Introduction
Ovarian cancer is the most lethal gynecological
cancer type and the fifth leading cause of cancer-related mortality
of women in the USA in 2016 (1).
In 2019, there were ~22,530 new cases of ovarian cancer, resulting
in 13,980 deaths (2). Since most
patients with ovarian cancer are diagnosed at an advanced stage,
chemotherapy is usually required before or after surgery (3). The combination of platinum and
taxane is regarded as the first-line approach (3). Despite intensive research during the
past 20 years, the 5-year survival rates have not sufficiently
improved, due to both intrinsic and acquired chemoresistance
(3). In addition, current
chemotherapeutic approaches may have several side effects that may
limit their application (4).
Identification of novel, more effective and less toxic therapeutic
targets is necessary.
Both IL-6 and IL-8 are inflammatory chemokines that
have been demonstrated to serve an important role in the
tumorigenesis of a variety of malignancies, including ovarian
cancer (5). IL-6 and IL-8 are
involved in tumor cell apoptosis and invasion, tumor growth and
metastasis (6-9). IL-6 and IL-8 upregulation has been
associated with chemoresistance in ovarian cancer (10). Certain clinical studies have
demonstrated that the IL-6 and IL-8 concentration in the peritoneal
fluid of patients with ovarian cancer is 100-1,000-fold higher than
that in the serum, and is associated with advanced stage, high
grade, lymph node metastasis and poor prognosis in ovarian cancer
(6,7,11).
Our previous study demonstrated that the
simultaneous inhibition of IL-6 and IL-8 can reduce the viability,
colony formation and migration of triple-negative breast and
pancreatic cancer cells; however, the potential mechanism was not
elucidated (5). Therefore, the
aim of the present study was not only to define the anticancer
activity of IL-6/IL-8 co-inhibition in ovarian cancer but also to
identify the mechanisms underlying IL-6 and IL-8 signaling. To
accomplish this, two agents with demonstrated activity in
pancreatic, triple-negative breast and colon cancer were used in
combination to treat ovarian cancer in vitro and in
vivo; Baze, a third-generation selective estrogen receptor
modulator approved by the Food and Drug Administration and novel
inhibitor of IL-6/glycoprotein 130 (GP130) protein-protein
interactions (12–15), and SCH, an IL-8/chemokine (CXC
motif) ligand 1/CXC chemokine receptor 2 (CXCR2) antagonist
(16).
Materials and methods
Materials
SCH was purchased from AdooQ Bioscience. Baze
acetate was purchased from Merck KGaA. The stock concentration of
SCH and Baze acetate was 20 mM reconstituted in DMSO (Merck KGaA)
and stored at −20°C. MTT was purchased from Sigma-Aldrich; Merck
KGaA, and a 5-mM stock concentration was prepared in
ddH2O and stored at −20°C. All primary and secondary
antibodies were purchased from Cell Signaling Technology, Inc.
Cell lines
SKOV3, CAOV3, OVCAR3 and A2780 human ovarian cancer
cell lines were purchased from American Type Culture Collection.
OV75 primary ovarian cancer cells were provided by Dr Jocelyn
Reader and Dr Dana Roque, who were the original suppliers (Division
of Gynecologic Oncology, University of Maryland School of Medicine,
Baltimore, MD, USA). OV75 cells are primary ovarian cancer cells
available for research labs from the University of Maryland. The
SKOV3, CAOV3 and A2780 cells were cultured in DMEM (Corning, Inc.)
supplemented with 10% FBS (Sigma-Aldrich; Merck KGaA) and 1%
penicillin/streptomycin (PS). OVCAR3 cells were cultured in RPMI
1640 medium (Corning, Inc.) with 10% FBS and 1% PS. OV75 cells were
cultured in HOSE Media [1:1 mixture of MCDB 105 (Sigma-Aldrich;
Merck KGaA) and Medium 199 (Thermo Fisher Scientific, Inc.)], 10%
FBS, 1% L-glutamine, 1% non-essential amino acids, 1% PS, sodium
bicarbonate at pH 7.4) as described previously (17). All cell lines were grown in a
humidified 37°C incubator with 5% CO2/95% air, and the
medium was replaced twice a week.
Measurement of IL-6 and -8
Immunoreactive IL-6 and IL-8 were measured using an
Quantikine ELISA kit (Human CXCL8/IL-8, cat. no. D8000C; human
IL-6, cat. no. D6050; R&D Systems, Inc.). Ovarian cancer cells
were seeded in 6-well plates at a confluency of 70%. The cell-free
culture supernatant of every well was collected and incubated, and
optical density (OD) was read at 450 nm. Each assay detects
cytokines as low as 5±7 pg/ml.
Western blot analysis
Cells were seeded in 10-cm plates at 70% confluency
and treated with DMSO, Baze, SCH or Baze + SCH (CAOV3, Baze 10
µM, SCH 50 µM; SKOV3, Baze 5 µM, SCH 50
µM) at 37°C overnight before being harvested, lysed in cold
Cell Lysis Buffer (0.5% 0.2 M PMSF, 0.5% 0.2 M NaF, 0.5% 0.2 M
NaPP, 0.5% 0.1 M Na3VO4 and 4% 25X CP1; Cell
Signaling Technology, Inc.) to collect the protein for western blot
analysis. The protein concentration was determined using a
Microplate BCA Protein Assay kit (cat. no. 23252; Thermo Fisher
Scientific, Inc.). The proteins (30 µg/lane) were separated
by 10% SDS-PAGE (30 µg per lane), transferred to a PVDF
membrane at 350 mA for 110 min, blocked with 5% milk/TBS-8%
Tween-20 (TBST) for 1 h at room temperature and incubated with
primary antibodies overnight at 4°C. The membranes were washed with
TBST three times (15 min each), blotted with the secondary antibody
for 1.5 h at room temperature. The visualization reagent (Western
Lighting Plus-ECL; PerkinElmer, Inc.) was added and membranes were
scanned using the Amersham Imager 600 (GE Healthcare, version
2.0.0).
The following primary antibodies were purchased from
Cell Signaling Technology, Inc. and diluted at 1:1,000 in 5% milk:
Phosphorylated (p)-STAT3 (Y705; rabbit mAb; cat. no. 9131S), STAT3
(rabbit mAb; cat. no. 4904S), p-S6 (rabbit mAb; cat. no. 4858S), S6
(rabbit mAb; cat. no. 2217S), p-AKT (rabbit mAb; cat. no. 4060S),
AKT (rabbit mAb; cat. no. 4691S), survivin (rabbit mAb; cat. no.
2808S) and GAPDH (rabbit mAb; cat. no. 2118S). The secondary
antibody was diluted at 1:10,000 in 5% milk (anti-rabbit IgG
HRP-linked antibody; cat. no. 7074; Cell Signaling Technology,
Inc.).
Cell proliferation assay
Ovarian cancer cells were seeded in 24-well plates
at the same cell density, depending on the growth ability of each
cell line (SKOV3, 2×103 cells per well; CAOV3,
3×103 cells per well; OVCAR3, 3×103 cells per
well), cultured overnight at 37°C and treated with DMSO, Baze, SCH
or Baze + SCH at 37°C until the end of the assay (CAOV3, Baze 5
µM, SCH 15 µM; SKOV3, Baze 2.5 µM, SCH 15
µM; OVCAR3, Baze 2.5 µM, SCH 15 µM). The cell
number of each well was then counted every 2 days after treatment
(on days 2, 4, 6 and 8) to generate the growth curves. The cell
density of cell growth assay is much lower than that of other
assays, so the doses of Baze were less than those of other assays
(CAOV3, Baze 5 µM; SKOV3, Baze 2.5 µM; OVCAR3, Baze
2.5 µM). However, the effects of different concentrations of
SCH (such as 10 or 15 µM) were similar. Therefore, the SCH
concentration remained constant in a number of experiments,
including the MTT and wound healing/cell migration assays (the
concentration of SCH here is 15 µM for each cell line).
MTT assay
A total of 3,000 cells in 100 µl medium per
well were seeded into 96-well microtiter plates and treated
overnight with DMSO, Baze, SCH or Baze + SCH (CAOV3, Baze 5
µM, SCH 15 µM; SKOV3, Baze 5 µM, SCH 15
µM; OVCAR3, Baze 5 µM, SCH 15 µM; OV75, Baze
10 µM, SCH 15 µM), followed by incubation at 37°C for
72 h. Each well was treated with 20 µl MTT, incubated for ~4
h and then combined with 150 µl N,N-dimethylformamide
solution, followed by further incubation overnight at room
temperature protected from the light. Cell viability was assessed
by measuring absorbance at 595 nm for each well. The cell viability
of DMSO control cells was set at 100% and the cell viability of
drug-treated cells was compared with that of DMSO-treated cells.
The combination index (CI) was then determined using CompuSyn
software (www.combosyn.com; ComboSyn, Inc.). CI
values of 1 indicate an additive effect, values of >1 an
antagonistic effect and values of <1 a synergistic effect, based
on the theorem of Chou (18).
Wound healing/cell migration assay
SKOV3 cells were seeded in 6-well plates and
incubated at 37°C overnight until they reached 100% confluence. A
100-µl pipette tip was used to scrape the monolayer of each
well, each well was washed twice with PBS, serum-free culture
medium containing different drugs (DMSO, Baze, SCH and their
combination) was added (SKOV3, Baze 5 µM, SCH 15 µM)
and images were captured under the light microscope.
Cells were incubated at 37°C and imaged again when
the wound in the DMSO control was well healed (17 h). Migration
inhibition was measured using ImageJ software 1.53e (National
Institutes of Health) and calculated using the following formula:
Percentage of wound healing = 100-[(final area/initial area) ×
100%] (19).
In vitro invasion assay using
Matrigel®
This assay was performed as described previously
(20). Matrigel was diluted at
1:8 with DMEM (without FBS and PS), added on the top of the
Transwell membrane and allowed to polymerize for a minimum of 30
min at 37°C. Next, 5×104 cells were added in 200
µl culture medium without FBS on the top of the Matrigel and
600 µl normal culture medium (DMEM or 1640 with 10% FBS) was
added to the bottom of the lower chamber in a 24-well plate. The
cells were treated with different drugs: DMSO, Baze, SCH or Baze +
SCH for 24 h at 37°C (CAOV3, Baze 5 µM, SCH 15 µM;
OVCAR3, Baze 10 µM, SCH 15 µM). The
Transwell® insert (permeable cell culture inserts;
CELLTREAT Scientific Products) was then removed from the plate and
the media and the remaining cells were carefully removed from the
top of the membrane without damaging it using a cotton-tipped
applicator. The Transwell insert was placed into 70% ethanol for 10
min and then stained with 0.2% crystal violet for 5-10 min at room
temperature, rinsed, dried and imaged under a light microscope.
Finally, the Transwell membrane was decolorized with 3% acetic acid
to completely elute the crystal violet, and 100 µl/well
eluent was added into a 96-well plate to measure the OD value (570
nm) on a microplate reader.
Ovarian peritoneal tumor growth
A peritoneal ovarian tumor mouse model was used to
evaluate the efficacy of Baze + SCH compared with monotherapy in
suppressing ovarian tumor growth. CAOV3-luciferase ovarian cancer
cells (1×107) in DMEM without FBS were injected
intraperitoneally into a total of 20 female athymic nude mice aged
6–8 weeks purchased from Jackson Laboratory (21,22). Mice were housed at ~37°C with a
12/12-h light/dark cycle and ad libitum access to food and
water. The development and growth of the tumors were monitored 1–2
times a week using an IVIS™ instrument (bioluminescent imaging;
PerkinElmer, Inc.) (23). After
the tumors had been measured, the mice were divided into four
different groups with 5 mice/group: i) Vehicle, DMSO; ii) Baze, 8.8
mg/kg/mouse (24,25); iii) SCH, 25 mg/kg/mouse (16); and iv) Baze + SCH. Both drugs were
administered daily by intraperitoneal injection. All mice were
monitored using an IVIS instrument. Once experiments had been
completed at 2 months after injection, tumors were harvested,
weighed and snap-frozen in liquid nitrogen, and stored at −80°C.
Tumor tissue was ground into powder, lysed and separated by
SDS-PAGE to examine the expression of the downstream targets of
IL-6 and IL-8 in different mouse groups, as described in the
western blot analysis subsection. The method of euthanasia used for
the mice was CO2 asphyxiation followed by cervical
dislocation (CO2 was introduced into the chamber at a
rate of 30-70% of the chamber volume per min to minimize distress).
The maximum tumor diameter was 14.5 mm and the volume obtained was
988.2675 mm3. The use of mice was approved on June 11,
2019 by the Institutional Animal Care & Use Committee of the
University of Maryland (Baltimore, MD, USA).
Bioluminescent imaging
The development and growth of tumors were monitored
by detecting the bioluminescence through an IVIS™ Imaging System,
which has a high-sensitivity cooled charge-coupled camera mounted
in a light-tight box (21,23).
The images were collected and analyzed using Living Image™ software
(Version:4.5.2.18424; PerkinElmer, Inc.). The mice were
anaesthetized and injected intraperitoneally with d-luciferin
potassium salt solution (150 mg/kg), and then placed into the
imaging chamber. Bioluminescent images were collected per week
(days 7, 14, 22, 29, 36, 44 and 62) throughout the duration of the
study.
Anesthesia
Mice were anesthetized using pharmaceutical grade
isoflurane dispensed from a precision vaporizer in 100%
O2 delivery gas at 3.5–4.5% to an induction chamber with
charcoal scavenger attached (scavenger was weighed before use and
weight recorded on side of cannister to determine disposal point by
weight gain). For Xenogen Imaging, there was a dedicated XGI8 gas
inhalation anesthesia apparatus next to the Xenogen IVIS-200
imager. Once recumbent 20–30 sec and respiratory rate was noted to
slow, isoflurane was stopped, the chamber was flushed with 100%
O2 (to reduce technician exposure to anesthetic when
chamber is opened) and the animal was removed to continue
anesthesia with isoflurane at 1.5–2.5% via face mask inside the
imaging chamber. After the imaging was performed, mice were moved
to a clean cage with bedding. Mice were monitored until able to
ambulate normally prior to return to their assigned husbandry
rooms.
Statistical analysis
The significance of associations was determined
using GraphPad Prism 7 software (GraphPad Software, Inc.). Data for
experiments performed in triplicate are presented as the mean ±
standard error of the mean. One-way ANOVA and Tukey's post hoc test
were used to analyze the statistical differences among groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
IL-6 and IL-8 levels in human ovarian
cancer cells
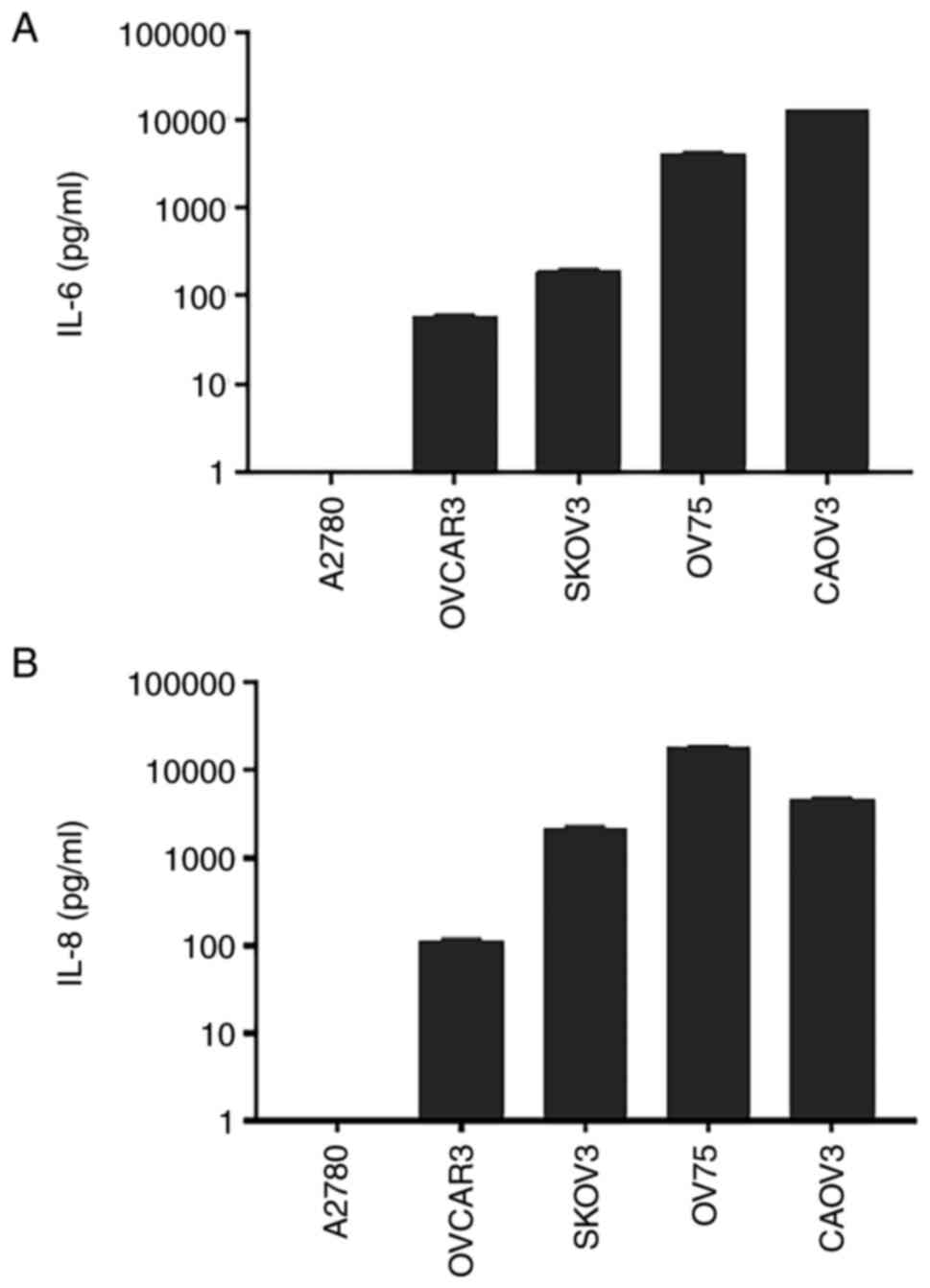
The secretion of IL-6 and IL-8 was examined in human
ovarian cancer cell lines. The present results demonstrated that
OVCAR3, SKOV3 and CAOV3 human ovarian cancer cell lines, and OV75
primary human ovarian cancer cells, secreted both IL-6 and IL-8 at
moderate to high levels [IL-6 (pg/ml): OVCAR3, 58.75±2.859; SKOV3,
191.4±7.469; OV75, 4,113±110.7; CAOV3, 12,517±11.15; IL-8 (pg/ml):
OVCAR3, 112.4±6.138; SKOV3, 2,215±68.58; OV75, 18,757±233.3; CAOV3,
4,776±53.34; Fig. 1]. As a
negative control, A2780 non-serous human ovarian endometroid
adenocarcinoma cells did not secrete detectable IL-6 and IL-8. In
general, endometrioid carcinoma is associated with an improved
prognosis compared with serous ovarian cancer (26). These results supported the current
hypothesis that IL-6 and IL-8 signaling pathways may serve as
potential therapeutic targets for the most aggressive human ovarian
cancer types.
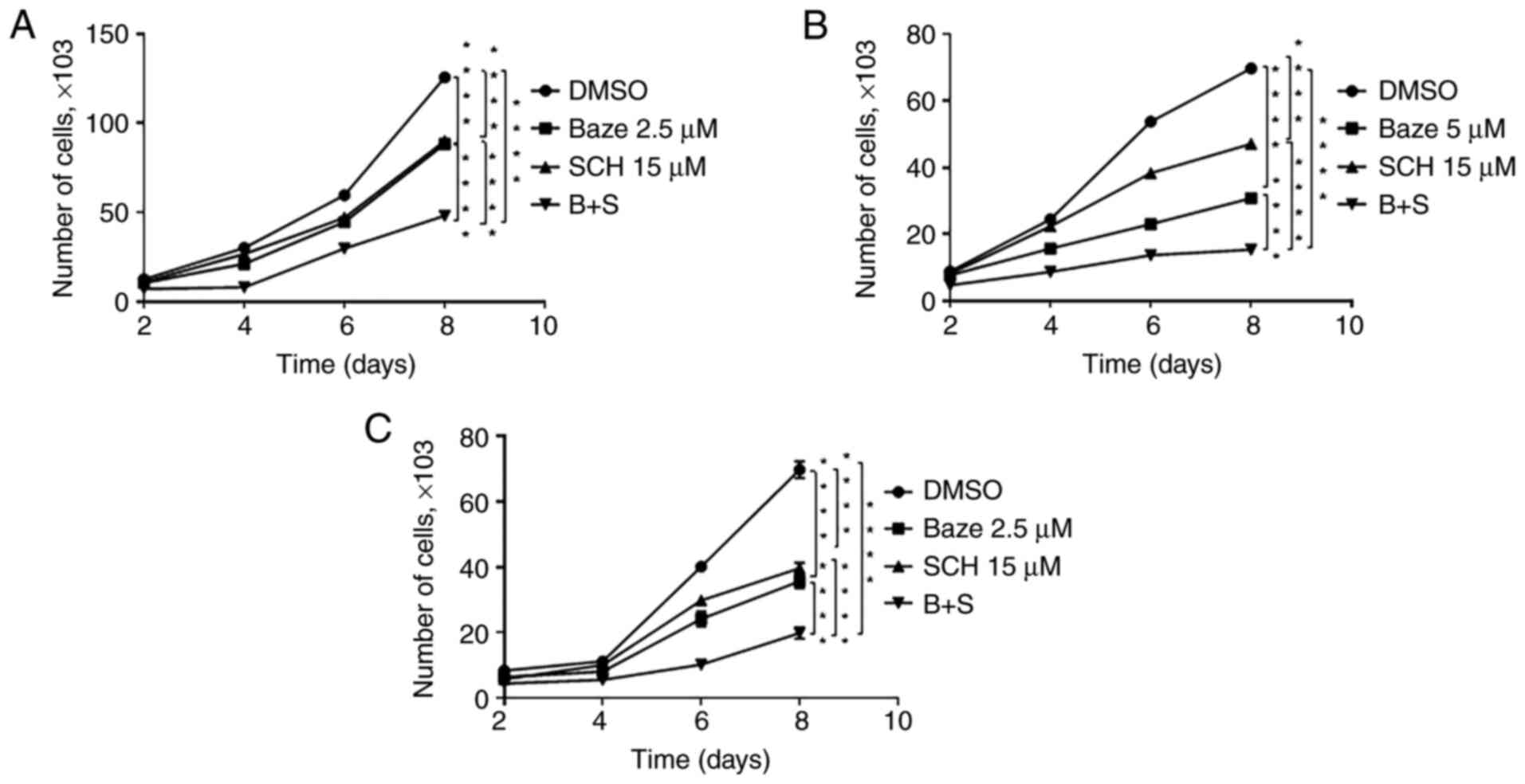
Baze + SCH treatment inhibits human
ovarian cancer cell proliferation
IL-6 and IL-8 signaling can both regulate cell
proliferation, adhesion, metastasis and invasion in human ovarian
cancer cells (4,27–29). Therefore, cell proliferation may
be decreased by IL-6 and IL-8 inhibitors (Baze and SCH). Thus, a
cell proliferation assay was first performed. As shown in Fig. 2, the proliferation of the ovarian
cancer cells was inhibited by each drug alone, but Baze + SCH
treatment exerted a stronger inhibitory effect compared with each
monotherapy.
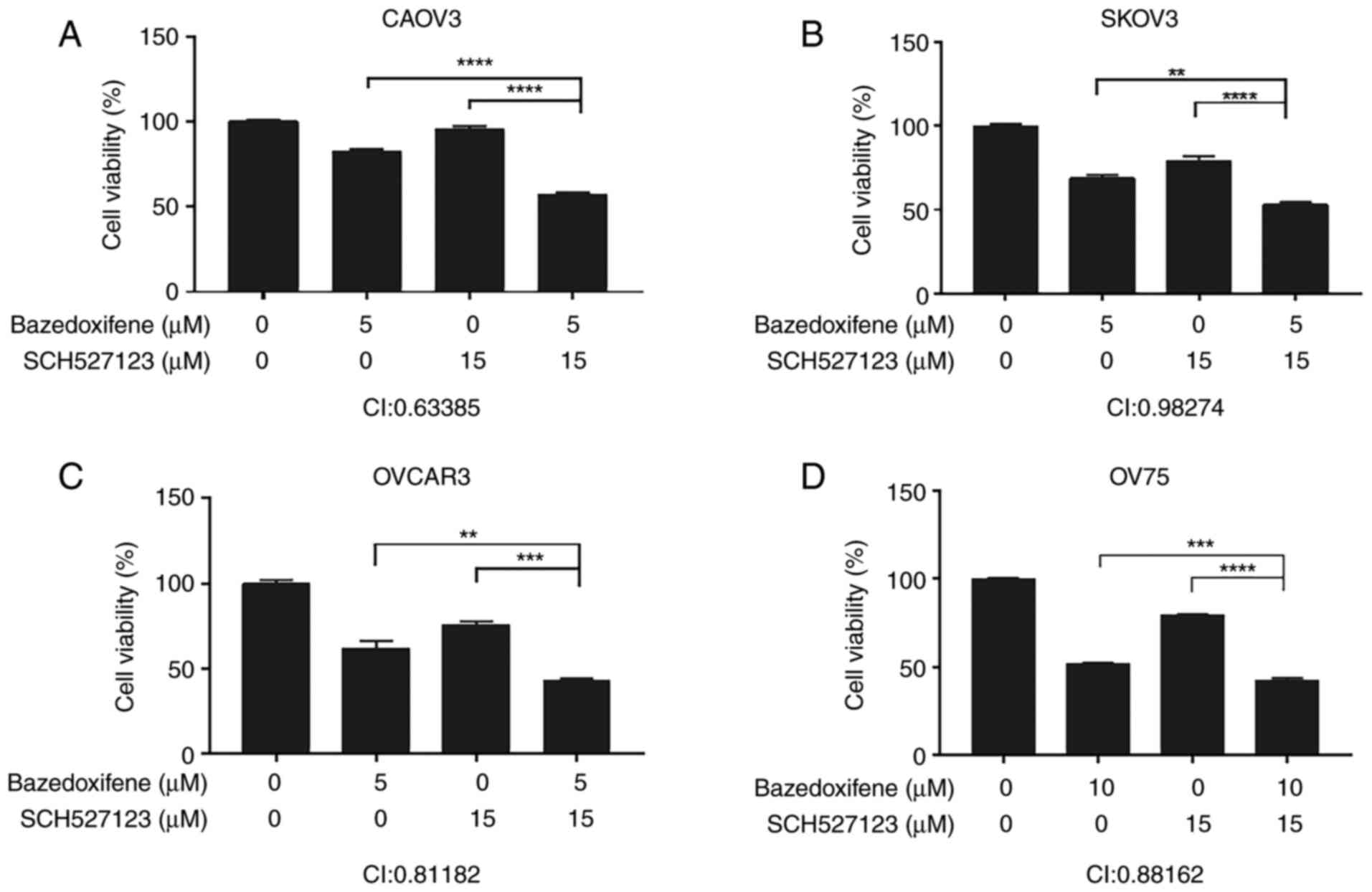
Baze + SCH synergistically inhibit human
ovarian cancer cell viability
It has been reported that IL-6 and IL-8 enhance the
proliferation of ovarian cancer cells (6). Therefore, the cell viability of
ovarian cancer cells may be inhibited by IL-6 and IL-8 inhibitors.
MTT assays were performed using Baze + SCH in OVCAR3, SKOV3, CAOV3
and OV75 human ovarian cancer cells As shown in Fig. 3, Baze + SCH treatment exerted a
stronger inhibitory effect on cell viability compared with each
monotherapy. Furthermore, the CI of each of the four ovarian cancer
cell lines was <1, indicating that the two drugs had a
synergistic effect.
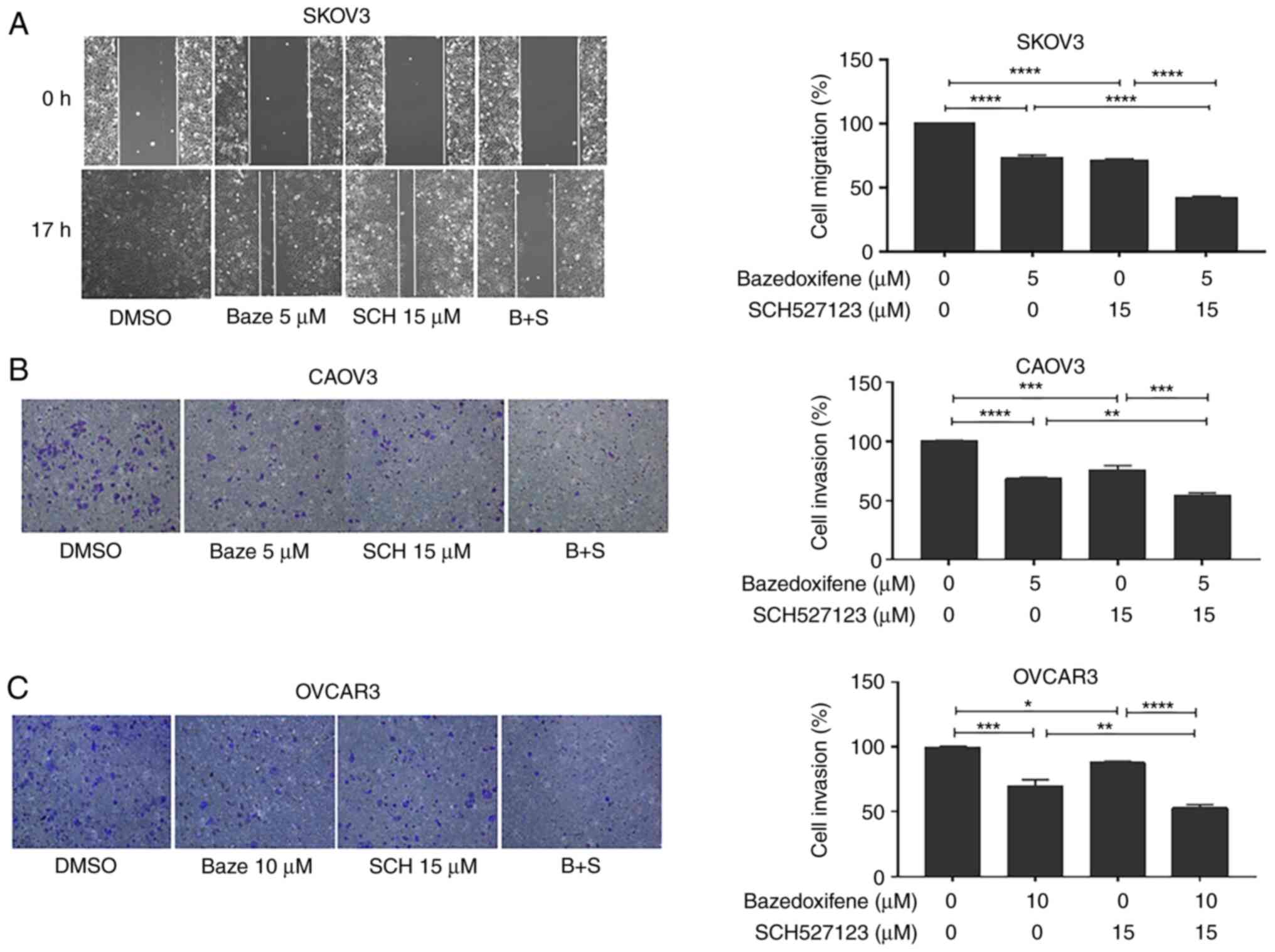
Baze + SCH treatment inhibits the
migration and invasion of human ovarian cancer cells
Cell migration and invasion are important steps in
tumor metastasis, which can indicate a poor prognosis (30,31). IL-6 and IL-8 are necessary and
sufficient to increase tumor cell migration (7). Therefore, cell migration and
invasion may be reduced by the inhibitors of IL-6 and IL-8. A cell
migration/wound healing assay was performed using the SKOV3 cell
line, as the monolayer phenotypes of the other two cell lines were
not suitable for this assay. An in vitro invasion assay was
also performed in OVCAR3 and CAOV3 cells using Matrigel, to detect
the inhibitory effect on cell invasion. Compared with the DMSO
control, cell migration (Baze, 72.73%; SCH, 71.82%; Baze + SCH,
41.82%; DMSO vs. Baze/SCH, Baze + SCH vs. Baze/SCH, P<0.0001;
Fig. 4A) and invasion (Fig. 4B and C) were inhibited by Baze and
SCH monotherapy (CAOV3: Baze, 68.73%; DMSO vs. Baze, P<0.0001;
SCH, 76.05%, DMSO vs. SCH, P=0.0002; Baze + SCH, 54.8%, Baze + SCH
vs. Baze, P<0.01; Baze + SCH vs. SCH, P<0.001; OVCAR3: Baze,
70.3%; DMSO vs. Baze, P<0.001; SCH, 87.98%, DMSO vs. SCH,
P<0.05; Baze + SCH, 53.06%, Baze + SCH vs. Baze, P<0.01; Baze
+ SCH vs. SCH, P<0.0001). Baze + SCH treatment resulted in a
greater inhibitory effect on cell migration and invasion compared
with each monotherapy. These results indicated that Baze + SCH may
be used to treat or prevent ovarian cancer cell invasion.
Baze + SCH synergistically inhibit the
expression of targeted genes downstream of the IL-6 and IL-8
pathways in human ovarian cancer cells
Since there are several interactions between the
IL-6 and IL-8 pathways (5,7),
western blot analysis was performed to determine which downstream
targeted genes were synergistically inhibited by these two drugs.
Two ovarian cancer cell lines were seeded in 10-cm plates and
treated with DMSO, a single drug or their combination. Protein
expression levels were analyzed. Compared with those in cells
treated with DMSO, the levels of p-STAT3, p-AKT, p-S6 and survivin
were decreased by monotherapy but this was not significant in most
cases (Fig. 5). The expression
levels following the combination treatment were significantly lower
than those of monotherapy and DMSO across all targets
(p-STAT3/STAT3, p-AKT/AKT, p-S6/S6 and survivin/GAPDH). These
results indicated that Baze + SCH synergistically inhibited the
expression of downstream targeted genes (p-STAT3/STAT3, p-AKT/AKT,
p-S6/S6 and survivin/GAPDH) of IL-6 and IL-8 pathways in human
ovarian cancer cells, which may have effects on cell viability,
migration and invasion.
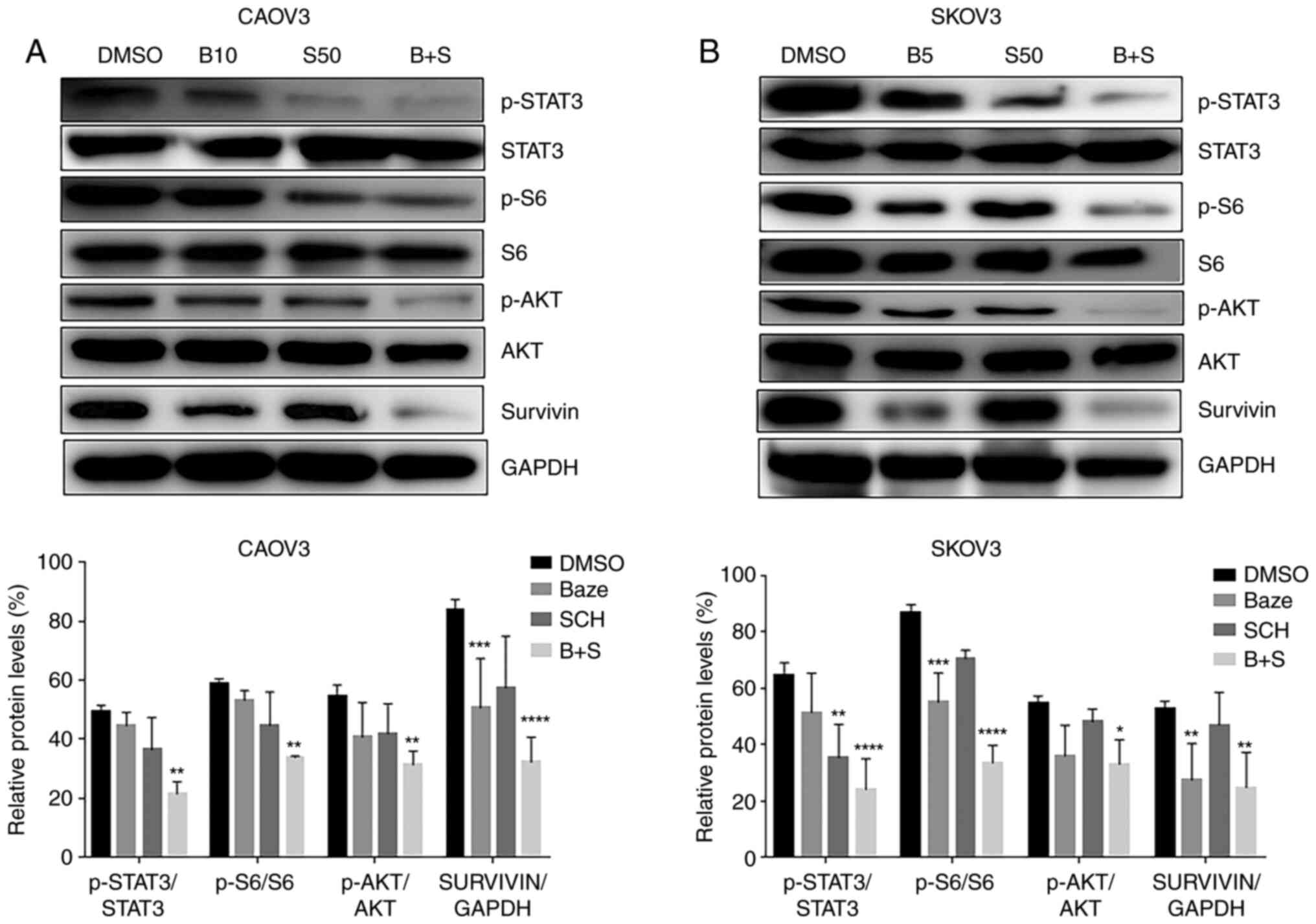 | Figure 5Baze and SCH inhibit the expression
of target genes downstream of the IL-6 and IL-8 pathways in human
ovarian cancer cells. SKOV3 and CAOV3 cells were treated with DMSO,
a single drug or their combination. The levels of p-STAT3, p-AKT,
p-S6 and survivin were determined by western blot analysis. (A)
CAOV3. (B) SKOV3. *P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001 (DMSO vs.
Baze, SCH or Baze + SCH). B+S, Baze + SCH; B10, BAZE 10 µM;
B5, BAZE 5 µM; BAZE, Baze; S50, SCH 50 µM;
p-phosphorylated; SCH, SCH. |
Baze + SCH treatment suppresses ovarian
cancer growth in vivo
The antitumor effect of Baze + SCH was investigated
in vivo. CAOV3-luciferase ovarian cancer cells
(1×107) were injected intraperitoneally into female
athymic nude mice. The mice were divided into four different
groups, with 5 mice per group: i) Vehicle, DMSO; ii) Baze, 8.8
mg/kg; iii) SCH, 25 mg/kg; iv) Baze + SCH. As shown in Fig. 6A and B, a significant reduction in
tumor growth was observed in the Baze or SCH single-treatment
groups. However, combination treatment with Baze + SCH exhibited a
greater inhibitory effect than monotherapy. In addition, compared
with the DMSO control group, combination treatment could
significantly reduce the levels of p-STAT3/STAT3, p-AKT/AKT,
p-S6/S6 and survivin/GAPDH in the tumor tissues (Fig. 6C).
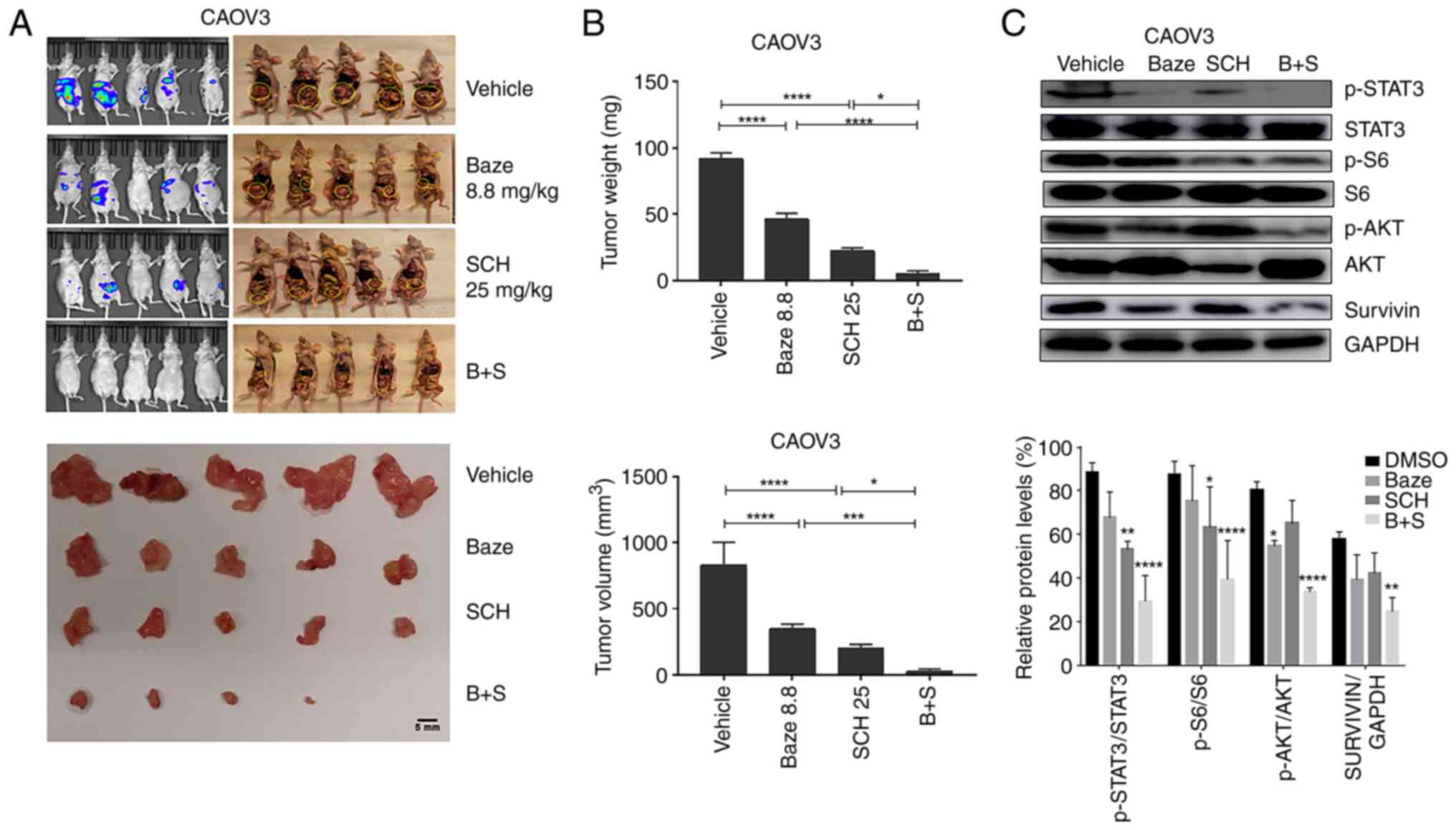 | Figure 6Baze and SCH suppress ovarian tumor
growth. (A) Images of CAOV3 ovarian tumors. Scale bar, 5 mm. (B)
Tumor weight and volume in different experimental groups.
*P<0.05, ***P<0.001 and
****P<0.0001. (C) Western blot analysis of the levels
of IL-6 and IL-8 downstream targets in representative tumors
(n=12). *P<0.05, **P<0.01 and
****P<0.0001 (DMSO vs. Baze, SCH or Baze + SCH). B+S,
Baze + SCH; Baze, Baze; p-, phosphorylated; Sch, SCH; Baze 8.8,
Baze 8.8 mg/kg; SCH25, SCH 25 mg/kg. |
In conclusion, the present in vitro and in
vivo findings indicated that the combined inhibition of IL-6
and IL-8 has the potential to suppress the growth of ovarian cancer
and prevent tumor invasion.
Discussion
Ovarian cancer has the highest mortality rate among
gynecological malignancies (2,32).
Detecting ovarian cancer early is challenging, and thus, >50% of
patients are diagnosed at advanced stages (33). Secondly, ~75% of patients with
ovarian cancer relapse due to intrinsic and acquired chemotherapy
resistance, which leads to cancer recurrence (2,33).
These are the two main reasons for the low 5-year overall survival
rate, which has remained at 20-35% in the past 20 years (2). Therefore, the continuous development
of novel treatment methods and drugs, such as targeted therapy,
which has the advantages of precise action and fewer side effects,
is crucial (33).
IL-6 is not only an inflammatory factor but also a
tumor-promoting factor, which serves an important role in
tumorigenesis, cancer cell survival and tumor progression in
multiple cancer types, including breast, cervical, colorectal,
oesophageal, head-and-neck, ovarian, pancreatic, prostate and renal
cancer (4,10,34,35). IL-6 binds to IL-6 receptor and
GP130 to form a signaling complex, which then activates down-
stream pathways (4,10,34,35). Janus kinase/STAT3 is the main
activated pathway which, in turn, leads to tyrosine phosphorylation
of STAT3 (4,10,34,35). p-STAT3 translocates into the
nucleus to induce the expression of target genes that promote tumor
cell proliferation, migration and invasion and suppress apoptosis
(4,10,34,35). In addition to the STAT3 signaling
pathway, several other pathways, such as the PI3K/AKT and MEK/MAPK
signaling pathways, may also be activated by IL-6 (34). IL-6 is one of the principal
oncogenic mediators in ovarian cancer (35). IL-6 is either constitutively
secreted directly by ovarian carcinoma cells or through secondary
inflammatory and tumor-infiltrating cells, including fibroblasts,
tumor-associated macrophages and T cells (36–38). Increasing evidence supports the
notion that high levels of serum IL-6 are associated with poor
prognosis, short survival, advanced disease and metastasis in
patients with ovarian cancer (4,27,39). Higher levels of serum and
peritoneal fluid IL-6 are also more commonly associated with
aggressive metastatic ovarian carcinomas (27). Autocrine production of IL-6
confers resistance to chemotherapeutic agents, such as cisplatin
and paclitaxel, in ovarian cancer cells (35,40). In addition, IL-6 signaling
regulates anchorage-independent proliferation, adhesion and
invasion in human ovarian cancer cells (4,27,28).
IL-8, also known as C-X-C motif chemokine ligand 8,
is a chemokine produced by macrophages, epithelial cancer cells and
other cells. IL-8 gene silencing decreases tumor growth through
antiangiogenic mechanisms (29).
IL-8 is upregulated in various human solid tumors (41) and is associated with advanced
tumor stage (P=0.019), high tumor grade (P=0.031) and poor survival
(29). The biological effects of
IL-8 are mediated by the binding of IL-8 to its receptors on the
surface of the cell membrane, two cell-surface G protein-coupled
receptors termed C-X-C motif chemokine receptor 1 (CXCR1) and CXCR2
(42,43). Following binding, PI3K/AKT, STAT3
and other signaling pathways may be activated, leading to tumor
progression through cell proliferation, migration, invasion and
epithelial-mesenchymal transition in several cancer types,
including ovarian cancer (8,9,29,44-48). Therefore, in addition to the IL-6
signaling pathway, the IL-8 signaling pathway is also an important
potential therapeutic target in ovarian cancer.
Since IL-6/GP130-targeted small molecule drugs are
still not available for clinical cancer therapy, IL-6/GP130 small
molecule inhibitors have been developed using multiple ligand
simultaneous docking and drug repurposing (49). In our previous study, Baze was
identified as a novel inhibitor of the IL-6/GP130 interaction using
a search of virtual hits on a drug database (49). Baze has also been approved as a
third-generation selective estrogen receptor modulator, as well as
a novel inhibitor of IL-6/GP130 (12–15,50). The direct binding of Baze to GP130
was supported by our previous study (49). It was also observed in our
previous study that Baze inhibits IL-6-mediated IL-6/IL-6Ra/GP130
heterotrimer in HPAC cells (data not shown), supporting the effects
of bazeodxifene on blocking IL-6 signaling, which has been
demonstrated to lead to growth suppression in breast (24) and colon cancer (25).
SCH is a potent allosteric CXCR1 and CXCR2
antagonist and has been shown to suppress tumor growth through the
inhibition of the NF-κB/AKT/MAPK signaling pathway (16,51). SCH has been found to target
IL-8/CXCR1/CXCR2 signaling in cancer cells, including in melanoma,
breast and pancreatic cancer cells (19,51,52). Furthermore, SCH alone has also
been tested in colon cancer cells and tumor models, and it was
found to have a limited in vivo activity on inhibiting tumor
growth and liver metastasis (16,53). SCH alone may have only a partial
tumor-suppressive activity in colon cancer, mainly due to the
inhibition of the IL-8 signaling pathway alone (without the
simultaneous inhibition of the IL-6 pathway), which was
insufficient for full tumor suppression (16,53). It may therefore be necessary to
target both IL-6 and IL-8 for cancer therapy.
In our previous study, it was demonstrated that the
combination of Baze and SCH for the treatment of triple-negative
breast and pancreatic cancer cells had synergistic inhibitory
effects (5); however, the
possible mechanism was not explored. In the present study, the
efficacy of Baze and SCH in ovarian cancer was examined, and it was
demonstrated that cell viability, migration and invasion were
inhibited. Tumor growth in vivo was also suppressed. Using
western blot analysis, the levels of p-AKT, p-STAT3, survivin and
p-S6 were found to be decreased by Baze + SCH. Baze + SCH caused a
greater inhibition of p-AKT, p-S6 and survivin levels compared with
monotherapy. Therefore, the dual inhibition of IL-6 and IL-8 may
inhibit cross-talk between the STAT3 (survivin) and AKT (S6)
signaling pathways. Notably, to the best of our knowledge, this was
the first study reporting the application of Baze + SCH in ovarian
cancer and its potential mechanism.
In conclusion, the present results indicated that
IL-6 and IL-8 are secreted by ovarian cancer cells and may confer
an aggressive phenotype. The combination of IL-6/GP130 inhibitor
Baze and IL-8/CXCL2 inhibitor SCH could synergistically inhibit
cell viability, and inhibited the migration and invasion of human
ovarian cancer cells in vitro In addition, Baze + SCH was
also found to suppress ovarian cancer growth in vivo.
Therefore, Baze + SCH treatment may be a potentially effective
approach for ovarian cancer therapy and should be studied
further.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
RZ and JL conceived the study. JL supervised the
study. RZ carried out the experiments and wrote the manuscript with
support from JL, JR and DMR. JR and DMR provided the OV75 cell line
and contributed to editing the manuscript. RZ and JL confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The use of mice was approved on June 11, 2019 by the
Institutional Animal Care & Use Committee of the University of
Maryland (approval no. 1488GCC: Collection of Tissue Samples from
Women with Gynecologic Malignancies and Healthy Controls for
Laboratory Research, Baltimore, MD, USA).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Dr Richard Eckert
(Department of Biochemistry and Molecular Biology, University of
Maryland School of Medicine, Baltimore, MD 21201, USA) for his kind
assistance in providing the microscope during the wound healing
assay.
Funding
The present study was supported by the University of Maryland
School of Medicine and Comprehensive Cancer Center start-up
fund.
Abbreviations:
|
GP130
|
glycoprotein 130;
|
|
PS
|
penicillin/streptomycin;
|
|
CI
|
combination index
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
National Comprehensive Cancer Network.
NCCN clinical practice guidelines in oncology (CN Guidelines®),
ovarian cancer. Version 5. 2020:2020, https://www2.tri-kobe.org/nccn/guideline/breast/english/ovarian.pdf.
|
|
3
|
Coleman RL, Monk BJ, Sood AK and Herzog
TJ: Latest research and treatment of advanced-stage epithelial
ovarian cancer. Nat Rev Clin Oncol. 10:211–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Browning L, Patel MR, Horvath EB, Tawara K
and Jorcyk CL: IL-6 and ovarian cancer: Inflammatory cytokines in
promotion of metastasis. Cancer Manag Res. 10:6685–6693. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fu S and Lin J: Blocking interleukin-6 and
interleukin-8 signaling inhibits cell viability, colony-forming
activity, and cell migration in human triple-negative breast cancer
and pancreatic cancer cells. Anticancer Res. 38:6271–6279. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Y, Yang J, Gao Y, Du Y, Bao L, Niu W
and Yao Z: Regulatory effect of e2, IL-6 and IL-8 on the growth of
epithelial ovarian cancer cells. Cell Mol Immunol. 2:365–372.
2005.PubMed/NCBI
|
|
7
|
Jayatilaka H, Tyle P, Chen JJ, Kwak M, Ju
J, Kim HJ, Lee JSH, Wu PH, Gilkes DM, Fan R and Wirtz D:
Synergistic IL-6 and IL-8 paracrine signalling pathway infers a
strategy to inhibit tumour cell migration. Nat Commun. 8:155842017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wen J, Zhao Z, Huang L, Wang L, Miao Y and
Wu J: IL-8 promotes cell migration through regulating EMT by
activating the Wnt/β-catenin pathway in ovarian cancer. J Cell Mol
Med. 24:1588–1598. 2020. View Article : Google Scholar
|
|
9
|
Yung MM, Tang HW, Cai PC, Leung TH, Ngu
SF, Chan KK, Xu D, Yang H, Ngan HY and Chan DW: GRO-α and IL-8
enhance ovarian cancer metastatic potential via the CXCR2-mediated
TAK1/NFκB signaling cascade. Theranostics. 8:1270–1285. 2018.
View Article : Google Scholar :
|
|
10
|
Zhu X, Shen H, Yin X, Long L, Chen X, Feng
F, Liu Y, Zhao P, Xu Y, Li M, et al: IL-6R/STAT3/miR-204 feedback
loop contributes to cisplatin resistance of epithelial ovarian
cancer cells. Oncotarget. 8:39154–39166. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feng L, Qi Q, Wang P, Chen H, Chen Z, Meng
Z and Liu L: Serum levels of IL-6, IL-8, and IL-10 are indicators
of prognosis in pancreatic cancer. J Int Med Res. 46:5228–5236.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yadav A, Kumar B, Teknos TN and Kumar P:
Bazedoxifene enhances the anti-tumor effects of cisplatin and
radiation treatment by blocking IL-6 signaling in head and neck
cancer. Oncotarget. 8:66912–66924. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gennari L, Merlotti D, De Paola V, Martini
G and Nuti R: Bazedoxifene for the prevention of postmenopausal
osteoporosis. Ther Clin Risk Manag. 4:1229–1242. 2008. View Article : Google Scholar
|
|
14
|
Komm BS, Kharode YP, Bodine PV, Harris HA,
Miller CP and Lyttle CR: Bazedoxifene acetate: A selective estrogen
receptor modulator with improved selectivity. Endocrinology.
146:3999–4008. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma H, Yan D, Wang Y, Shi W, Liu T, Zhao C,
Huo S, Duan J, Tao J, Zhai M, et al: Bazedoxifene exhibits growth
suppressive activity by targeting interleukin-6/glycoprotein
130/signal transducer and activator of transcription 3 signaling in
hepatocellular carcinoma. Cancer Sci. 110:950–961. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ning Y, Labonte MJ, Zhang W, Bohanes PO,
Gerger A, Yang D, Benhaim L, Paez D, Rosenberg DO, Nagulapalli
Venkata KC, et al: The CXCR2 antagonist, SCH-527123, shows
antitumor activity and sensitizes cells to oxaliplatin in
preclinical colon cancer models. Mol Cancer Ther. 11:1353–1364.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shin HY, Yang W, Lee EJ, Han GH, Cho H,
Chay DB and Kim JH: Establishment of five immortalized human
ovarian surface epithelial cell lines via SV40 T antigen or HPV
E6/E7 expression. PLoS One. 13:e02052972018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chou TC: Theoretical basis, experimental
design, and computerized simulation of synergism and antagonism in
drug combination studies. Pharmacol Rev. 58:621–681. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fu S, Chen X, Lin HJ and Lin J: Inhibition
of interleukin 8/C-X-C chemokine receptor 1,/2 signaling reduces
malignant features in human pancreatic cancer cells. Int J Oncol.
53:349–357. 2018.PubMed/NCBI
|
|
20
|
Hall DM and Brooks SA: In vitro invasion
assay using Matrigel™: A reconstituted basement membrane
preparation. Methods Mol Biol. 1070:1–11. 2014. View Article : Google Scholar
|
|
21
|
Vassileva V, Moriyama EH, De Souza R,
Grant J, Allen CJ, Wilson BC and Piquette-Miller M: Efficacy
assessment of sustained intraperitoneal paclitaxel therapy in a
murine model of ovarian cancer using bioluminescent imaging. Br J
Cancer. 99:2037–2043. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Westfall SD and Skinner MK: Inhibition of
phosphatidylinositol 3-kinase sensitizes ovarian cancer cells to
carboplatin and allows adjunct chemotherapy treatment. Mol Cancer
Ther. 4:1764–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu T, Close D, Handagama W, Marr E, Sayler
G and Ripp S: The expanding toolbox of in vivo bioluminescent
imaging. Front Oncol. 6:1502016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian J, Chen X, Fu S, Zhang R, Pan L, Cao
Y, Wu X, Xiao H, Lin HJ, Lo HW, et al: Bazedoxifene is a novel
IL-6/GP130 inhibitor for treating triple-negative breast cancer.
Breast Cancer Res Treat. 175:553–566. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei J, Ma L, Lai YH, Zhang R, Li H, Li C
and Lin J: Bazedoxifene as a novel GP130 inhibitor for colon cancer
therapy. J Exp Clin Cancer Res. 38:632019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bouchard-Fortier G, Panzarella T, Rosen B,
Chapman W and Gien LT: Endometrioid carcinoma of the ovary:
Outcomes compared to serous carcinoma after 10 years of follow-up.
J Obstet Gynaecol Can. 39:34–41. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Isobe A, Sawada K, Kinose Y, Ohyagi-Hara
C, Nakatsuka E, Makino H, Ogura T, Mizuno T, Suzuki N, Morii E, et
al: Interleukin 6 receptor is an independent prognostic factor and
a potential therapeutic target of ovarian cancer. PLoS One.
10:e01180802015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Li L, Guo X, Jin X, Sun W, Zhang X
and Xu RC: Interleukin-6 signaling regulates anchorage-independent
growth, proliferation, adhesion and invasion in human ovarian
cancer cells. Cytokine. 59:228–236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Merritt WM, Lin YG, Spannuth WA, Fletcher
MS, Kamat AA, Han LY, Landen CN, Jennings N, De Geest K, Langley
RR, et al: Effect of interleukin-8 gene silencing with
liposome-encapsulated small interfering RNA on ovarian cancer cell
growth. J Natl Cancer Inst. 100:359–372. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
van Zijl F, Krupitza G and Mikulits W:
Initial steps of metastasis: Cell invasion and endothelial
transmigration. Mutat Res. 728:23–34. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bravo-Cordero JJ, Hodgson L and Condeelis
J: Directed cell invasion and migration during metastasis. Curr
Opin Cell Biol. 24:277–283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Salomon-Perzyński A, Salomon-Perzyńska M,
Michalski B and Skrzypulec-Plinta V: High-grade serous ovarian
cancer: The clone wars. Arch Gynecol Obstet. 295:569–576. 2017.
View Article : Google Scholar
|
|
33
|
Mantia-Smaldone GM, Edwards RP and Vlad
AM: Targeted treatment of recurrent platinum-resistant ovarian
cancer: Current and emerging therapies. Cancer Manag Res. 3:25–38.
2011.PubMed/NCBI
|
|
34
|
Johnson DE, O'Keefe RA and Grandis JR:
Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev
Clin Oncol. 15:234–248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yousefi H, Momeny M, Ghaffari SH,
Parsanejad N, Poursheikhani A, Javadikooshesh S, Zarrinrad G,
Esmaeili F, Alishahi Z, Sabourinejad Z, et al: IL-6/IL-6R pathway
is a therapeutic target in chemoresistant ovarian cancer. Tumori.
105:84–91. 2019. View Article : Google Scholar
|
|
36
|
Watson JM, Sensintaffar JL, Berek JS and
Martinez-Maza O: Constitutive production of interleukin 6 by
ovarian cancer cell lines and by primary ovarian tumor cultures.
Cancer Res. 50:6959–6965. 1990.PubMed/NCBI
|
|
37
|
Yao X, Huang J, Zhong H, Shen N, Faggioni
R, Fung M and Yao Y: Targeting interleukin-6 in inflammatory
autoimmune diseases and cancers. Pharmacol Ther. 141:125–139. 2014.
View Article : Google Scholar
|
|
38
|
Tanaka T and Kishimoto T: The biology and
medical implications of interleukin-6. Cancer Immunol Res.
2:288–294. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lane D, Matte I, Rancourt C and Piché A:
Prognostic significance of IL-6 and IL-8 ascites levels in ovarian
cancer patients. BMC Cancer. 11:2102011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Y, Niu XL, Qu Y, Wu J, Zhu YQ, Sun WJ
and Li LZ: Autocrine production of interleukin-6 confers cisplatin
and paclitaxel resistance in ovarian cancer cells. Cancer Lett.
295:110–123. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Todorović-Raković N and Milovanović J:
Interleukin-8 in breast cancer progression. J Interferon Cytokine
Res. 33:563–570. 2013. View Article : Google Scholar
|
|
42
|
Waugh DJ and Wilson C: The interleukin-8
pathway in cancer. Clin Cancer Res. 14:6735–6741. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ha H, Debnath B and Neamati N: Role of the
CXCL8-CXCR1/2 axis in cancer and inflammatory diseases.
Theranostics. 7:1543–1588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li Y, Liu L, Yin Z, Xu H, Li S, Tao W,
Cheng H, Du L, Zhou X and Zhang B: Effect of targeted silencing of
IL-8 on in vitro migration and invasion of SKOV3 ovarian cancer
cells. Oncol Lett. 13:567–572. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Singh S, Wu S, Varney M, Singh AP and
Singh RK: CXCR1 and CXCR2 silencing modulates CXCL8-dependent
endothelial cell proliferation, migration and capillary-like
structure formation. Microvasc Res. 82:318–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang Y, Qu Y, Niu XL, Sun WJ, Zhang XL and
Li LZ: Autocrine production of interleukin-8 confers cisplatin and
paclitaxel resistance in ovarian cancer cells. Cytokine.
56:365–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang Y, Xu RC, Zhang XL, Niu XL, Qu Y, Li
LZ and Meng XY: Interleukin-8 secretion by ovarian cancer cells
increases anchorage-independent growth, proliferation, angiogenic
potential, adhesion and invasion. Cytokine. 59:145–155. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zheng T, Ma G, Tang M, Li Z and Xu R: IL-8
secreted from M2 macrophages promoted prostate tumorigenesis via
STAT3/MALAT1 pathway. Int J Mol Sci. 20:982018. View Article : Google Scholar
|
|
49
|
Li H, Xiao H, Lin L, Jou D, Kumari V, Lin
J and Li C: Drug design targeting protein-protein interactions
(PPIs) using multiple ligand simultaneous docking (MLSD) and drug
repositioning: Discovery of raloxifene and bazedoxifene as novel
inhibitors of IL-6/GP130 interface. J Med Chem. 57:632–641. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Romero IL, Lee W, Mitra AK, Gordon IO,
Zhao Y, Leonhardt P, Penicka CV, Mui KL, Krausz TN, Greene GL and
Lengyel E: The effects of 17β-estradiol and a selective estrogen
receptor modulator, bazedoxifene, on ovarian carcinogenesis.
Gynecol Oncol. 124:134–141. 2012. View Article : Google Scholar
|
|
51
|
Singh S, Sadanandam A, Nannuru KC, Varney
ML, Mayer-Ezell R, Bond R and Singh RK: Small-molecule antagonists
for CXCR2 and CXCR1 inhibit human melanoma growth by decreasing
tumor cell proliferation, survival, and angiogenesis. Clin Cancer
Res. 15:2380–2386. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Y, Liu J, Jiang Q, Deng J, Xu F, Chen
X, Cheng F, Zhang Y, Yao Y, Xia Z, et al: Human adipose-derived
mesenchymal stem cell-secreted CXCL1 and CXCL8 facilitate breast
tumor growth by promoting angiogenesis. Stem Cells. 35:2060–2070.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Varney ML, Singh S, Li A, Mayer-Ezell R,
Bond R and Singh RK: Small molecule antagonists for CXCR2 and CXCR1
inhibit human colon cancer liver metastases. Cancer Lett.
300:180–188. 2011. View Article : Google Scholar
|