genes, upon Notch signaling (188). Another example showed that the
lncRNA TUG1 accounted for the upregulation of mesenchymal markers
(involving vimentin) and the down-regulation of epithelial markers
in CC (337). In that regard, the
inhibitory action of lncRNA TUG1 against distinct miRNAs was shown
to contribute to the exacerbation of cancer progression.
Apart from the contribution of lncRNA TUG1 to
cancer through either chromatin remodeling or sequestration of
miRNAs, the lncRNA TUG1 has been reported to determine the outcome
of various malignancies, through its involvement in various
signaling cascades. For example, the lncRNA TUG1 exerted its
regulatory action on either osteosarcoma or glioma or gallbladder
carcinoma or oral squamous cell carcinoma through its potential to
interfere with either PI3K/Akt or Notch or transforming growth
factor-β (TGF-β) or Wnt/β-catenin (331,338,339,340). Interestingly, it was proved that
the therapeutic efficacy of cisplatin could be increased through
overexpression of lncRNA TUG1 in breast cancer, relying on
inhibiting the Wnt signaling pathway through regulation of
miR-197/nemo like kinase (NLK) (341). In another case, lncRNA TUG1
gathered considerable attention as an oncogene in colon cancer, due
to its capacity to enable the constitutive transmission of
Wnt/β-catenin signals (342).
Similarly, it was proved that lncRNA TUG1 enabled the pancreatic
cancer cells to acquire mesenchymal characteristics, by
competitively inhibiting TGF-β/Smad signaling pathway (343).
When introducing the contribution of the lncRNA
TUG1 to lung cancer, many independent studies highlighted the
downregulation of lncRNA TUG1 in the tissues derived from NSCLC
patients compared to control samples. Interestingly, patients with
a 2-year follow-up from lung cancer presented a marked decline of
lncRNA TUG1 depending on the tumor-node-metastasis (TNM) staging,
tumor size, and patient progression (344). The lower TUG1 expression was
related to increased tumor burden and the poorer overall survival
of patients, implying that lncRNA TUG1 can function as an
independent predictor for overall survival of NSCLC patients. The
expression pattern of lncRNA TUG1 was also confirmed in an
independent cohort study of patients, indicating the CUGBP
Elav-Like Family Member 1 (CELF1) as a possible target of lncRNA
TUG1 (345). Interestingly, it
was proved that the underlying molecular mechanisms of lncRNA TUG1
were based on inhibiting epigenetically homeobox B7 (HOXB7)
expression, through interference with either AKT or MAPK signaling
cascade (314). These data
suggested that TUG1 is regarded as a potential tumor-suppressive
lncRNA in NSCLC.
In gastric cancer, lncRNA TUG1 seemed to be
overexpressed, in turn leading to metastasis of gastric cancer into
lymph nodes (327). TCGA
validated the overexpression of lncRNA TUG1, which predicts for
poor prognosis of gastric cancer (346). In particular, the lncRNA TUG1 was
regarded as an unfavorable predictor of gastric cancer, because
patients diagnosed with gastric cancer displayed such increased
expression patterns depending on the following clinicopathological
features: the invasion depth of tumor, the tumor site and the tumor
stage (327). Focusing on the
functional properties of lncRNA TUG1, it was proved that lncRNA
TUG1 was important in cell cycle regulation of gastric cancer
cells, through the interaction of lncRNA TUG1 with either PRC2
complex or cyclin-dependent protein kinase inhibitors such as p15,
p16, p21, p27, and p57 (320). Of
particular interest was the competitive inhibitory association of
TUG1 and miR-381, affecting SRY-Box Transcription Factor 4 (SOX-4)
levels and leading to enhanced migratory capacities of gastric
cancer cells (328). In another
case, it was illustrated that the negative relationship between the
lncRNA TUG1 and miR-145-5p accelerated gastric cancer proliferation
(347).
In hepatocellular carcinoma, high lncRNA TUG1
expression was tightly related to cancer progression, suggesting
the diagnostic importance of the lncRNA TUG1 (311). It was reported that lncRNA TUG1
was positively related to the alpha-fetoprotein (AFP) gene.
Considering that AFP gene had high prognostic significance in
non-hepatitis B/non-hepatitis C HCC (NBNC-HCC), it was plausible to
suggest that the lncRNA TUG1 could be effectively used as an
unfavorable prognostic marker in patients with non-hepatitis
B/non-hepatitis C HCC (NBNC-HCC) (348). At the molecular level, the
tumor-promoting role of lncRNA TUG1 was observed to orchestrate
hepatocellular carcinoma environment, through the sequestration of
distinct miRNAs. Following lncRNA TUG1 silencing, TUG1 inhibited
its interactions with various miRNAs, causing their upregulation
and the subsequent cancer remission (349). In that direction, it was
supported that lncRNA TUG1 was a competitive inhibitor of
miR-216b-5p, through the assembly of distal-less homeobox 2 (DLX2),
thus leading to exacerbation of hepatocellular carcinoma (HCC)
(349), given that overexpression
of DLX2 is present in HCC patients with poor prognosis (350). In another study, it was suggested
that lncRNA TUG1 was a competitive inhibitor of miR-132-3p and its
target (Sox4), as shown in HepG2, Huh7, HccLM3 cells (351). Additional in vitro
experiments proved that lncRNA TUG1 could abrogate the expression
levels of miR-144, through activation of Janus kinase/signal
transducer and activator of transcription 3 (JAK/STAT3), thereby
contributing to tumor progression (352). Alternatively, the lncRNA TUG1
could reduce miR-142-3p expression levels, thereby positively
contributing to zinc finger e-box binding homeobox 1 (ZEB1)-induced
epithelial-mesenchymal transition and metastasis in hepatocarcinoma
cells (Huh7 and HepG2) (353).
Similarly, it was substantiated that there was an upregulation of
TUG1 according to the Barcelona Clinic Liver Cancer (BCLC) stage
and tumour size, so that liver tumor carcinogenesis was accelerated
through overexpression of lncRNA TUG1 (311). At transcriptional setting, the
nuclear transcription factor SP1 induced the mRNA expression of
lncRNA TUG1 and in parallel the lncRNA TUG1 functioned as an
inhibitor of PRC2 complex, preventing the binding of PRC2 complex
to the promoter of Kruppel-like factor 2 (KLF2) (311).
Apart from the sponging action of lncRNA TUG1
against miRNAs, it was proved that that lncRNA TUG1 was involved in
the regulation of host immune responses during hepatocarcinoma
progression. It was highlighted that there was a positive
relationship between lncRNA TUG1 and C-X-C chemokine receptor type
4 (CXCR4) protein, accounting for increased infiltration of immune
cells such as lymphocytes, neutrophils, and monocytes, through
activation of many downstream signaling pathways in the hepatic
microenvironment of cancer patients. It was evidenced that elevated
CXCR4 expression not only contributed to increased trafficking of
immune cells but also was associated with the augmented function of
immune cells in tissues isolated from patients with
hepatocarcinoma. In particular, the lncRNA TUG1 appeared to
increase specific markers in immune populations such as COX-2 in
macrophages, C-C chemokine receptor type 7 (CCR7) in neutrophils
and CD1c, neuropilin 1 (NRP1), and CD11c in dendritic cells through
its positive association with CXCR4 protein (354). Opposite results were observed in
catenin β1 (CTNNB1)-mutated hepatoblastoma (HB) cells, following
lncRNA TUG1 elimination. Behind the molecular mechanism underlying
the tight link of lncRNA TUG1 and CXCR4 protein, it was proved that
the sponging action of lncRNA TUG1 against miR-335-5p accounted for
the acceleration of malignant progression of CTNNB1-mutated HB
cells, through increased infiltration of pro-tumor immunocytes
(354).
Furthermore, a considerable advance was made in the
functional significance of lncRNA TUG1 in pancreatic development.
Yin et al (302) observed
that relatively low levels of TUG1 modulated apoptosis and insulin
secretion in pancreatic β cells in vitro and in vivo,
implying its participation in diabetes pathogenesis. Afterward,
researchers investigated the function of TUG1 in malignant
transformation of pancreatic cells given that pancreatic cancer
(PC) is known as the 'king of cancers' due to the shortage of early
diagnostic biomarkers and effective therapeutic methods in advanced
stages (355). In pancreatic
cancer patients, the expression levels of lncRNA TUG1 appeared to
be increased according to the clinical pathologic characteristics.
Interestingly, patients with advanced stages (3/4) of pancreatic
cancer presented the most important increase of lncRNA TUG1, which
correlated with their poor prognosis (356,357). There was a close association of
TUG1 with the advanced stage of PC patients and lymphatic
metastasis (357), suggesting its
significance in the regulatory network that determined the
pancreatic development. In another study, overexpression of TUG1
appeared to confer increased gemcitabine chemoresistance in
pancreatic ductal adenocarcinoma (PDAC) patients (358), taking into consideration that
gemcitabine (2',2'-difluorodeoxycitidine) is the first-line
chemotherapy for PDAC patients (359). It was proved that gemcitabine
combined with SCH772984 (an inhibitor of the ERK pathway) could
counteract the drug resistance driven by the overexpression of
lncRNA TUG1, thereby increasing gemcitabine therapeutic efficacy
(358).
Additional research findings confirmed that lncRNA
TUG1 expression levels culminated in all pancreatic cell lines
in vitro, exacerbating tumor progression (356). Some studies proved that sponging
action of lncRNA TUG1 against distinct miRNAs accounted for
dramatically increased tumor growth of pancreatic cells,
facilitating the acquisition of mesenchymal characteristics in
tumor cells. For example, lncRNA TUG1 was enriched in three
following pancreatic cell lines: SW1990, BxPC3, and PaTu8988 but
its levels were discriminated among cell lines, indicating that
lncRNA TUG1 exerted its oncogenic role in cell type-dependent
manner (343). The underlying
molecular mechanism of lncRNA TUG1 relied on inhibiting miR-382,
thus increasing the recruitment of EZH2 methyltransferase to genes
that were implicated in epithelial to mesenchymal transition (EMT)
(343).
In another case, the migratory potential of
pancreatic cells was validated through the association between
lncRNA TUG1 with miR-29c (360).
In tissues derived from pancreatic cancer patients, the lncRNA TUG1
appeared to be upregulated, thereby causing the reduced expression
levels of tumor suppressor miR-29c and exacerbating pancreatic
cancer progression (360).
Following the elimination of lncRNA TUG1, the expression levels of
miR-29c downstream targets including integrin subunit beta 1
(ITGB1), MMP-2, and MMP-9 were increased, thereby contributing to
pancreatic cancer remission (360). In addition, it was shown that the
silencing of lncRNA TUG1 prevented pancreatic cancer growth,
through inhibition of the Notch1 pathway and upregulation of
miR-299-3p. The positive relationship of lncRNA TUG1 with the
Notch1 pathway was especially important, given that abnormal
activation of the Notch1 pathway accounts for the exacerbation of
pancreatic carcinogenesis (361,362). In that regard, the
TUG1/miR-299-3p/Notch1 pathway was considered a promising
therapeutic approach for pancreatic carcinogenesis. Besides, the
lncRNA TUG1 was reported to act as competing endogenous RNA
(ceRNAs) to inhibit miR-299-3p, taking into consideration that
miR-299-3p has been detected at low levels in thyroid cancer (TC)
(363), hepatocellular carcinoma
(HCC) (364), and colon cancer
(365).
Besides, the oncogenic role of lncRNA TUG1 has been
evidenced not only through its potential to promote the acquisition
of mesenchymal characteristics by cancer cells, but also through
its capacity to interfere with the function of tumor suppressor
genes. For example, lncRNA TUG1 was illustrated to be recruited at
the promoters of Rho family GTPase 3 (RND3) and metallothionein 2A
(MT2A) genes, inhibiting the transcriptional expression of either
gene, through the assembly of EZH2 in tissues originated from
pancreatic cancer patients (366). The pancreatic cancer growth was
worsened through the link between the lncRNA TUG1 and RND3 protein,
considering that the RND3 gene is a target of tumor suppressor p53
(367). Conversely, the prolif
eration of pancreatic cancer cells was prevented by the
upregulation of RND3 and the concomitant elimination of lncRNA TUG1
(366). In other words, lncRNA
TUG1 has an inverse relationship with the RND3 gene, through the
recruitment of EZH2 methyltransferase, which functioned as a
scaffold protein, as shown by experiments that were performed in
tissues originated from pancreatic cancer patients (366). As a result, the poor prognostic
value of lncRNA TUG1 (356) and
the tumor-promoting effect of lncRNA TUG1 on cancer invasion
(360) raised the possibilities
to use lncRNA TUG1 as a novel therapeutic target for combating
pancreatic cancer. In that regard, lncRNA TUG1 was presented as a
novel potential therapeutic approach, which was extremely helpful
in ameliorating pancreatic cancer progression.
The tumor-promoting role of lncRNA TUG1 was proved
not only in pancreatic cancer but also it was expanded to other
cancer cell types through its interaction with the RND3 gene. In
esophageal squamous cell carcinoma (ESCC), the silencing of RND3
was reported to stimulate cell proliferation and cell cycle
progression. Conversely, the upregulation of RND3 reversed the
phenotype of cancer cells, by preventing cell proliferation and
leading to cell cycle arrest at the G0/G1 phase. The aforementioned
phenotype of cancer cells was ascribed to the upregulation of RND3
targets including phosphatase and tensin homolog (PTEN) and
cyclin-dependent kinase inhibitor 1B (CDKN1B/p27kip1),
in combination with downregulation of the signaling molecules such
as phosphorylated Akt (pAKT) and cell cycle protein D1 (CCND1)]
(134). In glioblastoma, RND3
appeared to be the determinant factor that causes inhibition of
CCND1 expres sion, accompanied by the activation of
RB1/retinoblastoma, thereby leading to cancer remission (368).
In colon cancer, it was substantiated that lncRNA
TUG1 gathered considerable attention as an oncogene. The
overexpression of lncRNA TUG1 was observed in tissues isolated from
88 patients with CRC to a high extent (64.77%, 57 of 88) compared
to that of healthy subjects. To assimilate conditions in a clinical
setting, it was revealed that LoVo and SW480 cells presented very
high lncRNA TUG1 expression, so a series of functional analysis was
focused on those cells (369).
Following the silencing of lncRNA TUG1, the growth inhibition of
LOVO and SW480 cells was observed and it was attributed to the
induction of cell apoptosis (369). In particular, elimination of
lncRNA TUG1 significantly induced G0/G1 arrest in CRC cells (LOVO
and SW480), disabling them to metastasize (369). When elucidating the importance of
the strong tumor-promoting role of lncRNA TUG1, it was evidenced
that lncRNA TUG1 caused the constitutive expression of
Wnt/β-catenin signaling pathway in colon cancer (342). The research group of Jiang
(312) also explained that lncRNA
TUG1 was the determinant factor of colon cancer proliferation and
migration, through its inverse association with tumor suppressor
p63. Another research group considered that the upregulation of
lncRNA TUG1 had strong potential to boost the migration of colon
cancer through epithelial-mesenchymal transition (EMT) (369), consistent with the classical
tumor-promoting role of lncRNA TUG1 on cancer cell invasion and
radioresistance via EMT (330,332). Indeed, TUG1 silencing reduced
migration, invasion, the acquisition of mesenchymal characteristics
by CRC cells in vitro, and in parallel TUG1 depletion
inhibited lung metastasis in vivo (370). In particular, the lncRNA TUG1
targeted its downstream target, the twist-related protein 1
(TWIST1), causing the metastasis of CRC cells to be attenuated via
TWIST1 knockdown, independently of TGF-β signaling (370). The silencing of lncRNA TUG1 could
alleviate all the invasive properties of CRC cells, through
inhibition of TGF-β/TUG1/TWIST1 signaling cascade (370). Accordingly, Sun et al
pointed out that enforced expression of lncRNA TUG1 could be of
utmost importance in potentiating invasiveness of colon cancer
cells, as TUG1-overexpression SW480 CRC cells formed more
metastatic nodules after injection into the spleens of nude mice
(371). Following TUG1
upregulation, reduced expression of the epithelial marker
E-cadherin and increased expression of mesenchymal markers
(N-cadherin, vimentin, and fibronectin) were detected in colon
cancer cells (371). Taken
together, the lncRNA TUG1 might serve as a prognostic biomarker and
a therapeutic target (371).
With regard to breast cancer, the role of lncRNA
TUG1 is presented as a promising therapeutic target according to
tumor staging. Results compiled from TCGA database showed that
cancer progression of the patients was markedly improved when
lncRNA TUG1 expression levels were increased (341). Tissues originated from 20
triple-negative breast cancer (TNBC), and multiple TNBC cell lines
presented low expression levels of lncRNA TUG1, suggesting the
tumor-suppressor role of lncRNA TUG1 (341). Interestingly, the therapeutic
efficacy of cisplatin was increased through the overexpression of
lncRNA TUG1 (341). The
underlying mechanism of action of lncRNA TUG1 was based on
inhibiting the Wnt signaling pathway through negative regulation of
miR-197/Nemo-like kinase (NLK) expression (341). The research group of Wang
(372) highlighted that the
lncRNA TUG1 overexpression was capable of alleviating breast
cancer, as demonstrated by experiments in several breast cancer
cell lines and in patient samples. In particular, the low lncRNA
TUG1 expression was tightly related to mutant p53 expression, as
evidenced by results in MDA-MB-231 cancer cells compared to those
derived from MCF7 breast cancer cells with wild-type p53 status. In
line with that, it was also shown that low lncRNA TUG1 expression
was correlated with lymph node metastasis via modulating cell cycle
regulators (cyclinD1/CDK4), which were capable of exerting their
action as oncogenes in breast cancer (373). On the contrary, Ren et al
(347) presented that the lncRNA
TUG1 functioned as an oncogene, given that TUG1 silencing abrogated
breast cancer proliferation, as demonstrated in breast cancer cell
lines (MDA-MB-231 and MDA-MB-436). Zhao and Ren (374) delineated the molecular mechanism
underlying the tumor-promoting role of lncRNA TUG1, showing that
the sponging action of lncRNA TUG1 against miR-9 expression
increased the proliferation of p53 wild type breast cancer cells
(such as MCF7 cells). In support of the above, RNA sequencing data
derived from TCGA database proved that lncRNA TUG1 was enriched in
a great proportion of patients bearing HER2-positive and basal-like
subtypes of breast cancer compared to matched controls.
Consequently, lncRNA TUG1 might exert a significant prognostic and
therapeutic value, monitoring patient responses in pancreatic
cancer (375).
Besides, the tumor suppressor role of lncRNA TUG1
was also highlighted in glioma. Most of the studies showed that
lncRNA TUG1 was generally downregulated in glioma tissues compared
to matched normal tissues (313).
When the low lncRNA TUG1 expression levels were increased, the
glioma progression was counteracted, confirming the tumor
suppressor role of lncRNA TUG1. The tumor-blocking action of the
lncRNA TUG1 relied on its capacity to sequester miR-26a, leading to
enrichment of phosphatase and tensin homolog (PTEN), thus causing
inducing mitochondrial apop-tosis in glioma cells (313). Even though most of the studies
illustrated the downregulation of lncRNA TUG1 in glioma patients,
other studies have substantiated the opposite trend. Katsushima
et al (338) proved that
the lncRNA TUG1 played a significant role in increasing the
self-renewal of glioma stem cells by sequestering miR-145 in the
cytoplasm and by inhibiting the expression of crucial
differentiation genes through the recruitment of PRC2 complex to
target genes. The inhibitory action of TUG1 against miR-145, was
responsible for the upregulation of SOX2 and c-Myc expression,
thereby promoting self-renewal in glioma stem cells (338). In essence, Notch signaling caused
lncRNA TUG1 overexpression in glioma stem cells, thereby increasing
the recruitment of PCR2 complex to neuronal
differentiation-associated genes and causing their epigenetic
silencing (338). In another
study, it was shown that lncRNA TUG1 exerted pro-tumorigenic
action, not only by triggering glioma progression but also by
playing a crucial role in metastasis. The lncRNA TUG1 was
considered as an important regulator of tumor angiogenesis, by
elevating VEGF expression and potentiating tumor growth via
augmenting tumor microvessel density (340). In more depth, the mechanism
underlying the angiogenesis-stimulatory action of lncRNA TUG1 was
based on inhibiting miR-299 in glioblastoma cells (340).
Since the blood-tumor barrier inhibits the delivery
of chemotherapeutic drugs to brain tumor tissue (376), the possibility of TUG1 to
increase the movement of chemotherapeutic drugs in brain tissues
was especially important. Cai et al (333) highlighted that inhibition of TUG1
enabled chemotherapeutic drugs to be permeabilized through
blood-tumor, to deal with glioma progression. In particular,
silencing of lncRNA TUG1 increased blood-tumor barrier
permeability, through reducing the expression of three junction
proteins, namely occludin, tight junction protein-1 (ZO-1) and
claudin-5 (333). Elucidating the
underlying molecular mechanism of TUG1, it was proved that TUG1
exerted its inhibitory action against miR-144, thereby targeting
heat shock transcription factor 2 (HSF2) (333).
Apart from the importance of lncRNA TUG1 in
multiple cancer types, its significance was evidenced in both sex-
dependent cancer types including ovarian and prostate. TCGA
database supported that the lncRNA TUG1 was remarkably upregulated
in tissues of epithelial ovarian cancer patients compared to paired
adjacent tissues. The overexpression of lncRNA TUG1 was positively
correlated with pathological grade, tumor size, supporting its high
prognostic value (377,378). As a general note, the most known
interacting partners of the lncRNA TUG1 were identified as the
miRNAs: miR-29c, miR-142, and miR-145 (332,379,380) exacerbating the unrestrained
growth of bladder cancer cells. For example, the overexpression of
TUG1 was reported to inhibit the miR-29c expression, accounting for
the uncontrolled, and the migration of bladder cancer cells (T24
and EJ) (379). It is important
to be mentioned that the oncogenic role of lncRNA TUG1 was
consistent with that in pancreatic cancer since the lncRNA TUG1
functioned as a tumor promoter in the pancreas through its
association with miR-29c (360).
Apart from the inhibitory effect of lncRNA TUG1 on miR-29c, TUG1
appeared to upregulate zinc finger e-box binding homeobox 2 (ZEB2)
transcription factor, through its competitive interaction with
miR-142, thereby enabling the increased cell proliferation and
migration of bladder cancer cells (380). Likewise, lncRNA TUG1 was shown to
function as a potent inhibitor of miR-145 expression, facilitating
bladder cancer cell metastasis by increasing the recruitment of
ZEB2 transcription factor to target EMT genes, thereby providing
new insights into the regulation of radioresistance mediated by
lncRNA TUG1 (332). The
significance of lncRNA TUG1 was also proved through its capacity to
amplify the radiosensitivity of bladder cancer through inhibition
of high-mobility group protein 1 (HMGB1) expression (381). As a result, the lncRNA TUG1 was
proposed as a promising therapeutic target and prognostic marker in
bladder cancer.
In that direction, it was shown that overexpression
of TUG1 was the most determinant factor for activating the
Wnt/β-catenin signaling pathway (380) through negative regulation of
miR-138-5p and subsequent upregulation of Sirtuin 1
(Sirt1)-nicotinamide adenosine dinucleotide (NAD)-dependent
deacetylase in CC cells (382).
In more depth, the increased activity of SIRT1 protein appeared to
be pronounced in inhibiting the expression of epithelial markers
such as E-cadherin, in turn exerting a positive effect on the
Wnt/β-catenin signaling pathway, through a positive feedback loop
(382). In line with the above,
additional experiments showed that lncRNA TUG1 caused the increased
invasion of human umbilical vein endothelial cells (HUVECs), by
upregulating leucine-rich alpha-2-glycoprotein 1 (LRG1) secretion
through transforming growth factor-beta (TGF-β) pathway (383), confirming its oncogenic role. In
particular, both SKOV3 and CAOV3 endothelial cell lines presented
strong pro-angiogenic effects via Smad1/5/8 signaling pathways,
thereby leading to binding of LRG1 protein to TGF-β accessory
receptor (384). In this way,
tumor-related angio-genesis was reduced after LRG1 elimination, as
manifested by the downregulation VEGF-a, angiopoietin-1 (Ang-1),
thereby reducing the signal transmission through TGF-β pathway
(385). Likewise, angiogenesis
was demonstrated to be increased in cerebral ischemia, through the
positive effect of LRG1 on the TGF-β1 pathway (386). If one considered that there was a
positive association of LRG1 and VEGF-α, it was plausible that
lncRNA TUG1 contributed to increased angio-genesis through LRG1
upregulation (384,385). In parallel, lncRNA TUG1 increased
hypoxia-inducible factor-1α (HIF-1α) expression, though LRG1
upregulation, by accelerating tumor angiogenesis (387). Besides, it was proved that
extracellular taurine triggered angiogenesis, through activation of
Akt, extracellular- signal-regulated kinase (ERK), and
steroid-receptor-coactivator/focal adhesion kinase (Src/FAK)
signaling cascades in vitro and in vivo (388). The cell cycle progression of
endothelial cells was regulated by Akt- and ERK-dependent cell
signaling pathways and the cell migration of endothelial cells was
orchestrated in an Src-dependent manner, without stimulating
inflammation and permeability in vitro and in vivo
(388).
Hence, the lncRNA TUG1 was identified to predict
poor prognosis of prostate cancer patients (389). Many researchers have provided
deep insight into the oncogenic role of TUG1, supporting that the
function of lncRNA TUG1 was mediated through its capacity to hinder
the expression levels of related miRNAs: miR-145-5p, miR-144, and
miR-381 (327,328,347). The molecular mechanism of lncRNA
TUG1 relied on triggering cancer cell proliferation through its
effects on the cell cycle of prostate cancer cells (320,346). Importantly, the lncRNA TUG1 was
of critical importance for prostate cancer progression in
vitro and in vivo, exerting its action through
miR-128-3p/YES1 axis (390). The
lncRNA TUG1 elicited its potential oncogenic role in prostate
cancer cells, by inhibiting miR-128-3p and its target YES1
(390).
Similarly, abnormal expression of TUG1 was observed
to predict poor prognosis of ESCC patients, serving as a potential
oncogene (391). In a cohort of
62 patients, the lncRNA TUG1 was significantly overexpressed in
ESCC tissues compared with paired adjacent normal tissues, and the
high expression level of TUG1 was related to family history and
upper segment of esophageal cancer (392). By loss of function experiments,
it was substantiated that the silencing of TUG1 limited the
proliferation and migration of ESCC cells and arrested the cell
cycle progression (392). Behind
its molecular targets, it was observed that lncRNA TUG1 potentiated
the EMT progression of ESSC cells, though its preventive action on
miR-148a-3p (393). The silencing
of lncRNA TUG1 was sufficient to reverse all the manifestations of
EMT progression, due to its inverse correlation with miR-148a-3p,
as shown by experiments in ESCC (EC9706 and OE19) cells (393). The significant regulatory
function of TUG1 against the migration of ESCC cells was supported
by the fact that expression of EMT-associated proteins (C-myc,
Cyclin D1, and catenin-beta 1/β-catenin) was under the control of
antagonistic interaction of TUG1 and miR-148a-3p (393).
In osteosarcoma, the overexpression of lncRNA TUG1
was observed according to tumor size, distant metastasis, TNM
staging, and overall and recurrence-free survival of patients,
suggesting that lncRNA exhibited strong prognostic value.
Interestingly, the lncRNA TUG1 was considered a superior biomarker
to alkaline phosphatase (ALP) in distinguishing osteosarcoma
patient cases from healthy controls (394). Focusing on its molecular
mechanisms, the lncRNA TUG1 functioned as an oncogene in
osteosarcoma through its capacity to bind to various miRNAs
(395). To begin with, the fact
that overexpression of lncRNA TUG1 in osteosarcoma cells was
accompanied by the transcriptional inhibition of miR-212-3p, which
in turn caused the relative downregulation of forkhead box A1
(FOXA1) which was a transcriptional target of miR-212-3p (396). The negative regulation of
miR-212-3p also exerted a great impact on affecting the expression
levels of sex-determining region Y box 4 (SOX4) (397), causing the overexpression of SOX4
which comprises an oncogene in various malignancies, including
osteosarcoma (398). The latest
research findings supported that the lncRNA TUG1 functioned as an
endogenous sponge, downregulating either miR-335-5p (399) or miR-9-5p (334) or miR-219a-5p (400) or miR-132-3p (397), thus promoting migration of
osteosarcoma cells. Similarly, the lncRNA TUG1 inhibited
miR-144-3p, causing nuclear translocation of β-catenin in
osteosarcoma cells (MG63, U2OS, HOS, and Saos-2), thus exacerbating
tumor progression (401). In
another study, the lncRNA TUG1 functioned as an oncogene,
competitively inhibiting miR-219a-5p (400). The attenuating action of lncRNA
TUG1 against miR-219a-5p seemed to potentiate osteosarcoma
progression, through the activation of either Wnt/β-catenin
signaling pathway or of Akt signal transduction pathway (400). In addition, it was proved that
lncRNA TUG1 acted as an endogenous sponge to directly bind to
miR-9-5p, inhibiting its expression. The lncRNA TUG1 overturned the
effect of miR-9-5p on the proliferation, colony formation, cell
cycle arrest, and apoptosis in osteosarcoma cells, which involved
in the activation of POU class 2 homeobox 1 (POU2F1) expression. As
a result, a novel TUG1/miR-9-5p/POU2F1 pathway was revealed, in
which lncRNA TUG1 acted as a competitive endogenous RNA by sponging
miR-9-5p, leading to down-regulation of POU2F1, thereby worsening
the progression of osteosarcoma. These findings corroborate the
importance of lncRNA-targeted therapy against human osteosarcoma
(137). The importance of lncNRA
TUG1 was also confirmed by its involvement in metastasis, given
that early osteosarcoma gives rise to robust progression and
aggressive metastasis. Indeed, Ma et al (394) supported that TUG1 expression was
also correlated to post-operative chemotherapy, tumour size and
Enneking surgical stage. As the TUG1 expression levels were
increased, lncRNA TUG1 was associated with poorer prognosis,
including shortened overall and progression-free survival,
independent of other clinicopathological parameters.
At the molecular setting, the lncRNA TUG1
expression was reported to be increased through the TGF-β pathway
elicited by cancer-associated fibroblasts (CAFs). The impaired TUG1
expression functioned as a miRNA 'sponge' to competitively confer
protection to the hypoxia inducible factor -1a (HIF-1a) mRNA 3'UTR
from the inhibitory action of miR-143-5p. It was proposed that the
lncRNA TUG1 might be a prognostic indicator for osteosarcoma and
could be a therapeutic target for osteosarcoma, through its potent
significance in determining osteosarcoma cell metastasis,
angiogenesis, and proliferation in vivo and in vitro
(402). As a result, the high
lncRNA TUG1 expression was regarded as a critical modulator in
orchestrating the metastasis of osteosarcoma cells.
In acute myeloid leukemia (AML), patients were
diagnosed with poor prognosis, when TUG1 expression levels were
high. The underlying mechanism of lncRNA TUG1 relied on interfering
with miR-34a and recruiting EZH2 methyltransferase, thus rendering
HL60/ADR cells insensitive to adriamycin (ADR) (403). As a result, the high expression
pattern of lncRNA TUG1 provided a clue about the diagnostic
importance of lncRNA TUG1 in ADR resistant cells of AML.
Accordingly, it was reported that multiple myeloma patients
presented showing significant downregulation of lncRNA TUG1 as
opposed to healthy subjects (404).
Cancer progression is affected by aberrant energy
metabolism, which can be elicited through the action of lncRNAs
(405). It has been reported that
cancer progression can be aggravated through abnormal high
glycolysis and enhanced glutamine metabolism (406). Taking into consideration that
lncRNA TUG1 was an important regulator of mitochondrial
bioenergetics (301), further
investigations revealed that lncRNA TUG1 was overexpressed in
clinical specimens isolated from cholangiocarcinoma (ICC) and its
upregulation was dependent on tumor stage (407). Also, its overexpression was
considered as an independent prognostic factor for patients with
poor outcome (407). Other
researchers suggested that lncRNA TUG1 functioned as a competitive
endogenous RNA against miR-145, causing metabolic reprogramming of
ICC cells (408). In particular,
it was proved that the overexpression of TUG1 was capable of
elevating glutamine consumption, α-Ketoglutaric acid (a-KG)
production, and ATP levels through miR-145 inhibition and
subsequent Sirt3/GDH elevation (408). As a result, the lncRNA TUG1 might
be a useful prognostic biomarker in ICC patients and a potentially
important therapeutic target to orchestrate metabolic reprogramming
in ICC, thereby recovering the glutamine metabolism in cancer
cells. Accordingly, the abnormal overexpression of lncRNA TUG1
appeared to play a crucial role in the progression of osteosar-coma
cells, through metabolic alterations (408). The lncRNA TUG1 seemed to affect
the osteosarcoma progression through its ability to augment glucose
consumption, and lactate production owing to the upregulation of
hexokinase-2 (409).
Taken together, many oncologists have embraced the
search of molecular mechanisms underlying the therapeutic index of
lncRNA TUG1 in a wide spectrum of cancer types, to overcome the
challenges such as toxicity, drug resistance, tumor heterogeneity
encountered by gold standard treatment decisions. The molecular
characterization of TUG1 targets enables physicians to define the
genomic changes in each tumor type, thereby contributing to the
design of selected tumor-targeted therapy based on the detailed
portrait of tumor types. It seems that lncRNA TUG1 signature is
heading toward the mainstream in precision medicine given that
lncRNAs are characterized by high sensitivity and specificity
against their targets (410).
Nonetheless, more research is urgently needed to elucidate the
crosstalk between lncRNA TUG1 and endogenous molecules, to render
the lncRNA TUG1 as an appealing therapeutic approach against
cancer.
7. The association of lncRNA TUG1 and
chemoresistance
During metastasis of cancer cells, the major
challenges are related to the biologic heterogeneity of tumor cells
and to the tumor microenvironment, which accounts for the
drug-resistant phenotype in cancer cells (411-413). Besides, it is important to be
noted that the ineffectiveness of multiple chemotherapeutic drugs
is attributed to the phenomenon of multidrug resistance (MDR). In
this MDR procedure, cancer cells acquire multiple aggressive
characteristics, enabling them resistant to the cytostatic or
cytotoxic action of potential drugs. Among them, limited drug
penetrance; enhanced drug efflux; affected membrane lipids (e.g.,
ceramides) (414); increased drug
metabolism; altered drug targets; detoxification by
compartmentalization; blocked programmed cell death (apoptosis);
induction of mechanisms that repair DNA damage (415); alterations in the cell cycle and
checkpoints have been reported as the most significant barriers
during MDR that should be addressed (416). For example, the expression of MDR
protein 1 (MDR1) and MDR-associated protein 1 (MRP1) expression can
be reduced in DOX or cisplatin-resistant BUC cells, through
Wnt/β-catenin pathway (417). In
this context, the upregulation of the Wnt/β-catenin pathway has
shown to reverse the effects of lncRNA TUG1 knockdown on Dox
resistance in T24/DR cells (418).
To overcome resistance liabilities, research has
focused on examining the functional role of lncRNA TUG1. The
pleiotropic nature of lncRNA has enabled it to be uniquely
tailored, with the ultimate aim of overcoming cancer cell drug
resistance originating from a redundancy of oncogenic signaling in
a wide spectrum of cancer types. In particular, the lncRNA TUG1 has
appeared to play a complicated role in cancer progression, exerting
either beneficial or detrimental effects on resistant cancer cells.
On one side, the lncRNA TUG1 has been extensively analyzed for its
capacity to confer resistance in various types of cancer cells in
response to classical therapeutic actions. On the other side, the
lncRNA TUG1 has been reported as a protective agent that enables
cancer cells to overcome resistance conferred by various
chemotherapeutic drugs.
In this direction, it has been reported that
overexpression of lncRNA TUG1 in esophageal squamous carcinoma
cells confers resistance to platinum combined with 5-FU or PTX
(391). For example, elevated
levels of lncRNA TUG1 appear in bladder urothelial cancer (BUC)
patients, indicating the poor response of patients to DOX
chemotherapy (418). Activation
of the Wnt/β-catenin pathway has been shown to contribute to DOX
resistance in BUC patients, as demonstrated by TCGA Pan-Cancer
(PANCAN) (418). Also, lncRNA
TUG1 has been reported to be overexpressed in colon cancer
patients, given that lncRNA TUG1 silencing is regarded as an
effective way to overcome MTX resistance conferred by colorectal
cancer cells. TUG1/miR-186/CPEB2 is the main signaling cascade that
determines the reduced sensitivity of CRC cells to MTX (419).
It has also been proven that lncRNA TUG1
significantly contributes to ADR resistance in urothelial carcinoma
of the bladder (UCB) growth, thanks to its positive relationship
with master transcription factor of anti-oxidant response
NF-erythroid 2 (NF-E2)-related factor 2 (Nrf2) (403). Deficiency of either transcription
factor Nrf2 or lncRNA TUG1 enables UCB cells to overcome ADR-based
chemotherapy resistance (403).
The overexpression of TUG1 also plays an important role in inducing
PTX resistance in both SK-OV-3 and A2780/R ovarian cancer cells,
exerting its effect on increasing autophagy. In particular, lncRNA
TUG1 promotes autophagy, through its inhibitory effect on
miR-29b-3p, resulting in conferring PTX resistance to both SK-OV-3
and A2780/R ovarian cancer cells (420). The PTX resistance of ovarian
cancer cells can be overcome through the silencing of TUG1
(420). The potential of lncRNA
TUG1 to increase autophagy in ovarian cancer cells is considered
one of the important mechanisms by which the lncRNA TUG1
inactivates PTX therapeutic effect, thereby enabling ovarian cancer
cells resistant to PTX (421).
In addition to the above, CC tissues have presented
increased expression levels of TUG1 and the expression levels of
lncRNA TUG1 are tightly associated with cisplatin
(cis-Dichlorodiammineplatinum, DDP). The silencing of lncRNA TUG1
seems to hinder the proliferative rate of CC cells, through the
increased signal transmission of the MAPK pathway (p38 MAPK, JNK),
thereby accelerating the apo ptosis of CC cells (422). Following MAPK activation, the
expression levels of the RFX7 gene (transcriptional target of TUG1)
are inhibited, due to TUG1 silencing in CC cells. The contribution
of lncRNA TUG1 is of utmost importance because the majority of
patients commonly develop cisplatin resistance (423). Besides, it is important to
highlight that overexpression of TUG1 has a high prognostic value
in CC patients (423).
Similarly, ADR-resistant AML tissues and cells have
presented increased sensitivity to ADR response, through TUG1
silencing. It has been documented that there is a competitive
indirect interaction between lncRNA TUG1 and miR-34a, through the
action of one component of Polycomb complex (EZH2), which catalyzes
the epigenetic regulation of miR-34a. The increased sensitivity of
AML cells to ADR is accompanied by the silencing of lncRNA TUG1 as
well as the upregulation of miR-34a (403).
Consistent with the above, lncRNA TUG1 has been
reported to confer resistance to small cell lung cancer (SCLC)
cells through modulation of the expression of LIMK2b (a splice
variant of LIM-kinase 2) via binding with EZH2 (424). Similarly, Xu et al
observed that lncRNA TUG1 elimination facilitated the cisplatin
sensitivity of cisplatin-resistant ESCC (ECA109 or EC9706) cells
(425). The upregulation of TUG1
appeared to exacerbate cancer progression given that TUG1 lncRNA is
an epigenetic inhibitor of PDCD4 expression through recruitment of
EZH2 (426). In the same context,
the up-regulation of TUG1 was also identified to drive increased
migration of pancreatic ductal adenocarcinoma cells, thereby
reducing the gemcitabine chemosensitivity (358).
Last but not least, Liu et al (427) examined the expression levels of
various lncRNAs including TUG1 in glioma cell lines U87 and U251
upon treatment with resveratrol and DOX (427). It was demonstrated that the
expression pattern of lncRNA TUG1 was downregulated upon necrosis
induction in both cell lines but it was remained unchanged during
DNA damage-induced apoptosis.
8. The role of taurine or lncRNA TUG1 as a
prognostic marker
Towards evaluating new therapeutic agents, multiple
studies are underway identifying specific markers, that might
provide indicative clues for the increased susceptibility of cancer
patients to specific drugs and the design of a suitable
personalized medicine. Some criteria such as reliability,
reproducibility, noninvasiveness, simplicity, and cost-efficiency
are required to identify the ideal biomarker. In samples of
physiologic fluids, taurine can be easily isolated to be used for
diagnostic scopes. Taurine or lncRNA TUG1 can provide us with
specific information concerning an individual's nutrition or
disease status or medications when taurine or lncRNA TUG1 is
harvested from physiological fluids. As high-technology platforms
become available, the major question remains whether taurine or
lncRNA TUG1 can be used as a good prognostic biomarker or
diagnostic indicator to guide care for cancer patients.
Accumulating evidence has supported that TUG1
lncRNA has a prognostic significance in cancer patients. The lncRNA
TUG1 protein expression levels have been associated with
clinicopathologic features of patients harboring various tumor
types, indicating that TUG1 lncRNA is a strong indicator of poor
outcome in cancer patients. Interestingly, results of meta-analysis
derived from nine cancer types, have proposed that there is an
inverse relationship between lncRNA TUG1 and overall survival in
patients with cancer. In this regard, TUG1 is linked to the
unfavorable overall survival of cancer patients given that TUG1
expression is increased depending on the severity of
clinicopathological symptoms of cancer patients and can be used as
a new reliable biomarker for cancer patients (428). These data suggest that lncRNA
TUG1 can be added as a potential biomarker to the growing list of
tests for the management of cancer patients. However, some
limitations should not be addressed in the meta-analysis due to the
involvement of 12 studies. To acquire more reliable data, more
large-scale and well-designed studies need to be included in the
analysis to circumvent the existed heterogeneity across 12 studies,
as demonstrated by inconsistent threshold values of TUG1 lncRNA
protein expression (428).
With regards to taurine, proton MRS technique has
shown that there is a differential expression in 29 metabolites
between mice bearing B16F10 melanoma cells and control matched
mice. Among the metabolites with differential expression between
tumor and normal samples, taurine is involved. As a result, taurine
can be leveraged for either monitoring tumor progression or
evaluating the pharmacological efficacy of different therapeutic
schemes (234). As a result, the
introduction of taurine or lncRNA TUG1 heralds its use as a
predictive biomarker, by which correct patient stratification can
be accomplished during cancer therapy. The correct tumor patient
sampling is mandatory for predictive biomarker discovery.
9. Conclusions
In the past decade, there has been a tremendous
advance in our knowledge on the molecular mechanisms of taurine or
its haloamines against inflammatory disorders and distinct cancer
types. To manipulate taurine or its haloamines for therapeutic
purposes, it is essential to elucidate how the complex cellular
interplay between stimulatory and inhibitory signals is regulated.
In this direction, the anti-inflammatory or anti-cancer effect
elicited by taurine or its haloamines has been evaluated, thus
delineating the exact molecular mechanism caused by either agent.
In addition, some studies have presented that taurine functions as
a favorable agent for cancer chemoprevention, used either alone or
in combination with other drugs, by maximizing the therapeutic
outcome of chemotherapeutic drugs without increasing cytotoxicity.
At present, the most alluring reason to recommend taurine
supplementation is a taurine deficiency. Advances in the
understanding of cell procedures regulated by taurine or its
haloamines are needed to provide significant input to ignite our
minds, ensuring a bright future regarding the potential therapeutic
use of taurine and its derivatives in cancer and inflammation.
LncRNA TUG1 is regarded as a potential therapeutic
target or prognostic marker, exerting its biological action at
least in part through chromatin remodeling and sequestration of
microRNAs. The lncRNA TUG1 is presented to be an eminent non-coding
RNA of utmost importance to modulate targets, either enriching or
reliably attenuating gene expression. Despite the great accrual of
research findings, there is a certain paucity pertinent to the
detailed downstream molecular mechanisms mediated by TUG1. Certain
research technologies, such as high-throughput identification of
binding partners and integrative analysis of omics data can be
employed to elucidate the functional role of lncRNA TUG1 in a cell
type-dependent manner. Further analysis will provide us with
information that will enable the prognostic significance of TUG1
and the systemic modulation of TUG1 as a therapeutic option against
distinct cancer types. To use the lncRNA TUG1 as a therapeutic
target, some barriers should be bypassed such as the increased
vulnerability of lncRNA to degradation and its low efficiency of
delivery.
Taurine or its haloamines or its lncRNA TUG1
deserve further research studies that will substantiate into their
functional properties, their specificity, their stability, their
toxicity, prompting researchers to optimize either agent for
implementation in the clinical setting.
Acknowledgments
Not applicable.
Funding
This study was supported by I.K.Y State Scholarship
Foundation for S. Baliou's Ph.D. studies. The IKY code is
2018-050-0502-13155.
Availability of data and materials
Not applicable.
Authors' contributions
All the authors were involved in the conception and
design of the study. SB performed the literature search, wrote the
manuscript, critically analyzing the existing knowledge and
designed the figures; SB, AMK, DAS and VZ also contributed to
editing the manuscript. All authors approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
References
|
1
|
Leon R, Wu H, Jin Y, Wei J, Buddhala C,
Prentice H and Wu JY: Protective function of taurine in
glutamate-induced apoptosis in cultured neurons. J Neurosci Res.
87:1185–1194. 2009. View Article : Google Scholar
|
|
2
|
Chang CY, Shen CY, Kang CK, Sher YP, Sheu
WHH, Chang CC and Lee TH: Taurine protects HK-2 cells from oxidized
LDL-induced cytotoxicity via the ROS-mediated mitochondrial and
p53-related apoptotic pathways. Toxicol Appl Pharmacol.
279:351–363. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schaffer S, Azuma J, Takahashi K and
Mozaffari M: Why is taurine cytoprotective? Adv Exp Med Biol.
526:307–321. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marcinkiewicz J and Kontny E: Taurine and
inflammatory diseases. Amino Acids. 46:7–20. 2014. View Article : Google Scholar :
|
|
5
|
Schuller-Levis GB and Park E: Taurine and
its chloramine: Modulators of immunity. Neurochem Res. 29:117–126.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maher SG, Condron CE, Bouchier-Hayes DJ
and Toomey DM: Taurine attenuates CD3/interleukin-2-induced T cell
apoptosis in an in vitro model of activation-induced cell death
(AICD). Clin Exp Immunol. 139:279–286. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fukuda K, Hirai Y, Yoshida H, Nakajima T
and Usui T: Free amino acid content of lymphocytes and granulocytes
compared. Clin Chem. 28:1758–1761. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Capuozzo E, Pecci L, Baseggio Conrado A
and Fontana M: Thiotaurine prevents apoptosis of human neutrophils:
A putative role in inflammation. Adv Exp Med Biol. 775:227–236.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Redmond HP, Stapleton PP, Neary P and
Bouchier-Hayes D: Immunonutrition: The role of taurine. Nutrition.
14:599–604. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
William R, Watson G, Redmond HP, Wang JH
and Bouchier-Hayes D: Mechanisms involved in sodium
arse-nite-induced apoptosis of human neutrophils. J Leukoc Biol.
60:625–632. 1996. View Article : Google Scholar
|
|
11
|
Condron CM, Toomey DM, Casey RG, Creagh T
and Bouchier-Hayes DJ: Taurine protects against PMN dysfunction and
death in urine. Urol Res. 32:338–345. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Masuda M, Horisaka K and Koeda T: Effects
of taurine on neutrophil function in hyperlipidemic rats. Jpn J
Pharmacol. 40:478–480. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schaffer SW, Azuma J and Mozaffari M: Role
of antioxidant activity of taurine in diabetes. Can J Physiol
Pharmacol. 87:91–99. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Zhao N, Zhang F, Yue W and Liang
M: Effect of taurine on leucocyte function. Eur J Pharmacol.
616:275–280. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pasantes-Morales H and Cruz C: Taurine and
hypotaurine inhibit light-induced lipid peroxidation and protect
rod outer segment structure. Brain Res. 330:154–157. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Son M, Kim HK, Kim WB, Yang J and Kim BK:
Protective effect of taurine on indomethacin-induced gastric
mucosal injury. Adv Exp Med Biol. 403:147–155. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marcinkiewicz J, Grabowska A, Bereta J and
Stelmaszynska T: Taurine chloramine, a product of activated
neutrophils, inhibits in vitro the generation of nitric oxide and
other macrophage inflammatory mediators. J Leukoc Biol. 58:667–674.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakajima Y, Osuka K, Seki Y, Gupta RC,
Hara M, Takayasu M and Wakabayashi T: Taurine reduces inflammatory
responses after spinal cord injury. J Neurotrauma. 27:403–410.
2010. View Article : Google Scholar
|
|
19
|
Zhang F, Mao Y, Qiao H, Jiang H, Zhao H,
Chen X, Tong L and Sun X: Protective effects of taurine against
endotoxin-induced acute liver injury after hepatic ischemia
reperfusion. Amino Acids. 38:237–245. 2010. View Article : Google Scholar
|
|
20
|
Elson CO, Sartor RB, Tennyson GS and
Riddell RH: Experimental models of inflammatory bowel disease.
Gastroenterology. 109:1344–1367. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abdih H, Kelly CJ, Bouchier-Hayes D, Barry
M and Kearns S: Taurine prevents interleukin-2-induced acute lung
injury in rats. Eur Surg Res. 32:347–352. 2000. View Article : Google Scholar
|
|
22
|
Sun M, Zhao Y, Gu Y and Xu C:
Anti-inflammatory mechanism of taurine against ischemic stroke is
related to down-regulation of PARP and NF-κB. Amino Acids.
42:1735–1747. 2012. View Article : Google Scholar
|
|
23
|
Son MW, Ko JI, Doh HM, Kim WB, Park TS,
Shim MJ and Kim BK: Protective effect of taurine on TNBS-induced
inflammatory bowel disease in rats. Arch Pharm Res. 21:531–536.
1998. View Article : Google Scholar
|
|
24
|
Shimizu M, Zhao Z, Ishimoto Y and Satsu H:
Dietary taurine attenuates dextran sulfate sodium (DSS)-induced
experimental colitis in mice. Adv Exp Med Biol. 643:265–271. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao Z, Satsu H, Fujisawa M, Hori M,
Ishimoto Y, Totsuka M, Nambu A, Kakuta S, Ozaki H and Shimizu M:
Attenuation by dietary taurine of dextran sulfate sodium-induced
colitis in mice and of THP-1-induced damage to intestinal Caco-2
cell monolayers. Amino Acids. 35:217–224. 2008. View Article : Google Scholar
|
|
26
|
Yin Y, Wen K, Wu Y, Kang Y and Lou J:
Inhibition of sodium current by taurine magnesium coordination
compound prevents cesium chloride-induced arrhythmias. Biol Trace
Elem Res. 146:192–198. 2012. View Article : Google Scholar
|
|
27
|
Wang Q, Fan W, Cai Y, Wu Q, Mo L, Huang Z
and Huang H: Protective effects of taurine in traumatic brain
injury via mitochondria and cerebral blood flow. Amino Acids.
48:2169–2177. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Megaraj V, Iida T, Jungsuwadee P, Hofmann
AF and Vore M: Hepatobiliary disposition of
3alpha,6alpha,7alpha,12alpha-tetra-hydroxy-cholanoyl taurine: A
substrate for multiple canalicular transporters. Drug Metab Dispos.
38:1723–1730. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guler L, Tavlasoglu M, Yucel O, Guler A,
Sahin MA, Kurkluoglu M, Sirin Y, Eken A, Gamsizkan M, Dakak M, et
al: Taurine attenuates lung ischemia-reperfusion injury after lung
transplantation in rats. J Anesth. 28:347–353. 2014. View Article : Google Scholar
|
|
30
|
Park SH, Lee H, Park KK, Kim HW and Park
T: Taurine-responsive genes related to signal transduction as
identified by cDNA micro-array analyses of HepG2 cells. J Med Food.
9:33–41. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Song XD, Chen CZ, Dong B, Shi YY, Zhang W,
Yan LS and Luo GA: Study on the intervening mechanism of taurine on
streptozotocin-induced diabetic cataracts. Zhonghua Yan Ke Za Zhi.
39:605–609. 2003.In Chinese.
|
|
32
|
Schuller-Levis G, Mehta PD, Rudelli R and
Sturman J: Immunologic consequences of taurine deficiency in cats.
J Leukoc Biol. 47:321–331. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sapronov NS, Khnychenko LK and
Polevshchikov AV: Effects of new taurine derivatives on primary
immune response in rats. Bull Exp Biol Med. 131:142–144. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Negoro S and Hara H: The effect of taurine
on the age-related decline of the immune response in mice: The
restorative effect on the T cell proliferative response to
costimulation with ionomycin and phorbol myristate acetate. Adv Exp
Med Biol. 315:229–239. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tappaz ML: Taurine biosynthetic enzymes
and taurine transporter: Molecular identification and regulations.
Neurochem Res. 29:83–96. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Park E, Park SY, Dobkin C and
Schuller-Levis G: A novel cysteine sulfinic acid decarboxylase
knock-out mouse: Comparison between newborn and weanling mice. Adv
Exp Med Biol. 803:3–16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vogelstein B and Kinzler KW: Cancer genes
and the pathways they control. Nat Med. 10:789–799. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo J, Solimini NL and Elledge SJ:
Principles of cancer therapy: Oncogene and non-oncogene addiction.
Cell. 136:823–837. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sardesai VM: Role of antioxidants in
health maintenance. Nutr Clin Pract. 10:19–25. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Szatrowski TP and Nathan CF: Production of
large amounts of hydrogen peroxide by human tumor cells. Cancer
Res. 51:794–798. 1991.PubMed/NCBI
|
|
41
|
Trachootham D, Alexandre J and Huang P:
Targeting cancer cells by ROS-mediated mechanisms: A radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang X, Tu S, Wang Y, Xu B and Wan F:
Mechanism of taurine-induced apoptosis in human colon cancer cells.
Acta Biochim Biophys Sin (Shanghai). 46:261–272. 2014. View Article : Google Scholar
|
|
43
|
Liu Z, Xia Y, Zhang X, Liu L, Tu S, Zhu W,
Yu L, Wan H, Yu B and Wan F: Roles of the MST1-JNK signaling
pathway in apoptosis of colorectal cancer cells induced by Taurine.
Libyan J Med. 13:15003462018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tu S, Zhang XL, Wan HF, Xia YQ, Liu ZQ,
Yang XH and Wan FS: Effect of taurine on cell proliferation and
apoptosis human lung cancer A549 cells. Oncol Lett. 15:5473–5480.
2018.PubMed/NCBI
|
|
45
|
Wang AS, Lodi A, Rivera LB,
Izquierdo-Garcia JL, Firpo MA, Mulvihill SJ, Tempero MA, Bergers G
and Ronen SM: HR-MAS MRS of the pancreas reveals reduced lipid and
elevated lactate and taurine associated with early pancreatic
cancer. NMR Biomed. 27:1361–1370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Opstad KS, Bell BA, Griffiths JR and Howe
FA: Taurine: A potential marker of apoptosis in gliomas. Br J
Cancer. 100:789–794. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yu J and Kim AK: Effect of taurine on
antioxidant enzyme system in B16F10 melanoma cells. Adv Exp Med
Biol. 643:491–499. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vanitha MK, Anandakumar P and
Sakthisekaran D: Taurine abrogates mammary carcinogenesis through
induction of apoptosis in Sprague-Dawley rats. J Biochem Mol
Toxicol. 32:e222042018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Vanitha MK, Baskaran K, Periyasamy K,
Selvaraj S, Ilakkia A, Saravanan D, Venkateswari R, Revathi Mani B,
Anandakumar P and Sakthisekaran D: Modulatory effect of taurine on
7,12-Dimethylbenz(a)Anthracene-induced alterations in
detoxifi-cation enzyme system, membrane bound enzymes, glycoprotein
profile and proliferative cell nuclear antigen in rat breast
tissue. J Biochem Mol Toxicol. 30:414–423. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang X, Lu H, Wang Y, Liu C, Zhu W, Zheng
S and Wan F: Taurine induces the apoptosis of breast cancer cells
by regulating apoptosis-related proteins of mitochondria. Int J Mol
Med. 35:218–226. 2015. View Article : Google Scholar
|
|
51
|
Choi EJ, Tang Y, Lee CB, Cheong SH, Sung
SH, Oh MR, Young Jang SY, Park PJ and Kim EK: Effect of taurine on
in vitro migration of MCF-7 and MDA-MB-231 human breast carcinoma
cells. Adv Exp Med Biol. 803:191–201. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
He F, Ma N, Midorikawa K, Hiraku Y, Oikawa
S, Zhang Z, Huang G, Takeuchi K and Murata M: Taurine exhibits an
apoptosis-inducing effect on human nasopharyngeal carcinoma cells
through PTEN/Akt pathways in vitro. Amino Acids. 50:1749–1758.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tang Y, Choi E-J, Cheong SH, Hwang YJ,
Arokiyaraj S, Park PJ, Moon SH and Kim EK: Effect of taurine on
prostate-specific antigen level and migration in human prostate
cancer cells. Adv Exp Med Biol. 803:203–214. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chatzakos V, Slätis K, Djureinovic T,
Helleday T and Hunt MC: N-acyl taurines are anti-proliferative in
prostate cancer cells. Lipids. 47:355–361. 2012. View Article : Google Scholar
|
|
55
|
Li H, Ruan WJ, Liu LQ, Wan HF, Yang XH,
Zhu WF, Yu LH, Zhang XL and Wan FS: Impact of taurine on the
proliferation and apoptosis of human cervical carcinoma cells and
its mechanism. Chin Med J (Engl). 132:948–956. 2019. View Article : Google Scholar
|
|
56
|
Srivastava S, Roy R, Singh S, Kumar P,
Dalela D and Sankhwar SN: Taurine - a possible fingerprint
biomarker in non-muscle invasive bladder cancer: A pilot study by
1H NMR spectroscopy. Cancer Biomark. 6:11–20. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang X, Du W, Shen F and Wang J: Research
on effects of taurine on the transplanted tumor of mice. Wei Sheng
Yan Jiu. 26:321–324. 1997.In Chinese.
|
|
58
|
Yousef HN and Aboelwafa HR: The potential
protective role of taurine against 5-fluorouracil-induced
nephrotoxicity in adult male rats. Exp Toxicol Pathol. 69:265–274.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Vanitha MK, Priya KD, Baskaran K,
Periyasamy K, Saravanan D, Venkateswari R, Mani BR, Ilakkia A,
Selvaraj S, Menaka R, et al: Taurine regulates mitochondrial
function during 7,12-dimethyl Benz[a]anthracene induced
experimental mammary carcinogenesis. J Pharmacopuncture. 18:68–74.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tu S, Zhang X, Luo D, Liu Z, Yang X, Wan
H, Yu L, Li H and Wan F: Effect of taurine on the proliferation and
apoptosis of human hepatocellular carcinoma HepG2 cells. Exp Ther
Med. 10:193–200. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sadzuka Y, Matsuura M and Sonobe T: The
effect of taurine, a novel biochemical modulator, on the antitumor
activity of doxo-rubicin. Biol Pharm Bull. 32:1584–1587. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Daigeler A, Chromik AM, Geisler A, Bulut
D, Hilgert C, Krieg A, Klein-Hitpass L, Lehnhardt M, Uhl W and
Mittelkötter U: Synergistic apoptotic effects of taurolidine and
TRAIL on squamous carcinoma cells of the esophagus. Int J Oncol.
32:1205–1220. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Jacobi CA, Menenakos C and Braumann C:
Taurolidine - a new drug with anti-tumor and anti-angiogenic
effects. Anticancer Drugs. 16:917–921. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Rodak R, Kubota H, Ishihara H, Eugster HP,
Könü D, Möhler H, Yonekawa Y and Frei K: Induction of reactive
oxygen intermediates-dependent programmed cell death in human
malignant ex vivo glioma cells and inhibition of the vascular
endothelial growth factor production by taurolidine. J Neurosurg.
102:1055–1068. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Refai NS, Louka ML, Halim HY and Montasser
I: Long non-coding RNAs (CASC2 and TUG1) in hepatocellular
carcinoma: Clinical significance. J Gene Med. 21:e31122019.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Matés JM, Segura JA, Alonso FJ and Márquez
J: Oxidative stress in apoptosis and cancer: An update. Arch
Toxicol. 86:1649–1656. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Mates JM, Segura JA, Alonso FJ and Marquez
J: Sulphur-containing non enzymatic antioxidants: Therapeutic tools
against cancer. Front Biosci (Schol Ed). 4:722–748. 2012.
View Article : Google Scholar
|
|
68
|
Okamoto K, Sugie S, Ohnishi M, Makita H,
Kawamori T, Watanabe T, Tanaka T and Mori H: Chemopreventive
effects of taurine on diethylnitrosamine and phenobarbital-induced
hepato-carcinogenesis in male F344 rats. Jpn J Cancer Res.
87:30–36. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Abd-Allah AR, Gado AM, Al-Majed AA,
Al-Yahya AA and Al-Shabanah OA: Protective effect of taurine
against cyclophosphamide-induced urinary bladder toxicity in rats.
Clin Exp Pharmacol Physiol. 32:167–172. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Islambulchilar M, Asvadi I, Sanaat Z,
Esfahani A and Sattari M: Effect of taurine on attenuating
chemotherapy-induced adverse effects in acute lymphoblastic
leukemia. J Cancer Res Ther. 11:426–432. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Desai TK, Maliakkal J, Kinzie JL,
Ehrinpreis MN, Luk GD and Cejka J: Taurine deficiency after
intensive chemotherapy and/or radiation. Am J Clin Nutr.
55:708–711. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Tabassum H, Parvez S, Rehman H, Dev
Banerjee B, Siemen D and Raisuddin S: Nephrotoxicity and its
prevention by taurine in tamoxifen induced oxidative stress in
mice. Hum Exp Toxicol. 26:509–518. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Tabassum H, Rehman H, Banerjee BD,
Raisuddin S and Parvez S: Attenuation of tamoxifen-induced
hepatotoxicity by taurine in mice. Clin Chim Acta. 370:129–136.
2018. View Article : Google Scholar
|
|
74
|
Parvez S, Tabassum H, Banerjee BD and
Raisuddin S: Taurine prevents tamoxifen-induced mitochondrial
oxidative damage in mice. Basic Clin Pharmacol Toxicol.
102:382–387. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Han X, Yue J and Chesney RW: Functional
TauT protects against acute kidney injury. J Am Soc Nephrol.
20:1323–1332. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Al-Asmari A, Al-Zahrani A, Khan A,
Al-Shahrani H and Ali Al Amri M: Taurine ameliorates
5-flourouracil-induced intestinal mucositis, hepatorenal and
reproductive organ damage in Wistar rats: A biochemical and
histological study. Hum Exp Toxicol. 35:10–20. 2018. View Article : Google Scholar
|
|
77
|
Das J, Ghosh J, Manna P and Sil PC:
Taurine protects rat testes against doxorubicin-induced oxidative
stress as well as p53, Fas and caspase 12-mediated apoptosis. Amino
Acids. 42:1839–1855. 2012. View Article : Google Scholar
|
|
78
|
Das J, Ghosh J, Manna P and Sil PC:
Taurine suppresses doxo-rubicin-triggered oxidative stress and
cardiac apoptosis in rat via up-regulation of PI3-K/Akt and
inhibition of p53, p38-JNK. Biochem Pharmacol. 81:891–909. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Neary PM, Hallihan P, Wang JH, Pfirrmann
RW, Bouchier-Hayes DJ and Redmond HP: The evolving role of
taurolidine in cancer therapy. Ann Surg Oncol. 17:1135–1143. 2010.
View Article : Google Scholar
|
|
80
|
Mobley JA and Brueggemeier RW: Estrogen
receptor-mediated regulation of oxidative stress and DNA damage in
breast cancer. Carcinogenesis. 25:3–9. 2004. View Article : Google Scholar
|
|
81
|
Huang S, Chong N, Lewis NE, Jia W, Xie G
and Garmire LX: Novel personalized pathway-based metabolomics
models reveal key metabolic pathways for breast cancer diagnosis.
Genome Med. 8:342016. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Sitter B, Bathen TF, Singstad TE, Fjøsne
HE, Lundgren S, Halgunset J and Gribbestad IS: Quantification of
metabolites in breast cancer patients with different clinical
prognosis using HR MAS MR spectroscopy. NMR Biomed. 23:424–431.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
El Agouza IM, Eissa SS, El Houseini MM,
El-Nashar DE and Abd El Hameed OM: Taurine: A novel tumor marker
for enhanced detection of breast cancer among female patients.
Angiogenesis. 14:321–330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
He YU, Li QQ and Guo SC: Taurine
attenuates dimethylbenz[a] anthracene-induced breast tumorigenesis
in rats: A plasma metabolomic study. Anticancer Res. 36:533–543.
2016.PubMed/NCBI
|
|
85
|
Zhou DN, Deng YF, Li RH, Yin P and Ye CS:
Concurrent alterations of RAGE, RECK, and MMP9 protein expression
are relevant to Epstein-Barr virus infection, metastasis, and
survival in nasopharyngeal carcinoma. Int J Clin Exp Pathol.
7:3245–3254. 2014.PubMed/NCBI
|
|
86
|
Shennan D and Thomson J: Estrogen
regulation and ion dependence of taurine uptake by MCF-7 human
breast cancer cells. Cell Mol Biol Lett. 12:396–406. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Pine MJ, Kim U and Ip C: Free amino acid
pools of rodent mammary tumors. J Natl Cancer Inst. 69:729–735.
1982.PubMed/NCBI
|
|
88
|
Brown NS and Bicknell R: Hypoxia and
oxidative stress in breast cancer. Oxidative stress: Its effects on
the growth, metastatic potential and response to therapy of breast
cancer. Breast Cancer. 3:323–327. 2001. View Article : Google Scholar
|
|
89
|
Ambrosone CB, Marshall JR, Vena JE,
Laughlin R, Graham S, Nemoto T and Freudenheim JL: Interaction of
family history of breast cancer and dietary antioxidants with
breast cancer risk (New York, United States). Cancer Causes.
6:407–415. 1995. View Article : Google Scholar
|
|
90
|
Freudenheim JL, Marshall JR, Vena JE,
Laughlin R, Brasure JR, Swanson MK, Nemoto T and Graham S:
Premenopausal breast cancer risk and intake of vegetables, fruits,
and related nutrients. J Natl Cancer Inst. 88:340–348. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Kubota A, Meguid MM and Hitch DC: Amino
acid profiles correlate diagnostically with organ site in three
kinds of malignant tumors. Cancer. 69:2343–2348. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Maeda J, Higashiyama M, Imaizumi A,
Nakayama T, Yamamoto H, Daimon T, Yamakado M, Imamura F and Kodama
K: Possibility of multivariate function composed of plasma amino
acid profiles as a novel screening index for non-small cell lung
cancer: A case control study. BMC Cancer. 10:6902010. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Satsu H, Ishimoto Y, Nakano T, Mochizuki
T, Iwanaga T and Shimizu M: Induction by activated macrophage-like
THP-1 cells of apoptotic and necrotic cell death in intestinal
epithelial Caco-2 monolayers via tumor necrosis factor-alpha. Exp
Cell. 312:3909–3919. 2006. View Article : Google Scholar
|
|
94
|
Wang H, Tso VK, Slupsky CM and Fedorak RN:
Metabolomics and detection of colorectal cancer in humans: A
systematic review. Future Oncol. 6:1395–1406. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Scheel C, Eaton EN, Li SH, Chaffer CL,
Reinhardt F, Kah KJ, Bell G, Guo W, Rubin J, Richardson AL and
Weinberg RA: Paracrine and autocrine signals induce and maintain
mesen-chymal and stem cell states in the breast. Cell. 145:926–940.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Turman MV, Kingsley PJ, Rouzer CA, Cravatt
BF and Marnett LJ: Oxidative metabolism of a fatty acid amide
hydrolase-regulated lipid, arachidonoyltaurine. Biochemistry.
47:3917–3925. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
McKinney MK and Cravatt BF: Structure and
function of fatty acid amide hydrolase. Annu Rev. 74:411–432.
2005.
|
|
98
|
Saghatelian A, McKinney MK, Bandell M,
Patapoutian A and Cravatt BF: A FAAH-regulated class of N-acyl
taurines that activates TRP ion channels. Biochemistry.
45:9007–9015. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ueki I, Roman HB, Hirschberger LL, Junior
C and Stipanuk MH: Extrahepatic tissues compensate for loss of
hepatic taurine synthesis in mice with liver-specific knockout of
cysteine dioxy-genase. Am J Physiol Endocrinol Metab.
302:E1292–E1299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Timbrell JA, Seabra V and Waterfield CJ:
The in vivo and in vitro protective properties of taurine. Gen
Pharmacol. 26:453–462. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Heidari R, Babaei H and Eghbal MA:
Ameliorative effects of taurine against methimazole-induced
cytotoxicity in isolated rat hepatocytes. Sci Pharm. 80:987–999.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Boşgelmez İİ, Söylemezoğlu T and Güvendik
G: The protective and antidotal effects of taurine on hexavalent
chromium-induced oxidative stress in mice liver tissue. Biol Trace
Elem Res. 125:46–58. 2008. View Article : Google Scholar
|
|
103
|
Heidari R, Babaei H and Eghbal MA:
Amodiaquine-induced toxicity in isolated rat hepatocytes and the
cytoprotective effects of taurine and/or N-acetyl cysteine. Res
Pharm Sci. 9:97–105. 2014.
|
|
104
|
Sinha M, Manna P and Sil PC: Taurine, a
conditionally essential amino acid, ameliorates arsenic-induced
cytotoxicity in murine hepatocytes. Toxicol In Vitro. 21:1419–1428.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Vissers MC and Fantone JC: Inhibition of
hypochlorous acid-mediated reactions by desferrioxamine.
Implications for the mechanism of cellular injury by neutrophils.
Free Radic Biol Med. 8:331–337. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Riordan JD, Feddersen CR, Tschida BR,
Beckmann PJ, Keng VW, Linden MA, Amin K, Stipp CS, Largaespada DA
and Dupuy AJ: Chronic liver injury alters driver mutation profiles
in hepatocellular carcinoma in mice. Hepatology. 67:924–939. 2018.
View Article : Google Scholar :
|
|
107
|
Glauert HP, Calfee-Mason K, Stemm DN,
Tharappel JC and Spear BT: Dietary antioxidants in the prevention
of hepatocar-cinogenesis: A review. Mol Nutr Food Res. 54:875–896.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Bansal AK, Trivedi R, Soni GL and
Bhatnagar D: Hepatic and renal oxidative stress in acute toxicity
of N-nitrosodiethylamine in rats. Indian J Exp Biol. 38:916–920.
2000.
|
|
109
|
Kang JS, Wanibuchi H, Morimura K, Gonzalez
FJ and Fukushima S: Role of CYP2E1 in diethylnitrosamine-induced
hepatocarcinogenesis in vivo. Cancer Res. 67:11141–11116. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Başaran-Küçükgergin C, Bingül I, Tekkeşin
MS, Olgaç V, Doğru-Abbasoğlu S and Uysal M: Effects of carnosine,
taurine, and betaine pretreatments on diethylnitrosamine-induced
oxida-tive stress and tissue injury in rat liver. Toxicol Ind
Health. 32:1405–1413. 2016. View Article : Google Scholar
|
|
111
|
Chang YY, Chou CH, Chiu CH, Yang KT, Lin
YL, Weng WL and Chen YC: Preventive effects of taurine on
development of hepatic steatosis induced by a high-fat/cholesterol
dietary habit. J Agric Food Chem. 59:450–457. 2011. View Article : Google Scholar
|
|
112
|
Kalaz EB, Çoban J, Aydın AF, Doğan-Ekici
I, Doğru- Abbasoğlu S, Öztezcan S and Uysal M: Carnosine and
taurine treatments decreased oxidative stress and tissue damage
induced by D-galactose in rat liver. J Physiol Biochem. 70:15–25.
2014. View Article : Google Scholar
|
|
113
|
Kerai MD, Waterfield CJ, Kenyon SH, Asker
DS and Timbrell JA: The effect of taurine depletion by beta-alanine
treatment on the susceptibility to ethanol-induced hepatic
dysfunction in rats. Alcohol. 36:29–38. 2001. View Article : Google Scholar
|
|
114
|
You JS and Chang KJ: Taurine protects the
liver against lipid peroxidation and membrane disintegration during
rat hepatocar-cinogenesis. Adv Exp Med Biol. 442:105–112. 1998.
View Article : Google Scholar
|
|
115
|
Liu Y, Li F, Zhang L, Wu J, Wang Y and Yu
H: Taurine alleviates lipopolysaccharide-induced liver injury by
anti-inflammation and antioxidants in rats. Mol Med Rep.
16:6512–6517. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Seabra V, Stachlewitz RF and Thurman RG:
Taurine blunts LPS-induced increases in intracellular calcium and
TNF-alpha production by Kupffer cells. J Leukoc Biol. 64:615–621.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wu G, Yang Q, Yu Y, Lin S, Feng Y, Lv Q,
Yang J and Hu J: Taurine inhibits kupffer cells activation induced
by lipopolysac-charide in alcoholic liver damaged rats. Adv Exp Med
Biol. 975:789–800. 2017. View Article : Google Scholar
|
|
118
|
Kim SK and Kim YC: Attenuation of
bacterial lipopolysaccha-ride-induced hepatotoxicity by betaine or
taurine in rats. Food Chem Toxicol. 40:545–549. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Abd-Rabou AA, Zoheir KMA and Ahmed HH:
Potential impact of curcumin and taurine on human hepatoma cells
using Huh-7 cell line. Clin. 45:1519–1521. 2012.
|
|
120
|
El-Houseini ME, El-Agoza IA, Sakr MM and
El-Malky GM: Novel protective role of curcumin and taurine
combination against experimental hepatocarcinogenesis. Exp Ther
Med. 13:29–36. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Kim YS, Cheong SH, Hwang JW, Lodhi G, Lee
KH, Choi DK, Song H, Lee SH, Park DJ, Ahn CB, et al: Effect of
taurine on viability and proliferation of murine melanoma B16F10
cells. Adv Exp Med Bio. 803:167–177. 2015. View Article : Google Scholar
|
|
122
|
Finnegan N, Toomey D, Condron C, Redmond
HP, Da Costa M and Bouchier-Hayes DJ: Potentiation of the
therapeutic index of interleukin-2 immunotherapy by combination
with taurine in a syngeneic murine tumour model. Ir J Med Sci.
171:85–88. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Son YI, Dallal RM and Lotze MT: Combined
treatment with interleukin-18 and low-dose interleukin-2 induced
regression of a murine sarcoma and memory response. J Immunother
Hagerstown Md. 26:234–240. 2003. View Article : Google Scholar
|
|
124
|
Da Costa ML, Redmond HP and Bouchier-Hayes
DJ: Taurolidine improves survival by abrogating the accelerated
development and proliferation of solid tumors and development of
organ metastases from circulating tumor cells released following
surgery. J Surg Res. 101:111–119. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Logotheti S, Khoury N, Vlahopoulos SA,
Skourti E, Papaevangeliou D, Liloglou T, Gorgoulis V, Budunova I,
Kyriakopoulos AM and Zoumpourlis V: N-bromotaurine surrogates for
loss of antiproliferative response and enhances cisplatin efficacy
in cancer cells with impaired glucocorticoid receptor. Transl Res.
173:58–73.e2. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Gottardi W and Nagl M: N-chlorotaurine, a
natural antiseptic with outstanding tolerability. J Antimicrob
Chemother. 65:399–409. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Carr C, Ng J and Wigmore T: The side
effects of chemotherapeutic agents. Curr Anaesth Crit Care.
19:70–79. 2008. View Article : Google Scholar
|
|
128
|
Bergkvist K and Wengström Y: Symptom
experiences during chemotherapy treatment-With focus on nausea and
vomiting. Eur J Oncol Nurs. 10:21–29. 2006. View Article : Google Scholar
|
|
129
|
Kim KS, Tsuji M, Kimura T and Sezaki H:
Effect of taurine on the gastrointestinal absorption of drugs -
Ionic requirement for the action. J Pharmacobiodyn. 5:172–178.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zeybek A, Ercan F, Çetinel Ş, Çikler E,
Sağlam B and Şener G: Taurine ameliorates water avoidance
stress-induced degenerations of gastrointestinal tract and liver.
Dig Dis Sci. 51:1853–1861. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Abe M, Takahashi M, Takeuchi K and Fukuda
M: Studies on the significance of taurine in radiation injury.
Radiat Res. 33:5631968. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Sener G, Sehirli O, Cetinel S, Midillioğlu
S, Gedik N and Ayanoğlu-Dülger G: Protective effect of taurine
against alendronate-induced gastric damage in rats. Fundam Clin
Pharmacol. 19:93–100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Saransaari P and Oja SS: Taurine and
neural cell damage. Amino Acids. 19:509–526. 2000. View Article : Google Scholar
|
|
134
|
Waters E, Wang JH, Redmond HP, Wu QD, Kay
E and Bouchier-Hayes D: Role of taurine in preventing
acetaminophen-induced hepatic injury in the rat. Am J Physiol
Gastrointest Liver Physiol. 280:G1274–G1279. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Erdem A, Gündogan NÜ, Usubütün A, Kılınç
K, Erdem ŞR, Kara A and Bozkurt A: The protective effect of taurine
against gentamicin-induced acute tubular necrosis in rats. Nephrol
Dial Transplant. 15:1175–1182. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Piao J, Meng F, Fang H, Piao F, Jin B, Li
M and Li W: Effect of taurine on thymus differentiation of
dex-induced immunosup-pressive mice. Adv Exp Med Biol.
1155:381–390. 2019. View Article : Google Scholar
|
|
137
|
Hamaguchi T, Azuma J, Awata N, Ohta H,
Takihara K, Harada H, Kishimoto S and Sperelakis N: Reduction of
doxo-rubicin-induced cardiotoxicity in mice by taurine. Res Commun
Chem Pathol Pharmacol. 59:21–30. 1988.PubMed/NCBI
|
|
138
|
Refik Mas M, Comert B, Oncu K, Vural SA,
Akay C, Tasci I, Ozkomur E, Serdar M, Mas N, Alcigir G and Yener N:
The effect of taurine treatment on oxidative stress in experimental
liver fibrosis. Hepatol Res. 28:207–215. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Kato T, Tsunekawa M, Wang S, Yamashita T
and Ma N: Effect of taurine on iNOS-mediated DNA damage in
drug-induced renal injury. Adv Exp Med Biol. 975:717–727. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Gewirtz D: A critical evaluation of the
mechanisms of action proposed for the antitumor effects of the
anthracycline antibiotics adriamycin and daunorubicin. Biochem
Pharmacol. 57:727–741. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Milic V and Dragojevic V:
Doxorubicin-induced oxida-tive injury of cardiomyocytes - do we
have right strategies for prevention? Cardiotoxicity of oncologic
treatments. Fiuza M: InTech; 2012, http://www.intechopen.com/books/cardiotoxicity-of-oncologic-treatments/doxorubicin-induced-oxidative-injury-of-cardiomyocytes-do-we-have-right-strategies-for-prevention-.
View Article : Google Scholar
|
|
142
|
Ujhazy P, Zaleskis G, Mihich E, Ehrke MJ
and Berleth ES: Doxorubicin induces specific immune functions and
cytokine expression in peritoneal cells. Cancer Immunol Immunother.
52:463–472. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Kim YS, Kim EK, Hwang JW, Kim WS, Shin WB,
Natarajan SB, Moon SH, Jeon BT and Park PJ: Taurine attenuates
doxoru-bicin-induced toxicity on B16F10 cells. Adv Exp Med Biol.
975:1179–1190. 2017. View Article : Google Scholar
|
|
144
|
Grenier MA and Lipshultz SE: Epidemiology
of anthracycline cardiotoxicity in children and adults. Semin
Oncol. 25(Suppl 10): S72–S85. 1998.
|
|
145
|
Volkova M and Russell R III: Anthracycline
cardiotoxicity: Prevalence, pathogenesis and treatment. Curr
Cardiol Rev. 7:214–220. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Comereski CR, Peden WM, Davidson TJ,
Warner GL, Hirth RS and Frantz JD: BR96-doxorubicin conjugate
(BMS-182248) versus doxorubicin: A comparative toxicity assessment
in rats. Toxicol Pathol. 22:473–488. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Nagai K, Fukuno S, Oda A and Konishi H:
Protective effects of taurine on doxorubicin-induced acute
hepatotoxicity through suppression of oxidative stress and
apoptotic responses. Anticancer Drugs. 27:17–23. 2016. View Article : Google Scholar
|
|
148
|
Kim YS, Sung SH, Tang Y, Choi EJ, Choi YJ,
Hwang YJ, Park PJ and Kim EK: Protective effect of taurine on mice
with doxorubicin-induced acute kidney injury. Adv Exp Med Biol.
975:1191–1201. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Adedara IA, Ojuade TJD, Olabiyi BF, Idris
UF, Onibiyo EM, Ajeigbe OF and Farombi EO: Taurine ameliorates
renal oxida-tive damage and thyroid dysfunction in rats chronically
exposed to fluoride. Biol Trace Elem Res. 175:388–395. 2017.
View Article : Google Scholar
|
|
150
|
Mohamed RH, Karam RA and Amer MG:
Epicatechin attenuates doxorubicin-induced brain toxicity: Critical
role of TNF-α, iNOS and NF-κB. Brain Res Bull. 86:22–28. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Gradishar WJ and Vokes EE: 5-Fluorouracil
cardiotoxicity: A critical review. Ann Oncol. 1:409–414. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
D'Souza UJ and Narayana K: Induction of
seminiferous tubular atrophy by single dose of 5-fluorouracil
(5-FU) in Wistar rats. Indian J Physiol Pharmacol. 45:87–94.
2001.PubMed/NCBI
|
|
153
|
Narayana K, D'Souza UJ, Sanyal AK and Rao
KP: 5-fluoro-uracil (5-FU) induces the formation of giant cells and
sloughing of seminiferous epithelium in the rat testis. Indian J
Physiol Pharmacol. 44:317–322. 2000.PubMed/NCBI
|
|
154
|
Cheah KY, Howarth GS, Yazbeck R, Wright
TH, Whitford EJ, Payne C, Butler RN and Bastian SE: Grape seed
extract protects IEC-6 cells from chemotherapy-induced cytotoxicity
and improves parameters of small intestinal mucositis in rats with
experimentally-induced mucositis. Cancer Biol Ther. 8:382–390.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Son JY, Shin JW, Wang JH, Park HJ, Kim HG,
Raghavendran HR and Son CG: Chemotherapy-induced myelotoxicity and
incidence of lung metastasis in an animal model. Hum Exp Toxicol.
30:649–655. 2011. View Article : Google Scholar
|
|
156
|
Tsibiribi P, Bui-Xuan C, Bui-Xuan B,
Lombard-Bohas C, Duperret S, Belkhiria M, Tabib A, Maujean G,
Descotes J and Timour Q: Cardiac lesions induced by 5-fluorouracil
in the rabbit. Hum Exp Toxicol. 25:305–309. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Rashid S, Ali N, Nafees S, Hasan SK and
Sultana S: Mitigation of 5-Fluorouracil induced renal toxicity by
chrysin via targeting oxidative stress and apoptosis in wistar
rats. Food Chem Toxicol. 66:185–193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Heidari R, Rasti M, Shirazi Yeganeh B,
Niknahad H, Saeedi A and Najibi A: Sulfasalazine-induced renal and
hepatic injury in rats and the protective role of taurine.
Bioimpacts. 6:3–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Han X: Targeting Taurine Transporter
(TauT) for cancer immu-notherapy of p53 mutation mediated cancers -
molecular basis and preclinical implication. Adv Exp Med Biol.
1155:543–553. 2019. View Article : Google Scholar
|
|
160
|
Nazarewicz RR, Zenebe WJ, Parihar A,
Larson SK, Alidema E, Choi J and Ghafourifar P: Tamoxifen induces
oxidative stress and mitochondrial apoptosis via stimulating
mitochondrial nitric oxide synthase. Cancer Res. 67:1282–1290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Deng X, Liang J, Lin ZX, Wu FS, Zhang YP
and Zhang ZW: Natural taurine promotes apoptosis of human hepatic
stel-late cells in proteomics analysis. World J Gastroenterol.
16:1916–1923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Alam SS, Hafiz NA and Abd El-Rahim AH:
Protective role of taurine against genotoxic damage in mice treated
with methotrexate and tamoxfine. Environ Toxicol Pharmacol.
31:143–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Fontanelli R, Spatti G, Raspagliesi F,
Zunino F and Di Re F: A preoperative single course of high-dose
cisplatin and bleomycin with glutathione protection in bulky stage
IB/II carcinoma of the cervix. Ann Oncol. 3:117–122. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Panici PB, Greggi S, Scambia G, Ragusa G,
Baiocchi G, Battaglia F, Coronetta F and Mancuso S: High-dose
cisplatin and bleomycin neoadjuvant chemotherapy plus radical
surgery in locally advanced cervical carcinoma: A preliminary
report. Gynecol Oncol. 41:212–216. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Pabla N and Dong Z: Cisplatin
nephrotoxicity: Mechanisms and renoprotective strategies. Kidney
Int. 73:994–1007. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Ries F and Klastersky J: Nephrotoxicity
induced by cancer chemotherapy with special emphasis on cisplatin
toxicity. Am J Kidney Dis. 8:368–379. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Townsend DM, Deng M, Zhang L, Lapus MG and
Hanigan MH: Metabolism of cisplatin to a nephrotoxin in proximal
tubule cells. J Am Soc Nephrol. 14:1–10. 2003. View Article : Google Scholar
|
|
168
|
Francescato HDC, Costa RS, Scavone C and
Coimbra TM: Parthenolide reduces cisplatin-induced renal damage.
Toxicology. 230:64–75. 2007. View Article : Google Scholar
|
|
169
|
Yao X, Panichpisal K, Kurtzman N and
Nugent K: Cisplatin nephrotoxicity: A review. Am J Med Sci.
334:115–124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Chtourou Y, Aouey B, Aroui S, Kebieche M
and Fetoui H: Anti-apoptotic and anti-inflammatory effects of
naringin on cisplatin-induced renal injury in the rat. Chem Biol
Interact. 243:1–9. 2016. View Article : Google Scholar
|
|
171
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics 2018: Ovarian cancer statistics 2018. CA
Cancer J Clin. 68:284–296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Han X and Chesney RW: Regulation of TauT
by cisplatin in LLC-PK1 renal cells. Pediatr Nephrol. 20:1067–1072.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Shalby AB, Assaf N and Ahmed HH: Possible
mechanisms for N-acetyl cysteine and taurine in ameliorating acute
renal failure induced by cisplatin in rats. Toxicol Mech Methods.
21:538–546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Kim T and Kim AK: Taurine enhances
anticancer activity of cisplatin in human cervical cancer cells.
Adv Exp Med Biol. 776:189–198. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Tsunekawa M, Wang S, Kato T, Yamashita T
and Ma N: Taurine administration mitigates cisplatin induced acute
neph-rotoxicity by decreasing DNA damage and inflammation: An
Immunocytochemical Study. Adv Exp Med Biol. 975:703–716. 2017.
View Article : Google Scholar
|
|
176
|
Sørensen BH, Thorsteinsdottir UA and
Lambert IH: Acquired cisplatin resistance in human ovarian A2780
cancer cells correlates with shift in taurine homeostasis and
ability to volume regulate. Am J Physiol Cell Physiol.
307:C1071–C1080. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Sugiura H, Okita S, Kato T, Naka T,
Kawanishi S, Ohnishi S, Oshida Y and Ma N: Protection by taurine
against INOS-dependent DNA damage in heavily exercised skeletal
muscle by inhibition of the NF-κB signaling pathway. Adv Exp Med
Biol. 775:237–246. 2013. View Article : Google Scholar
|
|
178
|
Ma N, Kato T, Isogai T, Gu Y and Yamashita
T: The potential effects of taurine in mitigation of radiation
nephropathy. Adv Exp Med Biol. 1155:497–505. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Owoeye O, Adedara IA and Farombi EO:
Pretreatment with taurine prevented brain injury and exploratory
behaviour associated with administration of anticancer drug
cisplatin in rats. Biomed Pharmacother. 102:375–384. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Bishnu A, Sakpal A, Ghosh N, Choudhury P,
Chaudhury K and Ray P: Long term treatment of metformin impedes
development of chemoresistance by regulating cancer stem cell
differentiation through taurine generation in ovarian cancer cells.
Int J Biochem Cell Biol. 107:116–127. 2019. View Article : Google Scholar
|
|
181
|
Badary OA: Taurine attenuates fanconi
syndrome induced by ifosfamide without compromising its antitumor
activity. Oncol Res. 10:355–360. 1998.
|
|
182
|
Han X and Chesney RW: The role of taurine
in renal disorders. Amino Acids. 43:2249–2263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Bouckenooghe T, Remacle C and Reusens B:
Is taurine a functional nutrient? Curr Opin Clin Nutr Metab Care.
9:728–733. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Dotan E, Aggarwal C and Smith MR: Impact
of rituximab (Rituxan) on the treatment of B-cell Non-Hodgkin's
lymphoma. P T. 35:148–157. 2010.PubMed/NCBI
|
|
185
|
Han YM, Awng N, Nu LH, Thway NM and
McLiesh P: Orthopaedic nursing in developing nations: A
collaboration between the Republic of the Union of Myanmar (Burma)
and Australia. Int J Orthop Trauma Nurs. 27:41–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Dong JF, Zheng X-Q and Rui HB: Effect of
taurine on immune function in mice with T-cell lymphoma during
chemotherapy. Asian Pac J Trop Med. 10:1090–1094. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
D'souza M, Jaimini A, Bansal A, Tripathi
M, Sharma R, Mondal A and Tripathi RP: FDG-PET/CT in lymphoma.
Indian J Radiol Imaging. 23:354–365. 2013. View Article : Google Scholar
|
|
188
|
Marcinkiewicz J, Grabowska A, Bereta J,
Bryniarski K and Nowak B: Taurine chloramine down-regulates the
generation of murine neutrophil inflammatory mediators.
Immunopharmacology. 40:27–38. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Klebanoff SJ:
Myeloperoxidase-halide-hydrogen peroxide antibacterial system. J
Bacteriol. 95:2131–2138. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Thomas EL: Myeloperoxidase-hydrogen
peroxide-chloride antimicrobial system: Effect of exogenous amines
on antibacterial action against Escherichia coli. Infect Immun.
25:110–116. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Babior BM: Oxygen-dependent microbial
killing by phagocytes (first of two parts). N Engl J Med.
298:659–668. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Henderson JP, Byun J, Williams MV, Mueller
DM, McCormick ML and Heinecke JW: Production of brominating
intermediates by myeloperoxidase. A transhalogenation pathway for
generating mutagenic nucleobases during inflammation. J Biol Chem.
276:7867–7875. 2001. View Article : Google Scholar
|
|
193
|
Klebanoff SJ: Myeloperoxidase: Friend and
foe. J Leukoc Biol. 77:598–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
van Dalen CJ and Kettle AJ: Substrates and
products of eosino-phil peroxidase. Biochem J. 358:233–239. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Weiss SJ, Klein R, Slivka A and Wei M:
Chlorination of taurine by human neutrophils. Evidence for
hypochlorous acid generation. J Clin Invest. 70:598–607. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Marcinkiewicz J, Strus M, Walczewska M,
Machul A and Mikołajczyk D: Influence of taurine haloamines (TauCl
and TauBr) on the development of pseudomonas aeruginosa biofilm: A
preliminary study. Adv Exp Med Biol. 775:269–283. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Tokunaga S, Kanayama A and Miyamoto Y:
Modification of IkappaBalpha by taurine bromamine inhibits tumor
necrosis factor alpha-induced NF-kappaB activation. Inflamm Res.
56:479–486. 2007. View Article : Google Scholar
|
|
198
|
Midwinter RG, Peskin AV, Vissers MC and
Winterbourn CC: Extracellular oxidation by taurine chloramine
activates ERK via the epidermal growth factor receptor. J Biol
Chem. 279:32205–32211. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Grisham MB, Jefferson MM, Melton DF and
Thomas EL: Chlorination of endogenous amines by isolated
neutrophils. Ammonia-dependent bactericidal, cytotoxic, and
cytolytic activities of the chloramines. J Biol Chem.
259:10404–10413. 1984.PubMed/NCBI
|
|
200
|
Thomas EL, Grisham MB and Jefferson MM:
Myeloperoxidase- dependent effect of amines on functions of
isolated neutrophils. J Clin Invest. 72:441–454. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Kim C and Kang IS: Taurine chloramine, a
taurine metabolite from activated neutrophils, inhibits
osteoclastogenesis by suppressing NFATc1 expression. Adv Exp Med
Biol. 803:99–107. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Walczewska M, Peruń A, Białecka A, Śróttek
M, Jamróz W, Dorożyński P, Jachowicz R, Kulinowski P, Nagl M,
Gottardi W and Marcinkiewicz J: Comparative analysis of
microbicidal and anti-inflammatory properties of novel taurine
bromamine derivatives and bromamine T. Adv Exp Med Biol.
975:515–534. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Gottardi W, Hagleitner M and Nagl M: N,
N-Dichlorotaurine: Chemical and bactericidal properties. Arch Pharm
(Weinheim). 338:473–483. 2005. View Article : Google Scholar
|
|
204
|
Gottardi W and Nagl M: Chlorine covers on
living bacteria: The initial step in antimicrobial action of active
chlorine compounds. J Antimicrob Chemother. 55:475–482. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Roos D, Eckmann CM and Yazdanbakhsh M:
Killing of schis-tosomula by taurine chloramine and taurine
bromamine. Am J Trop Med Hyg. 37:106–110. 1987. View Article : Google Scholar
|
|
206
|
Nagl M, Nguyen VA, Gottardi W, Ulmer H and
Höpfl R: Tolerability and efficacy of N-chlorotaurine in comparison
with chloramine T for the treatment of chronic leg ulcers with a
purulent coating: A randomized phase II study. Br J Dermatol.
149:590–597. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Marcinkiewicz J, Wojas-Pelc A, Walczewska
M, Lipko- Godlewska S, Jachowicz R, Maciejewska A, Białecka A and
Kasprowicz A: Topical taurine bromamine, a new candidate in the
treatment of moderate inflammatory acne vulgaris: A pilot study.
Eur J Dermatol. 18:433–439. 2008.PubMed/NCBI
|
|
208
|
Nagl M, Hess MW, Pfaller K, Hengster P and
Gottardi W: Bactericidal activity of micromolar N-chlorotaurine:
Evidence for its antimicrobial function in the human defense
system. Antimicrob Agents Chemother. 44:2507–2513. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Nagl M, Teuchner B, Pöttinger E, Ulmer H
and Gottardi W: Tolerance of N-chlorotaurine, a new antimicrobial
agent, in infectious conjunctivitis - a phase II pilot study.
Ophthalmologica. 214:111–114. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Neher A, Gstöttner M, Nagl M, Scholtz A
and Gunkel AR: N-chlorotaurine - a new safe substance for
postoperative ear care. Auris Nasus Larynx. 34:19–22. 2007.
View Article : Google Scholar
|
|
211
|
Neher A, Nagl M, Appenroth E, Gstöttner M,
Wischatta M, Reisigl F, Schindler M, Ulmer H and Stephan K: Acute
otitis externa: Efficacy and tolerability of n-chlorotaurine, a
novel endogenous antiseptic agent. Laryngoscope. 114:850–854. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Mainnemare A, Mégarbane B, Soueidan A,
Daniel A and Chapple IL: Hypochlorous acid and
taurine-N-monochloramine in periodontal diseases. J Dent Res.
83:823–831. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Marcinkiewicz J: Taurine bromamine (TauBr)
- its role in immunity and new perspectives for clinical use. J
Biomed Sci. 17(Suppl 1): S32010. View Article : Google Scholar :
|
|
214
|
Eitzinger C, Ehrlenbach S, Lindner H,
Kremser L, Gottardi W, Debabov D, Anderson M, Nagl M and Orth D:
N-chlorotaurine, a long-lived oxidant produced by human leukocytes,
inactivates Shiga toxin of enterohemorrhagic Escherichia coli. PLoS
One. 7:e471052012. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Kim C, Jang JS, Cho MR, Agarawal SR and
Cha YN: Taurine chloramine induces heme oxygenase-1 expression via
Nrf2 activation in murine macrophages. Int Immunopharmacol.
10:440–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Schuller-Levis GB and Park E: Taurine: New
implications for an old amino acid. FEMS Microbiol Lett.
226:195–202. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Green TR, Fellman JH, Eicher AL and Pratt
KL: Antioxidant role and subcellular location of hypotaurine and
taurine in human neutrophils. Biochim Biophys Acta. 1073:91–97.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Jeon SH, Lee MY, Rahman MM, Kim SJ, Kim
GB, Park SY, Hong CU, Kim SZ, Kim JS and Kang HS: The antioxidant,
taurine reduced lipopolysaccharide (LPS)-induced generation of ROS,
and activation of MAPKs and Bax in cultured pneumo-cytes. Pulm
Pharmacol Ther. 22:562–566. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Oliveira MWS, Minotto JB, de Oliveira MR,
Zanotto-Filho A, Behr GA, Rocha RF, Moreira JC and Klamt F:
Scavenging and antioxidant potential of physiological taurine
concentrations against different reactive oxygen/nitrogen species.
Pharmacol Rep. 62:185–193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Marcinkiewicz J, Mak M, Bobek M, Biedroń
R, Białecka A, Koprowski M, Kontny E and Maśliński W: Is there a
role of taurine bromamine in inflammation? Interactive effects with
nitrite and hydrogen peroxide. Inflamm Res. 54:42–49. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Kontny E, Chorąży-Massalska M, Rudnicka W,
Marcinkiewicz J and Maśliński W: Comparison of taurine chloramine
and taurine bromamine effects on rheumatoid arthritis synoviocytes.
Amino Acids. 32:447–452. 2007. View Article : Google Scholar
|
|
222
|
Park E, Schuller-Levis G, Jia JH and Quinn
MR: Preactivation exposure of RAW 264.7 cells to taurine chloramine
attenuates subsequent production of nitric oxide and expression of
iNOS mRNA. J Leukoc Biol. 61:161–166. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Park E, Jia J, Quinn MR and Schuller-Levis
G: Taurine chlora-mine inhibits lymphocyte proliferation and
decreases cytokine production in activated human leukocytes. Clin
Immunol. 102:179–184. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Kontny E, Maśliński W and Marcinkiewicz J:
Anti-inflammatory activities of taurine chloramine: Implication for
immunoregulation and pathogenesis of rheumatoid arthritis. Adv Exp
Med Biol. 526:329–340. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Olszanecki R and Marcinkiewicz J: Taurine
chloramine and taurine bromamine induce heme oxygenase-1 in resting
and LPS-stimulated J774.2 macrophages. Amino Acids. 27:29–35. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Kim K, Choi HM, Oh D, Kim C, Jeong JS, Yoo
M and Yang HI: Effect of taurine chloramine on the production of
matrix metal-loproteinases (MMPs) in adiponectin- or
IL-1beta-stimulated fibroblast-like synoviocytes. J Biomed Sci.
17(Suppl 1): S272010. View Article : Google Scholar :
|
|
227
|
Olszanecki R, Kurnyta M, Biedroń R,
Chorobik P, Bereta M and Marcinkiewicz J: The role of heme
oxygenase-1 in down regulation of PGE2 production by taurine
chloramine and taurine bromamine in J774.2 macrophages. Amino
Acids. 35:359–364. 2008. View Article : Google Scholar
|
|
228
|
Araujo JA, Zhang M and Yin F: Heme
oxygenase-1, oxidation, inflammation, and atherosclerosis. Front
Pharmacol. 3:1192012. View Article : Google Scholar : PubMed/NCBI
|
|
229
|
Idelman G, Smith DLH and Zucker SD:
Bilirubin inhibits the up-regulation of inducible nitric oxide
synthase by scavenging reactive oxygen species generated by the
toll-like receptor 4-dependent activation of NADPH oxidase. Redox
Biol. 5:398–408. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Jong CJ, Azuma J and Schaffer S: Mechanism
underlying the antioxidant activity of taurine: Prevention of
mitochondrial oxidant production. Amino Acids. 42:2223–2232. 2012.
View Article : Google Scholar
|
|
231
|
Tallan HH, Jacobson E, Wright CE,
Schneidman K and Gaull GE: Taurine uptake by cultured human
lymphoblastoid cells. Life Sci. 33:1853–1860. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Kim C, Chung JK, Jeong JM, Chang YS, Lee
YJ, Kim YJ, Lee MC, Koh CS and Kim BK: Uptake of taurine and
taurine chloramine in murine macrophages and their distribution in
mice with experimental inflammation. Adv Exp Med Biol. 442:169–176.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Kwaśny-Krochin B, Bobek M, Kontny E,
Gluszko P, Biedroń R, Chain BM, Maśliński W and Marcinkiewicz J:
Effect of taurine chloramine, the product of activated neutrophils,
on the development of collagen-induced arthritis in DBA 1/J mice.
Amino Acids. 23:419–426. 2002. View Article : Google Scholar
|
|
234
|
Chung Y-L, Wassif WS, Bell JD, Hurley M
and Scott DL: Urinary levels of creatine and other metabolites in
the assessment of polymyositis and dermatomyositis. Rheumatology
(Oxford). 42:298–303. 2003. View Article : Google Scholar
|
|
235
|
Kim H, Jeon H, Kong H, Yang Y, Choi B, Kim
YM, Neckers L and Jung Y: A molecular mechanism for the
anti-inflammatory effect of taurine-conjugated 5-aminosalicylic
acid in inflamed colon. Mol Pharmacol. 69:1405–1412. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Quinn MR, Park E and Schuller-Levis G:
Taurine chloramine inhibits prostaglandin E2 production in
activated raw 264.7 cells by post-transcriptional effects on
inducible cyclooxygenase expression. Immunol Lett. 50:185–188.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Park E, Schuller-Levis G and Quinn MR:
Taurine chloramine inhibits production of nitric oxide and
TNF-alpha in activated RAW 264.7 cells by mechanisms that involve
transcriptional and translational events. J Immunol. 154:4778–4784.
1995.PubMed/NCBI
|
|
238
|
Kim C, Park E, Quinn MR and Schuller-Levis
G: The production of superoxide anion and nitric oxide by cultured
murine leukocytes and the accumulation of TNF-alpha in the
conditioned media is inhibited by taurine chloramine.
Immunopharmacology. 34:89–95. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Barua M, Liu Y and Quinn MR: Taurine
chloramine inhibits inducible nitric oxide synthase and TNF-alpha
gene expression in activated alveolar macrophages: Decreased
NF-kappaB activation and IkappaB kinase activity. J Immunol.
167:2275–2281. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Park E, Quinn MR, Wright CE and
Schuller-Levis G: Taurine chloramine inhibits the synthesis of
nitric oxide and the release of tumor necrosis factor in activated
RAW 264.7 cells. J Leukoc Biol. 54:119–124. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Sun Jang J, Piao S, Cha YN and Kim C:
Taurine chloramine activates Nrf2, increases HO-1 expression and
protects cells from death caused by hydrogen peroxide. J Clin
Biochem Nutr. 45:37–43. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Fazzino F, Obregón F, Morles M, Rojas A,
Arocha L, Mata S and Lima L: Taurine transporter in lymphocytes of
patients with major depression treated with venlafaxine plus
psychotherapy. Adv Exp Med Biol. 643:217–224. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Wirleitner B, Neurauter G, Nagl M and
Fuchs D: Down-regulatory effect of N-chlorotaurine on tryptophan
degradation and neopterin production in human PBMC. Immunol Lett.
93:143–149. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
244
|
Kanayama A, Inoue JI, Sugita-Konishi Y,
Shimizu M and Miyamoto Y: Oxidation of Ikappa Balpha at methionine
45 is one cause of taurine chloramine-induced inhibition of
NF-kappa B activation. J Biol Chem. 277:24049–24056. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Liu Y and Quinn MR: Chemokine production
by rat alveolar macrophages is inhibited by taurine chloramine.
Immunol Lett. 80:27–32. 2002. View Article : Google Scholar
|
|
246
|
Cobb MH, Xu S, Hepler JE, Hutchison M,
Frost J and Robbins DJ: Regulation of the MAP kinase cascade. Cell
Mol Biol Res. 40:253–256. 1994.PubMed/NCBI
|
|
247
|
Crews CM, Alessandrini A and Erikson RL:
The primary structure of MEK, a protein kinase that phosphorylates
the ERK gene product. Science. 258:478–480. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
248
|
Kim JW and Kim C: Inhibition of
LPS-induced NO production by taurine chloramine in macrophages is
mediated though Ras-ERK-NF-kappaB. Biochem Pharmacol. 70:1352–1360.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
249
|
Bhat NR, Zhang P, Lee JC and Hogan EL:
Extracellular signal-regulated kinase and p38 subgroups of
mitogen-activated protein kinases regulate inducible nitric oxide
synthase and tumor necrosis factor-alpha gene expression in
endotoxin-stimulated primary glial cultures. J Neurosci.
18:1633–1641. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
250
|
Ajizian SJ, English BK and Meals EA:
Specific inhibitors of p38 and extracellular signal-regulated
kinase mitogen-activated protein kinase pathways block inducible
nitric oxide synthase and tumor necrosis factor accumulation in
murine macrophages stimulated with lipopolysaccharide and
interferon-gamma. J Infect Dis. 179:939–944. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Chen CC and Wang JK: p38 but not p44/42
mitogen-activated protein kinase is required for nitric oxide
synthase induction mediated by lipopolysaccharide in RAW 264.7
macrophages. Mol Pharmacol. 55:481–488. 1999.PubMed/NCBI
|
|
252
|
Kontny E, Szczepańska K, Kowalczewski J,
Kurowska M, Janicka I, Marcinkiewicz J and Maśliński W: The
mechanism of taurine chloramine inhibition of cytokine
(interleukin-6, interleukin-8) production by rheumatoid arthritis
fibroblast-like synoviocytes. Arthritis Rheum. 43:2169–2177. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
253
|
Wojtecka-Lukasik E, Gujski M, Roguska K,
Maslinska D and Maslinski S: Taurine chloramine modifies adjuvant
arthritis in rats. Inflamm Res. 54(Suppl 1): S21–S22. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
254
|
Verdrengh M and Tarkowski A: Inhibition of
septic arthritis by local administration of taurine chloramine, a
product of activated neutrophils. J Rheumatol. 32:1513–1517.
2005.PubMed/NCBI
|
|
255
|
Kontny E, Plebanczyk M, Lisowska B,
Olszewska M, Maldyk P and Maslinski W: Comparison of rheumatoid
articular adipose and synovial tissue reactivity to proinflammatory
stimuli: Contribution to adipocytokine network. Ann Rheum Dis.
71:262–267. 2012. View Article : Google Scholar
|
|
256
|
Kontny E, Grabowska A, Kowalczewski J,
Kurowska M, Janicka I, Marcinkiewicz J and Maśliński W: Taurine
chlora-mine inhibition of cell proliferation and cytokine
production by rheumatoid arthritis fibroblast-like synoviocytes.
Arthritis Rheum. 42:2552–2560. 1999. View Article : Google Scholar
|
|
257
|
Kontny E, Rudnicka W, Kowalczewski J,
Marcinkiewicz J and Maslinski W: Selective inhibition of
cyclooxygenase 2-generated prostaglandin E2 synthesis in rheumatoid
arthritis synoviocytes by taurine chloramine. Arthritis Rheum.
48:1551–1555. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
258
|
Kontny E, Rudnicka W, Chorąży-Massalska M,
Marcinkiewicz J and Maśliński W: Taurine chloramine inhibits
proliferation of rheumatoid arthritis synoviocytes by triggering a
p53-dependent pathway. Inflamm Res. 55:446–455. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
259
|
Kim KS, Park EK, Ju SM, Jung HS, Bang JS,
Kim C, Lee YA, Hong SJ, Lee SH, Yang HI and Yoo MC: Taurine
chloramine differentially inhibits matrix metalloproteinase 1 and
13 synthesis in interleukin-1beta stimulated fibroblast-like
synovio-cytes. Arthritis Res Ther. 9:R802007. View Article : Google Scholar
|
|
260
|
Wang Y, Cha YN, Kim KS and Kim C: Taurine
chloramine inhibits osteoclastogenesis and splenic lymphocyte
proliferation in mice with collagen-induced arthritis. Eur J
Pharmacol. 668:325–330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
261
|
Davies EV, Williams BD and Campbell AK:
Synovial fluid polymorphonuclear leucocytes from patients with
rheumatoid arthritis have reduced MPO and NADPH-oxidase activity.
Br J Rheumatol. 29:415–421. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
262
|
Kanayama A and Miyamoto Y: Apoptosis
triggered by phago-cytosis-related oxidative stress through
FLIPS downregulation and JNK activation. J Leukoc Biol.
82:1344–1352. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
263
|
Emerson DK, McCormick ML, Schmidt JA and
Knudson CM: Taurine monochloramine activates a cell death pathway
involving Bax and Caspase-9. J Biol Chem. 280:3233–3241. 2005.
View Article : Google Scholar
|
|
264
|
Vile GF, Rothwell LA and Kettle AJ:
Initiation of rapid, P53-dependent growth arrest in cultured human
skin fibroblasts by reactive chlorine species. Arch Biochem
Biophys. 377:122–128. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
265
|
Klamt F and Shacter E: Taurine chloramine,
an oxidant derived from neutrophils, induces apoptosis in human B
lymphoma cells through mitochondrial damage. J Biol Chem.
280:21346–21352. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
266
|
Pilz M, Holinka J, Vavken P, Marian B and
Krepler P: Taurine chloramine induces apoptosis in human
osteosarcoma cell lines. J Orthop Res. 30:2046–2051. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
267
|
Gupta R, Seki Y and Yosida J: Role of
taurine in spinal cord injury. Curr Neurovasc Res. 3:225–235. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
268
|
Kulakowski EC and Maturo J: Hypoglycemic
properties of taurine: Not mediated by enhanced insulin release.
Biochem Pharmacol. 33:2835–2838. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
269
|
McCartney AC and Browne MK: Clinical
studies on administration of taurolin in severe sepsis: A
preliminary study. Prog Clin Biol Res. 272:361–371. 1988.PubMed/NCBI
|
|
270
|
Staubach KH: Adjuvant therapy of
peritonitis with taurolidine. Modulation of mediator liberation.
Langenbecks Arch Chir. 382(Suppl 1): S26–S30. 1997.In German.
View Article : Google Scholar
|
|
271
|
Wesch G, Petermann C and Linder MM: Drug
therapy of peritonitis. 6-year experience with the chemotherapeutic
agent and anti-endotoxin taurolin. Fortschr Med. 101:545–550.
1983.In German. PubMed/NCBI
|
|
272
|
Jurewitsch B and Jeejeebhoy KN:
Taurolidine lock: The key to prevention of recurrent
catheter-related bloodstream infections. Clin Nutr. 24:462–465.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Liu Y, Zhang AQ, Cao L, Xia HT and Ma JJ:
Taurolidine lock solutions for the prevention of catheter-related
bloodstream infections: A systematic review and meta-analysis of
random-ized controlled trials. PLoS One. 8:e794172013. View Article : Google Scholar
|
|
274
|
Hayes KC, Stephan ZF and Sturman JA:
Growth depression in taurine-depleted infant monkeys. J Nutr.
110:2058–2064. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
275
|
Braumann C, Winkler G, Rogalla P,
Menenakos C and Jacobi CA: Prevention of disease progression in a
patient with a gastric cancer-re-recurrence. Outcome after
intravenous treatment with the novel antineoplastic agent
taurolidine. Report of a case. World J Surg Onco. 4:342006.
View Article : Google Scholar
|
|
276
|
Stendel R, Picht T, Schilling A,
Heidenreich J, Loddenkemper C, Jänisch W and Brock M: Treatment of
glioblastoma with intravenous taurolidine. First clinical
experience Anticancer Res. 24:1143–1147. 2004.
|
|
277
|
Teuchner B, Nagl M, Schidlbauer A, Ishiko
H, Dragosits E, Ulmer H, Aoki K, Ohno S, Mizuki N, Gottardi W and
Larcher C: Tolerability and efficacy of N-chlorotaurine in epidemic
kera-toconjunctivitis-A double-blind, randomized, phase-2 clinical
trial. J Ocul Pharmacol Ther. 21:157–165. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
278
|
Pasich E, Walczewska M, Białecka A, Peruń
A, Kasprowicz A and Marcinkiewicz J: Taurine haloamines and
biofilm: II. Efficacy of taurine bromamine and chlorhexidine
against selected microorganisms of oral biofilm. Adv Exp Med Biol.
803:133–143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
279
|
Kyriakopoulos A, Logotheti S,
Marcinkiewicz J and Nagl M: N-chlorotaurine and N-bromotaurine
combination regimen for the cure of valacyclovir-unresponsive
herpes zoster comorbidity in a multiple sclerosis patient. Int J
Med Pharm Case Rep. 7:1–6. 2016. View Article : Google Scholar
|
|
280
|
Kyriakopoulos AM, Nagl M, Orth-Höller D,
Marcinkiewicz J, Baliou S and Zoumbourlis V: Successful treatment
of a unique chronic multi-bacterial scalp infection with
N-chlorotaurine, N-bromotaurine and bromamine T. Access Microbiol:
https://doi.org/10.1099/acmi.0.000126.
|
|
281
|
Zhai X, Zhao J, Wang Y, Wei X, Li G, Yang
Y, Chen Z, Bai Y, Wang Q, Chen X and Li M: Bibliometric analysis of
global scientific research on lncRNA: A swiftly expanding trend.
Biomed Res Int. 2018:76250782018. View Article : Google Scholar : PubMed/NCBI
|
|
282
|
Di Gesualdo F, Capaccioli S and Lulli M: A
pathophysi-ological view of the long non-coding RNA world.
Oncotarget. 5:10976–10996. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
283
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
284
|
Thin KZ, Liu X, Feng X, Raveendran S and
Tu JC: LncRNA- DANCR: A valuable cancer related long non-coding RNA
for human cancers. Pathol Res Pract. 214:801–805. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
285
|
Dykes IM and Emanueli C: Transcriptional
and post-transcriptional gene regulation by long non-coding RNA.
Genomics Proteomics Bioinformatics. 15:177–186. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
286
|
Dey BK, Mueller AC and Dutta A: Long
non-coding RNAs as emerging regulators of differentiation,
development, and disease. Transcription. 5:e9440142014. View Article : Google Scholar : PubMed/NCBI
|
|
287
|
Li CH and Chen Y: Targeting long
non-coding RNAs in cancers: Progress and prospects. Int J Biochem
Cell Biol. 45:1895–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
288
|
Herriges MJ, Swarr DT, Morley MP, Rathi
KS, Peng T, Stewart KM and Morrisey EE: Long noncoding RNAs are
spatially correlated with transcription factors and regulate lung
development. Genes Dev. 28:1363–1379. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
289
|
Kołat D, Hammouz R, Bednarek A and
Płuciennik E: Exosomes as carriers transporting long non-coding
RNAs: Molecular characteristics and their function in cancer
(Review). Mol Med Rep. 20:851–862. 2019.
|
|
290
|
Xiao Y, Zhang J and Deng L: Prediction of
lncRNA-protein interactions using HeteSim scores based on
heterogeneous networks. Sci Rep. 7:36642017. View Article : Google Scholar : PubMed/NCBI
|
|
291
|
Sun B, Liu C, Li H, Zhang L, Luo G, Liang
S and Lü M: Research progress on the interactions between long
non-coding RNAs and microRNAs in human cancer. Oncol Lett.
19:595–605. 2020.PubMed/NCBI
|
|
292
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
293
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
294
|
Mishra K and Kanduri C: Understanding long
noncoding RNA and chromatin interactions: What we know so far.
Noncoding RNA. 5:542019.
|
|
295
|
Tasharrofi B and Ghafouri-Fard S: Long
non-coding RNAs as regulators of the mitogen-activated protein
kinase (MAPK) pathway in cancer. Klin Onkol. 31:95–102. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
296
|
Young TL, Matsuda T and Cepko CL: The
noncoding RNA taurine upregulated gene 1 is required for
differentiation of the murine retina. Curr Biol. 15:501–512. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
297
|
Santoro M, Nociti V, Lucchini M, De Fino
C, Losavio FA and Mirabella M: Expression profile of long
non-coding RNAs in serum of patients with multiple sclerosis. J Mol
Neurosci. 59:18–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
298
|
Zang XJ, Li L, Du X, Yang B and Mei CL:
LncRNA TUG1 inhibits the proliferation and fibrosis of mesangial
cells in diabetic nephropathy via inhibiting the PI3K/AKT pathway.
Eur Rev Med Pharmacol Sci. 23:7519–7525. 2019.PubMed/NCBI
|
|
299
|
Li SY and Susztak K: The long noncoding
RNA tug1 connects metabolic changes with kidney disease in
podocytes. J Clin Invest. 126:4072–4075. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
300
|
Gu W, Yuan Y, Wang L, Yang H, Li S, Tang Z
and Li Q: Long non-coding RNA TUG1 promotes airway remodelling by
suppressing the miR-145-5p/DUSP6 axis in cigarette smoke-induced
COPD. J Cell Mol Med. 23:7200–7209. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
301
|
Long J, Badal SS, Ye Z, Wang Y, Ayanga BA,
Galvan DL, Green NH, Chang BH, Overbeek PA and Danesh FR: Long
noncoding RNA Tug1 regulates mitochondrial bioenergetics in
diabetic nephropathy. J Clin Invest. 126:4205–4218. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
302
|
Yin DD, Zhang EB, You LH, Wang N, Wang LT,
Jin FY, Zhu YN, Cao LH, Yuan QX, De W and Tang W: Downregulation of
lncRNA TUG1 affects apoptosis and insulin secretion in mouse
pancreatic β cells. Cell Physiol Biochem. 35:1892–1904. 2015.
View Article : Google Scholar
|
|
303
|
Li Z, Shen J, Chan MTV and Wu WKK: TUG1: A
pivotal onco-genic long non-coding RNA of human cancers. Cell
Prolif. 49:471–475. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
304
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
305
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
306
|
Li B, Shen S, Zhang W, Qi T, Hu Q and
Cheng Y: Long non-coding RNA TUG1 as a potential novel biomarker
for predicting the clinical outcome of cancer patients: A
meta-analysis. Clin Lab. 64:2018. View Article : Google Scholar
|
|
307
|
van Heesch S, van Iterson M, Jacobi J,
Boymans S, Essers PB, de Bruijn E, Hao W, MacInnes AW, Cuppen E and
Simonis M: Extensive localization of long noncoding RNAs to the
cytosol and mono- and polyribosomal complexes. Genome Biol.
15:R62014. View Article : Google Scholar : PubMed/NCBI
|
|
308
|
Long J, Menggen Q, Wuren Q, Shi Q and Pi
X: Long noncoding RNA taurine-upregulated gene1 (TUG1) promotes
tumor growth and metastasis through
TUG1/Mir-129-5p/astrocyte-elevated gene-1 (AEG-1) axis in malignant
melanoma. Med Sci Monit. 24:1547–1559. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
309
|
Han Y, Liu Y, Gui Y and Cai Z: Long
intergenic non-coding RNA TUG1 is overexpressed in urothelial
carcinoma of the bladder. J Surg Oncol. 107:555–559. 2013.
View Article : Google Scholar
|
|
310
|
Zhang Q, Geng PL, Yin P, Wang XL, Jia JP
and Yao J: Down-regulation of long non-coding RNA TUG1 inhibits
osteo-sarcoma cell proliferation and promotes apoptosis. Asian Pac
J Cancer Prev. 14:2311–2315. 2013. View Article : Google Scholar
|
|
311
|
Huang MD, Chen WM, Qi FZ, Sun M, Xu TP, Ma
P and Shu YQ: Long non-coding RNA TUG1 is up-regulated in
hepatocellular carcinoma and promotes cell growth and apoptosis by
epigeneti-cally silencing of KLF2. Mol Cancer. 14:1652015.
View Article : Google Scholar
|
|
312
|
Zhai HY, Sui MH, Yu X, Qu Z, Hu JC, Sun
HQ, Zheng HT, Zhou K and Jiang LX: Overexpression of long
non-coding RNA TUG1 promotes colon cancer progression. Med Sci
Monit. 22:3281–3287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
313
|
Li J, Zhang M, An G and Ma Q: LncRNA TUG1
acts as a tumor suppressor in human glioma by promoting cell
apoptosis. Exp Biol Med (Maywood). 241:644–649. 2016. View Article : Google Scholar
|
|
314
|
Zhang EB, Yin DD, Sun M, Kong R, Liu XH,
You LH, Han L, Xia R, Wang KM, Yang JS, et al: P53-regulated long
non-coding RNA TUG1 affects cell proliferation in human non-small
cell lung cancer, partly through epigenetically regulating HOXB7
expression. Cell Death Dis. 5:e12432014. View Article : Google Scholar : PubMed/NCBI
|
|
315
|
Kondo Y, Shinjo K and Katsushima K: Long
non-coding RNAs as an epigenetic regulator in human cancers. Cancer
Sci. 108:1927–1933. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
316
|
Kim KH and Roberts CWM: Targeting EZH2 in
cancer. Nat Med. 22:128–134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
317
|
Sun CC, Li SJ, Li G, Hua RX, Zhou XH and
Li DJ: Long intergenic noncoding RNA 00511 acts as an oncogene in
non-small-cell lung cancer by binding to EZH2 and suppressing p57.
Mol Ther Nucleic Acids. 5:e3852016. View Article : Google Scholar : PubMed/NCBI
|
|
318
|
Cao R, Wang L, Wang H, Xia L,
Erdjument-Bromage H, Tempst P, Jones RS and Zhang Y: Role of
histone H3 lysine 27 methylation in polycomb-group silencing.
Science. 298:1039–1043. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
319
|
Liu H, Zhou G, Fu X, Cui H, Pu G, Xiao Y,
Sun W, Dong X, Zhang L, Cao S, et al: Long noncoding RNA TUG1 is a
diagnostic factor in lung adenocarcinoma and suppresses apoptosis
via epigenetic silencing of BAX. Oncotarget. 8:101899–101910. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
320
|
Zhang E, He X, Yin D, Han L, Qiu M, Xu T,
Xia R, Xu L, Yin R and De W: Increased expression of long noncoding
RNA TUG1 predicts a poor prognosis of gastric cancer and regulates
cell proliferation by epigenetically silencing of p57. Cell Death
Dis. 7:e21092016. View Article : Google Scholar : PubMed/NCBI
|
|
321
|
Ding B, Lou W, Xu L and Fan W: Non-coding
RNA in drug resistance of hepatocellular carcinoma. Biosci Rep.
38:BSR201809152018. View Article : Google Scholar : PubMed/NCBI
|
|
322
|
Yang L, Lin C, Liu W, Zhang J, Ohgi KA,
Grinstein JD, Dorrestein PC and Rosenfeld MG: ncRNA- and Pc2
methylation-dependent gene relocation between nuclear structures
mediates gene activation programs. Cell. 147:773–788. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
323
|
Thomson DW and Dinger ME: Endogenous
microRNA sponges: Evidence and controversy. Nat Rev Genet.
17:272–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
324
|
Ma L, Bajic VB and Zhang Z: On the
classification of long non-coding RNAs. RNA Biol. 10:925–933. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
325
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
326
|
Tsai KW, Wu CW, Hu LY, Li SC, Liao YL, Lai
CH, Kao HW, Fang WL, Huang KH, Chan WC and Lin WC: Epigenetic
regulation of miR-34b and miR-129 expression in gastric cancer. Int
J Cancer. 129:2600–2610. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
327
|
Ji TT, Huang X, Jin J, Pan SH and Zhuge
XJ: Inhibition of long non-coding RNA TUG1 on gastric cancer cell
transference and invasion through regulating and controlling the
expression of miR-144/c-met axis. Asian Pac J Trop Med. 9:508–512.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
328
|
Zhang M, Huang S and Long D: MiR-381
inhibits migration and invasion in human gastric carcinoma through
downregulating SOX4. Oncol Lett. 14:3760–3766. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
329
|
Li J, Zhang Q, Fan X, Mo W, Dai W, Feng J,
Wu L, Liu T, Li S, Xu S, et al: The long noncoding RNA TUG1 acts as
a competing endogenous RNA to regulate the Hedgehog pathway by
targeting miR-132 in hepatocellular carcinoma. Oncotarget.
8:65932–65945. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
330
|
Sun J, Hu J, Wang G, Yang Z, Zhao C, Zhang
X and Wang J: LncRNA TUG1 promoted KIAA1199 expression via miR-600
to accelerate cell metastasis and epithelial-mesenchymal transition
in colorectal cancer. J Exp Clin Cancer Res. 37:1062018. View Article : Google Scholar : PubMed/NCBI
|
|
331
|
Ma F, Wang S, Cai Q, Jin L, Zhou D, Ding J
and Quan ZW: Long non-coding RNA TUG1 promotes cell proliferation
and metastasis by negatively regulating miR-300 in gallbladder
carcinoma. Biomed Pharmacother. 88:863–869. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
332
|
Tan J, Qiu K, Li M and Liang Y:
Double-negative feedback loop between long non-coding RNA TUG1 and
miR-145 promotes epithelial to mesenchymal transition and
radioresistance in human bladder cancer cells. FEBS Lett. 589(20 Pt
B): 3175–3181. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
333
|
Cai H, Xue Y, Wang P, Wang Z, Li Z, Hu Y,
Li Z, Shang X and Liu Y: The long noncoding RNA TUG1 regulates
blood-tumor barrier permeability by targeting miR-144. Oncotarget.
6:19759–19779. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
334
|
Xie CH, Cao YM, Huang Y, Shi QW, Guo JH,
Fan ZW, Li JG, Chen BW and Wu BY: Long non-coding RNA TUG1
contributes to tumorigenesis of human osteosarcoma by sponging
miR-9-5p and regulating POU2F1 expression. Tumor Biol.
37:15031–15041. 2016. View Article : Google Scholar
|
|
335
|
Dong R, Liu GB, Liu BH, Chen G, Li K,
Zheng S and Dong KR: Targeting long non-coding RNA-TUG1 inhibits
tumor growth and angiogenesis in hepatoblastoma. Cell Death Dis.
7:e22782016. View Article : Google Scholar : PubMed/NCBI
|
|
336
|
Cai H, Liu X, Zheng J, Xue Y, Ma J, Li Z,
Xi Z, Li Z, Bao M and Liu Y: Long non-coding RNA taurine
upregulated 1 enhances tumor-induced angiogenesis through
inhibiting microRNA-299 in human glioblastoma. Oncogene.
36:318–331. 2017. View Article : Google Scholar
|
|
337
|
Hu Y, Sun X, Mao C, Guo G, Ye S, Xu J, Zou
R, Chen J, Wang L, Duan P and Xue X: Upregulation of long noncoding
RNA TUG1 promotes cervical cancer cell proliferation and migration.
Cancer Med. 6:471–482. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
338
|
Katsushima K, Natsume A, Ohka F, Shinjo K,
Hatanaka A, Ichimura N, Sato S, Takahashi S, Kimura H, Totoki Y, et
al: Targeting the Notch-regulated non-coding RNA TUG1 for glioma
treatment. Nat Commun. 7:136162016. View Article : Google Scholar : PubMed/NCBI
|
|
339
|
Yun-Bo F, Xiao-Po L, Xiao-Li L, Guo-Long
C, Pei Z and Fa-Ming T: LncRNA TUG1 is upregulated and promotes
cell proliferation in osteosarcoma. Open Med (Wars). 11:163–167.
2016. View Article : Google Scholar
|
|
340
|
Liang S, Zhang S, Wang P, Yang C, Shang C,
Yang J and Wang J: LncRNA, TUG1 regulates the oral squamous cell
carcinoma progression possibly via interacting with Wnt/β-catenin
signaling. Gene. 608:49–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
341
|
Tang T, Cheng Y, She Q, Jiang Y, Chen Y,
Yang W and Li Y: Long non-coding RNA TUG1 sponges miR-197 to
enhance cisplatin sensitivity in triple negative breast cancer.
Biomed Pharmacother. 107:338–346. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
342
|
Xiao CH, Yu HZ, Guo CY, Wu ZM, Cao HY, Li
WB and Yuan JF: Long non-coding RNA TUG1 promotes the proliferation
of colorectal cancer cells through regulating Wnt/β-catenin
pathway. Oncol Lett. 16:5317–5324. 2018.PubMed/NCBI
|
|
343
|
Qin CF and Zhao FL: Long non-coding RNA
TUG1 can promote proliferation and migration of pancreatic cancer
via EMT pathway. Eur Rev Med Pharmacol Sci. 21:2377–2384.
2017.PubMed/NCBI
|
|
344
|
Zhang CG, Yin DD, Sun SY and Han L: The
use of lncRNA analysis for stratification management of prognostic
risk in patients with NSCLC. Eur Rev Med Pharmacol Sci. 21:115–119.
2017.PubMed/NCBI
|
|
345
|
Lin PC, Huang HD, Chang CC, Chang YS, Yen
JC, Lee CC, Chang WH, Liu TC and Chang JG: Long noncoding RNA TUG1
is downregulated in non-small cell lung cancer and can regulate
CELF1 on binding to PRC2. BMC Cancer. 16:5832016. View Article : Google Scholar : PubMed/NCBI
|
|
346
|
Baratieh Z, Khalaj Z, Honardoost MA,
Emadi-Baygi M, Khanahmad H, Salehi M and Nikpour P: Aberrant
expression of PlncRNA-1 and TUG1: Potential biomarkers for gastric
cancer diagnosis and clinically monitoring cancer progression.
Biomark Med. 11:1077–1090. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
347
|
Ren K, Li Z, Li Y, Zhang W and Han X: Long
noncoding RNA taurine-upregulated gene 1 promotes cell
proliferation and invasion in gastric cancer via negatively
modulating miRNA-145-5p. Oncol Res. 25:789–798. 2017. View Article : Google Scholar
|
|
348
|
Lin YH, Wu MH, Huang YH, Yeh CT and Lin
KH: TUG1 is a regulator of AFP and serves as prognostic marker in
non-hepatitis B non-hepatitis C hepatocellular carcinoma. Cells.
9:2622020. View Article : Google Scholar :
|
|
349
|
Dai Q, Deng J, Zhou J, Wang Z, Yuan X, Pan
S and Zhang HB: Long non-coding RNA TUG1 promotes cell progression
in hepatocellular carcinoma via regulating miR-216b-5p/DLX2 axis.
Cancer Cell Int. 20:82020. View Article : Google Scholar : PubMed/NCBI
|
|
350
|
Fang T, Fang Y, Xu X, He M, Zhao Z, Huang
P, Yuan F, Guo M, Yang B and Xia J: Actinidia chinensis planch root
extract attenuates proliferation and metastasis of hepatocellular
carcinoma by inhibiting epithelial-mesenchymal transition. J
Ethnopharmacol. 231:474–485. 2019. View Article : Google Scholar
|
|
351
|
Huang J, Lu D, Xiang T, Wu X, Ge S, Wang
Y, Wang J and Cheng N: MicroRNA-132-3p regulates cell
proliferation, apop-tosis, migration and invasion of liver cancer
by targeting Sox4. Oncol Lett. 19:3173–3180. 2020.PubMed/NCBI
|
|
352
|
Lv J, Kong Y, Gao Z, Liu Y, Zhu P and Yu
Z: LncRNA TUG1 interacting with miR-144 contributes to
proliferation, migration and tumorigenesis through activating the
JAK2/STAT3 pathway in hepatocellular carcinoma. Int J Biochem Cell
Biol. 101:19–28. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
353
|
He C, Liu Z, Jin L, Zhang F, Peng X, Xiao
Y, Wang X, Lyu Q and Cai XJ: lncRNA TUG1-mediated mir-142-3p
downregulation contributes to metastasis and the
epithelial-to-mesenchymal transition of hepatocellular carcinoma by
targeting ZEB1. Cell Physiol Biochem. 48:1928–1941. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
354
|
Xie F, Zhang L, Yao Q, Shan L, Liu J, Dong
N and Liang J: TUG1 promoted tumor progression by sponging
miR-335-5p and regulating CXCR4-mediated infiltration of pro-tumor
immunocytes in CTNNB1-mutated hepatoblastoma. Onco Targets Ther.
13:3105–3115. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
355
|
An N and Cheng D: The long noncoding RNA
HOST2 promotes gemcitabine resistance in human pancreatic cancer
cells. Pathol Oncol Res. 26:425–431. 2020. View Article : Google Scholar
|
|
356
|
Zhao L, Sun H, Kong H, Chen Z, Chen B and
Zhou M: The lncrna-TUG1/EZH2 axis promotes pancreatic cancer cell
proliferation, migration and EMT phenotype formation through
sponging mir-382. Cell Physiol Biochem. 42:2145–2158. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
357
|
Xu K and Zhang L: Inhibition of
TUG1/miRNA-299-3p axis represses pancreatic cancer malignant
progression via suppression of the notch1 pathway. Dig Dis Sci.
65:1748–1760. 2020. View Article : Google Scholar
|
|
358
|
Yang F and Li X, Zhang L, Cheng L and Li
X: LncRNA TUG1 promoted viability and associated with gemcitabine
resistant in pancreatic ductal adenocarcinoma. J Pharmacol Sci.
137:116–121. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
359
|
Burris HA III, Moore MJ, Andersen J, Green
MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo
AM, Tarassoff P, et al: Improvements in survival and clinical
benefit with gemcitabine as first-line therapy for patients with
advanced pancreas cancer: A randomized trial. J Clin Oncol.
15:2403–2413. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
360
|
Lu Y, Tang L, Zhang Z, Li S, Liang S, Ji
L, Yang B, Liu Y and Wei W: Long noncoding RNA TUG1/miR-29c axis
affects cell proliferation, invasion, and migration in human
pancreatic cancer. Dis Markers. 2018:68570422018. View Article : Google Scholar :
|
|
361
|
Miyamoto Y, Maitra A, Ghosh B, Zechner U,
Argani P, Iacobuzio-Donahue CA, Sriuranpong V, Iso T, Meszoely IM,
Wolfe MS, et al: Notch mediates TGFα-induced changes in epithelial
differentiation during pancreatic tumorigenesis. Cancer Cell.
3:565–576. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
362
|
Hu H, Zhou L, Awadallah A and Xin W:
Significance of Notch1-signaling pathway in human pancreatic
development and carcinogenesis. Appl Immunohistochem Mol Morphol.
21:242–247. 2013.
|
|
363
|
Chen X, Qi M, Yang Q and Li JY: MiR-299-3p
functions as a tumor suppressor in thyroid cancer by regulating
SHOC2. Eur Rev Med Pharmacol Sci. 23:232–240. 2019.PubMed/NCBI
|
|
364
|
Dang S, Zhou J, Wang Z, Wang K, Dai S and
He S: MiR-299-3p functions as a tumor suppressor via targeting
Sirtuin 5 in hepato-cellular carcinoma. Biomed Pharmacother.
106:966–975. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
365
|
Wang JY, Jiang JB, Li Y, Wang YL and Dai
Y: MicroRNA-299-3p suppresses proliferation and invasion by
targeting VEGFA in human colon carcinoma. Biomed Pharmacother.
93:1047–1054. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
366
|
Hui B, Xu Y, Zhao B, Ji H, Ma Z, Xu S, He
ZY, Wang K and Lu J: Overexpressed long noncoding RNA TUG1 affects
the cell cycle, proliferation, and apoptosis of pancreatic cancer
partly through suppressing RND3 and MT2A. Onco Targets Ther.
12:1043–1057. 2019. View Article : Google Scholar :
|
|
367
|
Zhu Y, Zhou J, Xia H, Chen X, Qiu M, Huang
J, Liu S, Tang Q, Lang N, Liu Z, et al: The Rho GTPase RhoE is a
p53-regulated candidate tumor suppressor in cancer cells. Int J
Oncol. 44:896–904. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
368
|
Poch E, Miñambres R, Mocholí E, Ivorra C,
Pérez-Aragó A, Guerri C, Pérez-Roger I and Guasch RM: RhoE
interferes with Rb inactivation and regulates the proliferation and
survival of the U87 human glioblastoma cell line. Exp Cell Res.
313:719–731. 2007. View Article : Google Scholar
|
|
369
|
Wang L, Zhao Z, Feng W, Ye Z, Dai W, Zhang
C, Peng J and Wu K: Long non-coding RNA TUG1 promotes colorectal
cancer metastasis via EMT pathway. Oncotarget. 7:51713–51719. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
370
|
Shen X, Hu X, Mao J, Wu Y, Liu H, Shen J,
Yu J and Chen W: The long noncoding RNA TUG1 is required for
TGF-β/TWIST1/EMT-mediated metastasis in colorectal cancer cells.
Cell Death Dis. 11:652020. View Article : Google Scholar
|
|
371
|
Sun J, Ding C, Yang Z, Liu T, Zhang X,
Zhao C and Wang J: The long non-coding RNA TUG1 indicates a poor
prognosis for colorectal cancer and promotes metastasis by
affecting epithelial-mesenchymal transition. J Transl Med.
14:422016. View Article : Google Scholar : PubMed/NCBI
|
|
372
|
Fan S, Yang Z, Ke Z, Huang K, Liu N, Fang
X and Wang K: Downregulation of the long non-coding RNA TUG1 is
associated with cell proliferation, migration, and invasion in
breast cancer. Biomed Pharmacother. 95:1636–1643. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
373
|
Caldon CE, Daly RJ, Sutherland RL and
Musgrove EA: Cell cycle control in breast cancer cells. J Cell
Biochem. 97:261–274. 2006. View Article : Google Scholar
|
|
374
|
Zhao X and Ren G: LncRNA
taurine-upregulated gene 1 promotes cell proliferation by
inhibiting microRNA-9 in MCF-7 cells. J Breast Cancer. 19:349–357.
2016. View Article : Google Scholar
|
|
375
|
Gradia DF, Mathias C, Coutinho R, Cavalli
IJ, Ribeiro EMSF and de Oliveira JC: Long non-coding RNA TUG1
expression is associated with different subtypes in human breast
cancer. Noncoding RNA. 3:262017.
|
|
376
|
Neuwelt EA, Barnett PA, Bigner DD and
Frenkel EP: Effects of adrenal cortical steroids and osmotic
blood-brain barrier opening on methotrexate delivery to gliomas in
the rodent: The factor of the blood-brain barrier. Proc Natl Acad
Sci USA. 79:4420–4423. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
377
|
Li TH, Zhang JJ, Liu SX and Chen Y: Long
non-coding RNA taurine-upregulated gene 1 predicts unfavorable
prognosis, promotes cells proliferation, and inhibits cells
apoptosis in epithelial ovarian cancer. Medicine (Baltimore).
97:e05752018. View Article : Google Scholar
|
|
378
|
Iliev R, Kleinova R, Juracek J, Dolezel J,
Ozanova Z, Fedorko M, Pacik D, Svoboda M, Stanik M and Slaby O:
Overexpression of long non-coding RNA TUG1 predicts poor prognosis
and promotes cancer cell proliferation and migration in high-grade
muscle-invasive bladder cancer. Tumour Biol. 37:13385–13390. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
379
|
Guo P, Zhang G, Meng J, He Q, Li Z and
Guan Y: Upregulation of long noncoding RNA TUG1 promotes bladder
cancer cell proliferation, migration, and invasion by inhibiting
miR-29c. Oncol Res. 26:1083–1091. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
380
|
Liu Q, Liu H, Cheng H, Li Y, Li X and Zhu
C: Downregulation of long noncoding RNA TUG1 inhibits proliferation
and induces apoptosis through the TUG1/miR-142/ZEB2 axis in bladder
cancer cells. Onco Targets Ther. 10:2461–2471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
381
|
Jiang H, Hu X, Zhang H and Li W:
Down-regulation of LncRNA TUG1 enhances radiosensitivity in bladder
cancer via suppressing HMGB1 expression. Radiat Oncol. 12:652017.
View Article : Google Scholar : PubMed/NCBI
|
|
382
|
Zhu J, Shi H, Liu H, Wang X and Li F: Long
non-coding RNA TUG1 promotes cervical cancer progression by
regulating the miR-138-5p-SIRT1 axis. Oncotarget. 8:65253–65264.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
383
|
Fan M, Li C, He P, Fu Y, Li M and Zhao X:
Knockdown of long noncoding RNA-taurine-upregulated gene 1 inhibits
tumor angiogenesis in ovarian cancer by regulating leucine-rich
α-2-glycoprotein-1. Anticancer Drugs. 30:562–570. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
384
|
Wang X, Abraham S, McKenzie JAG, Jeffs N,
Swire M, Tripathi VB, Luhmann UFO, Lange CAK, Zhai Z, Arthur HM, et
al: LRG1 promotes angiogenesis by modulating endothelial TGF-β
signalling. Nature. 499:306–311. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
385
|
Goel S, Duda DG, Xu L, Munn LL, Boucher Y,
Fukumura D and Jain RK: Normalization of the vasculature for
treatment of cancer and other diseases. Physiol Rev. 91:1071–1121.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
386
|
Meng H, Song Y, Zhu J, Liu Q, Lu P, Ye N,
Zhang Z, Pang Y, Qi J and Wu H: LRG1 promotes angiogenesis through
upregulating the TGF-β1 pathway in ischemic rat brain. Mol Med Rep.
14:5535–5543. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
387
|
Zhang J, Zhu L, Fang J, Ge Z and Li X:
LRG1 modulates epithelial-mesenchymal transition and angiogenesis
in colorectal cancer via HIF-1α activation. J Exp Clin Cancer Res.
35:292016. View Article : Google Scholar
|
|
388
|
Baek YY, Cho DH, Choe J, Lee H, Jeoung D,
Ha KS, Won MH, Kwon YG and Kim YM: Extracellular taurine induces
angio-genesis by activating ERK-, Akt-, and FAK-dependent signal
pathways. Eur J Pharmacol. 674:188–199. 2012. View Article : Google Scholar
|
|
389
|
Xu T, Liu CL, Li T, Zhang YH and Zhao YH:
LncRNA TUG1 aggravates the progression of prostate cancer and
predicts the poor prognosis. Eur Rev Med Pharmacol Sci.
23:4698–4705. 2019.PubMed/NCBI
|
|
390
|
Hao SD, Ma JX, Liu Y, Liu PJ and Qin Y:
Long non-coding TUG1 accelerates prostate cancer progression
through regulating miR-128-3p/YES1 axis. Eur Rev Med Pharmacol Sci.
24:619–632. 2020.PubMed/NCBI
|
|
391
|
Jiang L, Wang W, Li G, Sun C, Ren Z, Sheng
H, Gao H, Wang C and Yu H: High TUG1 expression is associated with
chemotherapy resistance and poor prognosis in esophageal squamous
cell carcinoma. Cancer Chemother Pharmacol. 78:333–339. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
392
|
Xu Y, Wang J, Qiu M and Xu L, Li M, Jiang
F, Yin R and Xu L: Upregulation of the long noncoding RNA TUG1
promotes proliferation and migration of esophageal squamous cell
carcinoma. Tumor Biol. 36:1643–1651. 2015. View Article : Google Scholar
|
|
393
|
Tang Y, Yang P, Zhu Y and Su Y: LncRNA
TUG1 contributes to ESCC progression via regulating miR-148a-3p/
MCL-1/Wnt/β-catenin axis in vitro. Thorac Cancer. 11:82–94. 2020.
View Article : Google Scholar
|
|
394
|
Ma B, Li M, Zhang L, Huang M, Lei JB, Fu
GH, Liu CX, Lai QW, Chen QQ and Wang YL: Upregulation of long
non-coding RNA TUG1 correlates with poor prognosis and disease
status in osteosarcoma. Tumor Biol. 37:4445–4455. 2016. View Article : Google Scholar
|
|
395
|
Wang H, Yu Y, Fan S and Luo L: Knockdown
of Long noncoding RNA TUG1 inhibits the proliferation and cellular
invasion of osteosarcoma cells by sponging miR-153. Oncol Res.
26:665–673. 2018. View Article : Google Scholar
|
|
396
|
Xie C, Chen B, Wu B, Guo J and Cao Y:
LncRNA TUG1 promotes cell proliferation and suppresses apoptosis in
osteosarcoma by regulating miR-212-3p/FOXA1 axis. Biomed
Pharmacother. 97:1645–1653. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
397
|
Li G, Liu K and Du X: Long Non-Coding RNA
TUG1 promotes proliferation and inhibits apoptosis of osteosarcoma
cells by sponging miR-132-3p and upregulating SOX4 expression.
Yonsei Med J. 59:226–235. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
398
|
Liu Y, Li Y, Liu J, Wu Y and Zhu Q:
MicroRNA-132 inhibits cell growth and metastasis in osteosarcoma
cell lines possibly by targeting Sox4. Int J Oncol. 47:1672–1684.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
399
|
Wang Y, Yang T, Zhang Z, Lu M, Zhao W,
Zeng X and Zhang W: Long non-coding RNA TUG1 promotes migration and
invasion by acting as a ceRNA of miR-335-5p in osteosarcoma cells.
Cancer Sci. 108:859–867. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
400
|
Li Y, Zhang T, Zhang Y, Zhao X and Wang W:
Targeting the FOXM1-regulated long noncoding RNA TUG1 in
osteosar-coma. Cancer Sci. 109:3093–3104. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
401
|
Cao J, Han X, Qi X, Jin X and Li X: TUG1
promotes osteo-sarcoma tumorigenesis by upregulating EZH2
expression via miR-144-3p. Int J Oncol. 51:1115–1123. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
402
|
Yu X, Hu L, Li S, Shen J, Wang D, Xu R and
Yang H: Long non-coding RNA Taurine upregulated gene 1 promotes
osteo-sarcoma cell metastasis by mediating HIF-1α via miR-143-5p.
Cell Death Dis. 10:2802019. View Article : Google Scholar
|
|
403
|
Li Q, Song W and Wang J: TUG1 confers
Adriamycin resistance in acute myeloid leukemia by epigenetically
suppressing miR-34a expression via EZH2. Biomed Pharmacother.
109:1793–1801. 2019. View Article : Google Scholar
|
|
404
|
Isin M, Ozgur E, Cetin G, Erten N, Aktan
M, Gezer U and Dalay N: Investigation of circulating lncRNAs in
B-cell neoplasms. Clin Chim Acta. 431:255–259. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
405
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
406
|
Yang L, Moss T, Mangala LS, Marini J, Zhao
H, Wahlig S, Armaiz-Pena G, Jiang D, Achreja A, Win J, et al:
Metabolic shifts toward glutamine regulate tumor growth, invasion
and bioenergetics in ovarian cancer. Mol Syst Biol. 10:7282014.
View Article : Google Scholar : PubMed/NCBI
|
|
407
|
Wang J, Xie H, Ling Q, Lu D, Lv Z, Zhuang
R, Liu Z, Wei X, Zhou L, Xu X and Zheng S: Coding-noncoding gene
expression in intrahepatic cholangiocarcinoma. Transl Res.
168:107–121. 2016. View Article : Google Scholar
|
|
408
|
Zeng B, Ye H, Chen J, Cheng D, Cai C, Chen
G, Chen X, Xin H, Tang C and Zeng J: LncRNA TUG1 sponges miR-145 to
promote cancer progression and regulate glutamine metabolism via
Sirt3/GDH axis. Oncotarget. 8:113650–113661. 2017. View Article : Google Scholar
|
|
409
|
Han X and Yang Y, Sun Y, Qin L and Yang Y:
LncRNA TUG1 affects cell viability by regulating glycolysis in
osteosarcoma cells. Gene. 674:87–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
410
|
Kunej T, Obsteter J, Pogacar Z, Horvat S
and Calin GA: The decalog of long non-coding RNA involvement in
cancer diagnosis and monitoring. Crit Rev Clin Lab Sci. 51:344–357.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
411
|
Dutour A, Leclers D, Monteil J, Paraf F,
Charissoux JL, Rousseau R and Rigaud M: Non-invasive imaging
correlates with histological and molecular characteristics of an
osteosar-coma model: Application for early detection and follow-up
of MDR phenotype. Anticancer Res. 27(6B): 4171–4178. 2007.
|
|
412
|
Fidler IJ: The biology of brain
metastasis: Challenges for therapy. Cancer J. 21:284–293. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
413
|
La Porta CA: Drug resistance in melanoma:
New perspectives. Curr Med Chem. 14:387–391. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
414
|
Liu YY, Han TY, Giuliano AE and Cabot MC:
Ceramide glyco-sylation potentiates cellular multidrug resistance.
FASEB J. 15:719–730. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
415
|
Synold TW, Dussault I and Forman BM: The
orphan nuclear receptor SXR coordinately regulates drug metabolism
and efflux. Nat Med. 7:584–590. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
416
|
Gottesman MM: Mechanisms of cancer drug
resistance. Annu Rev Med. 53:615–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
417
|
Liu Z and Zhang H: LncRNA plasmacytoma
variant trans-location 1 is an oncogene in bladder urothelial
carcinoma. Oncotarget. 8:64273–64282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
418
|
Xie D, Zhang H, Hu X and Shang C:
Knockdown of long non-coding RNA Taurine Up-Regulated 1 inhibited
doxorubicin resistance of bladder urothelial carcinoma via
Wnt/β-catenin pathway. Oncotarget. 8:88689–88696. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
419
|
Li C, Gao Y, Li Y and Ding D: TUG1
mediates methotrexate resistance in colorectal cancer via
miR-186/CPEB2 axis. Biochem Biophys Res Commun. 491:552–557. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
420
|
Gu L, Li Q, Liu H, Lu X and Zhu M: Long
noncoding RNA TUG1 promotes autophagy-associated paclitaxel
resistance by sponging miR-29b-3p in ovarian cancer cells.
OncoTargets Ther. 13:2007–2019. 2020. View Article : Google Scholar
|
|
421
|
Xi G, Hu X, Wu B, Jiang H, Young CY, Pang
Y and Yuan H: Autophagy inhibition promotes paclitaxel-induced
apoptosis in cancer cells. Cancer Lett. 307:141–148. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
422
|
Wei X, Zhou Y, Qiu J, Wang X, Xia Y and
Sui L: Low expression of TUG1 promotes cisplatin sensitivity in
cervical cancer by activating the MAPK pathway. J BUON.
24:1020–1026. 2019.PubMed/NCBI
|
|
423
|
Köberle B, Tomicic MT, Usanova S and Kaina
B: Cisplatin resistance: Preclinical findings and clinical
implications. Biochim Biophys Acta. 1806:172–182. 2010.PubMed/NCBI
|
|
424
|
Niu Y, Ma F, Huang W, Fang S, Li M, Wei T
and Guo L: Long non-coding RNA TUG1 is involved in cell growth and
chemo-resistance of small cell lung cancer by regulating LIMK2b via
EZH2. Mol Cancer. 16:52017. View Article : Google Scholar
|
|
425
|
Xu C, Guo Y, Liu H, Chen G, Yan Y and Liu
T: TUG1 confers cisplatin resistance in esophageal squamous cell
carcinoma by epigenetically suppressing PDCD4 expression via EZH2.
Cell Biosci. 8:612018. View Article : Google Scholar : PubMed/NCBI
|
|
426
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
427
|
Liu Q, Sun S, Yu W, Jiang J, Zhuo F, Qiu
G, Xu S and Jiang X: Altered expression of long non-coding RNAs
during genotoxic stress-induced cell death in human glioma cells. J
Neurooncol. 122:283–292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
428
|
Wang X, Chen X, Zhang D, Yang G, Yang Z,
Yin Z and Zhao S: Prognostic and clinicopathological role of long
non-coding RNA taurine upregulated 1 in various human malignancies:
A systemic review and meta-analysis. Tumor Biol.
39:10104283177143612017. View Article : Google Scholar
|