1. Structure and function of primary
cilia
The primary cilium is a feature of almost all cell
types, most commonly of epithelial cells. Primary cilia are
cellular non-motile structures, composed of two centrioles, that
extend from the basal body in the apical membrane. The cytoskeleton
of the cilium, also referred to as axoneme, is composed of 9
post-translationally acetylated microtubule doublets in a 9+0
arrangement, lacking the central microtubule pair as in motile
cilia (1,2). Although all types of cilia have the
same microtubule-based structure, their roles differ. Indeed,
motile cilia are responsible for processes including cerebrospinal
fluid flow and mucus clearance, whereas non-motile primary cilia
have evolved in sensory organelles. Primary cilia receive numerous
extracellular signals, such as mechanical stimuli, light and
low-molecular-weight molecules (1-3). In
addition to its structural role as major microtubule organization
centre in mammalian cells, the basal body is also responsible for
protein trafficking through the cilium. This protein exchange is
regulated at the ciliary base comprising Y-linker proteins
(transition zone, Y-ankles and ciliary gate). These linkers
protrude from the outer microtubule pairs present in the transition
zone to the ciliary membrane. The majority of the proteins located
in the ciliary membrane have yet to be investigated for their
functions; however, it is known that the ciliary membrane
encompasses several receptor proteins, ion channels, protein
transporters and sensory proteins. A number of these proteins are
mutated in ciliopathies, i.e., diseases caused by dysregulation of
primary cilia structure and function, such as autosomal dominant
polycystic kidney disease, nephronophthisis and Meckel-Gruber
syndrome (1-3).
2. Assembly and disassembly of primary
cilia: IFT and cell cycle
Two pathways of ciliogenesis have been described by
Sorokin, namely the extracellular and intracellular pathways
(4). In the extracellular pathway,
the mother centriole anchors to the plasma membrane upon nucleation
of axonemal microtubules. As the cilium derived directly from the
mitotic spindle, and is the first cilium to emerge from the cell,
it is named the 'primary cilium'. This type of ciliogenesis has
been observed in all cells, including those that later develop a
ciliated border. On the other hand, in the intracellular pathway,
the axoneme elongation originates from the cytoplasm after
association of the mother centriole with ciliary vesicles derived
from the Golgi apparatus. This type of ciliogenesis has been
observed in epithelial cells with a ciliated border (4). The assembly/disassembly of the
primary cilium and trafficking of intermediary molecules of
signalling pathways are dependent upon intraflagellar transport
(IFT). IFT proceeds in two directions: i) Anterograde transport
from the cilium base to the tip, catalysed by the dynein 2/1b
protein; and ii) retrograde transport from the tip to the cilium
base, catalysed by kinesin-2 family proteins (KIF3A, KIF3B and
KIF17). Proteins are transported to the base of the cilium by
several pathways. This transport step involves IFT20 protein,
exocyst-related small Rab GTPases and the BBSome complex, formed by
seven Bardet-Biedl syndrome (BBS) proteins (BBS1, BBS2, BBS4, BBS5,
BBS7, BBS8 and BBS9) that are located in the basal body and are
involved in trafficking cargo to the primary cilium (5,6).
Jonassen et al identified that mutations in IFT proteins in
murine models resulted in polycystic kidney disease (7).
Furthermore, assembly and disassembly of cilia are
correlated with cell cycle status. As the mother centriole
functions in the mitotic apparatus, it must be released from the
plasma membrane and the primary cilia must disassemble prior to
mitosis. In most cell types, the cilia are disassembled in the M
phase, re-assembled upon entry in the G1 phase, and they continue
to grow in the GO phase (8).
However, Tucker et al have also observed ciliary resorption
prior to the S phase in cells emerging from the quiescent state
(9). Hence, ciliogenesis and its
obliteration are crucial for proliferating cells and several
signalling pathways are responsible for their regulation. The
resorption of primary cilia has been well studied in human RPE1
(telomerase reverse transcriptase-immortalized retinal pigment
epithelial) cell cultures. It has been demonstrated that growth
factor-stimulated cell cultures, previously deprived of serum,
caused ciliary disassembly through the activation of human enhancer
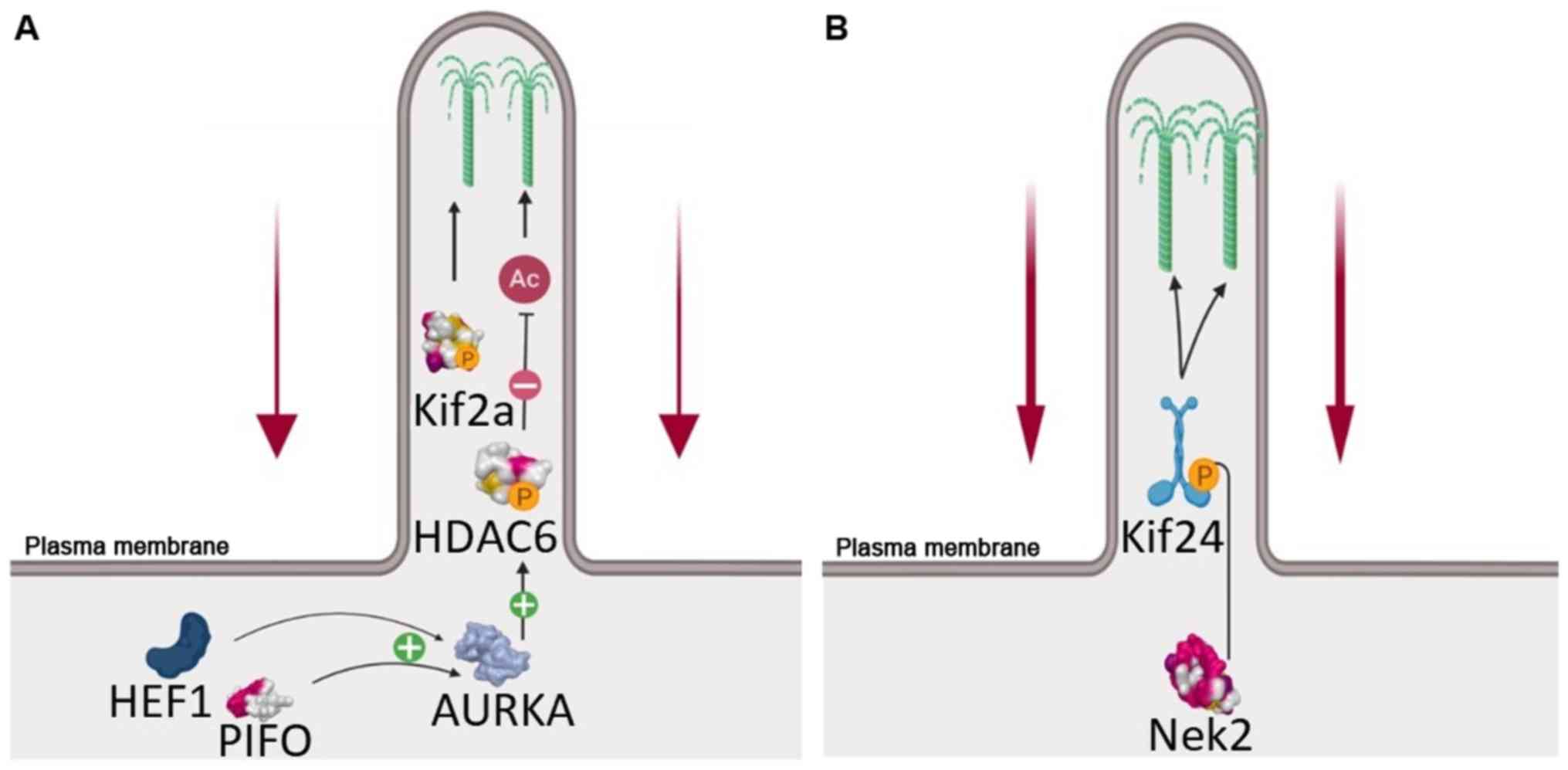
of filamentation 1 (HEF1) and Aurora A kinase (AURKA), which
sequentially activate histone deacetylase (HDAC)6 in the axoneme.
Through deacetylation of axonemal tubules, HDAC6 promotes ciliary
resorption (6). The phosphorylated
KIF2A, a member of the kinesin-13 protein family, exhibits
microtubule depolymerizing activity at the mother centriole,
causing disassembly of the primary cilium coupled to the cell cycle
(6). However, other proteins
involved in this process have recently emerged, including the
Pitchfork (PIFO) and dynein light chain Tctex-type 1 proteins. The
PIFO protein is required specifically to control cilia retraction,
as well as the liberation and duplication of the basal
body/centrosome. It may act by stimulating AURKA activity at the
basal body in a cell cycle-dependent manner. Cytoplasmic dynein 1
acts as a motor for the intracellular retrograde motility of
vesicles and organelles along the microtubules (6,8,10).
The primary cilia exhibit an interesting ability to
adjust their morphology and size to external stimuli. The cilium
can enlarge itself to improve its signal recognition capacity and,
vice versa, the resorption of cilia results in reduced
responsiveness to signals. The key mediators in these processes are
second messengers and IFT. In cell types with apicobasal polarity,
primary cilia are always localized to the apical surface. This is
very important, since the apical membrane is directed towards the
lumen of ducts; therefore, cilia are exposed to various chemical
and mechanical signals (3).
Ciliogenesis appears to be autonomously regulated
within cilium, since IFT is regulated through ciliary cAMP
concentration in response to external signals, but also stimuli
derived from the cell cycle may cause irreversible transition, such
as assembly/disassembly of cilia at the entry and exit from the
quiescent state (3,11). Based on the experimental evidence
obtained to date, it appears that ciliary resorption occurs in two
waves, i) the G1 resorption, regulated mainly by AURKA-HDAC6 and
polo-like kinase 1 (PLK1)-KIF2A loop; and ii) G2/M resorption,
driven by NIMA-related kinase 2 (NEK2)-KIF24 (6). The NEK2 protein is a S/G2 kinase,
which localizes to the distal portion of the mother centriole and
is essential for correct cilium resorption at the G2/M transition
(12). The substrate for this
kinase is KIF24, a kinesin-13 family protein, the phosphorylation
of which stimulates its microtubule-depolymerizing activity and
prevents the extension of cilia in proliferating cells (13). As shown in Fig. 1, the AURKA-HDAC6 and NEK2-KIF24
complexes play distinct, consecutive roles during cilia resorption
as cells re-enter the cell cycle from the quiescent state (6).
3. Primary cilia as mediators of signalling
pathways
The primary cilium serves as a central hub for
numerous signalling pathways implicated in a variety of biological
processes, also known to be dysregulated in pancreatic ductal
adenocarcinoma (PDAC), including the Hedgehog, Wnt and mammalian
target of rapamycin (mTOR) pathways, which have been extensively
investigated (1,2,5).
Since the primary cilium acts like a cellular antenna for several
ligands, it is enriched of all the components, both receptors and
effectors, of the Hedgehog, Wnt and mTOR signalling pathways.
Moreover, the IFT motor proteins allow the trafficking of these
components within the cilium and, therefore, they can modulate the
activity of these signalling pathways (1,2,5).
In numerous tumours characterized by Hedgehog
dysregulation, it is reported that tumour cells lack primary cilia
in comparison with normal neighbouring cells (14,15).
The consequence of the absence of the primary cilium is
context-dependent, as it may lead to the induction or the
suppression of Hedgehog pathway-dependent tumorigenesis (1,2,5). The
role of primary cilia in canonical Wnt signalling remains a subject
of discussion, but it has been acknowledged that normal
ciliogenesis is fundamental for the non-canonical Wnt/planar cell
polarity pathway. Studies on the mTOR pathway in renal cells have
proposed a model of cilium-dependent mTOR signalling. In
particular, it has been reported that flow shear stress causes
cilium bending, which consequently inhibits mTOR complex 1 (mTORCl)
(2,5). Upon cilium bending caused by fluid
flux, the Leishmania protein kinase is transported to the basal
body where another kinase, AMP-activated protein kinase, is
phosphorylated, which, in turn, leads to the activation of the
tuberous sclerosis (TSC)1-TSC2 complex, Rheb and, finally, the
inhibition of mTORC1 (2).
4. Loss of primary cilia and tumour
development
Various genomic and proteomic studies have
identified ~1,600 genes essential for ciliary structure and
function in mammals (16). Loss of
primary cilia has been observed in various tumours, suggesting its
involvement in carcinogenesis through aberrant signalling pathways.
Development of tumours characterized by primary cilia absence is
probably due to alterations in signalling pathways, which likely
lead cells into a hyperproliferative state and promote migration
and invasion (17).
Some initial results have already been proposed; for
example, in cholangiocarcinoma (CCA), ciliary expression was found
to be decreased both in human samples and human cell lines
(17). It has been proposed that
HDAC6 plays an important role in the regulation of ciliogenesis in
CCA. It is believed that CCA and PDAC cells share a common
mechanism of cilia loss, since K-Ras is responsible for HDAC6
activation (8,15,18).
Other tumours, such as prostate, colorectal, breast and melanoma,
similarly display loss of primary cilia, but the underlying
mechanisms remain to be elucidated (19-22).
The reduction in the number of primary cilia has been also observed
in ovarian cancer, renal cancer and glioblastoma. The underlying
mechanism causing reduction of cilia in ovarian cancer is
associated with overexpression and constant localization of AURKA
to the ciliary basal body (23).
HDAC6 plays an important role in the reduction of cilia number in
renal cell carcinoma; in particular, it is activated as a
consequence of an interaction among AURKA, p-catenin and HEF1 upon
binding of von Hippel-Lindau to the microtubules (24).
5. PDAC and cilium
The pancreatic acinar cells undergo metaplastic
acinar-to-ductal metaplasia, which lead to pancreatic
intraepithelial neoplasia (PanIN), which may develop into PDAC. The
majority of the PDAC cases harbour K-Ras gene mutations.
Furthermore, the majority of human PanIN and PDAC lesions are
devoid of primary cilia (25).
The study of Quilichini et al addressed the
function of pancreatic ducts and primary cilia in postnatal
pancreatic tissue homeostasis (26). The authors focused on hepatocyte
nuclear factor (Hnf)1b, a transcription factor that controls
several genes important for pancreatic duct morphogenesis during
development. Pancreatic cells with mutated Hnf1b exhibit a major
decrease in expression of genes involved in cystic disease, the
ductal cells do not have primary cilia and have a 1.4-fold higher
proliferation rate compared with control cells. The effects of
Hnf1b inactivation (loss of primary cilia, duct proliferation and
dilatation) generate fibrosis, inflammatory infiltration,
lipomatosis and activation of epithelial-to-mesenchymal transition,
consequently leading to chronic pancreatitis and PanlNs.
Furthermore, a significant decrease in the expression of polycystic
kidney and hepatic disease 1 (fibrocystin), cystin 1, secreted
phosphoprotein 1, prospero homeobox 1 and IFT88 (Tg737/IFT88) was
observed. These genes are involved in maintenance of the ciliary
and/or tubular architecture (26).
It has been demonstrated that PDAC cells lack
primary cilia, independently of their proliferation status
(15), by studies that have used
human tissue specimens, the PANC-1 cell line and murine models.
Instead, it was more recently demonstrated that primary cilia in
the epithelium are gradually lost during pancreatic carcinogenesis,
and this loss is accompanied by gain of primary cilia in the
surrounding stroma (Table I).
Moreover, the BxPc3, Capan1, MiaPaCa2, and PANC-1 PDAC cell lines
do not lose primary cilia under conditions of nutrient or oxygen
deprivation (27).
 | Table IFraction of pancreatic cells with
primary cilia. |
Table I
Fraction of pancreatic cells with
primary cilia.
| Type of tissue | Type of cell
|
|---|
| Epithelial (%) | Stromal (%) |
|---|
| Normal
pancreas | 15.5-31.8 | 12.0 |
| Chronic
pancreatitis | 20.6 | 25.0 |
| Pancreatic
intraepithelial neoplasia | | |
| 1A | 17.8 | 24.8 |
| 1B | 14.0 | 28.5 |
| 2 | 5.7 | 29.4 |
| 3 | 2.9 | 22.3 |
| Pancreatic ductal
adenocarcinoma (no. of samples) | | |
| 7 | 0.0 | 31.5 |
| 4 | 0.96-2.3 | 31.5 |
| 2 | 4.65-5.85 | 31.5 |
| Intraductal
papillary mucinous neoplasia (no. of samples) | | |
| Benign (4) | 0.0 | 8.3 |
| Malignant
(8) | 0.0 | 8.3 |
At the molecular level, these studies demonstrated
that, by inhibiting the downstream effectors of the K-Ras pathway,
mitogen-activated protein kinase kinase (MEK) and phosphoinositide
3-kinase (PI3K), ciliogenesis was fully restored, suggesting that
K-Ras may be involved in the regulation of ciliogenesis in PDAC
(15,28). Although some studies have
previously reported that AURKA drives primary cilia resorption by
activation of HDAC6, this does not appear to be the case in PDAC.
Indeed, Kobayashi et al, in an attempt to explain how loss
of primary cilia affects PDAC development, revealed that HDAC6 is
not involved in ciliation of PANC-1 cells. Depletion experiments
with siRNA molecules suggested that HDAC2 is responsible for
inhibition of primary ciliogenesis. Similar results have been
obtained from other PDAC cell lines, such as CFPAC1 and KrasPDEC,
suggesting that HDAC2 inhibition is a common characteristic of PDAC
cells (28). HDAC2 is known to be
overexpressed in PDAC lesions in comparison with normal duct
epithelial cells. Furthermore, AURKA was also identified as a
downstream target of K-Ras and HDAC2 in the suppression of
ciliogenesis. It is possible that HDAC2 positively controls
transcription of AURKA, which is overexpressed or amplified in
several types of cancer, including PDAC (29). Moreover, HDAC6 phosphorylated by
AURKA is involved in ciliary axoneme microtubule deacetylation,
leading to disassembly of primary cilia in non-tumour cells.
However, in PDAC models, inhibition of HDAC6 was not efficient
against ciliation and treatment of PANC-1 cells with AURKA
inhibitors led to a significant restoration of primary cilia
(28). However, the role of
primary cilia in PDAC development remains elusive.
AURKA regulates cell cycle entry and progression to
the M phase, as well as ciliary resorption. Additionally, this
protein kinase is involved in the mechanism of axonemal disassembly
during cell exit from the GO phase. When associated with other
proteins, namely HEF1 (NEDD9) and Pitchfork (PIFO), AURKA exhibits
increased catalytic activity and induces HDAC6 phosphorylation.
This reaction may cause ciliary destabilization and resorption.
HEF1 activation of AURKA has been observed after 1-2 h, and after
18-24 h upon treatment with serum, suggesting that the
HEF1-AURKA-HDAC6 loop is a key player in ciliary disassembly, even
in the first phase of resorption (G1 resorption). However, this
phenomenon has yet to be fully elucidated. The role of HDAC6 in
ciliogenesis has yet to be clearly determined due to the opposite
results of studies on mice with HDAC6 mutations who do not present
ciliopathies (8). Furthermore,
other AURKA regulators have been identified, such as calmodulin and
Ca2+. Inositol polyphosphate-5-phosphatase E (INPP5E), a
ciliary lipid phosphatase, removes the 5-phosphate group from
PI(3,4,5)3P
and PI(4,5) P2, and has been correlated to AURKA
activity for the stability of primary cilia. Furthermore, it has
been recognized that INPP5E maintains PI(4,5)P2 at
low or minimum levels in order to ensure correct trafficking of
Hedgehog proteins and, hence, it is implicated in Hedgehog
signalling regulation in primary cilia (30-32).
The control of the disassembly and resorption of primary cilia is
shown in Fig. 2. In particular,
AURKA can phosphorylate INPP5E, subsequently increasing its
5-phosphatase activity, and its highest affinity is for PI(3,4,5)P3
(31-33). INPP5E interacts with the p85
subunit of PI3K, leading to the inhibition of the PI3K/AKT
signalling pathway (34). In fact,
293 cells stably transfected with the INPP5E gene were found to be
highly susceptible to FAS-induced apoptosis and cell growth was
suppressed (35). Accumulation of
PI(4,5)P2 in the distal half of the cilium led
to decapitation by controlling intraciliary F-actin assembly,
followed by resorption of the primary cilium (36). PI(4,5)P2
accumulation caused by INPP5E mutations negatively regulates the
Sonic Hedgehog (SHH) pathway (32,37,38).
However, in order to cause ciliary resorption, it is necessary that
AURKA phosphorylates KIF2A and HDAC6 (or HDAC2 in PDAC), but not
INPP5E.
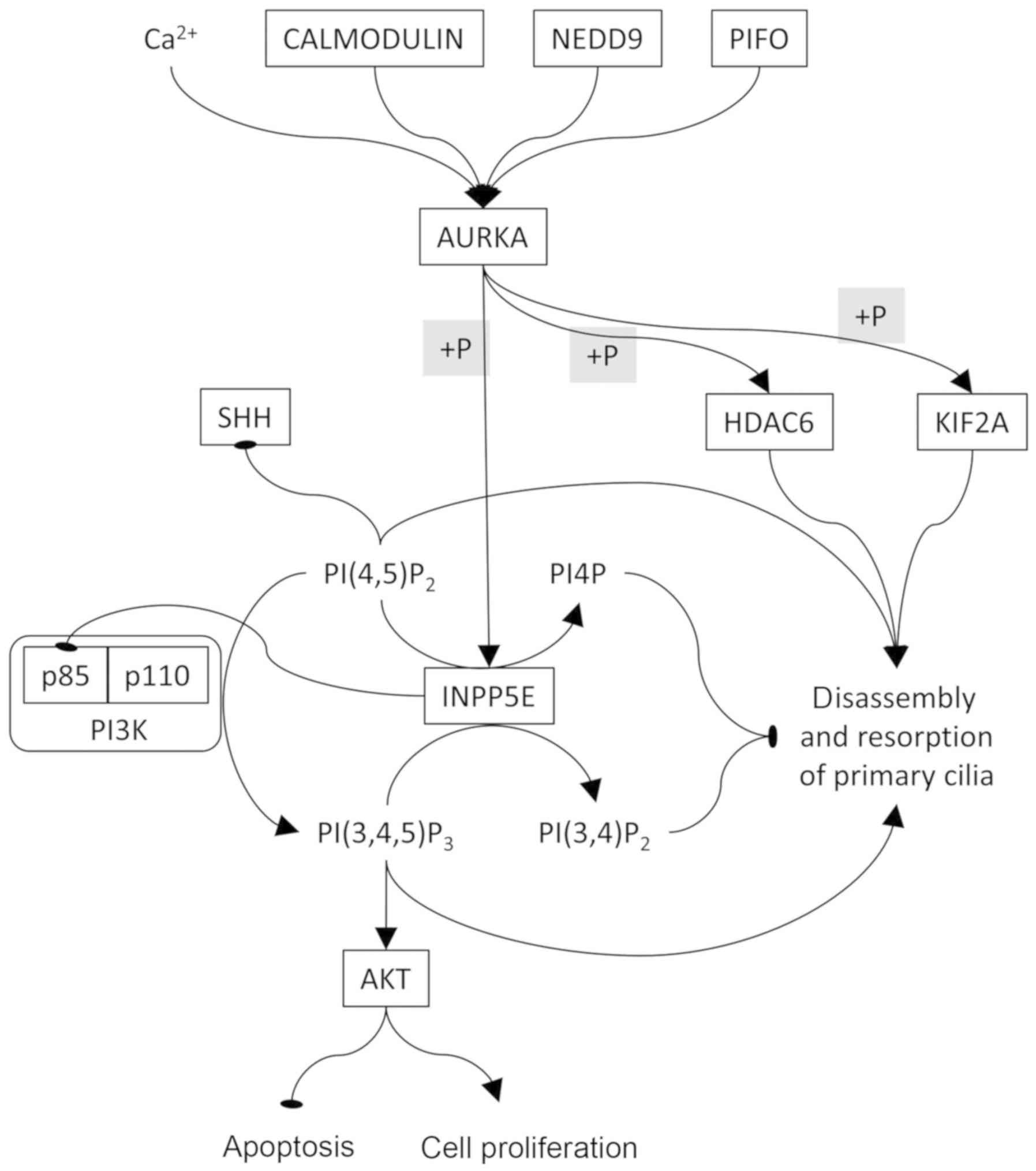 | Figure 2AURKA is the key element controlling
the disassembly and resorption of the primary cilium. AURKA is
controlled by calcium and various proteins, including calmodulin,
NEDD and PIFO. AURKA activates INPP5E by phosphorylation. INPP5E
converts PI(4,5)P2 to PI4P and PI(3,4,5)P3 to
PI(3,4)P2. PI3K converts PI(4,5)P2 to
PI(3,4,5)P3
which, in turn, activates AKT, leading to resistance to apoptosis
and cell proliferation. In order to allow disassembly and
resorption of the primary cilium, AURKA must negatively regulate
INPP5E. Oval-ending connectors indicate inhibition. AURKA, Aurora A
kinase; PIFO, Pitchfork; NEDD, neural precursor cell expressed
developmentally downregulated protein; PI, phosphatidylinositol;
INPP5E, inositol polyphosphate-5-phosphatase E; HDAC6, histone
deacetylase 6; KIF, kinesin family member; SHH, Sonic Hedgehog. |
Seeley et al studied the abundance and
distribution of primary cilia in 17 patients with PDAC to establish
whether primary ciliogenesis is disrupted in cancer (15). The abundance of primary cilia in
histologically normal tissues (negative control samples from each
patient) generally varied. On the other hand, almost all pancreatic
cancer samples were devoid of primary cilia, independently of
tumour stage, suggesting that absence of primary cilia is a
conserved characteristic of tumour cells. Authors have reported
lack of primary cilia in PanIN-1, 2 and 3 lesions; therefore, the
arrest in ciliary formation would occur in the earliest stages of
PDAC development. Since centro-acinar and ductal pancreatic cells
are ciliated, PanIN formation would involve ciliogenesis arrest of
either cell type. Based on the results, the authors concluded that
i) lack of primary cilia is independent of proliferation status,
ii) arrest of ciliary formation is reversible and iii)
overactivation of the K-Ras pathway arrests ciliogenesis in PDAC
(15). Primary cilia have been
restored in PDAC upon blocking K-Ras signalling with PI3K and MEK
inhibitors. This result suggested that activated K-Ras signalling
is directly involved in the inhibition of ciliogenesis. Since
overstimulation of the Hedgehog pathway is a hallmark of PDAC, and
absence of primary cilia upregulates this pathway, it has been
hypothesized that deciliation is involved in tumour progression via
upregulation of the Hedgehog pathway (25).
In the study of Emoto et al, primary cilia
were identified in 100 PDAC patients who received no therapy prior
to initial surgery (39). Among
the cases presenting primary cilia (25 out of 100), almost all (24
out of 25) had lymph node metastasis, and this frequency was higher
than in cases where primary cilia were absent. There was a
statistically significant association between overall survival (OS)
and the tumour size, grade, lymph node metastasis and presence of
primary cilia. In particular, patients with cancer with primary
cilia had a poorer outcome compared with patients without primary
cilia (39). It has been reported
that SHH is highly expressed in PDAC tissues, in PanIN lesions and
in several pancreatic cancer cells deriving from primary tumours
and metastases, suggesting that the Hedgehog pathway plays a key
role in the early and late phases of PDAC tumorigenesis (40). Similarly, Emoto et al
demonstrated high levels of SHH expression in the PANC-1 and Aspc-1
pancreatic cancer cell lines (39). SHH can activate stromal cells,
abundant in primary cilia, which may consequently stimulate
epithelial cell proliferation and, therefore, tumorigenesis
(39). By overexpressing SHH in
mouse models, it has been demonstrated that pancreatic primary
tumours were larger and the rates of tumour invasion and metastasis
to the liver, spleen and peritoneum were higher (41). In addition, primary cilia and the
Hedgehog receptor SMO were co-present in stromal cells, both in
primary tumours and metastases, but were absent in primary tumour
cells and metastatic cells. Instead, SHH production follows an
opposite pattern. Moreover, SHH enhances the paracrine expression
of matrix metalloproteinase 9. These results suggest that SHH acts
on stromal cells, enriched in primary cilia, in order to regulate
the tumour microenvironment and promote metastasis formation
(41).
Deng et al investigated the biological
functions of cilia during the malignant transformation by knocking
down Tg737/IFT88 and KIF3A (a subunit of the kinesin-II complex),
two crucial components of cilia, in mouse embryonic fibroblasts and
HPDE6C7 (an immortalized epithelial cell line derived from normal
human pancreatic duct epithelial cells) (42). This study demonstrated that the
mevalonate (MVA) pathway is activated upon inactivation of
ciliogenesis and may accelerate the oncogene-induced transformation
of normal cells both in vitro and in vivo. Another
important characteristic of the MVA pathway products are many
non-sterol isoprenoids, such as farnesyl pyrophosphate and
geranylgeranyl diphosphate. These two products are known mediators
of protein prenylation, and their substrates also include small G
proteins such as Ras, Rho and Rac, which are implicated in a number
of tumours, including PDAC (42).
6. Association between primary ciliary
regulatory genes and PDAC
The Xena Functional Genomics Explorer tool
(https://xenabrowser.net/) was used to explore the
associations between RNAseq expression of key genes in primary
cilium regulation and phenotypic variables in the pancreatic
adenocarcinoma dataset from The Cancer Genome Atlas of 178
patients. As shown in Table II,
by analysing the Kaplan-Meier curves and using log-rank tests, it
was revealed that the expression of the AURKA, PLK1, NEK2, HDAC6
and INPP5E genes was associated with OS and progression-free
survival (PFS). In particular, lower expression of AURKA, PLK1 and
NEK2, and higher expression of HDAC6 and INPP5E, were associated
with better prognosis. Instead, higher expression of PIFO was only
associated with better OS. The gene expression of AURKA, PLK1,
NEK2, KIF2A, NEDD9, calmodulin (CALM)1, CALM2 and CALM3 is
statistically significant higher in tumours compared with in normal
tissues (data derived from the Genotype-Tissue Expression database;
www.gtexportal.org). On the other hand, the
expression of PIFO, HDAC6 and INPP5E is lower in tumours compared
with that in normal tissues. By using one-way ANOVA, it was
demonstrated that the gene expression of AURKA, PLK1 and NEK2
increases proportionally to the tumour grade, whereas CALM1
displays an inverse proportionality. Finally, no gene expression
was found to be associated with pathological stage. These
expression data indicate that AURKA induces ciliary resorption, it
is a negative prognostic index, and it is more highly expressed in
tumours of higher histological grade. As regards INPP5E, which
blocks ciliary resorption, the data suggest that its higher
expression improves the prognosis. Of note, high PIFO expression,
which induces ciliary resorption, would be expected to worsen the
prognosis; instead, our data demonstrated an improvement of
prognosis.
| Table IISurvival and expression analyses of
primary cilium related genes in pancreatic ductal
adenocarcinoma. |
Table II
Survival and expression analyses of
primary cilium related genes in pancreatic ductal
adenocarcinoma.
| Genes | Survival
| Expression
|
|---|
| Association with OS
(P-value) | Association with
PFS (P-value) | Expression in
primary tumor vs. normal tissue (P-value) | Association with
histological tumor grade (P-value) | Association with
pathological stage (P-value) |
|---|
| AURKA | Inverse
(0.01774) | Inverse
(0.02812) | Higher
(4.995e-83) | Direct
(0.001278) | NS |
| PLK1 | Inverse
(0.007640) | Inverse
(0.005791) | Higher
(4.142e-71) | Direct
(0.000005639) | NS |
| NEK2 | Inverse
(0.003376) | Inverse
(0.01698) | Higher
(3.723e-104) | Direct
(0.000008372) | NS |
| KIF2A | NS | NS | Higher
(5.708e-8) | NS | NS |
| NEDD9 | NS | NS | Higher
(3.901e-38) | NS | NS |
| CALM1 | NS | NS | Higher
(3.047e-12) | Inverse
(0.000006607) | NS |
| CALM2 | NS | NS | Higher
(2.185e-31) | NS | NS |
| CALM3 | NS | NS | Higher
(6.951e-11) | NS | NS |
| PIFO | Direct
(0.03329) | NS | Lower
(2.528e-21) | NS | NS |
| HDAC6 | Direct
(0.005232) | Direct
(0.0009176) | Lower
(0.00002954) | NS | NS |
| INPP5E | Direct
(0.0006044) |
Direct(0.01807) | Lower
(4.150e-13) | NS | NS |
7. Therapeutic strategies targeting primary
cilia in PDAC
Currently, it is widely hypothesized that the
therapies targeted at ciliary restoration, collectively termed
ciliotherapies, may induce re-differentiation of tumour cells to
normal cells, reduce tumour growth, or even initiate apoptosis of
cancer cells. Of note, ciliotherapies would vary among different
tumour types. However, it has been reported that HDAC6 activation
is a common event among various tumours. Gradilone et al
reported successful in vitro and in vivo results by
targeting HDAC6 in CCA (17,18).
HDAC6 was targeted with shRNA or with the inhibitor tubastatin-A,
resulting in restored primary cilia formation, decreased cell
proliferation and anchorage-independent growth. Although some
studies have previously reported that activation of HDAC6 drives
primary cilia resorption, it does not appear to be the case in
PDAC. Kobayashi et al treated PANC-1 cells with the HDAC6
inhibitor tubacin, or with HDAC6 siRNA, but no effect on primary
cilia assembly was observed, although HDAC6 was efficiently
inhibited (28). Since primary
ciliogenesis in PANC-1 cells was induced by treatment with
trichostatin A, a pan-HDAC inhibitor, further inhibitors (i.e.,
valproic acid, MS-275 and FK228/depsipeptide) and specific siRNAs
have been used to determine the involvement of HDAC family members
in cilia formation, and HDAC2 was found to be responsible for
inhibition of primary ciliogenesis in PANC-1 cells and in other
PDAC cell lines (28). HDAC2
likely suppresses primary cilia formation by controlling the
expression of genes involved in cilia assembly and disassembly,
including AURKA. Indeed, PANC-1 cells treated with AURKA inhibitors
(alisertib and PHA-680632) exhibited marked restoration of primary
cilia (28). Recently, in a 3D
mouse pancreatic acinar cell culture that displays acinar-to-ductal
metaplasia (a pancreatic pre-neoplastic lesion), the inhibition of
AURKA by MLN8237 (alisertib) significantly increased ciliogenesis.
Furthermore, tubacin-induced inhibition confirmed that HDAC6 is not
involved in cilium disassembly in the pancreas (43).
Some clinical trials investigating the safety and
efficacy of certain HDAC inhibitors in patients with PDAC are
ongoing. The majority involves pan-HDAC inhibitors, such as
vorinostat (ClinicalTrials.gov IDs NCT00667082,
NCT00831493, NCT00983268, NCT02349867 and NCT00948688) and
panobinostat (NCT01056601), or the class 1 inhibitors entinostat
(NCT03925428, NCT03760614, NCT00020579 and NCT03250273) and
romidepsin (NCT00379639 and NCT04257448) in combination with other
chemotherapy drugs for PDAC. However, there are currently no
ongoing clinical trials on HDAC2-specific inhibitors in PDAC
patients. HDAC inhibitors, although they have shown promising
results in preclinical models, are associated with certain
difficulties in the clinical setting, likely due to their toxicity,
which is associated with low specificity towards a single HDAC
member (44). As regards AURKA,
some clinical trials are focusing on its oral inhibitor, alisertib,
in combination with gemcitabine or nab-paclitaxel (NCT01677559 and
NCT01924260). However, due to the paucity of published preclinical
and clinical results, the usefulness of HDAC or AURKA inhibitors in
PDAC treatment remains to be clearly determined.
8. Conclusion
Primary cilia contain several proteins that act as
receptors, ion channels and transporters; thus, they are able to
receive various extracellular signals (mechanical stimuli, light,
low-molecular-weight molecules, etc.). Assembly and disassembly of
cilia are correlated with cell cycle status. In particular, most
cell types undergo cilia disassembly in the M phase and re-assembly
upon entry in the G1 phase. Therefore, inhibition of ciliogenesis
is crucial for proliferating cells, but its exact regulation
mechanism remains unknown at present. Based on the evidence
obtained to date, it appears that ciliary resorption is regulated
by the AURKA-HDAC6 and PLK1-KIF2A loops in the G1 phase, and by the
NEK2-KIF24 loop in the G2/M phase.
Lack of primary cilia has been reported in tumour
cells characterized by dysregulation of certain signalling
pathways. The consequences of ciliary absence may differ depending
on the context; in some cases it may promote tumour growth, whereas
in others it may act as an inhibitor. However, some of these
pathways may work normally, even in the absence of cilia, although
many ciliary proteins have been associated with signalling
pathways. Therefore, it would be reasonable to suggest that
signalling pathways may include cilium-dependent and -independent
mechanisms. Loss of primary cilia has been observed in a number of
tumour samples, indicating their role in carcinogenesis due to
aberrant signalling, which may lead cells into a hyperproliferative
state and increase their migration and invasion abilities. PDAC
cells also lack primary cilia, independently of proliferation
status. It has been suggested that K-Ras may be involved in
ciliogenesis regulation in PDAC. Furthermore, HDAC2 may cause loss
of primary cilia by regulating AURKA transcription independently of
K-Ras.
In the present study, the gene regulation of AURKA
and INPP5E was reconstructed and it was demonstrated that
AURKA-mediated INPP5E phosphorylation blocks the resorption of
primary cilia. On the other hand, the lack of phosphorylation of
INPP5E activates the PI3K/AKT pathway, which promotes the
proliferation and survival of cancer cells. Through analysing gene
expression databases, it was highlighted which primary cilia
regulatory genes have a high probability to be associated to
prognosis, histological grade and pathological stage in patients
with PDAC. As expected, a higher expression of AURKA (which induces
primary cilia resorption) was found to be associated with lower OS
and PFS and with higher histological degree. Similarly, the
expression of INPP5E (which prevents primary cilia resorption) was
associated with higher OS and PFS. However, further experimental
studies are required to reach definitive conclusions on their
roles. Indeed, the databases used herein store RNAseq data and not
data on protein expression or enzymatic activity, which are yet to
be evaluated.
It would be interesting to determine the precise
downstream targets of AURKA phosphorylation, which may cause
deciliation of pancreatic cells and PDAC progression, possibly via
upregulation of the Hedgehog pathway. Many questions are to be
answered regarding primary cilia and ciliogenesis regulation. A
better understanding of this process would be useful for the
development of therapies targeted at cilia restoration. These
ciliotherapies may decrease tumour growth rate or stimulate
apoptosis of cancer cells. However, although several clinical
trials for HDAC and AURKA inhibitors are currently in progress,
there is yet no validated clinical application for PDAC.
Funding
No funding was received.
Availability of data and material
Not applicable.
Authors' contributions
BS and MG: writing and revision of the manuscript.
BS, MG and FP: reference collection, discussion. FP: conception and
design of the study. All authors have read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Higgins M, Obaidi I and McMorrow T:
Primary cilia and their role in cancer. Oncol Lett. 17:3041–3047.
2019.PubMed/NCBI
|
|
2
|
Pala R, Alomari N and Nauli SM: Primary
Cilium-dependent signaling mechanisms. Int J Mol Sci. 18:22722017.
View Article : Google Scholar :
|
|
3
|
Malicki JJ and Johnson CA: The Cilium:
Cellular antenna and central processing unit. Trends Cell Biol.
27:126–140. 2017. View Article : Google Scholar :
|
|
4
|
Sorokin SP: Reconstructions of centriole
formation and ciliogenesis in mammalian lungs. J Cell Sci.
3:207–230. 1968.PubMed/NCBI
|
|
5
|
Wheway G, Nazlamova L and Hancock JT:
Signaling through the primary cilium. Front Cell Dev Biol. 6:82018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Izawa I, Goto H, Kasahara K and Inagaki M:
Current topics of functional links between primary cilia and cell
cycle. Cilia. 4:122015. View Article : Google Scholar
|
|
7
|
Jonassen JA, San Agustin J, Follit JA and
Pazour GJ: Deletion of IFT20 in the mouse kidney causes
misorientation of the mitotic spindle and cystic kidney disease. J
Cell Biol. 183:377–384. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pugacheva EN, Jablonski SA, Hartman TR,
Henske EP and Golemis EA: HEF1-dependent Aurora A activation
induces disassembly of the primary cilium. Cell. 129:1351–1363.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tucker RW, Pardee AB and Fujiwara K:
Centriole ciliation is related to quiescence and DNA synthesis in
3T3 cells. Cell. 17:527–535. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li A, Saito M, Chuang J, Tseng Y, Dedesma
C, Tomizawa K, Kaitsuka T and Sung C: Ciliary transition zone
activation of phosphorylated Tctex-1 controls ciliary resorption,
S-phase entry and fate of neural progenitors. Nat Cell Biol.
13:402–411. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Breslin L, Prosser SL, Cuffe S and
Morrison CG: Ciliary abnormalities in senescent human fibroblasts
impair proliferative capacity. Cell Cycle. 13:2773–2779. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Spalluto C, Wilson DI and Hearn T: Nek2
localises to the distal portion of the mother centriole/basal body
and is required for timely cilium disassembly at the G2/M
transition. Eur J Cell Biol. 91:675–686. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim S, Lee K, Choi J, Ringstad N and
Dynlacht BD: Nek2 activation of Kif24 ensures cilium disassembly
during the cell cycle. Nat Commun. 6:80872015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moser JJ, Fritzler MJ and Rattner JB:
Primary ciliogenesis defects are associated with human
astrocytoma/glioblastoma cells. BMC Cancer. 9:4482009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Seeley ES, Carrière C, Goetze T,
Longnecker DS and Korc M: Pancreatic cancer and precursor
pancreatic intraepithelial neoplasia lesions are devoid of primary
cilia. Cancer Res. 69:422–430. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gherman A, Davis EE and Katsanis N: The
ciliary proteome database: An integrated community resource for the
genetic and functional dissection of cilia. Nat Genet. 38:961–962.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gradilone SA, Pisarello MJL and LaRusso
NF: Primary cilia in tumor biology: The primary cilium as a
therapeutic target in cholangiocarcinoma. Curr Drug Targets.
18:958–963. 2017. View Article : Google Scholar :
|
|
18
|
Gradilone SA, Radtke BN, Bogert PS, Huang
BQ, Gajdos GB and LaRusso NF: HDAC6 inhibition restores ciliary
expression and decreases tumor growth. Cancer Res. 73:2259–2270.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim J, Dabiri S and Seeley ES: Primary
cilium depletion typifies cutaneous melanoma in situ and malignant
melanoma. PLoS One. 6:e274102011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hassounah NB, Nagle R, Saboda K, Roe DJ,
Dalkin BL and McDermott KM: Primary cilia are lost in preinvasive
and invasive prostate cancer. PLoS One. 8:e685212013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yuan K, Frolova N, Xie Y, Wang D, Cook L,
Kwon YJ, Steg AD, Serra R and Frost AR: Primary cilia are decreased
in breast cancer: Analysis of a collection of human breast cancer
cell lines and tissues. J Histochem Cytochem. 58:857–870. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ho L, Ali SA, Al-Jazrawe M, Kandel R,
Wunder JS and Alman BA: Primary cilia attenuate hedgehog signalling
in neoplastic chondrocytes. Oncogene. 32:5388–5396. 2013.
View Article : Google Scholar
|
|
23
|
Egeberg DL, Lethan M, Manguso R, Schneider
L, Awan A, Jprgensen TS, Byskov AG, Pedersen LB and Christensen ST:
Primary cilia and aberrant cell signaling in epithelial ovarian
cancer. Cilia. 1:152012. View Article : Google Scholar
|
|
24
|
Dere R, Perkins AL, Bawa-Khalfe T, Jonasch
D and Walker CL: p-catenin links von Hippel-Lindau to aurora kinase
A and loss of primary cilia in renal cell carcinoma. J Am Soc
Nephrol. 26:553–564. 2015. View Article : Google Scholar
|
|
25
|
Kobayashi T and Itoh H: Loss of a primary
cilium in PDAC. Cell Cycle. 16:817–818. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Quilichini E, Fabre M, Dirami T, Stedman
A, De Vas M, Ozguc O, Pasek RC, Cereghini S, Morillon L, Guerra C,
et al: Pancreatic ductal deletion of Hnf1b disrupts exocrine
homeostasis, leads to pancreatitis, and facilitates tumorigenesis.
Cell Mol Gastroenterol Hepatol. 8:487–511. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schimmack S, Kneller S, Dadabaeva N,
Bergmann F, Taylor A, Hackert T, Werner J and Strobel O: Epithelial
to stromal Re-distribution of primary cilia during pancreatic
carcinogenesis. PLoS One. 11:e01642312016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kobayashi T, Nakazono K, Tokuda M, Mashima
Y, Dynlacht BD and Itoh H: HDAC2 promotes loss of primary cilia in
pancreatic ductal adenocarcinoma. EMBO Rep. 18:334–343. 2017.
View Article : Google Scholar :
|
|
29
|
Li D, Zhu J, Firozi PF, Abbruzzese JL,
Evans DB, Cleary K, Friess H and Sen S: Overexpression of oncogenic
STK15/BTAK/Aurora A kinase in human pancreatic cancer. Clin Cancer
Res. 9:991–997. 2003.PubMed/NCBI
|
|
30
|
Plotnikova OV, Nikonova AS, Loskutov YV,
Kozyulina PY, Pugacheva EN and Golemis EA: Calmodulin activation of
Aurora-A kinase (AURKA) is required during ciliary disassembly and
in mitosis. Mol Biol Cell. 23:2658–2670. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Plotnikova OV, Seo S, Cottle DL, Conduit
S, Hakim S, Dyson JM, Mitchell CA and Smyth IM: lNPP5E interacts
with aurka, linking phosphoinositide signaling to primary cilium
stability. J Cell Sci. 128:364–372. 2015. View Article : Google Scholar :
|
|
32
|
Garcia-Gonzalo FR, Phua SC, Roberson EC,
Garcia G, Abedin M, Schurmans S, Inoue T and Reiter JF:
Phosphoinositides regulate ciliary protein trafficking to modulate
hedgehog signaling. Dev Cell. 34:400–409. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kisseleva MV, Wilson MP and Majerus PW:
The isolation and characterization of a cDNA encoding
phospholipid-specific inositol polyphosphate 5-phosphatase. J Biol
Chem. 275:20110–20116. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bielas SL, Silhavy JL, Brancati F,
Kisseleva MV, Al-Gazali L, Sztriha L, Bayoumi RA, Zaki MS,
Abdel-Aleem A, Rosti RO, et al: Mutations in INPP5E, encoding
inositol polyphosphate-5-phosphatase E, link phosphatidyl inositol
signaling to the ciliopathies. Nat Genet. 41:1032–1036. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kisseleva MV, Cao L and Majerus PW:
Phosphoinositide-specific inositol polyphosphate 5-phosphatase IV
inhibits Akt/protein kinase B phosphorylation and leads to
apoptotic cell death. J Biol Chem. 277:6266–6272. 2002. View Article : Google Scholar
|
|
36
|
Phua SC, Chiba S, Suzuki M, Su E, Roberson
EC, Pusapati GV, Schurmans S, Setou M, Rohatgi R, Reiter JF, et al:
Dynamic remodeling of membrane composition drives cell cycle
through primary cilia excision. Cell. 168:264–279.e15. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mukhopadhyay S, Wen X, Ratti N, Loktev A,
Rangell L, Scales SJ and Jackson PK: The ciliary G-protein-coupled
receptor Gpr161 negatively regulates the sonic hedgehog pathway via
cAMP signaling. Cell. 152:210–223. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chavez M, Ena S, Van Sande J, de Kerchove
d'Exaerde A, Schurmans S and Schiffmann SN: Modulation of ciliary
phosphoinositide content regulates trafficking and sonic hedgehog
signaling output. Dev Cell. 34:338–350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Emoto K, Masugi Y, Yamazaki K, Effendi K,
Tsujikawa H, Tanabe M, Kitagawa Y and Sakamoto M: Presence of
primary cilia in cancer cells correlates with prognosis of
pancreatic ductal adenocarcinoma. Hum Pathol. 45:817–825. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Thayer SP, di Magliano MP, Heiser PW,
Nielsen CM, Roberts DJ, Lauwers GY, Qi YP, Gysin S, Fernandez-del
Castillo C, Yajnik V, et al: Hedgehog is an early and late mediator
of pancreatic cancer tumorigenesis. Nature. 425:851–856. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bailey J, Mohr A and Hollingsworth M:
Sonic hedgehog paracrine signaling regulates metastasis and
lymphangiogenesis in pancreatic cancer. Oncogene. 28:3513–3525.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Deng YZ, Cai Z, Shi S, Jiang H, Shang YR,
Ma N, Wang JJ, Guan DX, Chen TW, Rong YF, et al: Cilia loss
sensitizes cells to transformation by activating the mevalonate
pathway. J Exp Med. 215:177–195. 2018. View Article : Google Scholar :
|
|
43
|
Bangs FK, Miller P and O'Neill E:
Ciliogenesis and Hedgehog signalling are suppressed downstream of
KRAS during Acinar-ductal metaplasia in mouse. Dis Model Mech.
13:dmm0442892020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Baretti M, Ahuja N and Azad NS: Targeting
the epigenome of pancreatic cancer for therapy: Challenges and
opportunities. Ann Pancreat Cancer. 2:182019. View Article : Google Scholar
|