Introduction
Oral squamous cell carcinoma (OSCC) is a destructive
and lethal malignant tumour that accounts for >90% of all oral
cancers and is the sixth most common type of cancer worldwide
(1). Due to its anatomical
location, OSCC affects patients seriously, and most of them have
poor prognosis. Risk factors for OSCC include alcohol consumption,
tobacco smoking and adverse irritation of the oral mucosa (2,3).
Although great progress has been made in the treatment of OSCC, the
5-year survival rate has not improved much over the past two
decades (4,5). Therefore, further understanding of
the pathogenesis of OSCC may help to predict cancer progression and
provide novel molecular targets for cancer therapy.
MicroRNAs (miRNAs/miRs) belong to an endogenous
class of small non-coding RNAs that function through inhibiting the
translation levels of the target mRNAs (6,7).
Accumulating evidence has shown that miRNAs regulate cell
proliferation and apoptosis, and play key roles in OSCC
tumorigenesis, including miR-375, miR-139-5p, miR-155 and miR-21
(8-11). To date, downregulation of miR-198
has been found in various malignant tumours, including breast
cancer, lung cancer and hepatocellular carcinoma (12-14). Furthermore, miR-198 is an
independent prognostic factor in gastric cancer and glioma
(15). However, the role of
miR-198 in OSCC has not been fully investigated.
Epithelial-mesenchymal transition (EMT) is a
biological process in which epithelial cells transform into cells
with a mesenchymal phenotype through specific procedures. EMT plays
an important role in cancer metastasis. In the present study, the
expression of miR-198 in OSCC was examined. Furthermore, the
effects of miR-198 on the proliferation, invasion and EMT of OSCC
cells was analysed and the associated underlying mechanisms were
explored. These findings suggested that miR-198 acts as a potential
biomarker for OSCC.
Materials and methods
Tissue samples
A total of 80 OSCC tissues and adjacent non-tumour
tissues from patients at the School and Hospital of Stomatology,
China Medical University (Shenyang, China) between January 2013 and
January 2015 were selected. Patients with malignancies in other
organs and those who had received any anti-cancer therapy were
excluded. Clinical data including age, sex, primary tumour site,
differentiation, tumour node metastasis (TNM) classification and
recurrence were obtained from pathological and clinical records.
The ethics committee of the School and Hospital of Stomatology,
China Medical University approved the present study and written
informed consent was obtained from patients providing tissue
specimens.
Cell culture
Human OSCC cell lines (Cal-27, SCC-9 and SCC-25) and
human keratinocytes cell line (HaCaT cells) were purchased from the
American Type Culture Collection, and were maintained in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% foetal bovine
serum (FBS) and a 1% penicillin-streptomycin solution in a
humidified atmosphere containing 5% CO2 at 37°C. The
cell culture medium and FBS were purchased from Gibco (Thermo
Fisher Scientific, Inc.). The HaCaT cell line was originated from
normal skin tissue. The normal oral mucosal epithelial cell line is
not widely available, and the structure of the skin is very similar
to the structure of the oral mucosa, so HaCaT cells were used as
the control, as previously described (16,17). The HaCaT cell line was
authenticated by STR sequencing.
Cell transfection
miR-198 mimics, small interfering RNA (siRNA/si-)
targeting cyclin-dependent kinase 4 (CDK4) and negative controls
(NCs) were designed and synthesised by Shanghai GenePharma Co.,
Ltd. Cells (Cal-27 and SCC-9) were seeded into 96-well plates in
antibiotic-free growth medium at a density of 4×103
cells/well. miR-198 mimics (100 nmol/µl; 5′-GGU CCA GAG GGG
AGA UAG GUU C-3′) and the control (100 nmol/µl; 5′-UUC UCC
GAA CGU GUC ACG UAA U-3′) were transfected when the cells reached
70-80% confluence. si-CDK4 (80 nmol/µl; sense, 5′-CAG UUC
GUG AGG UGC UUU AC-3′ and antisense, 5′-GUA A AG CCA CCUC ACG AAC
UG-3′) and si-Ctrl (80 nmol/µl; sense, 5′-UUC UCC GAG CGU
GUC ACG UTT-3′ and antisense, 5′-ACG UGA CAC GUU CGG AGA ATT-3′)
were transfected using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) when cells reached 30-40%
confluence, in accordance with the manufacturer's protocol. By
inserting CDK4 cDNA into the pcDNA3.1 vector (Shanghai GenePharma
Co., Ltd.), the CDK4 overexpression plasmid (pcDNA-CDK4) was
generated and its sequence was confirmed by Shanghai GenePharma
Co., Ltd. An empty pcDNA3.1 vector was used as the NC. Using
Lipofectamine 3000, pcDNA-CDK4 (500 ng/µl) was transfected
into cells. After incubation at 37°C for 6 h, the culture medium
was changed. After transfection for 48 h, the subsequent
experimentation was carried out. Transfection efficiency was
analysed via reverse transcription-quantitative PCR (RT-qPCR).
RT-qPCR
Total RNA was extracted from frozen tissues and
Cal-27, SCC-9 and HaCat cells using TRIzol® (Takara
Biotechnology Co., Ltd.) according to the manufacturer's protocol.
Using standard spectrophotometric methods, RNA concentration was
quantified and its purity was determined.
To examine miR-198 expression, RNA (1 µg) was
reverse transcribed using Hairpin-it™ miRNA and a U6 snRNA
Normalisation Kit (Shanghai GenePharma Co., Ltd.). The RT reaction
system contained the following: 4 µl 5X MMLV RT buffer, 0.75
µl dNTP, 1.2 µl miRNA and U6 snRNA RT primer mix, 0.2
µl MML reverse transcriptase (200 U/µl)2,
1 µg RNA and RNase-free H2O, which added up to 20
µl. The following conditions were employed for RT: 25°C for
30 min, 42°C for 30 min and 85°C for 5 min. The qPCR reaction
system contained: 10 µl 2X Real-time PCR Master Mix, 0.4
µl miRNA and U6 snRNA specific primer set (10
µM)1, 0.4 µl ROX reference dye
(50X)3, 0.2 µl Taq DNA polymerase (5
U/µl), 2 µl miRNA RT product and sterilised
H2O, which added up to 20 µl. The thermocycling
conditions were as follows: 95°C for 3 min, 95°C for 12 sec and
62°C for 40 sec. Gene expression was normalised to U6 expression,
which was used as an internal control, and the relative expression
level was calculated using the 2−ΔΔCq method (18). The patients were divided into two
groups based on the median level of miR-198 expression: High
expression and low expression.
To determine the expression of E-cadherin,
N-cadherin, vimentin, MMP-2, MMP-9 and CDK4, cDNA served as the
template for PCR amplification using a cDNA synthesis kit (Takara
Biotechnology Co., Ltd.), according to the manufacturer's protocol.
The RT reaction system contained the following: 4 µl 5X
PrimeScript RT Master Mix, 1 µg RNA and RNase-free
H2O, which added up to 20 µl. The following
conditions were employed for RT: 37°C for 15 min and 85°C for 5
min. qPCR was conducted using SYBR Premix Ex Taq II kit (Takara
Biotechnology Co., Ltd.) with the following reaction system: 10
µl SYBR Premix Ex Taq II, 1 µl cDNA, 0.5 µl
forward primer, 0.5 µl reverse primer and 8 µl
sterile water. The thermocycling conditions were as follows: 95°C
for 1 min, 94°C for 30 sec, 58°C for 30 sec and 72°C for 10 sec.
Primers were designed and synthesised by Takara Biotechnology Co.,
Ltd. Gene expression was normalised to GAPDH expression, which was
used as an internal control, and the relative expression level was
calculated using the 2−ΔΔCq method.
Western blotting
Total protein from Cal-27 and SCC-9 cells were
extracted using RIPA lysis buffer (CoWin Biosciences) containing
PMSF (1 mM) and the concentration was determined using the
bicinchoninic acid method. Subsequently, 50 µg total protein
was separated via SDS-PAGE on a 10% gel, and subsequently separated
proteins were transferred onto PVDF membranes (Sigma-Aldrich; Merck
KGaA). Following blocking for 1 h with 5% non-fat milk at room
temperature, the membranes were incubated with primary antibodies
against E-cadherin (cat. no. 3195; 1:1,000; Cell Signaling
Technology, Inc.), N-cadherin (cat. no. 13116; 1:500; Cell
Signaling Technology, Inc.), vimentin (cat. no. 5741; 1:1,000; Cell
Signaling Technology, Inc.), MMP-2 (cat. no. 40994; 1:1,000; Cell
Signaling Technology, Inc.), MMP-9 (cat. no. 13667; 1:1,000; Cell
Signaling Technology, Inc.), CDK4 (cat. no. 12790; 1:1,000; Cell
Signaling Technology, Inc.) and GAPDH (cat. no. sc-47724; 1:5,000;
Santa Cruz Biotechnology, Inc.) overnight at 4°C. The membranes
were washed and then incubated with the secondary antibodies (cat.
nos. sc-2357 and sc-2005; 1:5,000; Santa Cruz Biotechnology, Inc.)
for 2 h at room temperature. The blots were visualised via ECL
(Pierce; Thermo Fisher Scientific, Inc.). ImageJ software (version
1.48; National Institutes of Health) was used for densitometry.
Bioinformatics analysis
The CDK4 expression data for OSCC was obtained from
the Gene Expression Profiling Interactive Analysis (GEPIA) online
database (http://gepia.cancer-pku.cn/). Tumour
and normal samples in the GEPIA database were derived from The
Cancer Genome Atlas (TCGA, https://tcga-data.nci.nih.gov/tcga/) and the
Genotype-Tissue Expression (GTEx, https://www.gtexportal.org/home/) projects.
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
(MTS) assay
OSCC Cal-27 and SCC-9 cells were transfected and
seeded on 96-well plates at a density of 2×103
cells/well. After culture for 24, 48, 72 or 96 h, cell
proliferation was tested via an MTS assay (Abcam), according to the
manufacturer's protocol. The absorbance of each well was measured
at 570 nm on a microplate reader.
Colony formation assay
OSCC Cal-27 and SCC-9 cells were transfected and
seeded on 6-well plates at a density of 1×103 cells/well
and cultured for 7 days. The medium was removed and cells were
washed with PBS twice, fixed in 100% methanol (1 ml) for 10 min and
stained with 10% crystal violet solution for 20 min at room
temperature, the colonies (defined as >10 cells) were counted
and photographed.
Flow cytometry analysis
OSCC Cal-27 and SCC-9 cells were transfected and
seeded on 6-well plates at a density of 4×105 cells/well
and cultured for 48 h. According to the manufacturer's protocols,
cells were incubated with Annexin V-Fluorescein
Isothiocyanate/Propidium Iodide Apoptosis Detection kit (Dojindo
Molecular Technologies, Inc.) in the dark. The results were
analysed using a BD FACSVerse™ flow cytometer (BD Biosciences)
equipped with the FlowJo version 10 software (FlowJo LLC). Early
and late apoptosis was assessed.
Transwell assay
OSCC Cal-27 and SCC-9 cells were transfected,
resuspended in serum-free DMEM, and plated onto the upper chamber
of the Transwell insert (Corning, Inc.) at a density of
5×104 cells/well. DMEM containing 10% FBS (500
µl) was added into the lower chamber to act as a
chemoattractant. For the invasion experiments, Transwell chambers
precoated with Matrigel (Corning, Inc.) at 4°C for 3 h. Cells in
the Transwell plates were cultured for 48 h at 37°C with 5%
CO2. The adherent cells on the upper surface of the
insert membrane were removed using cotton tips. Cells that migrated
into the lower chamber were stained with 10% crystal violet for 20
min at room temperature and quantitated by counting in five
different areas under a light microscope.
Enzyme-linked immunosorbent assay
(ELISA)
According to a previous study (19), the OSCC Cal-27 and SCC-9 cell
culture supernatant was collected after treatment. Using ELISA kits
(cat. nos. MMP200 and DMP900; R&D Systems, Inc.) the
concentrations of MMP-2 and MMP-9 were measured, according to the
manufacturer's protocol.
Dual-luciferase reporter assays
The potential binding sites of miR-198 and
3′-untranslated region (UTR) of CDK4 were predicted by TargetScan
7.1 (www.targetscan.org/vert_71). miR-198 mimics, miR NC
and CDK4-3′UTR mutant (CDK4-MUT) and CDK4-3′UTR wild-type (CDK4-WT)
plasmids were constructed by Shanghai GenePharma Co., Ltd. OSCC
Cal-27 and SCC-9 cells were seeded on 6-well plates at a density of
3×104 cells/well. After 24 h, miR-198 mimics (5′-GGU CCA
GAG GGG AGA UAG GUU C-3′) and miR NC (5′-UUC UCC GAA CGU GUC ACG
UAA U-3′) were co-transfected with CDK4-MUT or -WT plasmids into
OSCC cells using Lipofectamine 3000. After incubation for 48 h,
according to the manufacturer's protocol, a Dual-Luciferase
Reporter Assay System kit (Promega Corporation) was used. The
relative luciferase activity was determined by normalisation with
Renilla luciferase activity.
In vivo assays
A total of 24 male BALB/c nude mice (age, 4-6 weeks;
weight, 20-25 g) were purchased from Beijing Vital River Laboratory
Animal Technology Co., Ltd., (Charles River Laboratories). All
animals were raised at 22-26°C, 40-70% relative humidity and a 12 h
light/dark cycle. The animals were provided with water and food
freely. All animals were randomly divided into two groups
(experimental and control groups, 12 mice/group) and maintained in
a specific pathogen-free environment. For the xenograft experiment,
Cal-27 cells transfected with miR-198 mimics and miR-NC were
detached and resuspended in serum-free medium. Subsequently, Cal-27
cells (1 ml; 1×107 cells/ml) were implanted
subcutaneously in the flanks of BALB/c nude mice (20). The tumour size was checked
regularly at the indicated time points. The tumour-bearing mice
were sacrificed 6 weeks after the operation by cervical dislocation
and tumour weights were measured. The maximum tumour diameter was
13 mm, and volume was 1,100 mm3. All animal studies were
conducted at the Animal Center of China Medical University
(Shenyang, China) according to the protocols approved by the animal
ethics committee (Experimental animal welfare and ethics committee
of China Medical University, approval no. CMU2019182).
Statistical analysis
All statistical analysis was performed using SPSS
21.0 software (IBM Corp.). Data are represented as the mean ±
standard deviation based on at least three repeats. Comparisons
between tumour and adjacent normal tissue expression were performed
using a paired Student's t-test. For cell line experiments
involving two groups, an unpaired t-test was conducted. One-way
ANOVA followed by Tukey's post hoc test was performed to compare
the differences between multiple groups. Using the Kaplan-Meier
method and the log-rank test, survival curves were plotted and the
overall survival (OS) and disease-free survival (DFS) outcomes were
analysed. Pearson's correlation analysis was used to detect the
correlation between miR-198 and CDK4 expression in OSCC tissues.
Univariate and multivariate analyses were conducted according to
the Cox regression model. P<0.05 was considered to indicate a
statistically significant difference.
Results
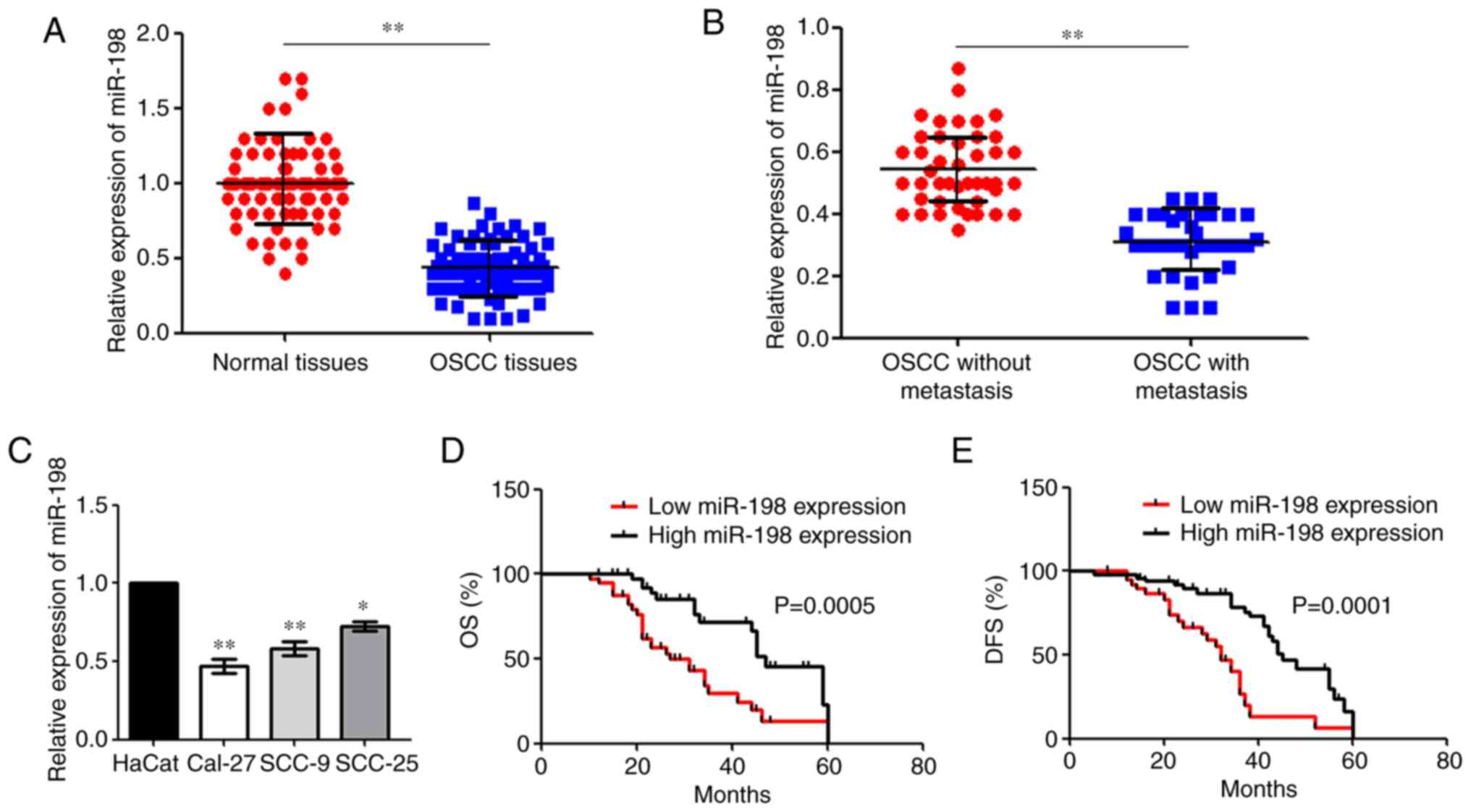
miR-198 is downregulated in OSCC
To investigate miR-198 expression in OSCC, miR-198
expression in OSCC tissues and the corresponding adjacent tissues
was primarily analysed using RT-qPCR. It was found that miR-198 was
downregulated in OSCC tissues compared with the normal tissues
(Fig. 1A). The level of miR-198
expression in patients with metastasis was significantly decreased
compared with patients with OSCC without metastasis (Fig. 1B). Moreover, RT-qPCR assays showed
that miR-198 expression was reduced in all OSCC cell lines compared
with HaCat cells (Fig. 1C).
Expression was the most significantly reduced in the Cal-27 and
SCC-9 cell lines, thus these cells were then chosen for further
assays. Patients with OSCC with low miR-198 expression had poorer
OS and DFS compared with patients with a high expression (Fig. 1D and E).
The cox proportional hazard model was used to
determine the impact of miR-198 expression and the
clinicopathological factors on the prognosis of patients with OSCC.
Univariate Cox regression analysis showed that the differentiation
(P=0.030 for OS; P=0.034 for DFS), TNM stage (P=0.039 for OS;
P=0.031 for DFS) and miR-198 expression (P=0.025 for OS; P=0.022
for DFS) were significantly associated with poor OS and DFS.
Multivariate analyses showed that the TNM stage (P=0.043 for OS;
P=0.033 for DFS) and miR-198 expression (P=0.033 for OS; P=0.027
for DFS) were associated with poor OS and DFS (Table I).
 | Table IUnivariate and multivariate Cox
proportional hazards model for OS and DFS in patients with oral
squamous cell carcinoma. |
Table I
Univariate and multivariate Cox
proportional hazards model for OS and DFS in patients with oral
squamous cell carcinoma.
A, OS
|
|---|
| Variables | Univariate
| Multivariate
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Sex (male vs.
female) | 1.441 | 0.412-2.113 | 0.549 | | | |
| Age, years (≤55 vs.
>55) | 1.559 | 0.712-2.176 | 0.432 | | | |
| Differentiation
(well/moderate vs. poor) | 3.554 | 0.558-3.569 | 0.030a | | | |
| Tumour size, cm (≤5
vs. >5) | 1.023 | 0.990-1.997 | 0.151 | | | |
| TNM stage (II+III
vs. I) | 2.814 | 0.998-3.698 | 0.039a | 2.612 | 1.122-3.006 | 0.043a |
| miR-198 expression
(high vs. low) | 3.996 | 1.345-5.885 | 0.025a | 3.221 | 1.443-4.113 | 0.033a |
| Tobacco usage (yes
vs. no) | 1.630 | 0.782-2.432 | 0.321 | | | |
|
B, DFS
|
| Variables | Univariate
| Multivariate
|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|
| Sex (male vs.
female) | 1.224 | 0.712-2.114 | 0.821 | | | |
| Age, years (≤55 vs.
>55) | 1.421 | 0.674-2.032 | 0.726 | | | |
| Differentiation
(well/moderate vs. poor) | 3.002 | 1.092-4.012 | 0.034a | | | |
| Tumour size, cm (≤5
vs. >5) | 1.123 | 0.598-2.912 | 0.915 | | | |
| TNM stage (II+III
vs. I) | 3.443 | 1.012-4.098 | 0.031a | 3.123 | 0.114-3.234 | 0.033a |
| miR-198 expression
(high vs. low) | 3.609 | 1.123-5.334 | 0.022a | 3.567 | 1.379-4.453 | 0.027a |
| Tobacco usage (yes
vs. no) | 1.768 | 0.543-4.119 | 0.564 | | | |
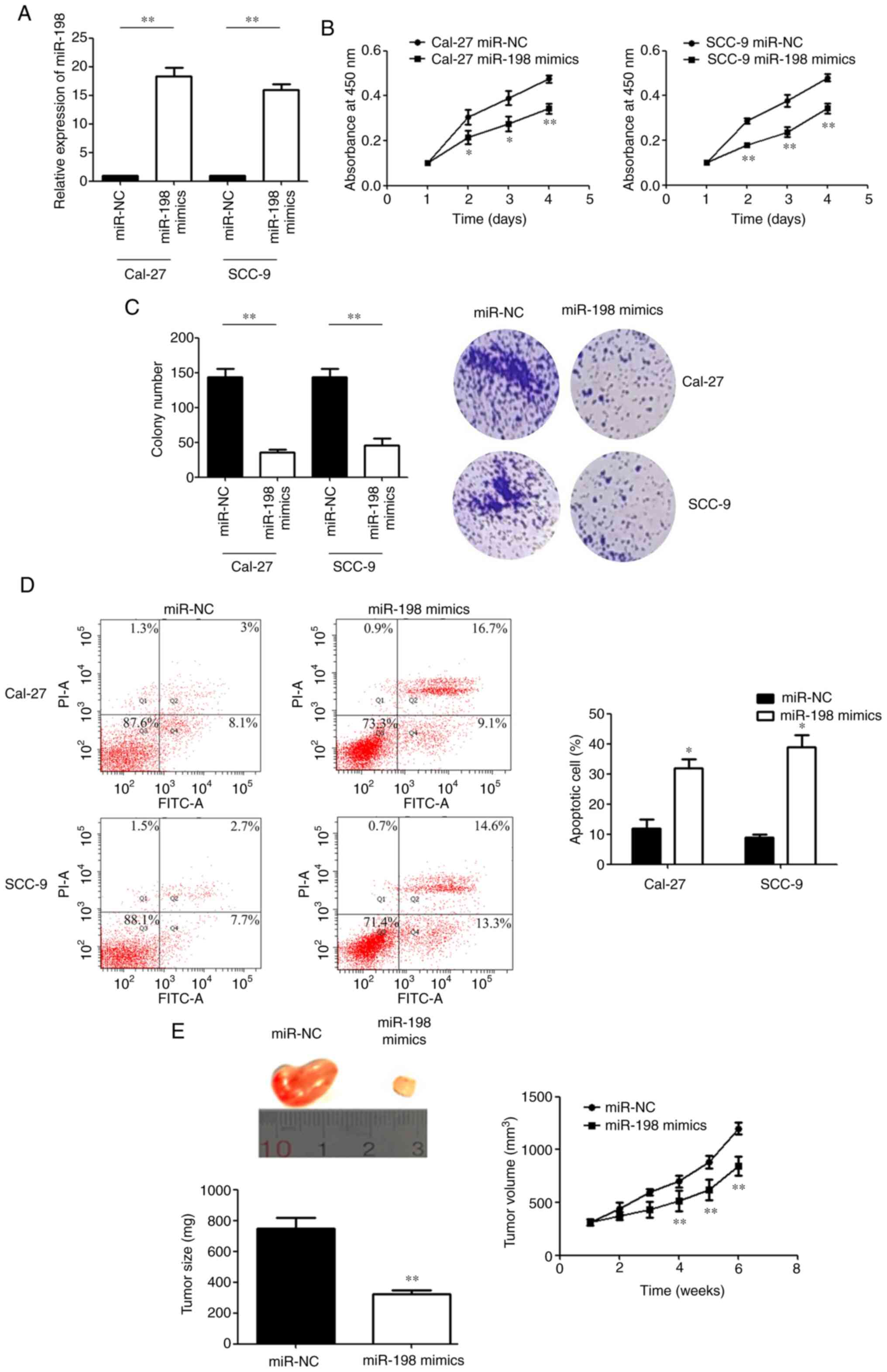
Upregulation of miR-198 suppresses the
proliferation and promotes the apoptosis of OSCC cells
RT-qPCR was used to detect transfection efficiency
after cells were transfected with miR-198 mimics (Fig. 2A). MTS assays showed that compared
with the miR-NC group, the proliferative activity of OSCC cells was
decreased after transfection with miR-198 mimics at different
observation time points (Fig.
2B). Consistent with this finding, the colony formation ability
of OSCC cells was significantly reduced after miR-198 mimic
transfection (Fig. 2C).
Subsequently, the flow cytometry results showed that the percentage
of apoptotic cells significantly increased after miR-198 mimics
transfection (Fig. 2D). Xenograft
experiments also showed a significant reduction in tumour size and
volume when miR-198 was overexpressed (Fig. 2E). Taken together, these data
indicated that miR-198 may function as a tumour suppressor in
OSCC.
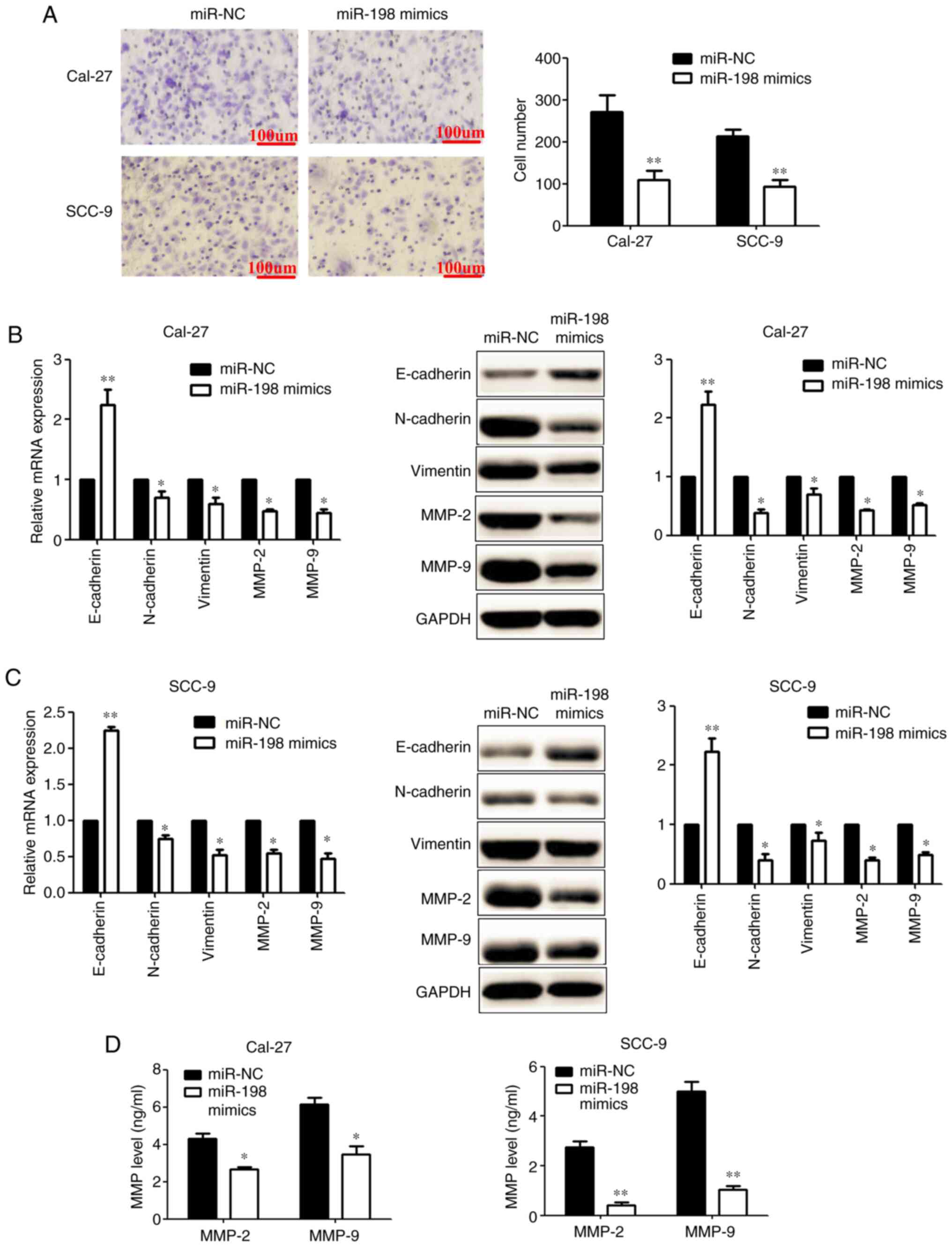
Upregulation of miR-198 suppresses
invasion and EMT in OSCC cells
Transwell assays showed that compared with the
miR-NC group, the number of invading OSCC cells was decreased
significantly after transfection with miR-198 mimics (Fig. 3A). RT-qPCR and western blotting
results showed that the overexpression of miR-198 significantly
increased the level of E-cadherin, and decreased the levels of
N-cadherin, vimentin, MMP-2 and MMP-9, in OSCC cells (Fig. 3B and C). ELISA showed that the
concentration of MMP-2 and MMP-9 was decreased significantly after
transfection with miR-198 mimics (Fig. 3D). Taken together, these data
indicated that miR-198 may function as a tumour suppressor in
OSCC.
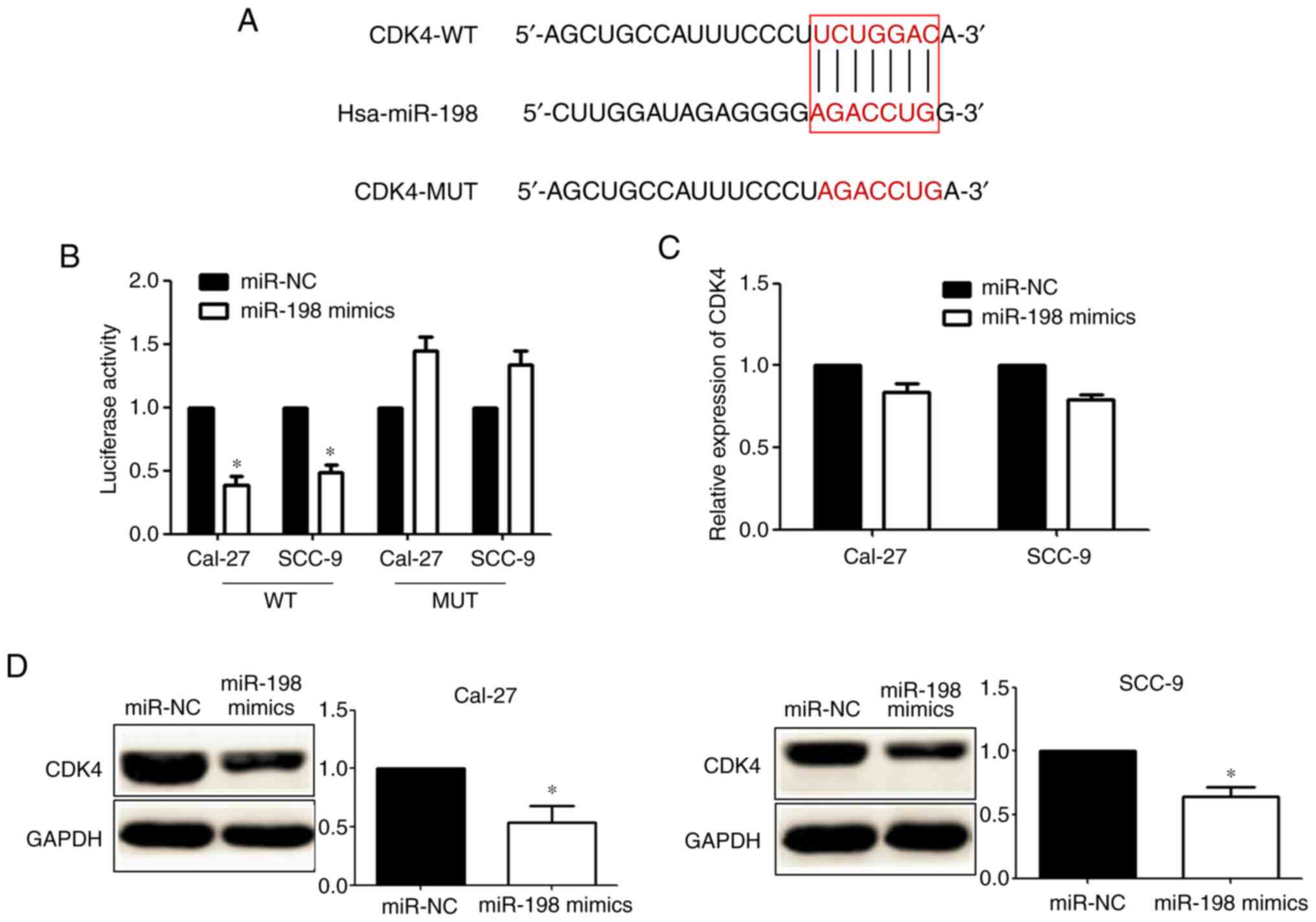
CDK4 is a direct target of miR-198 in
OSCC
TargetScan showed that there was a miR-198 binding
sequence in the 3′UTR of CDK4 (Fig.
4A). Dual-luciferase reporter assays showed that the
overexpression of miR-198 could inhibit CDK4-WT reporter activity,
but not the activity of the CDK4-MUT construct in OSCC cells,
demonstrating that miR-198 could specifically target the CDK4-3′UTR
by binding to the seed sequence (Fig.
4B). RT-qPCR showed that there were no obvious changes in CDK4
mRNA expression after transfection with miR-198 mimics (Fig. 4C). Western blotting showed that
transfection with miR-198 mimics significantly decreased the
expression of CDK4 protein (Fig.
4D). Taken together, these data indicated that miR-198 targets
CDK4 directly in OSCC cells.
CDK4 is upregulated in OSCC
From the GEPIA2 database, it was found that CDK4
expression was higher in OSCC tissues than that in controls
(Fig. 5A). RT-qPCR showed that
CDK4 mRNA expression was upregulated in OSCC tissues compared with
the normal tissues (Fig. 5B).
Moreover, RT-qPCR showed that there were higher expression levels
of CDK4 in OSCC cell lines compared with HaCat cells (Fig. 5C). Patients with OSCC with high
CDK4 expression had poorer OS and DFS than patients with a low
expression (Fig. 5D and E).
RT-qPCR analysis also showed that miR-198 was moderately negatively
correlated with CDK4 expression in patients with OSCC (Fig. 5F). Taken together, these data
indicated that CDK4 expression may be involved in OSCC
progression.
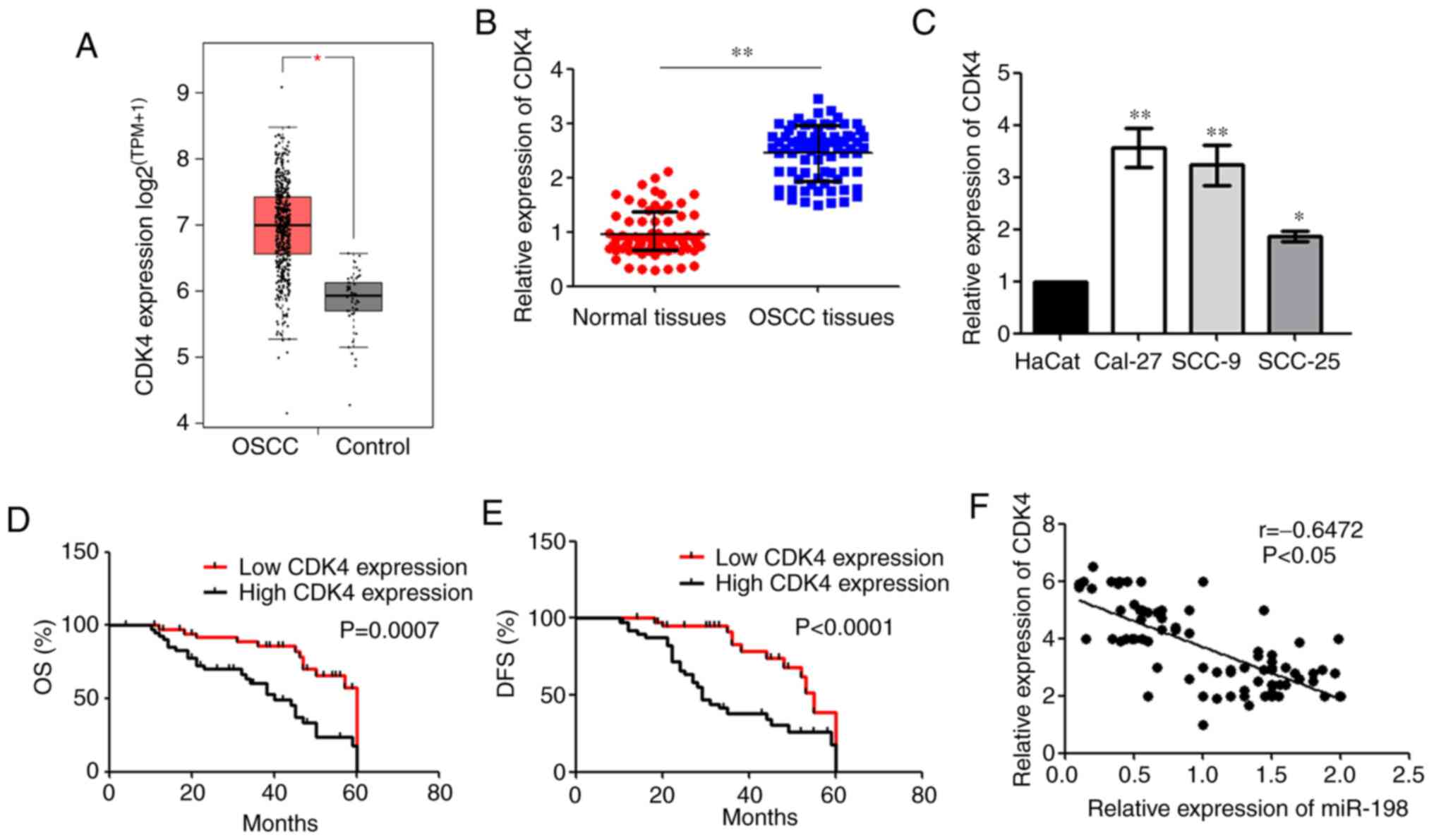 | Figure 5Expression of CDK4 in OSCC tissues
and cells. (A) The expression of CDK4 in OSCC and control (GEPIA2,
box plot, TPM). *P<0.05. (B) CDK4 expression in OSCC
and normal tissues, as determined via RT-qPCR.
**P<0.01. (C) CDK4 expression OSCC cell lines and the
HaCat cells were examined via RT-qPCR. *P<0.05,
**P<0.01 vs. HaCat cells. The Kaplan-Meier method was
used to compare the (D) OS and (E) DSF rates based on CDK4
expression. (F) A negative correlation was observed between the
expression levels of CDK4 and miR-198 in OSCC tissues. miR,
microRNA; OSCC, oral squamous cell carcinoma; CDK4,
cyclin-dependent kinase 4; RT-qPCR, reverse
transcription-quantitative PCR; OS, overall survival; DFS,
disease-free survival; TPM, transcripts per million. |
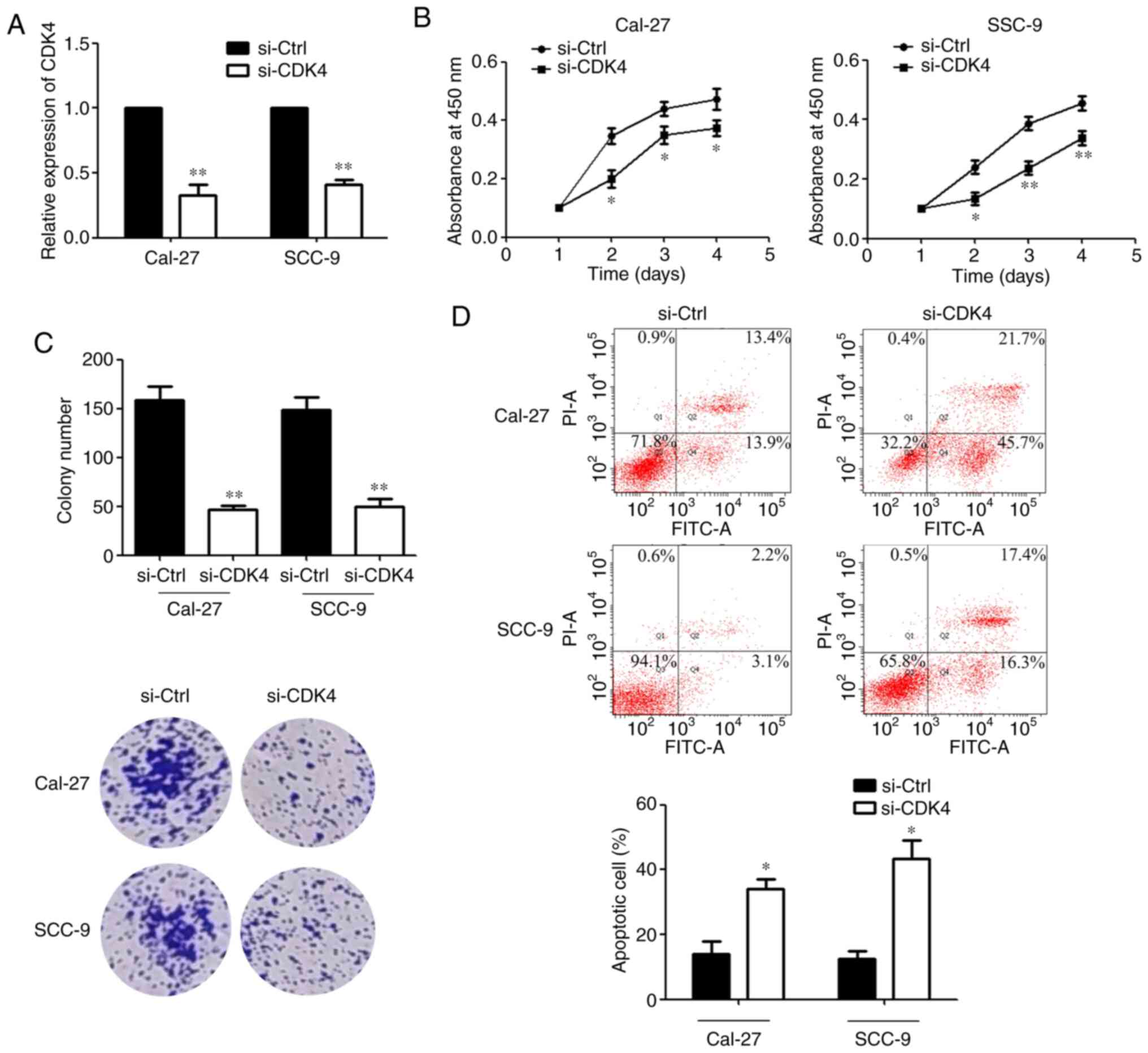
Downregulation of CDK4 suppresses the
proliferation and promotes the apoptosis of OSCC cells
RT-qPCR was performed to detect the transfection
efficiency after si-CDK4 was transfected (Fig. 6A). MTS assays showed that compared
with the si-Ctrl group, the proliferative activity of OSCC cells
was decreased after transfection of si-CDK4 at different
observation time points (Fig.
6B). Consistent with this finding, the colony formation ability
of OSCC cells was significantly reduced after transfection with
si-CDK4 (Fig. 6C). Subsequently,
the results of flow cytometry showed that the percentage of
apoptotic cells was significantly increased after transfection with
si-CDK4 (Fig. 6D). Taken
together, these data indicated that CDK4 may function as a tumour
promoter in OSCC.
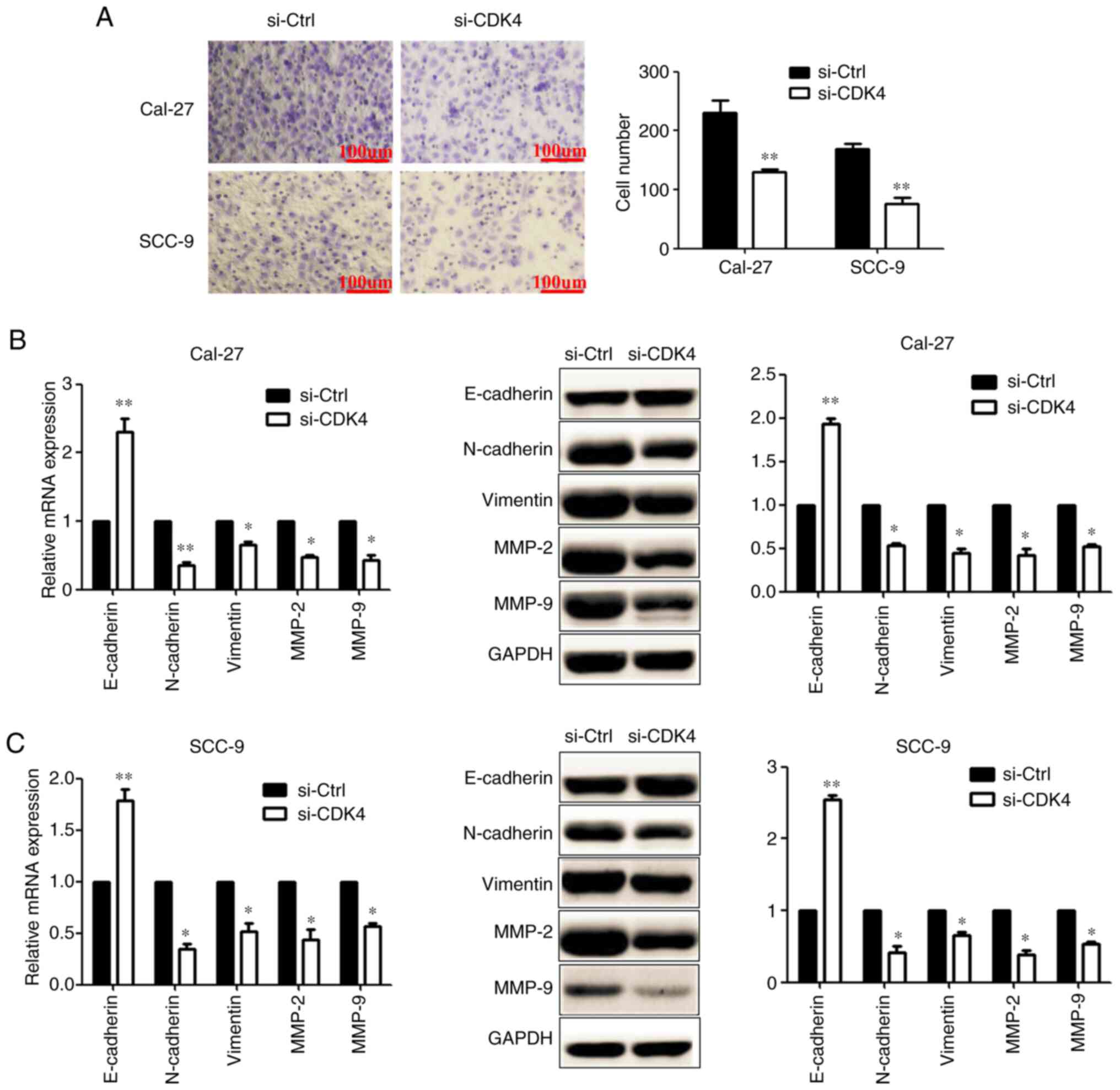
Downregulation of CDK4 suppresses
invasion and EMT in OSCC cells
Transwell assays showed that compared with the
si-Ctrl group, the number of invading OSCC cells was significantly
decreased after transfection with si-CDK4 (Fig. 7A). RT-qPCR and western blotting
results showed that knockdown of CDK4 expression significantly
increased the level of E-cadherin, and decreased the levels of
N-cadherin, vimentin, MMP-2 and MMP-9, in OSCC cells (Fig. 7B and C). Collectively, these data
indicated that CDK4 may function as a tumour promoter in OSCC.
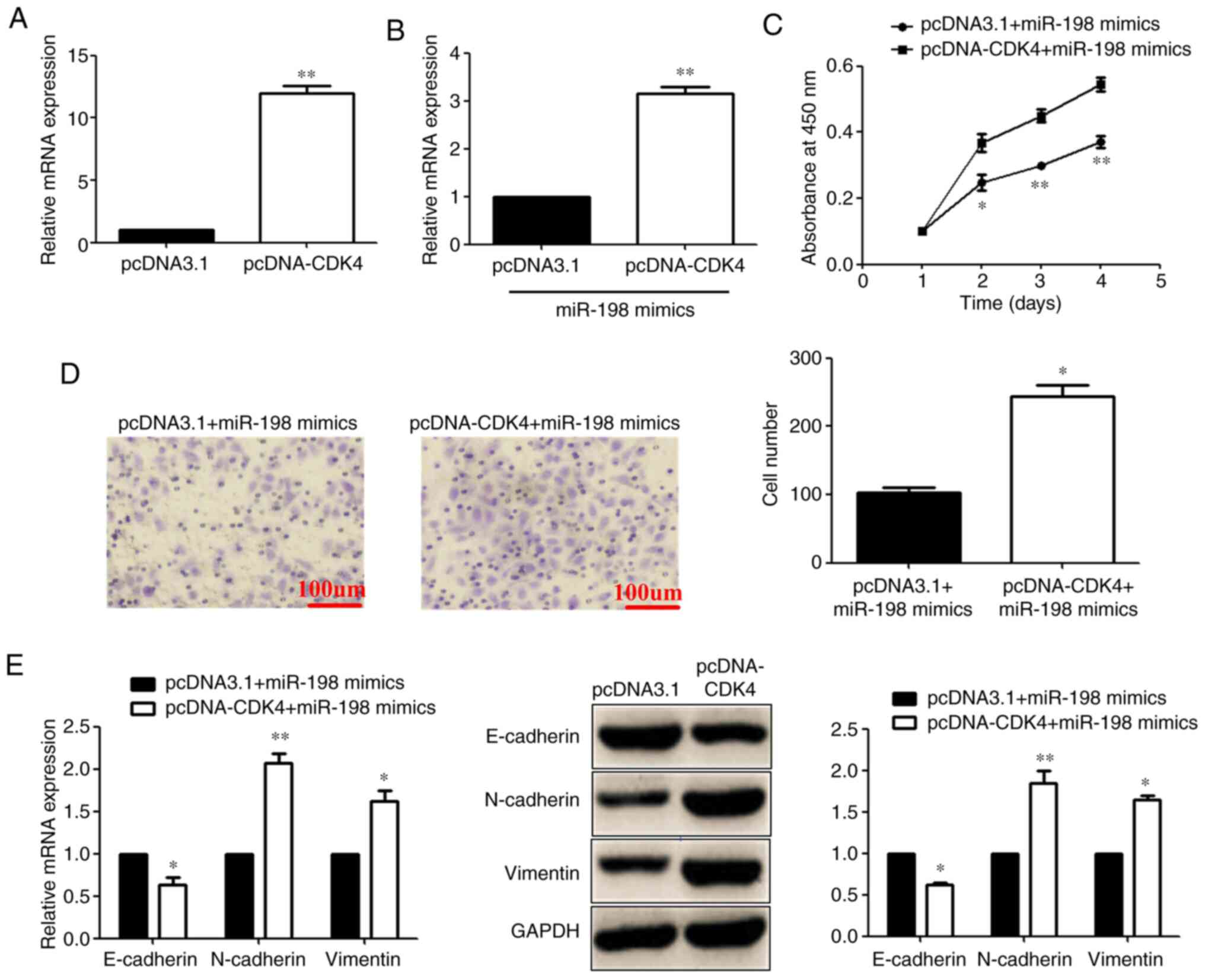
Addition of CDK4 reverses the
miR-198-mediated inhibitory effect on proliferation, invasion and
EMT in OSCC cells
A representation vector that encoded the entire CDK4
coding sequence and lacked the 3′UTR was constructed. RT-qPCR was
used to detect the transfection efficiency after pcDNA-CDK4 was
transfected (Fig. 8A). Then,
pcDNA-CDK4 or its NC was co-transfected with the miR-198 mimic into
Cal-27 cells. RT-qPCR showed that CDK4 expression was significantly
increased after co-transfection of pcDNA-CDK4 + miR-198 mimics
compared with the pcDNA3.1 + miR-198 mimic group (Fig. 8B). MTS assays showed that the
concomitant overexpression of miR-198 and CDK4 abrogated the
inhibitory effects of transfection with pcDNA3.1 + miR-198 mimic
(Fig. 8C). Transwell assays also
showed that CDK4 overexpression reversed the inhibitory effects of
transfection with pcDNA3.1 + miR-198 mimic on the invasion of
Cal-27 cells (Fig. 8D). Moreover,
RT-qPCR and western blotting showed that the overexpression of CDK4
decreased the level of E-cadherin, and increased N-cadherin and
vimentin, in Cal-27 cells compared with the pcDNA3.1 + miR-198
mimic group (Fig. 8E). Taken
together, these findings revealed that CDK4 reversed the
miR-198-mediated inhibitory effect in OSCC cells.
Discussion
Increasing evidence has demonstrated that abnormal
miRNA expression is detected in OSCC, and some of these miRNAs are
involved in modulating OSCC cell proliferation and metastasis
(21,22). Liang et al (23) found that miR-1297 expression is
significantly decreased in OSCC tumours compared with that in
adjacent non-tumour tissues, and the overexpression of miR-1297
could suppress the cell proliferation of SSC-4 cells in
vitro (23). In addition,
Nagai et al (24)
demonstrated that miR-205-5p could suppress the invasiveness of
OSCC by inhibiting metalloproteinase inhibitor 2 expression. The
present study confirmed a low expression of miR-198 in clinical
tumour tissues, which was consistent with the results of other
studies on several different types of tumours (12-14). Furthermore, the expression of
miR-198 in patients with metastasis was significantly lower than
that in patients without metastasis. OSCC metastasis can be
classified as lymphatic metastasis or vascular invasion, which
significantly impacts patient prognosis (25). Previous studies have reported that
miR-198 is a key molecule that is responsible for vascular invasion
(26-28). To investigate the effects of
miR-198 on OSCC prognosis, its prognostic ability in patients with
OSCC was examined. It was found that a low miR-198 expression was
significantly related to a poor OS and DFS in patients with
OSCC.
The previously identified targets of miR-198 vary
widely in structure and function depending on the cancer type
investigated. Yang et al (13) confirmed that miR-198 can inhibit
the proliferation of lung cancer cells and induce apoptosis by
targeting the fibroblast growth factor receptor in lung cancer.
Wang et al (20)
demonstrated that miR-198 targets fucosyltransferase 8 and can
inhibit tumour growth and metastasis in colorectal cancer.
Moreover, Ye et al (29)
found that miR-198 could inhibit livin expression, indicating that
miR-198 was associated with cell apoptosis by targeting livin in
prostate cancer cells. Through the investigation of non-cancer
cells, it has also been found that miR-198 overexpression can
inhibit cell proliferation in keratinocyte cell lines, and
significantly arrest cells at the G1 phase, as detected by cell
cycle analysis, by targeting cyclinD2 (30). In prostate cancer cell lines, by
targeting E3 ubiquitin-protein ligase MIB1, overexpression of
miR-198 has also been found to increase G0/G1 cell cycle arrest and
inhibit proliferation (31). In
the present study, using a dual-luciferase reporter assay, it was
confirmed that miR-198 suppressed CDK4 levels by binding to the
3′UTR of the oncogene. To the best of the authors' knowledge, this
study is the first to present the effects of miR-198 in OSCC cell
proliferation, apoptosis and invasion.
CDK genes belong to the serine/threonine kinase
family and are involved in the cell cycle pathway. CDK genes form a
complex with G1/S-specific cyclin-D1 and phosphorylates
retinoblastoma-associated protein gene (32). As a member of the CDK family, CDK4
is an essential signalling transduction molecule, which
participates in cell cycle and apoptosis by binding to cyclin
proteins (33). Its concentration
and activation are closely related to the transformation rate of
the G1/S phase of the cell cycle (34). The deregulation of CDK4 expression
is associated with several aspects of tumorigenesis, including cell
proliferation, cycle arrest and abnormal apoptosis (35). There is growing evidence that more
cells undergo apoptosis when CDK4 is knocked down in non-small cell
lung, oesophageal and breast cancer (36-38). In the present study, it was found
that CDK4 was highly expressed in most OSCC tissues and that the
proliferative ability of OSCC cells was inhibited after the
knockdown of CDK4, Moreover, metastasis and EMT were reduced.
Furthermore, the overexpression of CDK4 counteracted the
miR-198-mediated effects on cell behaviours, which also indicated
that the presence of CDK4 worsened the outcome in OSCC cells.
In conclusion, the results of the present study
showed that the overexpression of miR-198 could suppress tumour
growth and metastasis in OSCC cells by targeting CDK4. Therefore,
the miR-198/CDK4 axis may be considered as a novel prognostic
marker and therapeutic target in OSCC.
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YK designed the experiments. YZ and YS analysed the
data. YK and YS performed the experiments. YK wrote the paper. YK
and YZ confirm the authenticity of all the raw data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All research protocols were approved by the ethics
committee of the School and Hospital of Stomatology, China Medical
University (Shenyang, China) and written informed consent was
obtained from the donors.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of China (grant no. 82002886).
References
|
1
|
Zhang S, Wang X, Gupta A, Fang X, Wang L
and Zhang C: Expression of IL-17 with tumor budding as a prognostic
marker in oral squamous cell carcinoma. Am J Transl Res.
11:1876–1883. 2019.PubMed/NCBI
|
|
2
|
Wu K, Jiang Y, Zhou W, Zhang B, Li Y, Xie
F, Zhang J, Wang X, Yan M, Xu Q, et al: Long noncoding RNA RC3H2
facilitates cell proliferation and invasion by targeting
MicroRNA-101-3p/EZH2 Axis in OSCC. Mol Ther Nucleic Acids.
20:97–110. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen L, Zhang S, Wu J, Cui J, Zhong L,
Zeng L and Ge S: circRNA_100290 plays a role in oral cancer by
functioning as a sponge of the miR-29 family. Oncogene.
36:4551–4561. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao Z, Gao D, Ma T and Zhang L:
MicroRNA-141 suppresses growth and metastatic potential of head and
neck squamous cell carcinoma. Aging (Albany NY). 11:921–932. 2019.
View Article : Google Scholar
|
|
5
|
Wang Y, Guo W, Li Z, Wu Y, Jing C, Ren Y,
Zhao M, Kong L, Zhang C, Dong J, et al: Role of the EZH2/miR-200
axis in STAT3-mediated OSCC invasion. Int J Oncol. 52:1149–1164.
2018.PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Flynt AS and Lai EC: Biological principles
of microRNA-mediated regulation: Shared themes amid diversity. Nat
Rev Genet. 9:831–842. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang B, Li Y, Hou D, Shi Q, Yang S and Li
Q: MicroRNA-375 inhibits growth and enhances radiosensitivity in
oral squamous cell carcinoma by targeting insulin like growth
factor 1 receptor. Cell Physiol Biochem. 42:2105–2117. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang K, Jin J, Ma T and Zhai H: MiR-139-5p
inhibits the tumorigenesis and progression of oral squamous
carcinoma cells by targeting HOXA9. J Cell Mol Med. 21:3730–3740.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fu S, Chen HH, Cheng P, Zhang CB and Wu Y:
MiR-155 regulates oral squamous cell carcinoma Tca8113 cell
proliferation, cycle, and apoptosis via regulating p27Kip1. Eur Rev
Med Pharmacol Sci. 21:937–944. 2017.PubMed/NCBI
|
|
11
|
Yu EH, Tu HF, Wu CH, Yang CC and Chang KW:
MicroRNA-21 promotes perineural invasion and impacts survival in
patients with oral carcinoma. J Chin Med Assoc. 80:383–388. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu Y, Tang Z, Jiang B, Chen J and Fu Z:
MiR-198 functions as a tumor suppressor in breast cancer by
targeting CUB domain-containing protein 1. Oncol Lett.
13:1753–1760. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang J, Zhao H, Xin Y and Fan L:
MicroRNA-198 inhibits proliferation and induces apoptosis of lung
cancer cells via targeting FGFR1. J Cell Biochem. 115:987–995.
2014. View Article : Google Scholar
|
|
14
|
Tan S, Li R, Ding K, Lobie PE and Zhu T:
MiR-198 inhibits migration and invasion of hepatocellular carcinoma
cells by targeting the HGF/c-MET pathway. FEBS Lett. 585:2229–2234.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cui Z, Zheng X and Kong D: Decreased
miR-198 expression and its prognostic significance in human gastric
cancer. World J Surg Oncol. 14:332016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamamoto K, Kawaguchi M, Shimomura T,
Izumi A, Konari K, Honda A, Lin CY, Johnson MD, Yamashita Y,
Fukushima T and Kataoka H: Hepatocyte growth factor activator
inhibitor type-2 (HAI-2)/SPINT2 contributes to invasive growth of
oral squamous cell carcinoma cells. Oncotarget. 9:11691–11706.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu Q, Wu T, Chen X, Li H, Du Z, Hao Y,
Peng J, Tai S, Song M and Cheng B: The poor outcome of second
primary oral squamous cell carcinoma is attributed to Bmi1
upregulation. Cancer Med. 7:1056–1069. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Zhong F, Chen H, Han L, Jin Y and Wang W:
Curcumin attenuates lipopolysaccharide-induced renal inflammation.
Biol Pharm Bull. 34:226–232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang M, Wang J, Kong X, Chen H, Wang Y,
Qin M, Lin Y, Chen H, Xu J, Hong J, et al: MiR-198 represses tumor
growth and metastasis in colorectal cancer by targeting fucosyl
transferase 8. Sci Rep. 4:61452014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng CM, Shiah SG, Huang CC, Hsiao JR and
Chang JY: Up-regulation of miR-455-5p by the TGF-β-SMAD signalling
axis promotes the proliferation of oral squamous cancer cells by
targeting UBE2B. J Pathol. 240:38–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chang KW, Kao SY, Wu YH, Tsai MM, Tu HF,
Liu CJ, Lui MT and Lin SC: Passenger strand miRNA miR-31* regulates
the phenotypes of oral cancer cells by targeting RhoA. Oral Oncol.
49:27–33. 2013. View Article : Google Scholar
|
|
23
|
Liang L, Feng L and Wei B: MicroRNA-1297
involves in the progression of oral squamous cell carcinoma through
PTEN. Saudi J Biol Sci. 25:923–927. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nagai H, Hasegawa S, Uchida F, Terabe T,
Ishibashi Kanno N, Kato K, Yamagata K, Sakai S, Kawashiri S, Sato
H, et al: MicroRNA-205-5p suppresses the invasiveness of oral
squamous cell carcinoma by inhibiting TIMP-2 expression. Int J
Oncol. 52:841–850. 2018.PubMed/NCBI
|
|
25
|
Adel M, Kao HK, Hsu CL, Huang JJ, Lee LY,
Huang Y, Browne T, Tsang NM, Chang YL and Chang KP: Evaluation of
lymphatic and vascular invasion in relation to clinicopathological
factors and treatment outcome in oral cavity squamous cell
carcinoma. Medicine (Baltimore). 94:e15102015. View Article : Google Scholar
|
|
26
|
Mattiotti A, Prakash S, Barnett P and van
den Hoff MJB: Follistatin-like 1 in development and human diseases.
Cell Mol Life Sci. 75:2339–2354. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liao C, Huang X, Gong Y and Lin Q:
Discovery of core genes in colorectal cancer by weighted gene
co-expression network analysis. Oncol Lett. 18:3137–3149.
2019.PubMed/NCBI
|
|
28
|
Shi Y, Fang N, Li Y, Guo Z, Jiang W, He Y,
Ma Z and Chen Y: Circular RNA LPAR3 sponges microRNA-198 to
facilitate esophageal cancer migration, invasion, and metastasis.
Cancer Sci. 111:2824–2836. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ye L, Li S, Ye D, Yang D, Yue F, Guo Y,
Chen X, Chen F, Zhang J and Song X: Livin expression may be
regulated by miR-198 in human prostate cancer cell lines. Eur J
Cancer. 49:734–740. 2013. View Article : Google Scholar
|
|
30
|
Wang J, Dan G, Shangguan T, Hao H, Tang R,
Peng K, Zhao J, Sun H and Zou Z: MiR-198 represses the
proliferation of HaCaT cells by targeting cyclin D2. Int J Mol Sci.
16:17018–17028. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ray J, Hoey C, Huang X, Jeon J, Taeb S,
Downes MR, Boutros PC and Liu SK: MicroRNA-198 suppresses prostate
tumorigenesis by targeting MIB1. Oncol Rep. 42:1047–1056.
2019.PubMed/NCBI
|
|
32
|
Wood DJ and Endicott JA: Structural
insights into the functional diversity of the CDK-cyclin family.
Open Biol. 8:1801122018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hamilton E and Infante JR: Targeting
CDK4/6 in patients with cancer. Cancer Treat Rev. 45:129–138. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gao X, Leone GW and Wang H: Cyclin
D-CDK4/6 functions in cancer. Adv Cancer Res. 148:147–169. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sheppard KE and AbuHammad S: CDK4/6
inhibition in cancer: The cell cycle splicing connection. Mol Cell
Oncol. 6:e16736432019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Feng H, Ge F, Du L, Zhang Z and Liu D:
MiR-34b-3p represses cell proliferation, cell cycle progression and
cell apoptosis in non-small-cell lung cancer (NSCLC) by targeting
CDK4. J Cell Mol Med. 23:5282–5291. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lang B and Zhao S: MiR-486 functions as a
tumor suppressor in esophageal cancer by targeting CDK4/BCAS2.
Oncol Rep. 39:71–80. 2018.
|
|
38
|
Tarasewicz E, Hamdan R, Straehla J, Hardy
A, Nunez O, Zelivianski S, Dokic D and Jeruss JS: CDK4 inhibition
and doxorubicin mediate breast cancer cell apoptosis through Smad3
and survivin. Cancer Biol Ther. 15:1301–1311. 2014. View Article : Google Scholar : PubMed/NCBI
|