Introduction
Colorectal cancer (CRC) was the third most commonly
diagnosed malignancy and the second leading cause of cancer-related
deaths worldwide in 2020 (1). As a
primary treatment, enhancing the radio response of locally advanced
rectal cancer is essential to achieving local control and improving
long-term prognosis. Despite the effectiveness of radiotherapy in
treating cancer, damage caused by radiation to the surrounding
normal cells is often unavoidable (2). Therefore, it is necessary to design a
strategy for effectively treating the tumor while minimizing these
nontargeted side effects. One such approach includes the use of
radiosensitizers, which can render cancer cells more sensitive to
radiation, thereby enabling a lower and safer radiation dose to
achieve the same therapeutic response (3). Over the past decade, several studies
have addressed the efficacy of diverse radiosensitizers, including
fluorouracil (5-FU), oxaliplatin, capecitabine and bevacizumab, in
combination with radiotherapy for CRC. Specially, 5-FU achieved a
complete pathological response rate of ~20-30% (4-6).
However, in addition to their effects on cancer cells, these
radiosensitizers are also reportedly toxic to healthy cells
(7,8); therefore, while they may function to
enhance the efficiency of radiation, they do not effectively
minimize the adverse side effects.
Additional research has revealed the influence of
intestinal microbes derived from diet on the development of CRC
(9,10). As food flows through the colon, the
fiber can be fermented by gut microbiota to produce short-chain
fatty acids (SCFAs), such as propionate, acetate and butyrate,
which play multifunctional roles in the intestinal epithelial cells
(11). In particular, butyrate is
used as an energy source in normal colonocytes. Under the Warburg
effect, cancer cells rely on glucose as their primary energy
source, resulting in nuclear accumulation of butyrate that
functions as a histone deacetylase (HDAC) inhibitor (12-14).
Considering that other HDAC inhibitors, including suberoylanilide
hydroxamic acid (SAHA) and valproic acid, have been shown to
enhance the radiosensitivity of cancer cells (15-17),
we hypothesized that butyrate, along with other potential SCFAs,
may serve as effective, cancer cell-specific and safe
radiosensitizers.
To date, responses to radiotherapy have been
investigated exclusively in animal models owing to the low success
rate associated with the initiation of patient-derived cell
cultures (18,19). However, several studies have
demonstrated the value of 3D patient-derived organoid (PDO) culture
systems as useful and physiologically relevant tools to study the
treatment response (20,21). Organoids are self-organizing 3D
structures grown from stem cells to mimic in vivo
architecture and biology. Hence, organoids more closely mimic in
vivo tissues than cells grown in two-dimensional tissue
cultures, and are valuable tools for modeling human diseases,
including cancers (22).
Therefore, in the present study, PDOs from CRC
patients (CRC-PDO) were used as a model system for investigating
the response to radiotherapy, and to evaluate the influence of
combined radiotherapy with the SCFAs butyrate, propionate and
acetate as candidate radiosensitizers through regulation of HDAC
activity. Cumulatively, these results lay the foundation for
development of a novel combined radiation/SCFA strategy for CRC and
promote the utility of CRC-PDOs as a model system for investigating
tumor evolution and modulating drug response.
Materials and methods
Human specimens
A total of 7 rectal cancer samples and 1 colon
cancer tumor sample and the paired healthy tissue samples were
obtained from 8 patients diagnosed with CRC between December 2017
and May 2018 at the Korea Cancer Center Hospital (Seoul, Korea).
All tumors were staged according to the pathological
tumor/node/metastasis classification (8th edition) of the American
Joint Committee on Cancer (23).
Surgically resected and endoscopic biopsy samples were verified by
pathologists through hematoxylin and eosin staining as previously
described (24). The 7 rectal
samples were obtained during an endoscopic biopsy, and the 1 colon
cancer sample was obtained by low anterior resection. The mean age
of patients was 61.5 years (range, 53-85 years), and 4 patients
were male and 4 patients were female. Clinical data, including
patient information, such as age, sex and tumor locations, are
provided in Table SI.
This study was approved by Ethics Committee of Korea
Cancer Center Hospital (approval no. KIRAMS-2017-07-001). All
research was performed in accordance with the approved guidelines
and regulations of the institution. All samples were obtained from
patients who had provided written informed consent for the use of
their tissues for the purposes of research after the operation.
Organoid culture
Both tumor and adjacent normal tissues were
collected, from which healthy crypts and tumor cells were isolated,
as described previously (20,25),
with minor modifications. Briefly, normal intestinal tissue
fragments were incubated in 8 mM EDTA-phosphate buffered saline
(PBS) for 20 min on ice and further incubated for 8 min at 37°C.
Under these dissociation conditions, colonic crypts were mildly
digested, thereby physically separating their bottom and top
segments. Cancer tissues were incubated with collagenase type II
(Sigma-Aldrich; Merck KGaA), dispase type II (Roche Applied
Science) and Y-27632 (BioVision, Inc.) for 30 min at 37°C. Isolated
crypts and cells were washed with PBS and centrifuged at 300 × g
for 3 min at room temperature. The cells were then embedded in
Matrigel on ice (growth factor reduced, phenol red free; Corning,
Inc.) and seeded in 24-well plates, followed by addition of culture
medium. The composition of the CRC-PDO culture medium was 1× B27
(Gibco; Thermo Fisher Scientific, Inc.), 1.25 mM N-acetyl
cysteine (United States Pharmacopeia), 50 ng/ml human epidermal
growth factor (BioVision, Inc.), 50 ng/ml human Noggin (PeproTech,
Inc.), 10 nM gastrin (Sigma-Aldrich; Merck KGaA), 500 nM A83-01
(BioVision, Inc.) and 100 mg/ml primocin (InvivoGen). In
experiments that prevented the Warburg effect, organoids were grown
from high-glucose (3.151 g/l) to low-glucose (1 g/l) medium in same
composition. Wnt signaling-related factors are essential for the
growth of healthy colon organoids (normal-PDOs). However, in most
cases, CRC is associated with mutations that aberrantly activate
the Wnt signaling pathway (26).
Therefore, for selection of tumor cells, CRC-PDOs were cultured
without Wnt-related factors in the culture medium to ensure that
normal tissue-derived organoids did not contaminate tumor tissues
(27). The normal-PDOs were
cultured in IntestiCult™ Organoid Growth Medium (Stemcell
Technologies, Inc.) and CRC-PDO culture medium with CHIR99021
(Stemgent; ReproCELL) and R-spondin1 (R&D Systems, Inc.). To
prevent anoikis, 10 µM Y-27632 was added to the culture
medium for the first 2-3 days. Maintenance and freezing of
organoids was carried out as described previously with slight
modification (25). When organoids
were >200 µm, they were passaged by pipetting using
Gentle Cell Dissociation Reagent (Stemcell Technologies, Inc.)
according to the manufacturer's instructions.
Organoid viability
For further dissociation into single cells,
organoids were resuspended in TrypLE Express (Thermo Fisher
Scientific, Inc.) via pipetting with a p200 pipette, and incubated
at 37°C for 10 min. Cells were then pipetted multiple times to form
a homogeneous resuspension and centrifuged at 600 × g for 5 min at
4°C, followed by discarding of the supernatant to obtain pelleted
cells. The pellet was resuspended with Matrigel and divided into a
96-well plate (5,000 cells/10 µl Matrigel per well). After
the Matrigel was polymerized, 100 µl culture media was
added. Organoid viability was assessed as described previously
(28) with slight modifications.
Briefly, 3-[4,5-dimethylthiazol-2-yl]-2, 5 diphenyl tetrazolium
bromide (MTT) solution was added to the organoid culture at a final
concentration of 500 µg/ml. After incubation for 3 h, the
medium was discarded, and 20 µl 2% sodium dodecyl sulfate
solution was added to solubilize the Matrigel for 2 h. Next, 100
µl DMSO was added for 1 h to solubilize the reduced MTT, and
the optical density was measured on a BioTek Eon microplate
absorbance reader (BioTek Instruments, Inc.) at 562 nm. Matrigel
without organoids (10 µl) was used as the control.
Treatment of organoids with compounds and
radiation
To test radiosensitivity, the CRC-PDOs were
irradiated with Matrigel in the plates after treatment for 6 h with
1 mM acetate (Sigma-Aldrich; Merck KGaA), butyrate (Sigma-Aldrich;
Merck KGaA), propionate (Sigma-Aldrich; Merck KGaA) and/or 100 nM
FOXO inhibitor, AS1842856 (EMD Millipore) at 37°C using a 137Cs
γ-ray source (Atomic Energy of Canada Ltd.) at a dose rate of 3.81
Gy/min. The concentration of 1 mM SCFA was selected as described
previously (29). The organoids
were irradiated with three fractions of 5 Gy (one fraction per
day), and their size was analyzed by ImageJ software 1.48 V
(National Institutes of Health) on days 0 and 3. For analysis at
the 2nd passage, organoids were treated with butyrate and/or
ionizing radiation (IR). After 72 h, organoids were passaged by
pipetting using Gentle Cell Dissociation Reagent with a 1:2-1:4
split ratio. After 72 h, organoids were evaluated using the EVOS FL
Cell Imaging System (Thermo Fisher Scientific, Inc.).
Immunocytochemistry and
immunohistochemistry
Intestinal stem cells can differentiate into all
cell lineages of the intestinal epithelium, including goblet cells
(mucin 2+; Muc2+), entero-endocrine cells
(chromogranin A+; ChgA+) and enterocytes
(VL1+); meanwhile all proliferating cells express Ki-67
(30). PDOs were fixed in 4%
paraformaldehyde at room temperature for 24 h, embedded in paraffin
and then dissected into sections (5-µm). Following treatment
with Smartblock (CANDOR Bioscience) for 30 min at room temperature,
the slides were incubated with primary antibodies against
anti-Ki-67 (1:200; cat. no. ab16667; Abcam), anti-Muc2 (1:100; cat.
no. ab90007; Abcam) and anti-ChgA (1:100; cat. no. 20086;
ImmunoStar) at 4°C overnight. Slides were then incubated with Alexa
Fluor 594 goat antibody to rabbit IgG (H+L) (1:200; cat. no.
A11012; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature. Images were acquired using a EVOS FL Cell Imaging
System (Thermo Fisher Scientific, Inc.).
The cellular efficacy of recognizing double-strand
DNA breaks (DSB) and DNA damage response can be assessed by
observing the foci of γ-H2AX (31). For the detection of γ-H2AX,
organoids were seeded in 8-well glass chamber slides (LabTek
Services, Ltd.). The cells were fixed with 4% paraformaldehyde for
30 min at room temperature and permeabilized with 0.1% Triton X-100
in PBS. Unspecific epitopes on organoids were blocked with 5%
bovine serum albumin (Gene Depot) in Tris-buffered saline for 1 h
at room temperature and incubated with anti-γ-H2AX (1:100; cat. no.
05636; EMD Millipore) overnight at 4°C, followed by incubation with
Alexa Fluor 488 goat antibody to mouse IgG (H+L) (1:200; cat. no.
A11001; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature.
To characterize organoids and their tissue of
origin, immunohistochemistry was performed with colorectal markers
cytokeratin (CK)19, CK20 and CDX2, as described previously
(32). Organoids and tissues were
fixed in 4% paraformaldehyde at room temperature overnight and
embedded in paraffin blocks. Subsequently, 3-µm sections
were dewaxed in xylene and hydrated through a graded series of
alcohol. Endogenous peroxidase activity was quenched by incubating
the sections for 30 min in 0.3% H2O2 and
incubated in Smartblock (CANDOR Bioscience) for 30 min at room
temperature to reduced non-specific binding. Sections were
incubated for 1 h at 37°C with anti-CDX2 (1:200; cat. no. 235R-16;
Cell Marque; Sigma-Aldrich; Merck KGaA), CK20 (1:500; cat. no.
320M-16; Cell Marque; Sigma-Aldrich; Merck KGaA) and CK19 (1:400;
cat. no. ab15463; Abcam). Detection was performed on an
Envision/Horseradish Peroxidase system (Dako; Agilent Technologies,
Inc.), and counterstained with hematoxylin for 10 min at room
temperature. Finally, the sections were dehydrated through a graded
series of alcohol, cleared in xylene and mounted. Images were
acquired using the IX73 inverted microscope (Olympus Corporation;
magnification, ×40).
EdU staining
EdU staining was performed with a Click-iT Plus EdU
FACS kit (cat. no. C10424; Thermo Fisher Scientific, Inc.) and
Click-iT® Plus EdU Imaging kits (cat. no. C10639; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Images were acquired using the EVOS FL Cell Imaging
system fluorescence microscope (Thermo Fisher Scientific, Inc.;
magnification, ×200).
Lactate assays
Lactate concentration was determined using a Lactate
Assay kit (cat. no. K607-100; BioVision) according to the
manufacturer's instructions. Optical density was measured on a
BioTek Eon microplate absorbance reader (BioTek Instruments, Inc.)
at 570 nm 30 min after the addition of the substrate. The mean
values of lactate concentrations were calculated for each condition
based on the data obtained from three independent experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Organoids were harvested from Matrigel by first
washing them with cold PBS and incubating in cell recovery solution
(Corning, Inc.) at 4°C for 1 h (33). Total RNA was isolated using a
RNeasy mini kit (Qiagen GmbH). Subsequently, 2 µg RNA was
used for cDNA synthesis using RevertAid first strand cDNA kit
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Gene expression levels were quantified using the SYBR
Green PCR kit (Qiagen) and qPCRs were run on an ABI7500 Real-Time
PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The qPCR thermocycling conditions were as follows: 94°C for 5 min,
followed by 30 cycles at 94°C for 30 sec and 60°C for 30 sec, and a
final extension step at 72°C for 10 min. The amount of target mRNA
was normalized to that of GAPDH mRNA and analyzed as previously
described (34). Primers are
listed in Table SII. Cycle
threshold (Cq) values were obtained using an auto baseline, which
was applied to all amplicons of the same primer set by the
corresponding PCR instrument software (Applied Biosystems; Thermo
Fisher Scientific, Inc.). Triplicates with a Cq value difference of
>0.5 were not considered and excluded from the analysis.
Relative concentrations of cDNA for analyzing relative changes in
gene expression were calculated using the 2−ΔΔCq method
(35).
Western blotting
For western blot analysis, organoids were washed
with cold PBS and lysed in RIPA buffer (Thermo Fisher Scientific,
Inc.). Proteins were quantified using the Bradford method and 20
µg protein/lane was resolved by 8-13.5% SDS-PAGE. The
membranes were incubated with primary antibodies against cleaved
poly-ADP-ribose polymerase (c-PARP; 1:1,000; cat. no. 5625; Cell
Signaling Technology, Inc.), PARP (1:1,000; cat. no. 9542; Cell
Signaling Technology, Inc.), cleaved caspase 3 (c-caspase 3;
1:1,000; cat. no. 9664; Cell Signaling Technology, Inc.), caspase 3
(1:1,000; cat. no. 9662; Cell Signaling Technology, Inc.),
phosphorylated (p)-JNK (1:1,000; cat. no. 9251; Cell Signaling
Technology, Inc.), JNK (1:1,000; cat. no. 9252; Cell Signaling
Technology, Inc.), p-p38 (1:1,000; cat. no. 9211; Cell Signaling
Technology, Inc.), p38 (1:1,000; cat. no. 9212; Cell Signaling
Technology, Inc.), FOXO3A (1:1,000; cat. no. 2499; Cell Signaling
Technology, Inc.), LaminB (1:1,000; cat. no. SC-6216; Santa Cruz
Biotechnology, Inc.) and β-actin (1:5,000; cat. no. A5441;
Sigma-Aldrich; Merck KGaA) overnight at 4°C, followed by incubation
with corresponding horseradish peroxidase-conjugated secondary
antibodies (1:2,000; cat. nos. SC-2357 and SC-516102; Santa Cruz
Biotechnology, Inc.) for 1 h at room temperature. Proteins were
visualized using enhanced chemiluminescence (Thermo Fisher
Scientific, Inc.). Western blot images were analyzed using Bio-Rad
ChemiDoc (Bio-Rad Laboratories, Inc.).
Targeted next-generation sequencing
assay
To analyze the mutational status of tissues and
organoids, they were harvested using cell recovery solution. DNA
extraction and library construction were performed by Macrogen with
Axen Cancer Panel 2 (Macrogen) comprising 170 cancer-related genes.
The libraries were paired-end sequenced (2×150 bp) on a NextSeq 500
system (Illumina, Inc.) with high-throughput sequencing using
synthesis technology to a depth coverage of ~2,000×.
Statistical analysis
The data obtained from a minimum of three
independent experiments are expressed as the mean ± standard
deviation. Unpaired two-tailed Student's t-tests were applied to
determine significant differences between two groups. One-way ANOVA
followed by Tukey's test was performed to compare the means between
groups when performing multiple comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Statistical analysis was performed using GraphPad Prism 7 (GraphPad
Software, Inc.). Analysis of the mutation-annotated files was
conducted using the maftools R package V 3.5.2, which included the
generation of figure oncoplots (36).
Results
Butyrate exhibits anticancer effects
against CRC-PDO and shows high specificity
Immunocytochemistry confirmed that the established
CRC-PDOs were positive for all markers of intestinal epithelium
cell lineages (Muc2+, ChgA+, VL1+,
Ki-67+; Fig. S1). In
addition, the CRC-PDOs had the same genetic landscape as their
matching tumor tissues, as demonstrated in the oncoplot analysis
(Fig. 1A). Immunohistochemistry
analysis revealed that CK19, CK20 and CDX2 were expressed in
CRC-PDO #11 original tissues and organoids (Fig. 1B). Meanwhile other tissues and
organoids were weakly positive for CK19, whereas strong expression
of CK20 and CDX2 was detected in CRC-PDO #3 (Fig. S2). These features reflect those
present in the patient tissue samples. The tumor organoids were
selectively expanded by excluding Wnt-related factors from culture
medium (Fig. 1C), and
immunocytochemical analysis revealed that CRC-PDOs contained more
proliferating cells compared with normal-PDOs (P<0.001; Fig. 1D), confirming that the organoid
cultures reflect the characteristics of the primary tissue. Of the
three SCFAs screened, only butyrate significantly decreased the
organoid size (P=0.038), while propionate and acetate had no
effects (Figs. 1E and F, and
S3A and B). These observations
were supported by the MTT assay results (Fig. S3C). By contrast, butyrate did not
have any adverse effects on normal-PDOs (Fig. 1G and H). These findings suggest
that butyrate may exhibit selective antitumor activity on
CRC-PDO.
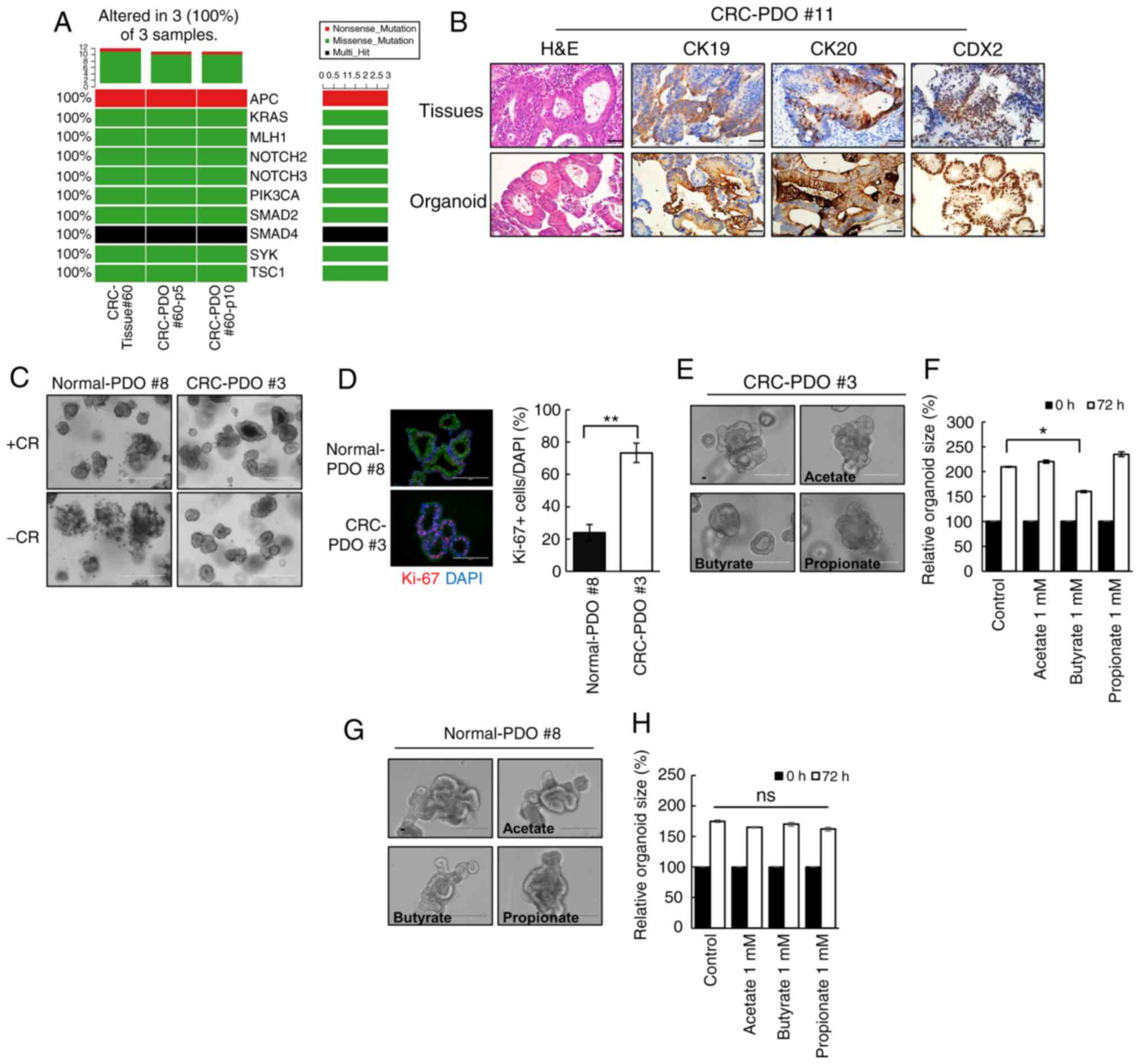 | Figure 1Butyrate exhibits selective
anticancer effects on CRC-PDOs. (A) Oncoplot of somatic cancer
driver alterations in CRC-tissue #60 and CRC-PDO tissue #60
belonging to passage 5 and 10, including most of the known major
cancer driver genes. (B) Immunohistochemical profile of FFPE
sections of organoids and corresponding tissues for CK19, CK20 and
CDX2 along with corresponding H&E staining. Magnification, ×40.
Scale bar, 50 µm. (C) Images of normal-PDO #8 and CRC-PDO #3
morphology with CHIR99021 and R-spondin (CR+) or without
(CR−). Scale bar, 400 µm. (D) Fluorescence
microscopy images of normal-PDO #8 and CRC-PDO #3. Blue, DAPI; red,
Ki-67; green, E-cadherin. Scale bar, 100 µm. (E) Morphology
of CRC-PDO #3 treated with SCFA after 3 days. Scale bar, 200
µm. (F) Relative CRC-PDO #3 size 3 days after treatment with
SCFA (n=3). Data are presented as mean ± SEM.
*P<0.05, **P<0.01. (G) Morphology of
normal-PDO #8 treated with SCFA after 3 days. Scale bar, 200
µm. (H) Relative normal-PDO #8 size 3 days after treatment
with SCFA (n=3). Data are presented as mean ± standard error of the
mean. *P<0.05, **P<0.01. ns, not
significant; H&E, hematoxylin and eosin; SCFA, short-chain
fatty acids; CRC, colorectal cancer; PDO, patient-derived organoid;
CK, cytokeratin. |
Butyrate enhances radiosensitivity in
CRC-PDO
Organoids co-treated with butyrate and IR
demonstrated a significant decrease in organoid size from 1.1-fold
to 0.6-fold as compared to those treated only with IR (P=0.04;
Fig. 2A and B). These observations
were supported by the MTT assay results, wherein a significant
reduction in organoid viability following butyrate and IR
combination treatment was observed (Fig. 2C). As cancer consists of both
tumorigenic and non-tumorigenic cancer cells, to further evaluate
the antitumor effect of butyrate, the number of tumor organoids in
the second passage were counted after PDO splitting. A total of 3
days after splitting, the number of formed tumor organoids
significantly decreased after butyrate and IR treatment compared
with that in the IR-only treatment group (Fig. 2D). These results suggested that
butyrate enhances the effect of radiation on CRC organoids. To
further elucidate if the loss of cell viability after butyrate and
IR exposure was accompanied by changes in cell cycle arrest, the
degree of EdU incorporation was measured by flow cytometry and it
was identified that 12% of the cells were in the S phase after IR
treatment, while only 5% were in the S phase after combination
treatment (P=0.003; Fig. 2E).
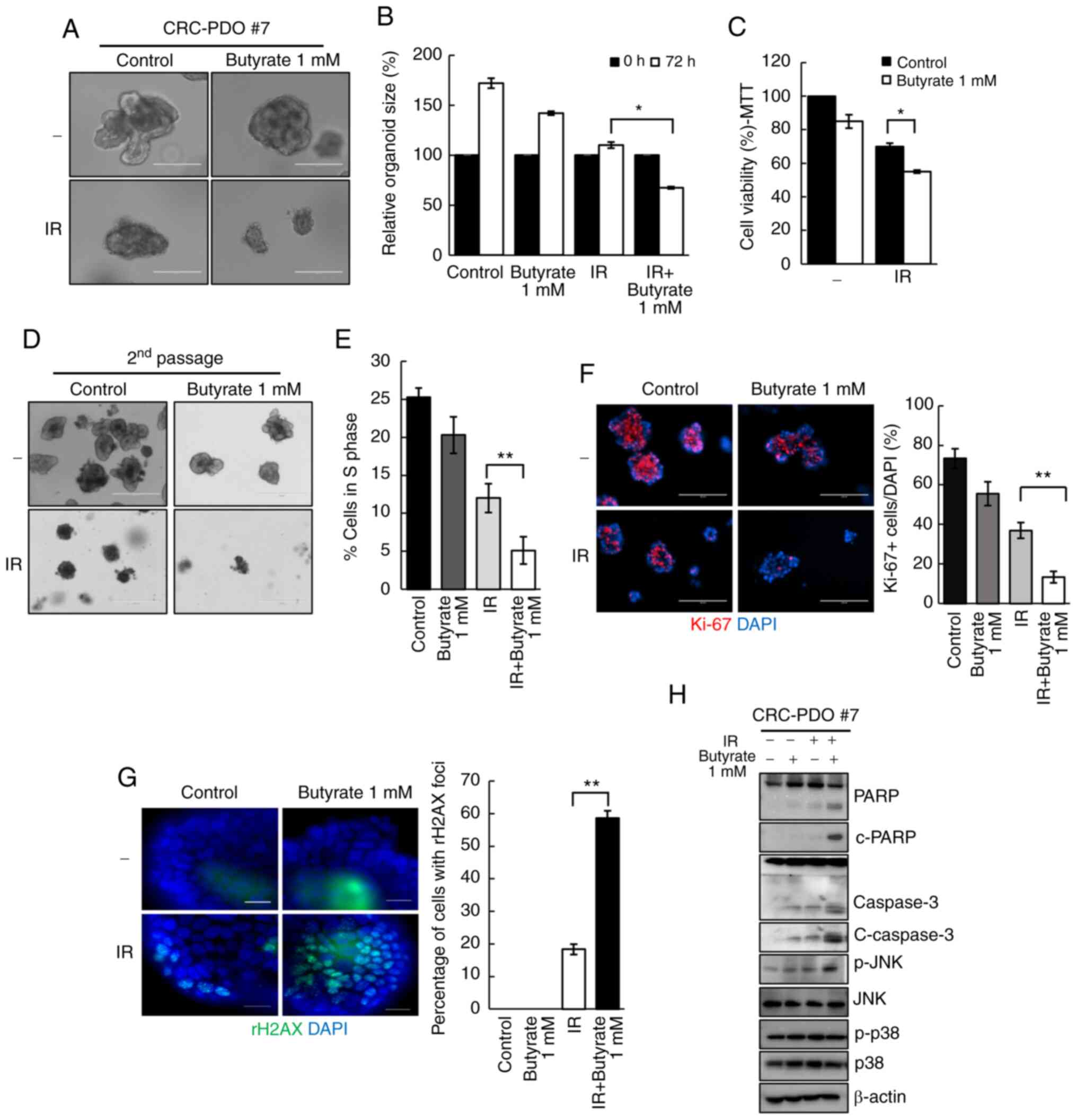 | Figure 2Butyrate enhances radiosensitivity in
CRC-PDOs. (A) Morphology and (B) organoid size (n=3) of CRC-PDO #7
irradiated thrice with 5 Gy with or without butyrate. Scale bar,
200 µm. (C) MTT cell viability assay of organoids described
in A (n=3). (D) Image of organoids after the second passage. Scale
bar, 400 µm. (E) Percentage of CRC-PDO in S phase (n=3). (F)
Left: Fluorescence microscopy images of organoids irradiated thrice
with 5 Gy with or without butyrate. Blue, DAPI; red, Ki-67. Scale
bar, 200 µm. Right: Statistical analysis representing
Ki-67-positive cells per DAPI staining cells (n=3). (G)
Fluorescence microscopy images showing γ-H2AX foci. Blue, DAPI;
green, γ-H2AX. Right: Statistical analysis representing γ-H2AX foci
(n=3). (H) Expression levels of PARP, c-PARP, caspase 3, c-caspase
3, p-JNK, JNK, p-p38 and p38 in the CRC-PDO. β-actin served as a
loading control. Data are presented as mean ± standard error of the
mean. *P<0.05, **P<0.01. CRC,
colorectal cancer; PDO, patient-derived organoid; PARP,
poly-ADP-ribose polymerase; c-, cleaved; p-, phosphorylated; IR,
irradiation. |
The effect of butyrate on the growth of CRC-PDOs was
then evaluated. Organoids after treatment with butyrate and IR had
a significantly lower number of Ki-67+ cells than those
treated with IR alone (P=0.005; Figs.
2F and S4A). To investigate
the effects of butyrate on radiation-induced DNA damage, γ-H2AX was
analyzed by western blotting and it was identified that combination
treatment with butyrate and radiation resulted in higher levels of
γ-H2AX compared with radiation alone at 6 and 24 h post-RT
(Fig. S4B). After irradiation,
18.3% of the cancer organoids were positive for γ-H2AX foci,
whereas the combination treatment induced 58.6% of organoids to
form γ-H2AX foci, indicating a significantly greater number of DSB
(P<0.001; Fig. 2G).
Next, to evaluate the cellular response to
radiation, the levels of apoptosis-related proteins were analyzed.
The levels of c-PARP and c-caspase 3, which are considered
hallmarks of apoptosis, were increased after IR and butyrate
treatment relative to the respective levels after IR treatment
alone. Moreover, p-JNK levels were increased after combination
treatments. There were no changes in the expression levels of p-p38
(Fig. 2H). These findings suggest
that butyrate acts as a radiosensitizer in CRC-PDO.
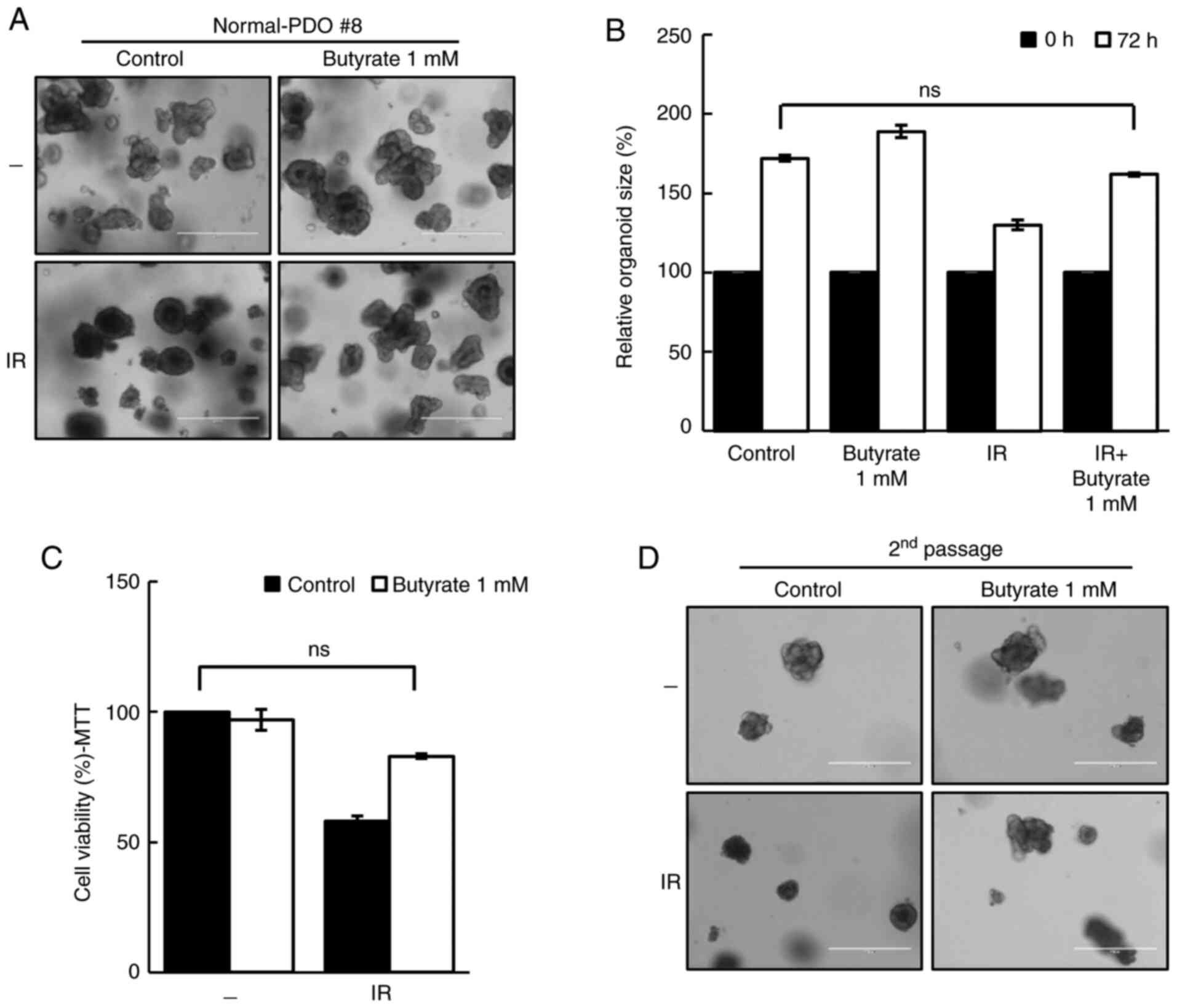
Butyrate does not act as a
radiosensitizer in normal-PDO
To evaluate the safety of butyrate, the response of
butyrate in normal-PDOs was further investigated. Although the
organoid size was reduced after irradiation, butyrate protected
normal-PDOs from the damaging effects of IR (Fig. 3A and B). The MTT assay and second
passage supported these results (Fig.
3C and D), thus suggesting that butyrate does not affect the
radiosensitivity in normal-PDO.
Butyrate enhances radiosensitivity via
the Warburg effect
Butyrate has previously been shown to selectively
suppress proliferation of cancer cell lines due to the difference
in the Warburg effect between normal colon cells and colon cancer
cells (37). Thus, to test whether
the Warburg effect may account for the differential
radiosensitizing effects of butyrate, the Warburg effect in
CRC-PDOs was inhibited by changing the culture media from
high-glucose (3.151 g/l) to low-glucose (1 g/l). Under these
conditions, glucose levels were too low to support aerobic
glycolysis, as evidenced by the detection of negligible levels of
lactate, i.e., the end-product of glycolysis (P=0.01; Fig. 4A). Without the Warburg effect in
the low-glucose condition, butyrate no longer affected the organoid
size and viability after irradiation (Fig. 4B and C). In addition, the number of
second passage organoids formed did not change after IR and
butyrate treatment under the low-glucose condition (Fig. 4D). As shown by EdU staining, the
proportion of S phase cells was reduced after IR and butyrate
treatment in high-glucose medium (P<0.001), but not in
low-glucose medium (Fig. 4E).
These results demonstrate that butyrate increases radiosensitivity
in CRC-PDOs in a Warburg effect-dependent manner.
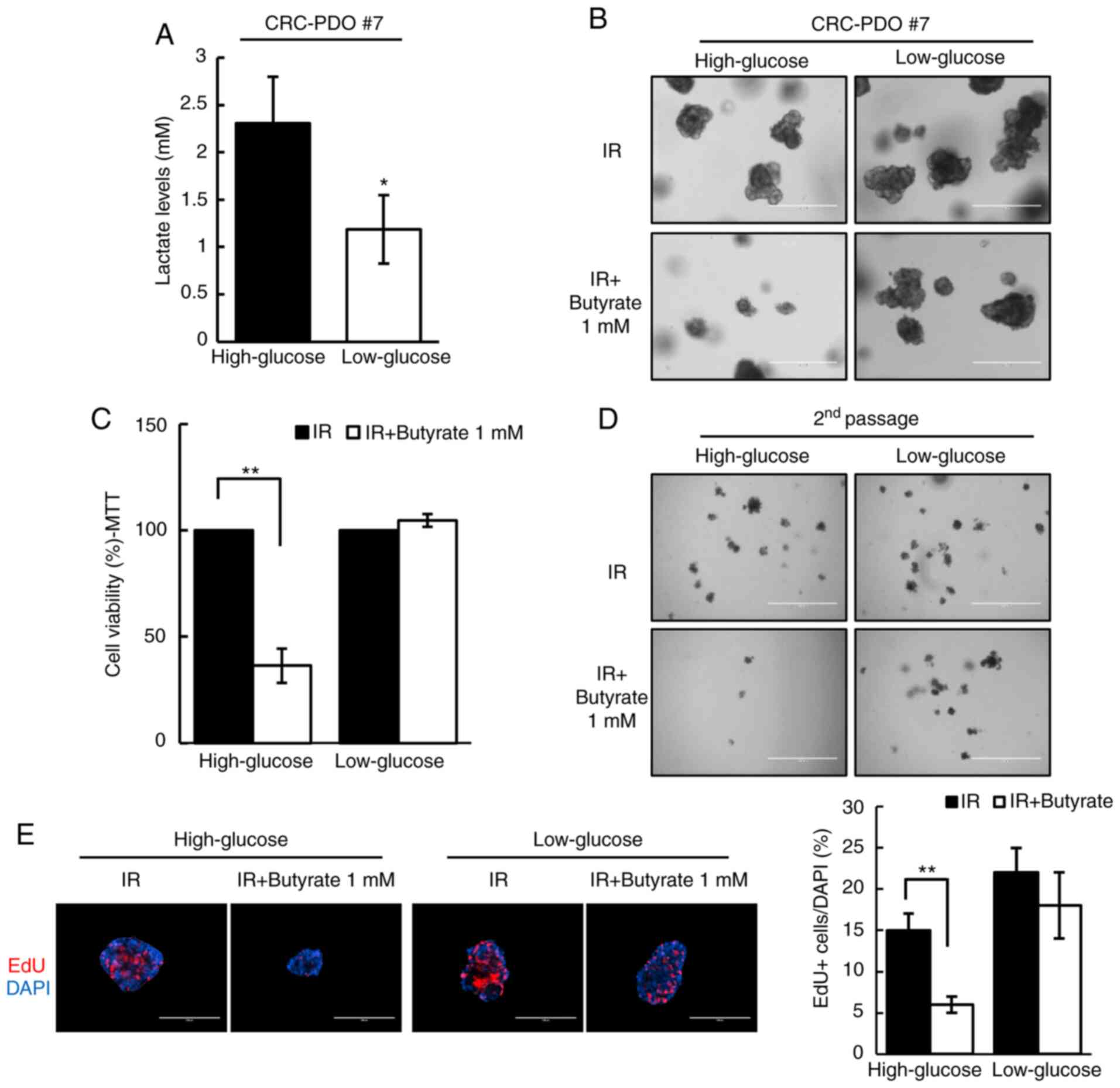 | Figure 4Radiosensitizing effects of butyrate
are dependent on glucose concentration. (A) Lactate levels in
CRC-PDO #7 grown in low- or high-glucose conditions (n=3). (B)
CRC-PDO #7 grown in low- or high-glucose conditions irradiated
thrice with 5 Gy with or without butyrate. Scale bar, 400
µm. (C) MTT cell viability assay of organoids described in
(B) (n=3). (D) Image of organoids after the second passage. Scale
bar, 1,000 µm. (E) Fluorescence microscopy images of EdU
incorporation in CRC-PDO #7 cultured under low- or high-glucose
conditions *P<0.05, **P<0.01. Scale
bar, 200 µm. Blue, DAPI; red, EdU. CRC, colorectal cancer;
PDO, patient-derived organoid; IR, irradiation. |
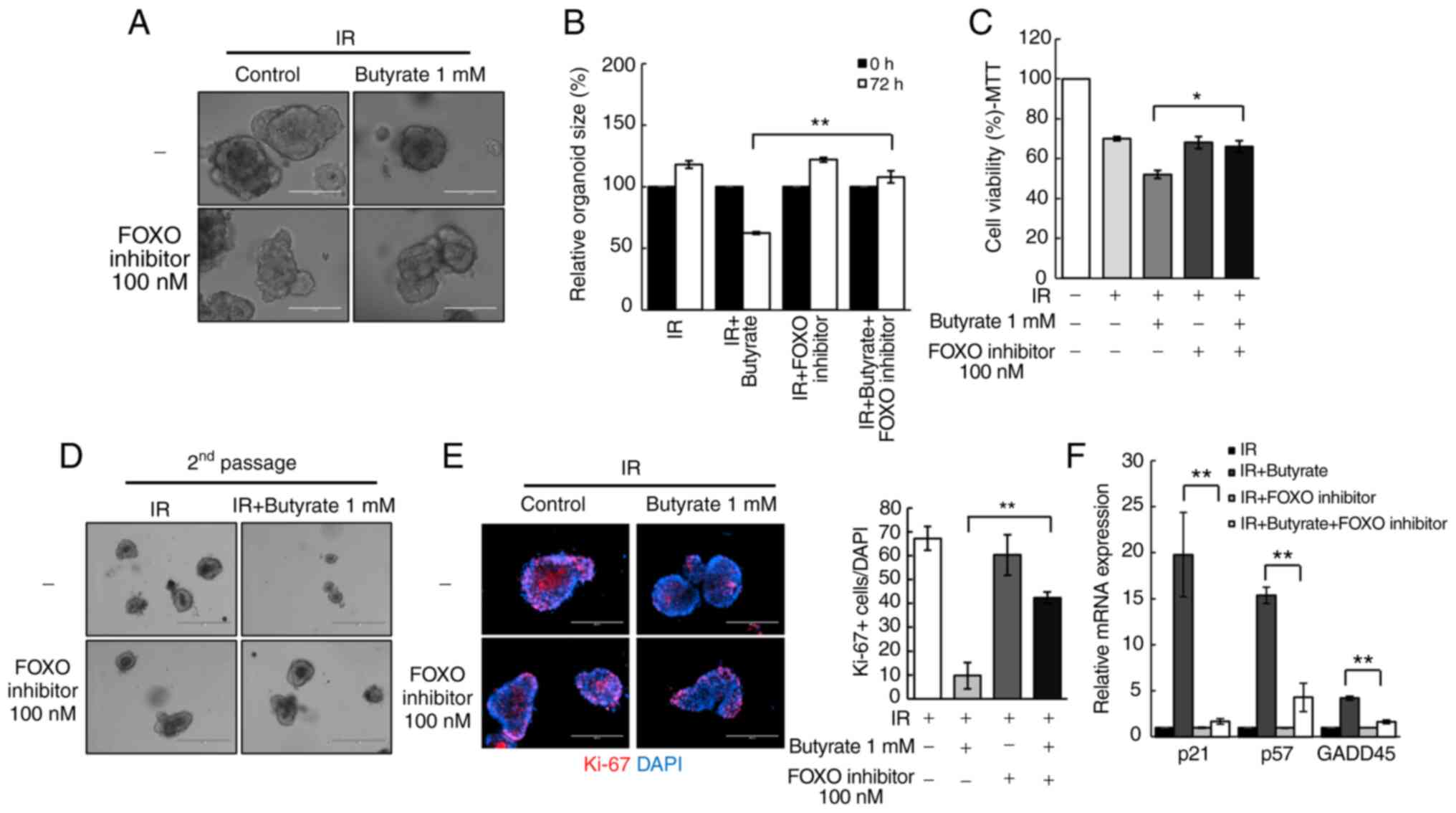
Butyrate regulates radio responses via
FOXO3A in CRC-PDOs
To investigate the mechanism of the potential
effects elicited by butyrate on radiosensitivity, the present study
focused on transcription factors that regulate cell cycle genes. In
particular, the influence of the transcription factor FOXO3A was
examined, as a previous study suggested that butyrate regulates its
transcriptional activity through HDAC inhibition (29). Treatment of CRC-PDOs with a FOXO
inhibitor rescued the radiosensitizing effect of butyrate in
CRC-PDOs (P<0.001; Fig. 5A and
B). In addition, the MTT assay showed that the significant
reduction in organoid viability following combination treatment,
was also rescued by the FOXO inhibitor (P=0.03; Fig. 5C) along with the
organoid-regenerating capacity analyzed by passaging (Fig. 5D). The FOXO inhibitor also reversed
the proliferation inhibitory effects of butyrate after irradiation
(P<0.001; Fig. 5E). RT-qPCR was
conducted to examine the expression levels of cell cycle-related
genes regulated by FOXO3A, namely GADD45, p21, and p57 (38,39).
Results show that butyrate and IR increased the mRNA expression
levels of GADD45 (P<0.001), p21 (P<0.001) and p57
(P<0.001) in CRC-PDOs, which were all decreased after treatment
with the FOXO inhibitor (Fig. 5F).
These findings suggest that butyrate increases radiosensitivity
through induction of p21, p57, and GADD45 regulated by FOXO3A.
Radiosensitizing effect of butyrate is
dependent on FOXO3A expression in CRC-PDOs
Although there was an overall reduction in organoid
size following combined butyrate and IR treatment, no difference
was observed in some cases (here-after referred to as
non-responsive CRC-PDOs; Fig. 6A).
EdU staining was, therefore, performed to compare the proliferative
capacity of responsive and non-responsive CRC-PDOs. After treatment
with butyrate and IR, the non-responsive CRC-PDO group had more
EdU-incorporated cells than the responsive CRC-PDO group
(P<0.001; Fig. 6B). Moreover,
responsive CRC-PDOs, and their original tissues, showed stronger
nuclear staining for FOXO3A than non-responsive-CRC-PDOs (Fig. 6C). Western blot analysis further
showed that all five responsive CRC-PDO cases exhibited increased
expression of FOXO3A compared with the non-responsive CRC-PDOs
(Fig. 6D), and the intensity of
FOXO3A expression was significantly increased compared with that in
the three non-responsive CRC-PDO (P=0.0084; Fig. 6E). RT-qPCR analysis further showed
that butyrate and IR combination treatment did not affect the
expression levels of p21 (P<0.001), p57 (P<0.001) or GADD45
(P=0.048) in non-responsive CRC-PDOs in contrast to the effects
observed in responsive CRC-PDOs (Fig.
6F). These results suggest that the radiosensitizing effect of
butyrate may be dependent on the level of FOXO3A expression in
CRC-PDOs.
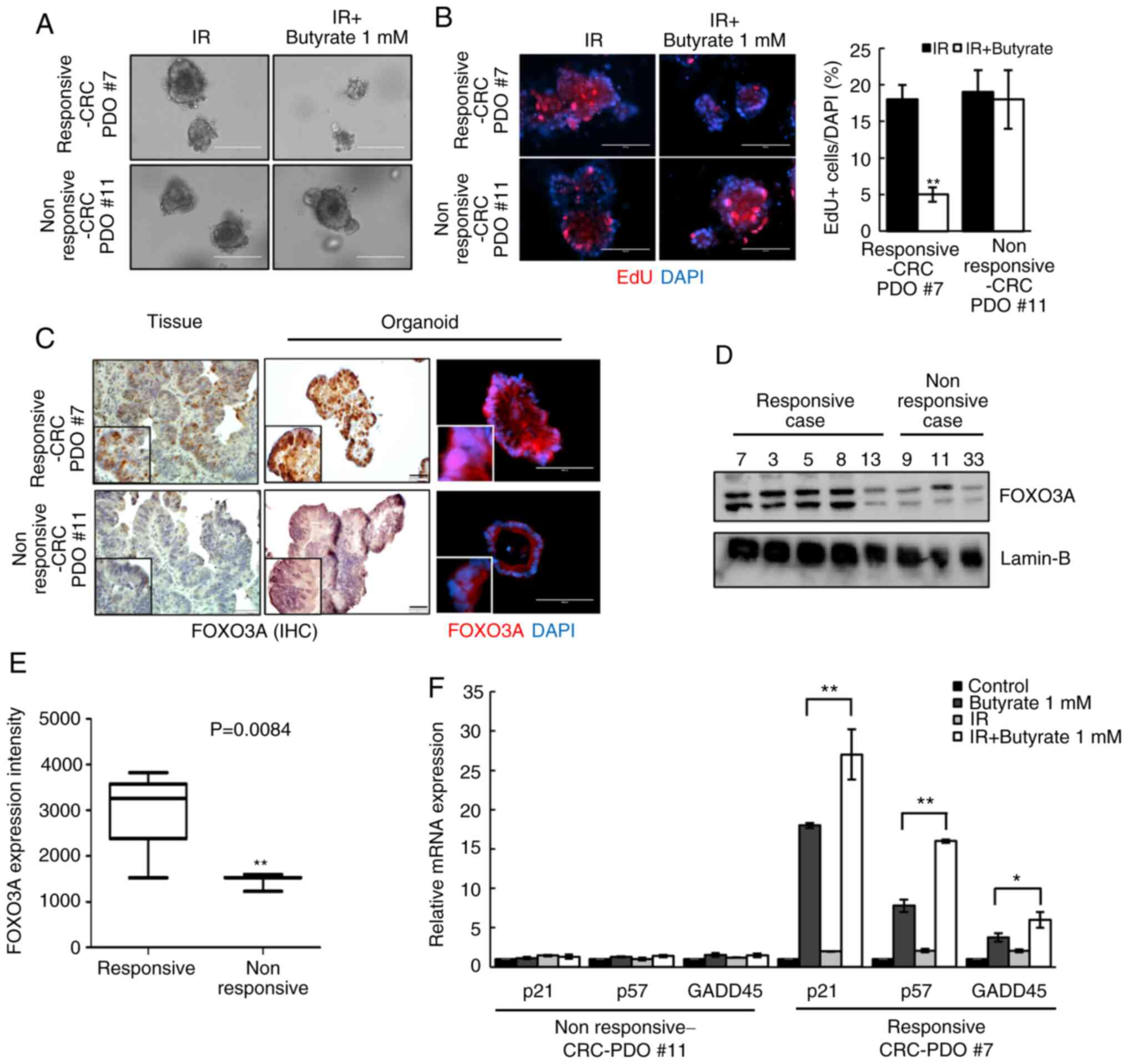 | Figure 6Radiosensitizing effects of butyrate
are dependent on FOXO3A expression level in CRC-PDOs. (A) Images of
CRC-PDO #7 irradiated thrice with 5 Gy with or without butyrate.
Scale bar, 200 µm. (B) Fluorescence microscopy images of EdU
incorporation into responsive-CRC-PDO #7 and non-responsive-CRC-PDO
#11 after irradiation with butyrate. Scale bar, 200 µm.
Blue, DAPI; red, EdU. (C) IHC and fluorescence microscopy images of
FOXO3a in tissues, responsive-CRC-PDO #7 and non-responsive-CRC-PDO
#11. Immunohistochemistry scale bar, 50 µm; fluorescence
microscopy scale bar, 100 µm. Blue, DAPI; red, FOXO3A. (D)
Protein expression of FOXO3A in the nuclear fraction of CRC-PDO.
Lamin-B served as a marker for the nuclear fraction. (E)
Box-and-whisker plots of FOXO3A expression in five cases of
responsive-CRC-PDO and three cases of non-responsive-CRC-PDO. (F)
CRC-PDO #7 was harvested at 24 h and analyzed by reverse
transcription-quantitative PCR for cell cycle-related genes. The
expression level of the target mRNA was normalized to that of GAPDH
mRNA (n=3). Data are presented as mean ± standard error of the
mean. *P<0.05, **P<0.01. CRC,
colorectal cancer; PDO, patient-derived organoid; IR, irradiation;
IHC, immunohistochemistry. |
Discussion
The present study demonstrated that butyrate has a
potent radiosensitizing effect on CRC-PDOs by increasing FOXO3A
transcriptional activity and inducing cell cycle arrest regulated
by p21, p57 and GADD45. However, butyrate protects against
IR-induced damage in normal colon PDOs. To the best of our
knowledge, this is the first study to suggest a role for butyrate
in regulating the radio response and cellular function in CRC-PDOs.
Previously, the underlying mechanism responsible for the anticancer
function of butyrate was reportedly associated with mediating
HDAC-dependent transcription activation (40,41).
In the normal colon, butyrate is used as an energy source. However,
since cancer cells rely on glucose as an energy source, butyrate
accumulates in the nucleus to inhibit proliferation, thus inducing
apoptosis only in cancer cells (13,37,42,43).
Hence, targeting tumor metabolism has been described as a potential
strategy for clinical applications (43,44).
This specificity makes butyrate particularly attractive as a novel
candidate radiosensitizer, as it would have minimal adverse effects
on normal cells.
The present study further demonstrated that certain
CRC-PDOs found to be non-responsive to butyrate and IR combination
treatment had relatively lower expression levels of FOXO3A compared
with responsive CRC-PDOs. Although responsive CRC-PDOs exhibited
higher expression of FOXO3A compared with those non-responsive,
variable expression was observed in both groups, highlighting the
heterogeneity of patient samples that causes natural variation
attributable to unique patient characteristics. Nevertheless, since
butyrate appears to enhance radiosensitivity through FOXO3A, its
expression may serve as a potential biomarker to predict the
effectiveness of butyrate on radiotherapy.
FOXO transcription factors are emerging as critical
transcriptional integrators among pathways regulating
differentiation, proliferation, survival and the cell cycle
(45-47). A previous study suggested that
AZD6244 enhances the expression of FOXO3A, and suppresses colon
cancer cell proliferation (48),
while others reported that FOXO3A enhances the radiosensitivity of
cancer cells through regulation of apoptosis and AMP-activated
kinase (AMPK) signaling (49,50).
Thus, FOXO3A is a vital regulator enhancing radiosensitivity. The
present study further found that the cell cycle-related genes
regulated by FOXO3A (p21, p57 and GADD45) were involved in the
mechanism through which butyrate inhibits cell growth, and cell
cycling in CRC-PDOs. These genes have been shown to negatively
regulate the cancer cell cycle (41-43).
Moreover, GADD45α inhibits the nitric oxide-regulated cytoplasmic
localization of APE1, thereby enhancing the radiosensitivity of
cervical cancer cells (51); while
the negative regulation of p57 by miRNA221/222 may also contribute
to radioresistance (52).
Therefore, butyrate enhanced radiosensitivity by inhibiting HDAC
and the FOXO3A/p21, p57 and GADD45 axis.
It was identified that butyrate enhanced the
radiosensitivity via the Warburg effect. However, only the lactate
levels in CRC-PDO were examined. AMPK negatively regulates the
Warburg effect in cancer cells and suppresses the growth of tumors
in vivo (53). Hence, it is
necessary to evaluate other signaling pathways, such as
AMP-activated kinase activation. Additionally, FOXO3A was recently
shown to be negatively correlated with the expression of a number
of glycolysis-associated genes and to inhibit glucose metabolism
and tumor cell proliferation in melanoma (54). Furthermore, transcriptional
activation of FOXO3A inhibits the Warburg effect in glioblastoma
cells (55). Together these
studies suggest that FOXO3A is associated with the Warburg effect,
which warrants further investigation.
Cancer cell lines have long served as the primary
work-horse model in radiobiology research. Indeed, some of the most
well-known radiosensitizers such as 5-FU, curcumin and docetaxel
(56-58) were discovered in cancer cell lines;
however, these effects were not reproduced in patients. These
inconsistent results are due to the highly heterogenous nature of
tumors, which are composed of a mixture of sub-clones and various
cell types, a characteristic that is not accurately reflected in
cancer cell lines. Thus, our PDO culture system can be exploited
for functional studies on radio responses within individual
patients that cannot be achieved by cell lines or animal studies.
Hence, we propose that PDOs will be a valuable tool for directly
testing the radiosensitivity of a tumor in a personalized treatment
approach.
Herein, it was demonstrated that 1 mM butyrate
sensitizes CRC organoids to the cytotoxic effects of radiotherapy,
while eliciting normal cell-protective effects. However, higher
concentrations of butyrate have limited applications due to
toxicity on normal intestinal cells (37). Hence, it is necessary to evaluate
the optimal dose before butyrate is used in clinical settings as a
radiosensitizer.
In conclusion, the results of the present study
suggest that butyrate suppresses proliferation of three-dimensional
CRC organoids and enhances radiation-induced cell death in CRC
organoids through FOXO3A. However, butyrate does not increase
radiation-induced cell death after irradiation in normal organoids.
Thus, it may enhance the efficacy of radiotherapy while protecting
the normal mucosa.
Supplementary Data
Acknowledgments
The biospecimens and data used in this study were
provided by the Radiation Tissue Resources Bank of Korea Cancer
Center Hospital (TB-2016-05).
Abbreviations:
|
CRC
|
colorectal cancer
|
|
HDAC
|
histone deacetylase
|
|
PDO
|
patient-derived organoid
|
|
SAHA
|
suberoylanilide hydroxamic acid
|
|
SCFAs
|
short-chain fatty acids
|
Funding
This study was supported by a grant of the Korea
Institute of Radiological and Medical Sciences, funded by the
Ministry of Science and ICT, Republic of Korea (grant no.
50542-2019).
Availability of data and materials
The datasets used or analyzed during this study are
available from the corresponding author upon reasonable
request.
Authors' contributions
MP, YH, and YK were involved in the conception and
design of the study. MP, JK, and HJS performed the experiments.
SMM, SBK and USS provided resources and analyzed the data. MP, YHH,
and YK wrote the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
This study was approved by Ethics Committee of Korea
Cancer Center Hospital (approval no. KIRAMS-2017-07-001). All
research was performed in accordance with the approved guidelines
and regulations of the institution. All samples were obtained from
patients who had provided written informed consent for the use of
their tissues for the purposes of research after the operation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD, Goding Sauer A,
Fedewa SA, Butterly LF, Anderson JC, Cercek A, Smith RA and Jemal
A: Colorectal cancer statistics, 2020. CA Cancer J Clin.
70:145–164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guren MG, Dueland S, Skovlund E, Fossa SD,
Poulsen JP and Tveit KM: Quality of life during radiotherapy for
rectal cancer. Eur J Cancer. 39:587–594. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang H, Mu X, He H and Zhang XD: Cancer
radiosensitizers. Trends Pharmacol Sci. 39:24–48. 2018. View Article : Google Scholar
|
|
4
|
Wolpin BM, Meyerhardt JA, Mamon HJ and
Mayer RJ: Adjuvant treatment of colorectal cancer. CA Cancer J
Clin. 57:168–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Willett CG, Duda DG, di Tomaso E, Boucher
Y, Ancukiewicz M, Sahani DV, Lahdenranta J, Chung DC, Fischman AJ,
Lauwers GY, et al: Efficacy, safety, and biomarkers of neoadjuvant
bevacizumab, radiation therapy, and fluorouracil in rectal cancer:
A multidisciplinary phase II study. J Clin Oncol. 27:3020–3026.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Crane CH, Eng C, Feig BW, Das P, Skibber
JM, Chang GJ, Wolff RA, Krishnan S, Hamilton S, Janjan NA, et al:
Phase II trial of neoadjuvant bevacizumab, capecitabine, and
radiotherapy for locally advanced rectal cancer. Int J Radiat Oncol
Biol Phys. 76:824–830. 2010. View Article : Google Scholar
|
|
7
|
Tomiak A, Vincent M, Kocha W, Taylor M,
Winquist E, Keith B, Sawyer M, Griffeth S, Whiston F and Stitt L:
Standard dose (Mayo regimen) 5-fluorouracil and low dose folinic
acid: Prohibitive toxicity? Am J Clin Oncol. 23:94–98. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meta-Analysis Group In Cancer; Levy E,
Piedbois P, Buyse M, Pignon JP, Rougier P, Ryan L, Hansen R, Zee B,
Weinerman B, et al: Toxicity of fluorouracil in patients with
advanced colorectal cancer: Effect of administration schedule and
prognostic factors. J Clin Oncol. 16:3537–3541. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cho M, Carter J, Harari S and Pei Z: The
interrelationships of the gut microbiome and inflammation in
colorectal carcinogenesis. Clin Lab Med. 34:699–710. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arthur JC and Jobin C: The struggle
within: Microbial influences on colorectal cancer. Inflamm Bowel
Dis. 17:396–409. 2011. View Article : Google Scholar
|
|
11
|
Kuwahara A: Contributions of colonic
short-chain fatty acid receptors in energy homeostasis. Front
Endocrinol (Lausanne). 5:1442014. View Article : Google Scholar
|
|
12
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bultman SJ: Molecular pathways:
Gene-environment interactions regulating dietary fiber induction of
proliferation and apoptosis via butyrate for cancer prevention.
Clin Cancer Res. 20:799–803. 2014. View Article : Google Scholar :
|
|
14
|
Li Q, Cao L, Tian Y, Zhang P, Ding C, Lu
W, Jia C, Shao C, Liu W, Wang D, et al: Butyrate suppresses the
proliferation of colorectal cancer cells via targeting pyruvate
kinase M2 and metabolic reprogramming. Mol Cell Proteomics.
17:1531–1545. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Folkvord S, Ree AH, Furre T, Halvorsen T
and Flatmark K: Radiosensitization by SAHA in experimental
colorectal carcinoma models-in vivo effects and relevance of
histone acetylation status. Int J Radiat Oncol Biol Phys.
74:546–552. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Wong P, Radany E and Wong JY: HDAC
inhibitor, valproic acid, induces p53-dependent radiosensitization
of colon cancer cells. Cancer Biother Radiopharm. 24:689–699. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Flatmark K, Nome RV, Folkvord S, Bratland
A, Rasmussen H, Ellefsen MS, Fodstad Ø and Ree AH:
Radiosensitization of colorectal carcinoma cell lines by histone
deacetylase inhibition. Radiat Oncol. 1:252006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Failli A, Consolini R, Legitimo A, Spisni
R, Castagna M, Romanini A, Crimaldi G and Miccoli P: The challenge
of culturing human colorectal tumor cells: Establishment of a cell
culture model by the comparison of different methodological
approaches. Tumori. 95:343–347. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dangles-Marie V, Pocard M, Richon S,
Weiswald LB, Assayag F, Saulnier P, Judde JG, Janneau JL, Auger N,
Validire P, et al: Establishment of human colon cancer cell lines
from fresh tumors versus xenografts: Comparison of success rate and
cell line features. Cancer Res. 67:398–407. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
van de Wetering M, Francies HE, Francis
JM, Bounova G, Iorio F, Pronk A, van Houdt W, van Gorp J,
Taylor-Weiner A, Kester L, et al: Prospective derivation of a
living organoid biobank of colorectal cancer patients. Cell.
161:933–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Walsh AJ, Cook RS, Sanders ME, Aurisicchio
L, Ciliberto G, Arteaga CL and Skala MC: Quantitative optical
imaging of primary tumor organoid metabolism predicts drug response
in breast cancer. Cancer Res. 74:5184–5194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Clevers H: Modeling development and
disease with organoids. Cell. 165:1586–1597. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Amin MB, Edge S, Greene F, Byrd DR,
Brookland RK, Washington MK, Gershenwald JE, Compton CC, Hess KR,
Sullivan DC, et al: AJCC Cancer Staging Manual. Springer
International Publishing; 2017, View Article : Google Scholar
|
|
24
|
Xie BY and Wu AW: Organoid culture of
isolated cells from patient-derived tissues with colorectal cancer.
Chin Med J (Engl). 129:2469–2475. 2016. View Article : Google Scholar
|
|
25
|
Sato T, Stange DE, Ferrante M, Vries RG,
Van Es JH, Van den Brink S, Van Houdt WJ, Pronk A, Van Gorp J,
Siersema PD and Clevers H: Long-term expansion of epithelial
organoids from human colon, adenoma, adenocarcinoma, and Barrett's
epithelium. Gastroenterology. 141:1762–1772. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fujii M, Shimokawa M, Date S, Takano A,
Matano M, Nanki K, Ohta Y, Toshimitsu K, Nakazato Y, Kawasaki K, et
al: A colorectal tumor organoid library demonstrates progressive
loss of niche factor requirements during tumorigenesis. Cell Stem
Cell. 18:827–838. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cancer Genome Atlas Network: Comprehensive
molecular characterization of human colon and rectal cancer.
Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Grabinger T, Luks L, Kostadinova F,
Zimberlin C, Medema JP, Leist M and Brunner T: Ex vivo culture of
intestinal crypt organoids as a model system for assessing cell
death induction in intestinal epithelial cells and enteropathy.
Cell Death Dis. 5:e12282014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kaiko GE, Ryu SH, Koues OI, Collins PL,
Solnica-Krezel L, Pearce EJ, Pearce EL, Oltz EM and Stappenbeck TS:
The colonic crypt protects stem cells from microbiota-derived
metabolites. Cell. 167:1708–1720. 2016. View Article : Google Scholar
|
|
30
|
Yin X, Farin HF, van Es JH, Clevers H,
Langer R and Karp JM: Niche-independent high-purity cultures of
Lgr5+ intestinal stem cells and their progeny. Nat Methods.
11:106–112. 2014. View Article : Google Scholar :
|
|
31
|
Kinner A, Wu W, Staudt C and Iliakis G:
Gamma-H2AX in recognition and signaling of DNA double-strand breaks
in the context of chromatin. Nucleic Acids Res. 36:5678–5694. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bozzi F, Mogavero A, Varinelli L, Belfiore
A, Manenti G, Caccia C, Volpi CC, Beznoussenko GV, Milione M, Leoni
V, et al: MIF/CD74 axis is a target for novel therapies in colon
carcinomatosis. J Exp Clin Cancer Res. 36:162017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miyoshi H and Stappenbeck TS: In vitro
expansion and genetic modification of gastrointestinal stem cells
in spheroid culture. Nat Protoc. 8:2471–2482. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Park M, Yoon HJ, Kang MC, Kwon J and Lee
HW: MiR-338-5p enhances the radiosensitivity of esophageal squamous
cell carcinoma by inducing apoptosis through targeting survivin.
Sci Rep. 7:109322017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
36
|
Mayakonda A, Lin DC, Assenov Y, Plass C
and Koeffler HP: Maftools: Efficient and comprehensive analysis of
somatic variants in cancer. Genome Res. 28:1747–1756. 2018.
View Article : Google Scholar :
|
|
37
|
Donohoe DR, Collins LB, Wali A, Bigler R,
Sun W and Bultman SJ: The warburg effect dictates the mechanism of
butyrate-mediated histone acetylation and cell proliferation. Mol
Cell. 48:612–626. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang H and Tindall DJ: Dynamic FoxO
transcription factors. J Cell Sci. 120:2479–2487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang X, Tang N, Hadden TJ and Rishi AK:
Akt, FoxO and regulation of apoptosis. Biochim Biophys Acta.
1813:1978–1986. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Thangaraju M, Cresci GA, Liu K, Ananth S,
Gnanaprakasam JP, Browning DD, Mellinger JD, Smith SB, Digby GJ,
Lambert NA, et al: GPR109A is a G-protein-coupled receptor for the
bacterial fermentation product butyrate and functions as a tumor
suppressor in colon. Cancer Res. 69:2826–2832. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tan HT, Tan S, Lin Q, Lim TK, Hew CL and
Chung MC: Quantitative and temporal proteome analysis of
butyrate-treated colorectal cancer cells. Mol Cell Proteomics.
7:1174–1185. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pouillart PR: Role of butyric acid and its
derivatives in the treatment of colorectal cancer and
hemoglobinopathies. Life Sci. 63:1739–1760. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Boroughs LK and DeBerardinis RJ: Metabolic
pathways promoting cancer cell survival and growth. Nat Cell Biol.
17:351–359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vernieri C, Casola S, Foiani M,
Pietrantonio F, de Braud F and Longo V: Targeting cancer
metabolism: Dietary and pharmacologic interventions. Cancer Discov.
6:1315–1333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Birkenkamp KU and Coffer PJ: Regulation of
cell survival and proliferation by the FOXO (Forkhead box, class O)
subfamily of Forkhead transcription factors. Biochem Soc Trans.
31:292–297. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Schmidt M, Fernandez de Mattos S, van der
Horst A, Klompmaker R, Kops GJ, Lam EW, Burgering BM and Medema RH:
Cell cycle inhibition by FoxO forkhead transcription factors
involves down-regulation of cyclin D. Mol Cell Biol. 22:7842–7852.
2002. View Article : Google Scholar
|
|
47
|
Wang Y, Zhou Y and Graves DT: FOXO
transcription factors: Their clinical significance and regulation.
Biomed Res Int. 2014:9253502014.PubMed/NCBI
|
|
48
|
Yang JY, Chang CJ, Xia W, Wang Y, Wong KK,
Engelman JA, Du Y, Andreeff M, Hortobagyi GN and Hung MC:
Activation of FOXO3a is sufficient to reverse mitogen-activated
protein/extra-cellular signal-regulated kinase kinase inhibitor
chemoresistance in human cancer. Cancer Res. 70:4709–4718. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang JY, Xia W and Hu MC: Ionizing
radiation activates expression of FOXO3a, Fas ligand, and Bim, and
induces cell apoptosis. Int J Oncol. 29:643–648. 2006.PubMed/NCBI
|
|
50
|
Yang Y, Pan W, Zhang S, Cao Y, Cheng H,
Chen J and Sun X: Metformin can enhance the radiosensitivity of
cholangiocarcinoma through AMPK-FOXO3a axis. Int J Clin Exp Med.
9:13539–13550. 2016.
|
|
51
|
Li Q, Wei X, Zhou ZW, Wang SN, Jin H, Chen
KJ, Luo J, Westover KD, Wang JM, Wang D, et al: GADD45α sensitizes
cervical cancer cells to radiotherapy via increasing cytoplasmic
APE1 level. Cell Death Dis. 9:5242018. View Article : Google Scholar
|
|
52
|
Milas L, Akimoto T, Hunter NR, Mason KA,
Buchmiller L, Yamakawa M, Muramatsu H and Ang KK: Relationship
between cyclin D1 expression and poor radioresponse of murine
carcinomas. Int J Radiat Oncol Biol Phys. 52:514–521. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Faubert B, Boily G, Izreig S, Griss T,
Samborska B, Dong Z, Dupuy F, Chambers C, Fuerth BJ, Viollet B, et
al: AMPK is a negative regulator of the warburg effect and
suppresses tumor growth in vivo. Cell Metab. 17:113–124. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Dong Z, Yang J, Li L, Tan L, Shi P, Zhang
J, Zhong X, Ge L, Wu Z and Cui H: FOXO3aSIRT6 axis suppresses
aerobic glycolysis in melanoma. Int J Oncol. 56:728–742.
2020.PubMed/NCBI
|
|
55
|
Dong Z, Zhong X, Lei Q, Chen F and Cui H:
Transcriptional activation of SIRT6 via FKHRL1/FOXO3a inhibits the
warburg effect in glioblastoma cells. Cell Signal. 60:100–113.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lawrence TS, Blackstock AW and McGinn C:
The mechanism of action of radiosensitization of conventional
chemotherapeutic agents. Semin Radiat Oncol. 13:13–21. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chendil D, Ranga RS, Meigooni D,
Sathishkumar S and Ahmed MM: Curcumin confers radiosensitizing
effect in prostate cancer cell line PC-3. Oncogene. 23:1599–1607.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dunne AL, Mothersill C, Robson T, Wilson
GD and Hirst DG: Radiosensitization of colon cancer cell lines by
docetaxel: Mechanisms of action. Oncol Res. 14:447–454. 2004.
View Article : Google Scholar : PubMed/NCBI
|