Introduction
As one of the most common malignant tumor types in
females, breast cancer has exhibited a gradual increase in its
incidence rate in recent years (1). In China, >300,000 females are
diagnosed with breast cancer every year. In the eastern coastal
areas and economically developed cities, the incidence rate of
breast cancer has increased significantly (2). With the popularization of new
treatment strategies and methods, the global mortality rate of
breast cancer has gradually decreased (3). However, in China, particularly in the
vast rural areas, the downward trend of breast cancer mortality is
not significant. An important biological feature of breast cancer
is its high metastatic capacity, which has become a major problem
that limits the survival of patients with breast cancer and is
associated with poor prognosis (4). It is of great significance to
understand the specific mechanisms of breast cancer metastasis.
The tripartite motif-containing (TRIM) protein
family is a member of the RING domain family of E3 ubiquitin
ligases, which are composed of a RING-finger domain, two B-box
domains and a C-terminal domain, and the presence of the RING
domain suggests that TRIM proteins act as E3 ubiquitin linkages.
Enzymes are involved in various physiological processes of cells
(5,6). To date, >70 TRIM proteins have
been found in humans and mice (7).
They are encoded by ~71 genes and are divided into subfamilies I to
XI according to their different domains (8). Previous studies have reported that
TRIM proteins are extensively involved in a variety of cellular
physiological processes (9-11).
TRIM35, a member of the TRIM family, was found to have an important
role in the malignant progression of tumors (12,13).
In hepatocellular carcinoma (HCC), low TRIM35 expression promotes
the malignant proliferation of HCC cells (14). Increasing the expression of TRIM35
may significantly inhibit cell proliferation, metastasis and
invasion of breast cancer (15).
However, the association between TRIM35 and the development of
breast cancer has remained elusive. The aim of the present study
was to examine the potential biological function of TRIM35 in the
carcinogenesis of breast cancer.
Normally differentiated cells mainly rely on
oxidative phosphorylation of mitochondria for cell energy supply,
while most tumor cells rely on aerobic glycolysis (16). This phenomenon is called the
Warburg effect. The efficiency of aerobic glycolysis to produce ATP
is low, but it gives tumor cells numerous advantages (17). Tumor cells grow rapidly, and
excessive growth frequently puts tumor cells in a state of hypoxia,
so they turn off the aerobic oxidation of mitochondria, and energy
is provided by the anaerobic fermentation of glucose (18). Cells provide energy by enhancing
anaerobic fermentation (19).
After glucose is metabolized to pyruvate, it no longer undergoes
aerobic oxidation through the tricarboxylic acid cycle of
mitochondria, but is transformed into lactic acid through lactate
dehydrogenase and discharged from cells (20). A study has indicated that Wilms
tumor 1 associated protein enhances the stability of HK2 mRNA by
binding to the 3'-UTR m6A site, which mediates the Warburg effect
in gastric cancer to promote carcinogenesis (21). In triple-negative breast cancer,
microRNA-210-3p regulates HIF-1α and p53 downstream glycolytic
genes promote aerobic glycolysis and enhance the tumorigenicity of
breast cancer cells (22). In
colorectal tumors, condensin-2 complex subunit D3 increases the
level of E2F transcription factor 1 (E2F1) and interacts with E2F1,
recruiting more E2F1 to the promoter region of pyruvate
dehydrogenase kinase 1 (PDK1) and PDK3 genes, resulting in the
inhibition of pyruvate dehydrogenase activity and tricarboxylic
acid cycle (23). It further
promotes the reprogramming of glucose metabolism and enhances the
Warburg effect in colorectal carcinogenesis and colorectal cancer
progression (23). The above
studies suggest that various mechanisms regulate the role of the
Warburg effect in tumors. However, whether TRIM35 is related to the
Warburg effect and the specific regulatory mechanism has remained
elusive and was thus investigated in the present study.
Materials and methods
Patient samples
A total of 59 samples from patients with breast
cancer (tumor tissues and paired normal adjacent tissues) were
retrospectively obtained from the Department of Breast Surgery,
West China Hospital, Sichuan University (Chengdu, China) between
March 2019 and June 2020). The primary cancer tissues and the
paired normal adjacent tissues were immediately snap-frozen in
liquid nitrogen and stored at −80°C until use for mRNA and protein
analyses. The following exclusion criteria were used: i) Patients
with incomplete pathological and clinical information; ii) patients
with recurrent disease; iii) patients that had received
preoperative radiation or chemotherapy, or any other treatment; iv)
patients with other preoperative complications; v) patients with
cognitive or mental disorders; and vi) pregnant women. The
following inclusion criteria were used: i) Breast cancer tissue
samples were all pathologically and confirmed as breast cancer. All
patients provided written informed consent to participate in the
study. The present study was approved by the Institutional Review
Board of West China Hospital, Sichuan University (Chengdu,
China).
Cell lines and cell culture
Human BC cell lines, including T47D, MDA-MB-231,
MCF-7 and BT-474, and the human normal mammary epithelial cell line
MCF-10A, were purchased from the American Type Culture Collection.
T47D, MDA-MB-231, MCF-7 and BT-474 cells were cultured in DMEM
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS
(Biological Industries). MCF-10A cells were cultured in Minimum
Essential Medium (cat. no. CC-3151; Lonza Group, Ltd.) supplemented
with Mammary Epithelial Cell Growth Medium, MEGM®
SingleQouts® (cat. no. CC-4136; Lonza Group, Ltd.) and
10% FBS (Biological Industries). Cells were cultured in a
humidified atmosphere containing 5% CO2 at 37°C.
Cell transfection
Cells were transfected with the following plasmids:
Overexpression TRIM35 (Oe-TRIM35), short hairpin (sh)RNA targeting
TRIM35 for knockdown (Sh-TRIM35) and the respective negative
controls (NCs) (Shanghai GenePharma, Co., Ltd.). Plasmids were
transfected into breast cancer cells with Lipofectamine®
3000 (Invitrogen; Thermo Fisher Scientific, Inc.). The shRNA
sequence De-TRIM35 was constructed into the pLVX-shRNA2-BSD
lentiviral vector (Takara Bio USA, Inc.). The target sequences of
the shRNAs were as follows: TRIM35 1, 5′-CTT GCA TCT GTG GAA TCT
GTA-3′; TRIM35 2, 5′-AGT GTC AAG GAA GAA CTG GAT-3′; TRIM35 3,
5′-GCG CTG GAC CAG CTA CCG CTT-3′ (this sequence was used to
produce the main results); NC shRNA, 5′-TTC TCC GAA CGT GTC ACG
TAA-3′. TRIM35-overexpressing lentiviruses (cat. no. CD527A-1;
System Biosciences, LLC) were constructed into the lentiviral
vector pCDH-EF1α-MCS-T2A-Puro (System Biosciences, LLC). For the NC
group, empty vector was used as instead of the overexpression
vector.
In another experiment, the cells were cultured in
DMEM (10% FBS) containing the pyruvate kinase M2 (PKM2) activator
TEPP-46 (5 µM) (cat. no. ML-265; Med Chem Express) for 24 h
at 37°C as directed by the manufacturer.
RNA extraction and reverse
transcription-quantitative (RT-q) PCR
Total RNA was extracted from breast cancer tumor
tissues, paired normal adjacent tissues and cell lines using TRIzol
reagent (Takara Bio, Inc.). The extracted RNA was reverse
transcribed to cDNA with PrimeScript RT Reagent (Takara Bio, Inc.)
according to the manufacturer's instructions. Real-time qPCR was
performed with a Prime-Script® RT Reagent Kit (Takara
Bio, Inc.) according to the manufacturer's instructions and a
LightCycler system (Roche Diagnostics) was used for detection with
the following thermocycling conditions: Initial denaturation step
at 95°C for 3 min, followed by 45 cycles of denaturation at 95°C
for 20 sec, annealing at 60°C for 30 sec and extension at 72°C for
30 sec. GAPDH was used as an internal control. Relative RNA
expression levels were calculated by the 2−ΔΔCq method
(24). The primer sequences were
as follows: TRIM35 forward, 5′-CAT CGC CAA GCA CAA TCA GG-3′ and
reverse, 5′-GCG TTT TCG GCT CTT GTG TT-3′; GAPDH forward, 5′-GGA
GCG ACA TCC GTC CAA AAT-3′ and reverse, 5′-GGC TGT TGT CAA TCT TCT
CAT GG-3′.
Western blot analysis
Total protein was isolated from breast cancer tumor
tissues, paired normal adjacent tissues and cell lines using
protein extraction buffer (RIPA lysis buffer; Beyotime Institute of
Biotechnology). The nuclear protein was extracted using the nuclear
protein extraction kit (cat. no. P0027; Beyotime Institute of
Biotechnology) according to the manufacturer's instructions. The
protein concentration was quantified using a BCA kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. Total protein was separated by 10% SDS-PAGE (20 µg
protein per lane) and transfer onto a PVDF membrane
(MilliporeSigma). The membranes were blocked for 1 h at room
temperature with 5% nonfat milk powder (Beyotime Institute of
Biotechnology). To detect the expression of PKM2 protein in
different conformations, the total protein extracted from the
sample could was not heat-denatured. Non-denatured gel sample
loading buffer (Beyotime Institute of Biotechnology) was used for
protein denaturation. In addition, protein electrophoresis was
performed using native PAGE running buffer (Beyotime Institute of
Biotechnology). The remaining experimental steps were the same as
those of conventional western blot experiments. The membranes were
probed at 4°C overnight with antibodies against TRIM35 (cat. no.
ab272582; 1:5,000 dilution; Abcam), PKM2 (cat. no. 15822-1-AP;
ProteinTech Group, Inc.), Bax (cat. no. ab32503; 1:1,000 dilution;
Abcam), Bcl-2 (cat. no. ab182858; 1:2,000 dilution; Abcam),
N-cadherin (cat. no. ab76011; 1:5,000 dilution; Abcam), Ki-67 (cat.
no. ab16667; 1:5,000 dilution; Abcam), E-cadherin (cat. no.
ab40772; 1:10,000 dilution; Abcam), twist1 (cat. no. ab175430;
1:1,000 dilution; Abcam) and β-actin (cat. no. ab8226; 1:5,000
dilution; Abcam). The membranes were then incubated with the
appropriate secondary antibodies (1:5,000 dilution; cat. nos.
bs-40296G-HRP and bs-40295G-HRP; Bioss) for 1 h at room
temperature. Finally, protein expression was analysed by
chemiluminescence reagents (Hyperfilm ECL).
The Cancer Genome Atlas (TCGA) and the
National Cancer Institute's Clinical Proteomic Tumor Analysis
Consortium (CPTAC) analysis
The UALCAN database (http://ualcan.path.uab.edu/cgi-bin/ualcan-res.pl)
was used to analyze the expression of TRIM35 in the TCGA and CPTAC
datasets.
Protein-protein interaction network
analysis
The Search Tool for the Retrieval of Interacting
Genes and proteins (STRING; https://cn.string-db.org) was used to analyze the
relationship between TRIM35 and PKM2.
Cell Counting Kit-8 (CCK-8) assay
Cells were transfected and then inoculated into
96-well plates (2×103 cells/well). After incubation for
the indicated durations, CCK-8 assay reagent (Dojindo Molecular
Technologies, Inc.) was added to each well containing cells and
plates were incubated according to the manufacturer's instructions.
The absorbance at 450 nm was recorded for each well using a
microplate reader to determine cell proliferation.
Cell migration and invasion assays
Cell invasion and migration assays were performed
using Transwell chambers (8 µm pore size; Costar; Corning,
Inc.). MDA-MB-231 and MCF-7 cells (1×105 cells/well)
were seeded into the upper chamber of the Transwell chambers with
high-glucose DMEM containing 1% FBS. Furthermore, 500 µl of
high-glucose DMEM containing 10% FBS was added to the matched lower
chamber. After incubation for 48 h, various proportions of
MDA-MB-231 and MCF-7 cells had moved to the lower chamber. The
cells were fixed with methanol at room temperature for 30 min and
stained with 0.1% crystal violet at room temperature for 20 min.
For the invasion assay, the inserts were precoated with
Matrigel® (1 mg/ml; Corning, Inc.).
Flow cytometric analysis of cell
apoptosis
For cell apoptosis, after transfection, MDA-MB-231
and MCF-7 cells were seeded into 6-well plates. Once they reached
confluence (60-70%), cells were collected and incubated with
Annexin V-FITC (5 µl) (Biogot Technology Co., Ltd.) and
propidium iodide (PI) solution (5 µl; Biogot Technology Co.,
Ltd.) at room temperature for 15 min according to the
manufacturer's instructions. Cells were subsequently suspended in
400 µl binding buffer (Biogot Technology Co., Ltd.). Cell
apoptosis progression was analyzed using flow cytometry (FACSAria;
BD Biosciences). All data were analyzed with ModFit version 4.0
(Verity Software House, Inc.)
Glucose uptake assay
After transfection for 36 h, MDA-MB-231 and MCF-7
cells were incubated in DMEM without any L-glucose or phenol red
for 8 h. The amount of glucose in the media was measured using a
Glucose Colorimetric Assay kit (cat. no. K606-100; BioVision, Inc.)
according to the manufacturer's instructions. Fresh DMEM (excluding
L-glucose and phenol red) was used for the negative control. A
total of three biological replicates were performed.
Intracellular pyruvate assay, lactate
assay and ATP assay
After transfection for 36 h, MDA-MB-231 and MCF-7
cells were incubated in phenol red-free DMEM without FBS for 4 h.
Subsequently, a pyruvate assay kit (cat. no. BC2205), lactate assay
kit (cat. no. BC2235) and ATP assay kit (cat. no. BC0300; all from
Beijing Solarbio Science & Technology Co., Ltd.) were used to
measure intracellular concentrations of pyruvate, lactate and ATP,
respectively, according to the manufacturer's instructions. The
relative absorbance was measured using a spectrophotometer (Thermo
Fisher Scientific, Inc.). The content of pyruvate, lactate and ATP
were calculated according to the product manual.
Mouse xenograft tumor model
Female BALB/c-nu mice (total number, n=20; age, 5
weeks; weight, 20-25 g) were purchased from the Shanghai
Experimental Animal Center and kept at the Experimental Animal
Center of West China Hospital of Sichuan University (Chengdu,
China). All animal experiments were performed in accordance with
the institutional guidelines and were approved by the Committee on
the Ethics of Animal Experiments of West China Hospital of Sichuan
University (Chengdu, China). For the in vivo xenograft
assay, MDA-MB-231 and MCF-7 cells (5×106) transfected
with TRIM35 overexpression vector or control were subcutaneously
injected into the mammary fat pads of the female athymic nude mice
(n=5 in each group). Tumor growth was recorded once every four days
with caliper measurements. Tumor volume was calculated according to
the following formula: Volume=(width2 × length)/2. At 30
days after injection, the mice were euthanized and tumors were
collected for analysis. For this, the mice were anesthetized with
1% pentobarbital sodium (45 mg/kg, intraperitoneal) and
subsequently euthanized by cervical dislocation (when the heart had
stopped completely, the mouse was determined to be dead). Body
weight loss >20% was considered a humane endpoint for
euthanasia.
Co-immunoprecipitation (Co-IP)
Cells were collected and lysed using RIPA Lysis
Buffer (cat. no. P0013B; Beyotime Institute of Biotechnology)
containing protease inhibitor (cat. no. P1011; Beyotime Institute
of Biotechnology). Subsequently, 3 µg of antibody was added
to the lysate and the mixture was incubated overnight at 4°C.
Protein A/G PLUS-Agarose beads (20 µl; cat. no. sc-2003;
Santa Cruz Biotechnology, Inc.) were then added to the mixture and
it was incubated on a rotator for 4 h at 4°C. The
bead-antibody-protein complexes were washed with precooled PBS
three times and then boiled for subsequent western blot analysis
(as described above). The antibodies used were as follows: PKM2
(cat. no. 15822-1-AP; 1:1,000 dilution; ProteinTech Group, Inc.)
and TRIM35 (cat. no. ab272582; 1:1,000 dilution; Abcam).
Ubiquitination assay
MDA-MB-231 and MCF-7 cells were lysed in 1%
SDS-containing RIPA buffer by sonication on ice. Lysates were then
treated with Protein A/G Plus-Agarose (cat. no. sc-2003; Santa Cruz
Biotechnology, Inc.) for 1 h at room temperature. Subsequently,
each sample was incubated with IgG (cat. no. 30000-0-AP; 1:5,00
dilution; ProteinTech Group, Inc.) overnight at 4°C. The nuclear
pellet was collected by centrifugation at 10,000 × g for 5 min at
4°C and subsequently washed four times using Protein A/G
Plus-Agarose beads (by absorbing the protein, washing the beads and
then releasing the protein again from the beads). The purified
proteins were separated by gradient SDS-PAGE (as in the protocol
for western blot analysis). Anti-PKM2 (cat. no. ab137852; 1:1,000
dilution; Abcam) or anti-ubiquitin antibody (cat. no. ab7780;
1:5,00 dilution; Abcam) was used for immunoblotting according to
the protocol for western blot analysis.
Immunohistochemistry
Tissues were fixed in 10% formalin for 12 h at room
temperature and embedded in paraffin for 8 h at room temperature.
The slice thickness of the tissue specimen is 4um. The tissue
specimen slides were deparaffinized twice with xylene for 15 min
each at room temperature and rehydrated three times in a descending
series of ethanol solutions (100, 100, 95 and 80%). Antigen
retrieval was performed in 10 mmol/l sodium citrate solution (pH
6.0) at 100°C for 16 min and the samples were then cooled for 30
min. Endogenous peroxidase activity was quenched using 3% hydrogen
peroxide and blocking non-specific binding was performed with 5%
goat serum (OriGene Technologies, Inc.) for 30 min at room
temperature. Subsequently, the slides were incubated with
anti-N-cadherin (cat. no. ab76011; 1:100 dilution; Abcam),
anti-twist1 (cat. no. ab175430; 1:1,000 dilution; Abcam),
anti-Ki-67 (cat. no. ab16667; 1:200 dilution; Abcam),
anti-E-cadherin (cat. no. ab40772; 1:500 dilution; Abcam),
anti-BCL-2 (cat. no. ab182858; 1:500 dilution; Abcam) and anti-BAX
(cat. no. ab32503; 1:100 dilution; Abcam) at 4°C overnight.
Following the primary antibody incubation, slides were incubated
with an anti-rabbit secondary IgG antibody (1:100; cat. no.
SAP-9100; OriGene Technologies, Inc.) at 37°C for 30 min. The
samples were stained with 3,3-diaminobenzidine and Mayer's
hematoxylin. Slides were visualized using a light microscope
(magnification, ×100 or ×200; Zeiss AG). Immunostaining results
were evaluated by two blinded pathologists. Staining was scored
according to the percentage of positive cells and staining
intensity. The scoring for staining intensity was as follows: 0,
negative; 1, weak; 2, moderate; and 3, strong. The scoring system
for the percentage of positive cells was as follows: Negative,
0-5%; 1, 6-25%; 2, 26-50%; 3, 51-75%; and 4, 76-100%. Sections with
a total combined score of <4 were classified as having low
expression, while sections with a score of ≥4 were considered to
have high expression. From each tissue sample, 5 regions were
randomly selected for evaluation.
Cell immunofluorescence
Cells were grown on coverslips that were coated with
lysin. Cells (60% confluency) were fixed with 4% paraformaldehyde
for 30 min at room temperature and permeabilized with 0.25% Triton
X-100 solution for 20 min at room temperature. Cells were washed
with PBS and then blocked in 5% bovine serum albumin (Beyotime
Institute of Biotechnology) for 1 h at room temperature. Coverslips
were incubated with antibodies against PKM2 (cat. no. 15822-1-AP;
1:1,000 dilution; ProteinTech Group, Inc.) overnight at 4°C. After
washing with PBS, cells were incubated with appropriate secondary
antibody (cat. no. ab150077; 1:5,00 dilution; Abcam) for 1 h at
room temperature and DAPI for 10 min at room temperature. Slides
were imaged using an inverted fluorescence microscope.
Statistical analysis
All data were analyzed using SPSS 22.0 software (IBM
Corporation) or GraphPad Prism version 7.0 (GraphPad Software,
Inc.). Values are expressed as the mean ± standard deviation.
Statistical differences (two groups) were analyzed by an unpaired
Student's t-test. A paired Student's t-test was used to analyze the
statistical difference of TRIM35 expression in breast cancer
tissues and adjacent tissues. ANOVA was used for multiple-group
comparisons and Tukey's test was used as the post-hoc test after
ANOVA. P<0.05 was considered to indicate a statistically
significant difference.
Results
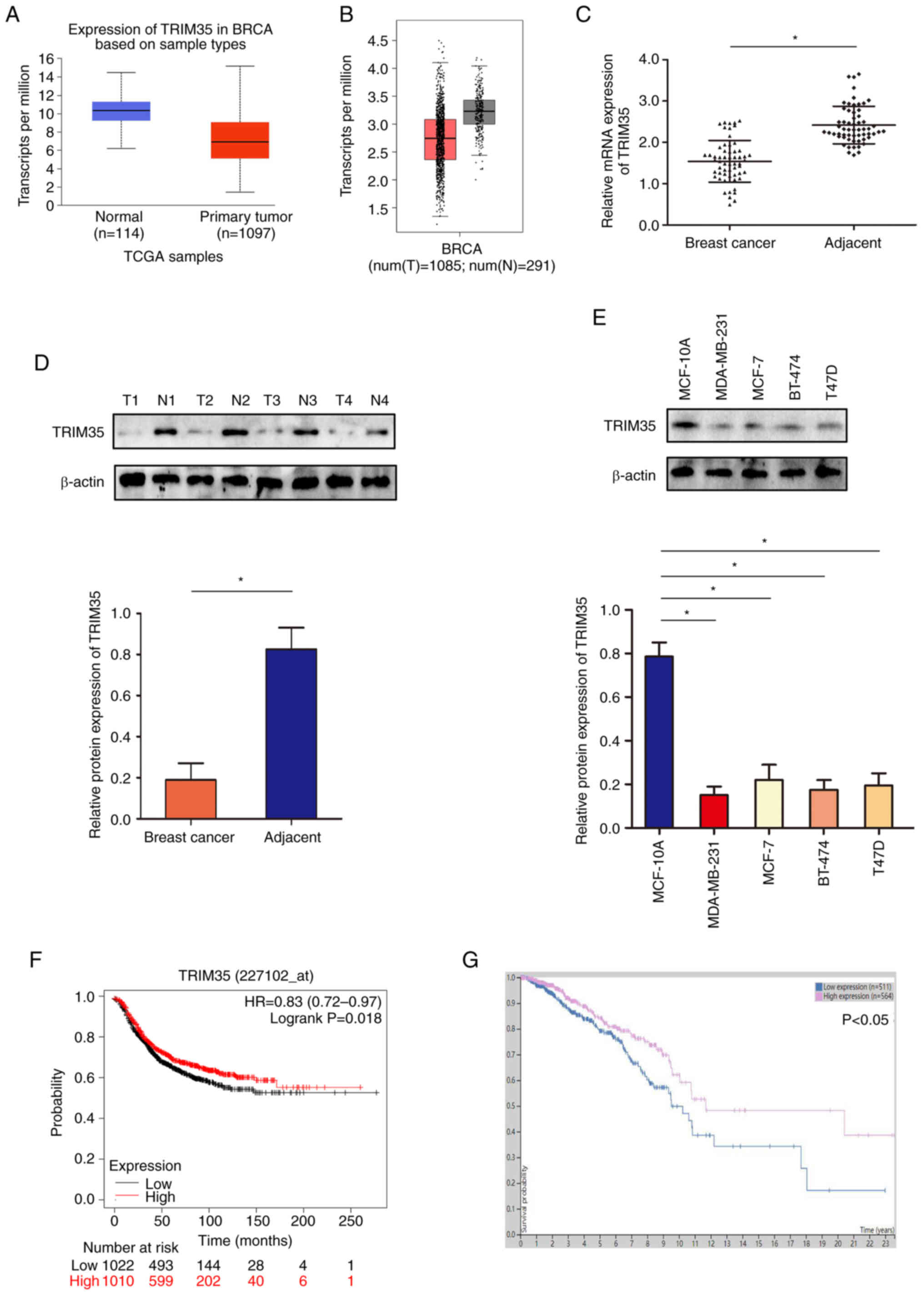
TRIM35 is downregulated in breast cancer
and low TRIM35 is associated with unfavorable survival in
patients
Analysis of the TCGA and GEPIA datasets indicated
that TRIM35 was low in breast cancer (Fig. 1A and B). In tissue samples
collected from patients with breast cancer (age range, 27-76 years;
all patients were female; demographic and clinicopathological data
provided in Table SI), western
blot and RT-qPCR analyses suggested that the expression of TRIM35
in breast cancer tissues was lower than that in adjacent tissues
(Fig. 1C and D). Furthermore, the
expression level of TRIM35 in breast cancer cell lines was
significantly lower than that in the normal breast cell line
(Fig. 1E). Through the analysis of
the relationship between the expression of TRIM35 and the prognosis
of patients with breast cancer in the GEO and TCGA databases, it
was revealed that those patients with low TRIM35 expression
exhibited significantly shorter overall survival than those with
high TRIM35 expression (Fig. 1F and
G).
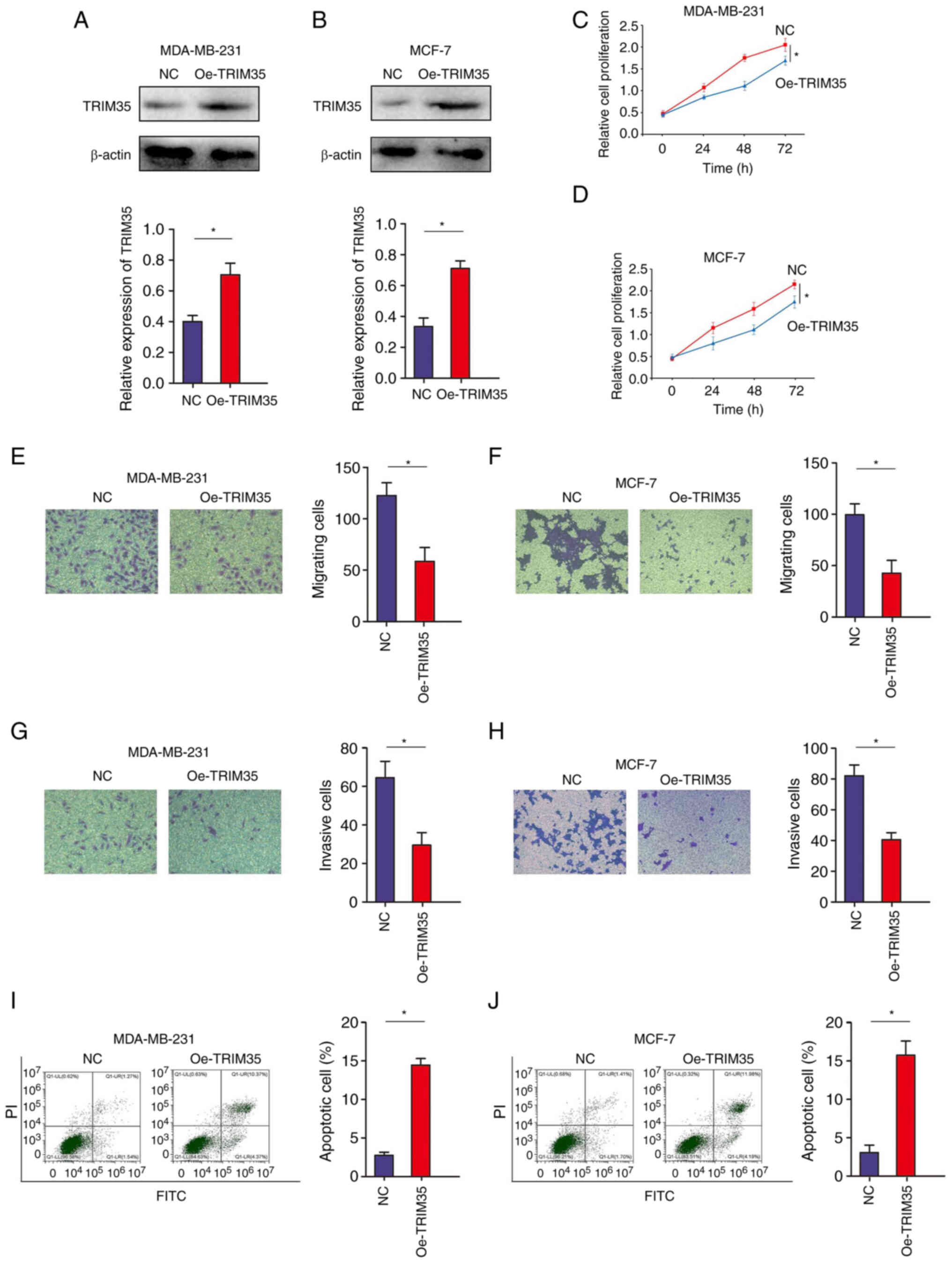
TRIM35 overexpression inhibits breast
cancer cell proliferation, migration and invasion and promotes
apoptosis
To further verify the role of TRIM35 in breast
cancer malignancy, gain-of-function experiments were performed in
MDA-MB-231 and MCF-7 cell lines. It was confirmed that TRIM35 was
effectively overexpressed in MDA-MB-231 and MCF-7 cell lines
(Fig. 2A and B). The CCK-8 assay
indicated that increasing the expression of TRIM35 inhibited number
of viable MDA-MB-231 and MCF-7 cells (Fig. 2C and D). The efficacy of TRIM35
overexpression and silencing using multiple clones are provided in
Figs. S1 and S2. Furthermore, a
Transwell assay suggested that increasing the expression of TRIM35
significantly inhibited the migration of MDA-MB-231 and MCF-7 cells
(Fig. 2E and F). In addition,
Transwell experiments indicated that increasing the expression of
TRIM35 significantly inhibited the invasion of MDA-MB-231 and MCF-7
cells (Fig. 2G and H). Increasing
the expression of TRIM35 promoted the apoptosis of MDA-MB-231 and
MCF-7 cells (Fig. 2I and J).
However, knockdown of TRIM35 promoted the malignant biological
behavior of breast cancer cells (Fig.
S3).
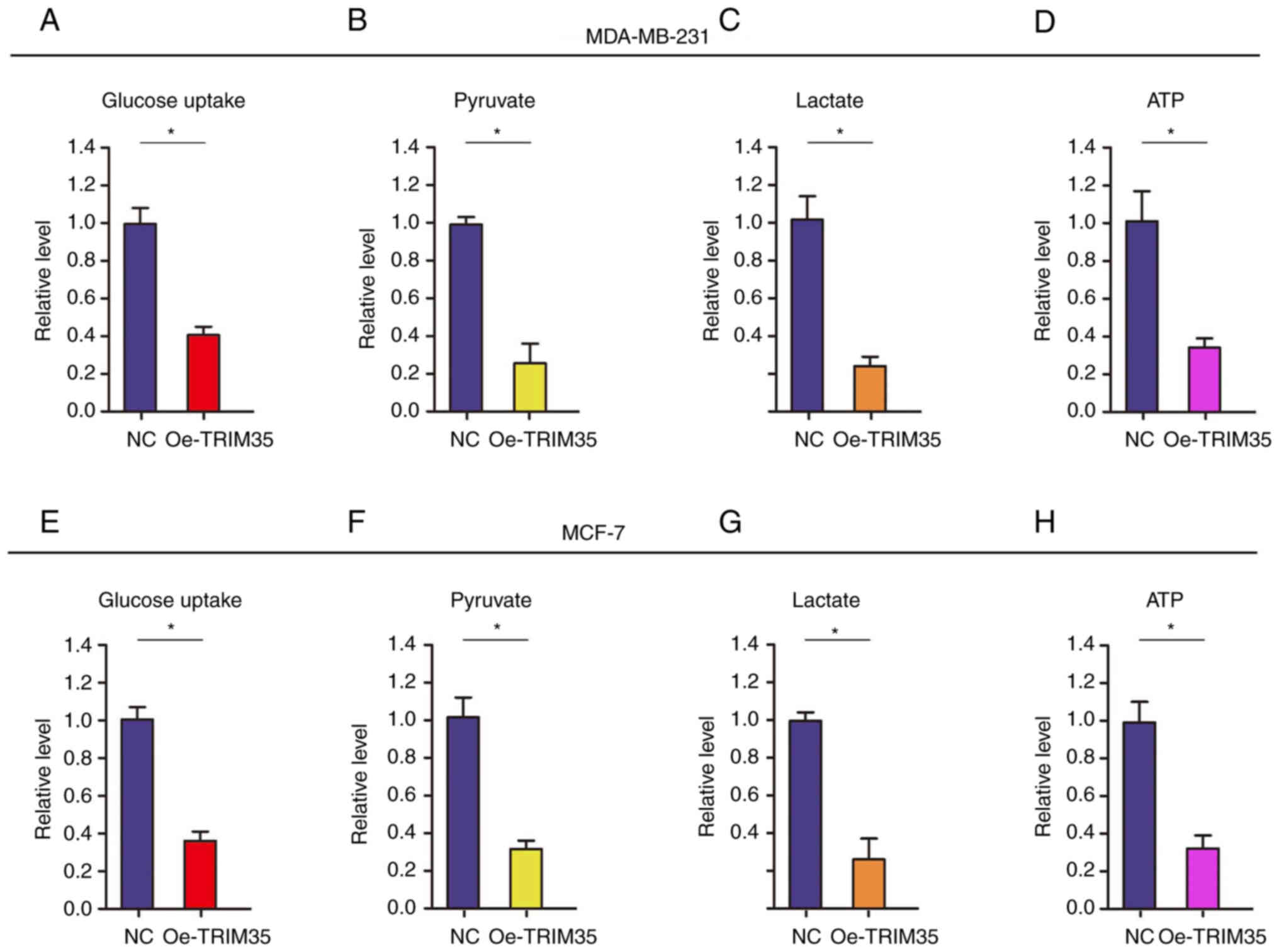
TRIM35 overexpression impedes energy
metabolism by restraining glycolysis in breast cancer cells
Metabolic reprogramming of tumor cells is an
important biological change, mainly manifested as enhanced
glycolysis, which provides effective energy and metabolites for the
rapid growth of tumor cells in different microenvironments
(25,26).
In the present study, it was analyzed whether TRIM35
affects glycolysis in breast cancer cells. The level of glucose
uptake was tested (Fig. 3),
revealing that it was significantly decreased after
TRIM35-overexpression in MDA-MB-231 and MCF-7 cells (Fig. 3A and E). In addition, the levels of
related metabolites, i.e. pyruvate and lactate, as well as the
production of ATP, were tested. The results revealed that the
levels of pyruvate and lactate, and the production of ATP were all
significantly decreased in TRIM35-overexpressing MDA-MB-231
(Fig. 3B-D) and MCF-7 cells
(Fig. 3F-H).
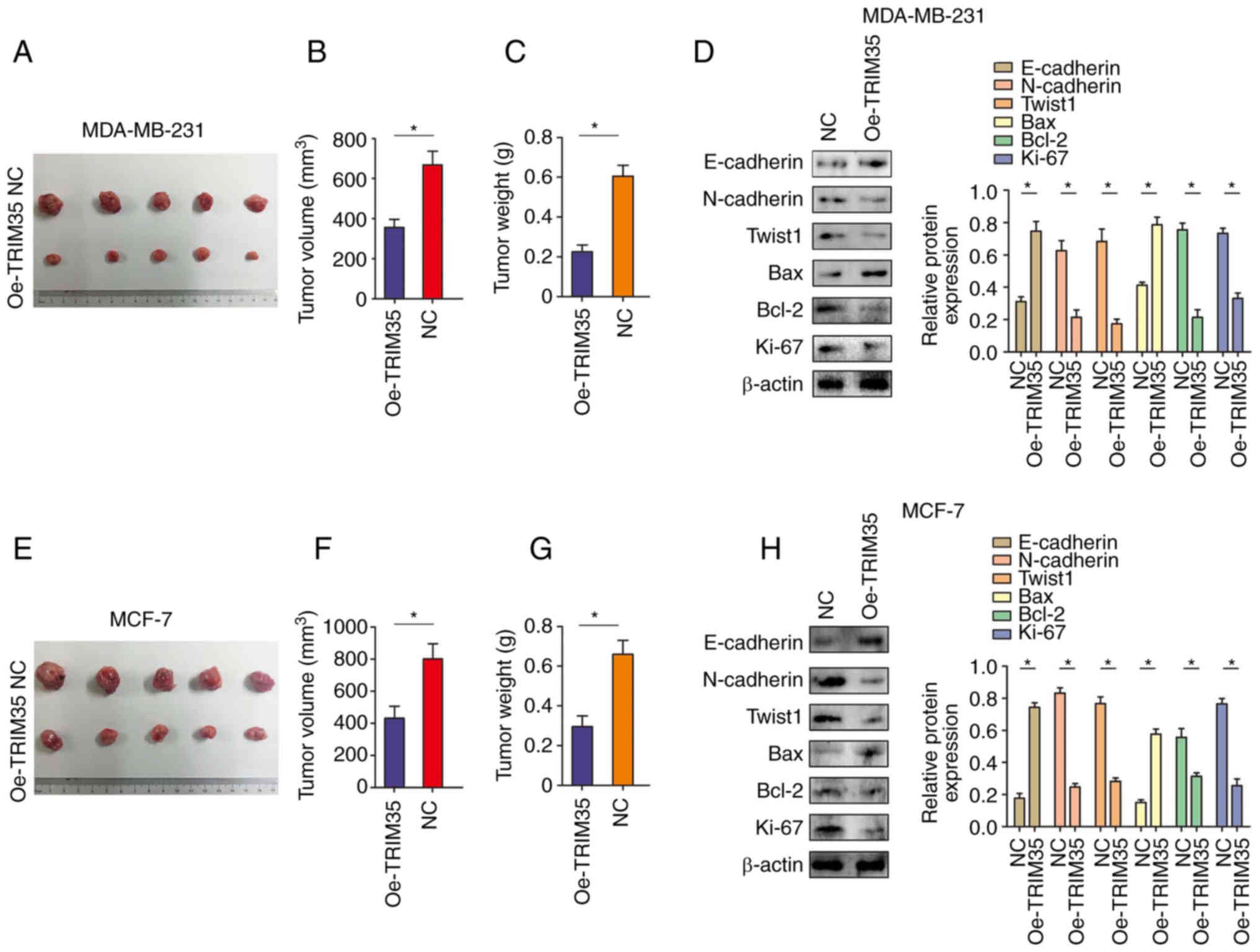
TRIM35 overexpression inhibits breast
cancer cell proliferation in vivo
The in vivo experiments indicated that
overexpression of TRIM35 significantly inhibited the growth of
tumors derived from MDA-MB-231 breast cancer cells (Fig. 4A-C). In addition, western blot
indicated that overexpression of TRIM35 significantly inhibited the
expression of N-cadherin, Twist1, Bcl-2 and Ki-67, while
significantly promoting the expression of E-cadherin and Bax in
MDA-MB-231-cell tumors (Fig. 4D).
In line with the above results, in tumors derived from MCF-7 cells
with overexpression of TRIM35, tumor growth was significantly
inhibited (Fig. 4E-G) and the
expression of N-cadherin, Twist1, Bcl-2 and Ki-67 were
significantly inhibited, while the expression levels of E-cadherin
and Bax were significantly promoted (Fig. 4H). Immunohistochemical staining
also confirmed the above results (Fig. S4).
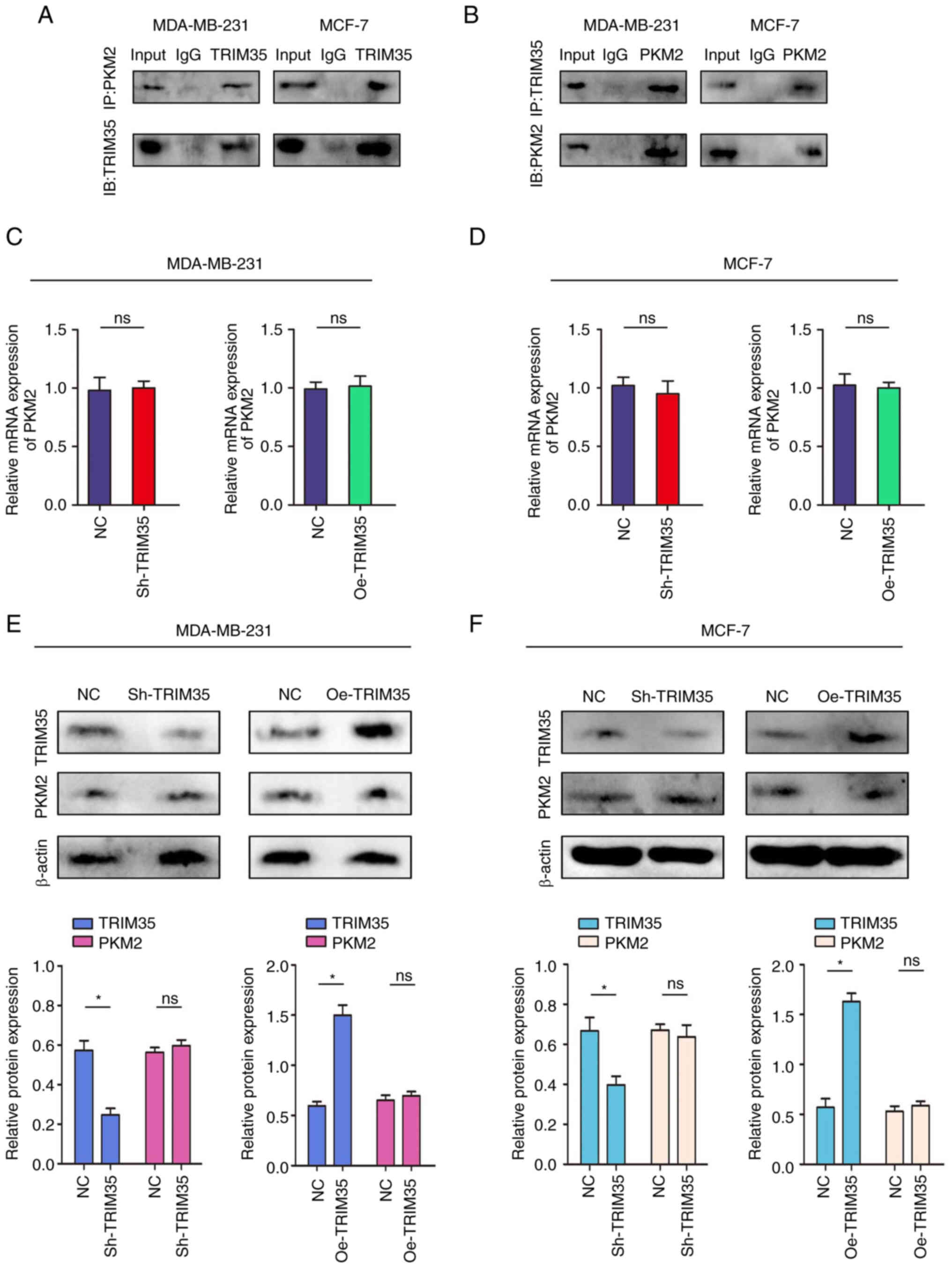
TRIM35 combines with PKM2 and
participates in the regulation of PKM2 in breast cancer
As a key protein of the Warburg effect, PKM2 is
widely involved in the glycolysis process of tumor cells (27,28).
The above-mentioned results of the present study indicated that
TRIM35 significantly regulates the glycolysis process in breast
cancer. Therefore, it was further examined whether TRIM35 has a
regulatory effect on PKM2.
The relationships between TRIM35 and PKM2 were
verified by using reciprocal Co-IP experiments carried out in
MDA-MB-231 and MCF-7 cells. The results confirmed the interaction
between endogenous TRIM35 and PKM2 (Fig. 5A and B). Subsequently, a
protein-protein interaction network prediction was performed,
suggesting that TRIM35 had a significant binding regulatory effect
on PKM2 (Fig. S5). However, no
changes in the mRNA or protein levels of PKM2 were observed after
changing the expression of TRIM35 (Fig. 5C-F).
TRIM35 regulates the ubiquitination of
PKM2, altering its tetramer-dimer ratio and promoting the Warburg
effect
PKM2 functions to convert phosphoenolpyruvate (PEP)
to pyruvate in the final step of glycolysis (29). Three dynamic types of PKM2 exist in
mammalian cells, namely monomeric, dimeric and tetrameric PKM2s,
but only dimeric and tetrameric PKM2s have pyruvate kinase activity
(30). Monomeric PKM2 is in an
inactive state. Dimeric PKM2 drives glucose-derived carbon towards
glycolysis (30). Tetrameric PKM2
promotes the transfer of glucose-derived carbons in the direction
of the respiratory chain (31).
The above-mentioned results of the present study indicated that
glycolytic function was inhibited after overexpression of TRIM35,
which also suggested that the changes in tetramers and dimers of
PKM2 may be the key target of TRIM35 in regulating PKM2.
Since TRIM35 is an ubiquitin E3 ligase, it was
further hypothesized that TRIM35 regulates the expression of PKM2
at the translational or post-translational level (32). Specifically, in the present study,
it was investigated whether TRIM35 affects the tetramer and dimer
changes of PKM2 by mediating changes in ubiquitination.
First, dimers of PKM2 were indicated to be highly
expressed in breast cancer cell lines (Fig. S6A-D). The ubiquitination levels of
PKM2 were enhanced in both the MDA-MB-231 and MCF-7 cell lines with
TRIM35 overexpression, which suggests that TRIM35 overexpression
allowed more PKM2 to enter the degradation process (Fig. 6A and B). In addition, the change in
ubiquitination has a regulatory role in the change in tetramers and
dimers of PKM2. Therefore, the effect of the change of
ubiquitination on the expression of PKM2 tetramer and dimer was
further analyzed. Overexpression of TRIM35 significantly increased
the expression of PKM2 tetramer, decreased the expression of PKM2
dimer and increased the ratio of PKM2 tetramer in MDA-MB-231 cells
(Fig. 6C-F). The same results were
also obtained in MCF-7 cells (Fig.
6G-J). Furthermore, knockdown of TRIM35 significantly decreased
the expression of PKM2 tetramer, increased the expression of PKM2
dimer and decreased the ratio of PKM2 tetramer in MDA-MB-231 and
MCF-7 cells (Fig. S7A-H). When
PKM2 dimer was reduced, PKM2 translocated from the nucleus to the
cytoplasm (Fig. S8A-D). The PKM2
activator TEPP-46 has been reported to reduce dimer formation and
promote tetramer formation (33).
Of note, treatment of breast cancer cells with TEPP-46
significantly inhibited number of viable cells (33), migration and invasion and promoted
cell apoptosis (Fig. S9).
Collectively, these data suggest that TRIM35 affects the expression
of the PKM2 tetramer and dimer by regulating the ubiquitination of
PKM2 and regulates the Warburg effect to promote the malignant
biological behavior of breast cancer (Fig. 6K).
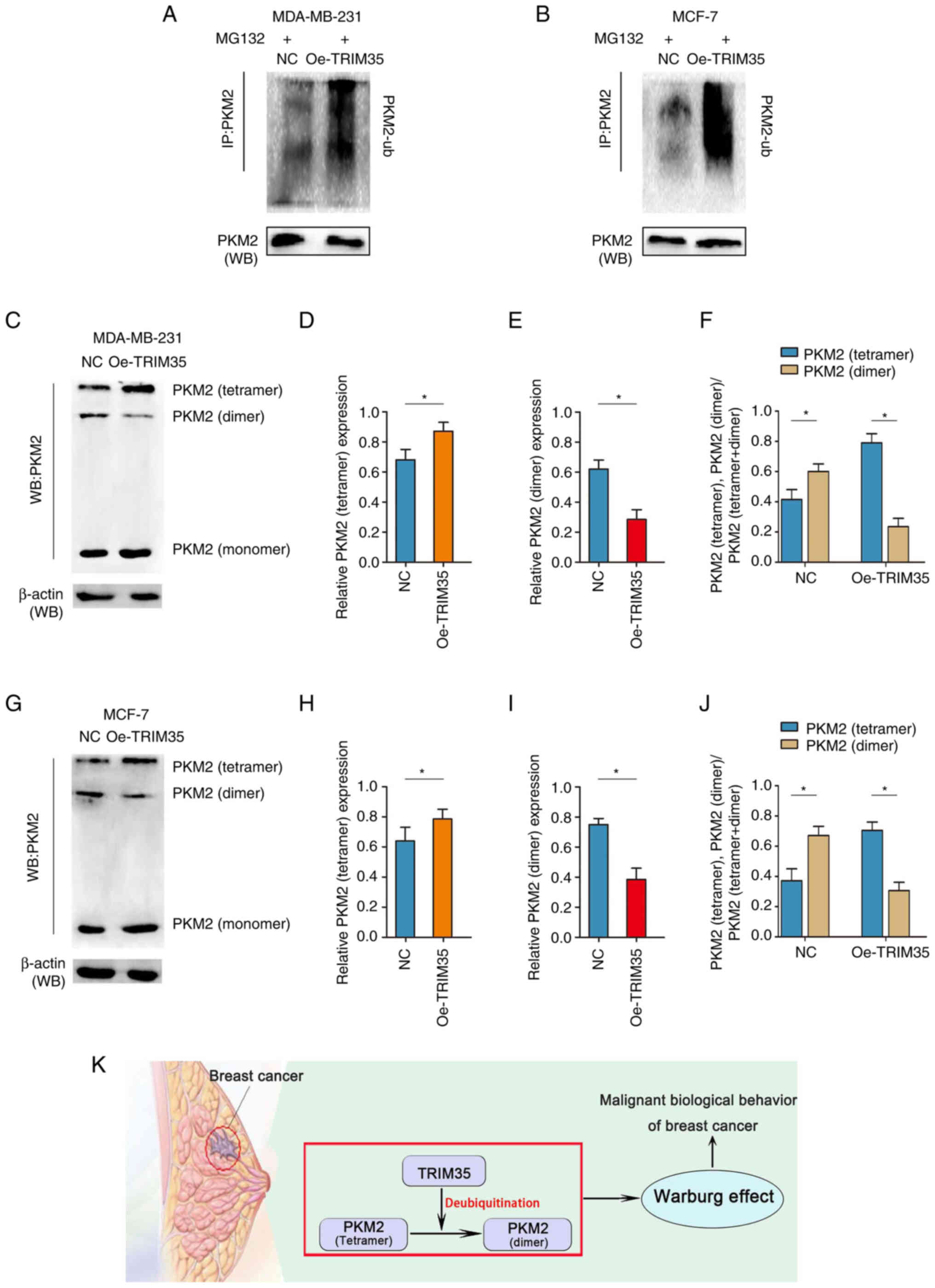 | Figure 6TRIM35 regulates the tetramer-dimer
transition of PKM2 through ubiquitination, thereby regulating the
Warburg effect. (A) The levels of PKM2-ub were examined by western
blot with anti-ubiquitin antibody in MDA-MB-231 cells. (B) The
levels of PKM2-ub were examined by western blot with anti-ubiquitin
antibody in MCF-7 cells. (C) Western blot analysis of the protein
expression levels of monomeric, dimeric and tetrameric PKM2 in
MDA-MB-231 cells. (D) Relative expression of tetrameric PKM2 in
MDA-MB-231 cells. (E) Relative expression of dimeric PKM2 in
MDA-MB-231 cells. (F) Proportion of PKM2 tetramers and PKM2 dimers
in MDA-MB-231 cells. (G) Western blot analysis of the protein
expression levels of monomeric, dimeric and tetrameric PKM2 in
MCF-7 cells. (H) Relative expression of tetrameric PKM2 in MCF-7
cells. (I) Relative expression of dimeric PKM2 in MCF-7 cells. (J)
Proportion of PKM2 tetramers and PKM2 dimers in MCF-7 cells. (K)
Schematic illustration of TRIM35 regulating the tetramer-dimer
transition of PKM2 through ubiquitination, thereby regulating the
Warburg effect. *P<0.05. TRIM35, tripartite
motif-containing 35; IP, immunoprecipitation; NC, negative control;
Oe, overexpression; PKM2, pyruvate kinase M2; PKM2-ub,
ubiquitinated PKM2 protein; WB, western blot. |
Discussion
Ubiquitination modification is one of the important
modifications of eukaryotic regulatory proteins after translation
(34). It affects cell
proliferation, differentiation, signal transduction and other
important physiological activities through the degradation,
localization and functional regulation of proteins (35). A series of processes of protein
ubiquitination modification are mainly completed by the ubiquitin
proteasome complex composed of ubiquitin activating enzyme E1,
ubiquitin binding enzyme E2 and ubiquitin ligase E3 (34). Most of the modified target proteins
are then degraded by the 26S proteasome (35). This process is called the ubiquitin
proteasome pathway. However, in recent years, an increasing number
of studies have indicated that ubiquitin modification is widely
involved in the assembly of signal pathway protein complexes and
the protein transport, as well as the activation and inactivation
of enzymes, which has made research on ubiquitin modification in
tumor signal transduction mechanisms a hot spot in this field
(36,37). Among the proteins that perform
ubiquitin modification, E3 ubiquitin ligase has become a potential
anti-tumor target due to its specific recognition of target
proteins. As a key protein of E3 ubiquitin ligase, TRIM35 is widely
involved in the regulation of various physiological and
pathological processes in tumors (38). In non-small cell lung cancer, the
expression of TRIM35 was observed to be increased in tumor samples,
and high expression of TRIM35 was closely related to poor clinical
prognosis of patients (39).
Inhibition of TRIM35 expression in non-small cell lung cancer cell
lines promoted cell proliferation, migration and invasion (39). Another study reported that E3
ubiquitin ligase TRIM35 is expressed at a low level in diffuse
large B-cell lymphoma (40).
Overexpression of TRIM35 inhibited the proliferation of diffuse
large B-cell lymphoma cells. A mechanistic study demonstrated that
TRIM35 mediates ubiquitination and degradation of 'clock', a key
regulator of the circadian rhythm (40). The present study further indicated
that the expression of TRIM35 was lower in breast cancer and its
low expression was associated with poor prognosis. A functional
study performed as part of the present paper indicated that
overexpression of TRIM35 significantly inhibited proliferation,
migration and invasion and promoted apoptosis of breast cancer
cells.
The Warburg effect is the main mode of ATP
production in tumor cells (41).
It changes from mitochondrial oxidative phosphorylation of glucose
to aerobic glycolysis (41). Its
essence is the rearrangement of the energy metabolism in tumor
cells (41). Pyruvate kinase is
the ultimate rate-limiting enzyme in glycolysis (42). PKM2 is the key enzyme in the
Warburg effect (42). It is able
catalyze PEP to transfer phosphate groups to adenosine diphosphate
to generate ATP (43). PKM2 has
two conformations in cells, a high enzyme activity tetramer and a
low enzyme activity dimer (44).
The low enzyme activity of the dimer is a necessary condition to
ensure the Warburg effect (44).
The low enzyme activity of PKM2 helps to make carbon atoms bind to
bioactive substances more rapidly, promote the rapid energy
production of glycolytic intermediates and enter the collateral
pathway, synthesize nucleic acids, amino acids and lipids without
accumulating reactive oxygen species, and provide energy and
metabolites for the malignant growth of tumor cells (44). Protein conformation and function
are closely related to posttranscriptional modification, which
frequently includes ubiquitination, phosphorylation and acetylation
(45). Zheng et al
(46) indicated that special at
rich sequence binding protein 1 was ubiquitinated through its
N-terminal ubiquitin-like domain, maintaining the tetramer
structure and having the role of an oncogene. French et al
(47) observed that ubiquitination
modification may promote the formation of casein kinase II tetramer
and inhibit the proliferation of tumor cells. The present results
are consistent with those of previous studies (43,44)
with regard to the expression of PKM2 dimer being inhibited, the
expression of PKM2 tetramer being promoted, and the proliferation,
invasion and migration of breast cancer cells being inhibited after
treatment of breast cancer cells with TEPP-46, while apoptosis was
increased. It was suggested that the conversion of PKM2 tetramer
and dimer has an important role in the malignant biological
behavior of breast cancer. But more importantly, the present
results suggested that TRIM35 is able to regulate the
ubiquitination modification of PKM2 and promote its transformation
from dimers to tetramers, inhibiting the Warburg effect and
reducing glycolysis in tumor cells. Of course, there are still
limitations to the present study. There are numerous key enzymes in
the glycolysis pathway involved in the Warburg effect, but in the
present study, only the regulatory mechanism of TRIM35 on PKM2 was
analyzed, while it was not analyzed whether TRIM35 has a regulatory
effect on other key enzymes. Future research by our group will
focus on the impact of TRIM35 on other key enzymes of the Warburg
effect.
Taken together, the results of the present study
indicated that the TRIM35-PKM2 axis promotes an aggressive
phenotype in tumors by enhancing glycolysis. TRIM35 is a potential
target for coordinating glycolysis in breast cancer. Protein
ubiquitination may be a key mechanism by which TRIM35 regulates the
transition between tetramers and dimers of PKM2.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HW and QL conceived and designed the study. XG, YJ
and ZW performed the experiments. HW and QL analyzed the data. HW
and QL wrote the manuscript. HW and QL checked and approved the
authenticity of the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The study was conducted with the approval of the
Ethics Committee of West China Hospital, Sichuan University
(Chengdu, China). Written informed consent was obtained from all
patients for the use of their tissue in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 82100655), Key Research and
Development Projects of the Sichuan Science and Technology
Department (grant no. 22QYCX0129) and Key Research and Development
Projects of the Sichuan Science and Technology Department (grant
no. 22ZDYF1209).
References
|
1
|
Harbeck N and Gnant M: Breast cancer.
Lancet. 389:1134–1150. 2017. View Article : Google Scholar
|
|
2
|
Fan L, Strasser-Weippl K, Li JJ, St Louis
J, Finkelstein DM, Yu KD, Chen WQ, Shao ZM and Goss PE: Breast
cancer in China. Lancet Oncol. 15:e279–e289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barzaman K, Karami J, Zarei Z,
Hosseinzadeh A, Kazemi MH, Moradi-Kalbolandi S, Safari E and
Farahmand L: Breast cancer: Biology, biomarkers, and treatments.
Int Immunopharmacol. 84:1065352020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Veronesi U, Boyle P, Goldhirsch A,
Orecchia R and Viale G: Breas cancer. Lancet. 365:1727–1741. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hatakeyama S: TRIM family proteins: Roles
in autophagy, immunity, and carcinogenesis. Trends Biochem Sci.
42:297–311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao G, Liu C, Wen X, Luan G, Xie L and
Guo X: The translational values of TRIM family in pan-cancers: From
functions and mechanisms to clinics. Pharmacol Ther.
227:1078812021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhan W and Zhang S: TRIM proteins in lung
cancer: Mechanisms, biomarkers and therapeutic targets. Life Sci.
268:1189852021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meroni G: TRIM E3 ubiquitin ligases in
rare genetic disorders. Adv Exp Med Biol. 1233:311–325. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Venuto S and Merla G: E3 ubiquitin ligase
TRIM proteins, cell cycle and mitosis. Cells. 8:5102019. View Article : Google Scholar :
|
|
10
|
Goyani S, Roy M and Singh R: TRIM-NHL as
RNA binding ubiquitin E3 ligase (RBUL): Implication in development
and disease pathogenesis. Biochim Biophys Acta Mol Basis Dis. Jan
6–2021.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Connacher RP and Goldstrohm AC: Molecular
and biological functions of TRIM-NHL RNA-binding proteins. Wiley
Interdiscip Rev RNA. 12. pp. e16202021, View Article : Google Scholar
|
|
12
|
Eberhardt W, Haeussler K, Nasrullah U and
Pfeilschifter J: Multifaceted roles of TRIM proteins in colorectal
carcinoma. Int J Mol Sci. 21:75322020. View Article : Google Scholar :
|
|
13
|
Watanabe M and Hatakeyama S: TRIM proteins
and diseases. J Biochem. 161:135–144. 2017.PubMed/NCBI
|
|
14
|
Chen Z, Wang Z, Guo W, Zhang Z, Zhao F,
Zhao Y, Jia D, Ding J, Wang H, Yao M and He X: TRIM35 Interacts
with pyruvate kinase isoform M2 to suppress the warburg effect and
tumorigenicity in hepatocellular carcinoma. Oncogene. 34:3946–3956.
2015. View Article : Google Scholar
|
|
15
|
Wang R, Huang KL and Xing LX: TRIM35
functions as a novel tumor suppressor in breast cancer by inducing
cell apoptosis through ubiquitination of PDK1. Neoplasma.
69:370–382. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shulman RG and Rothman DL: The glycogen
shunt maintains glycolytic homeostasis and the warburg effect in
cancer. Trends Cancer. 3:761–767. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lebelo MT, Joubert AM and Visagie MH:
Warburg effect and its role in tumourigenesis. Arch Pharm Res.
42:833–847. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kozal K, Jóźwiak P and Krześlak A:
Contemporary perspectives on the warburg effect inhibition in
cancer therapy. Cancer Control. Sep 23–2021.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Johar D, Elmehrath AO, Khalil RM, Elberry
MH, Zaky S, Shalabi SA and Bernstein LH: Protein networks linking
warburg and reverse warburg effects to cancer cell metabolism.
Biofactors. 47:713–728. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zam W, Ahmed I and Yousef H: The warburg
effect on cancer cells survival: The role of sugar starvation in
cancer therapy. Curr Rev Clin Exp Pharmacol. 16:30–38. 2021.
View Article : Google Scholar
|
|
21
|
Yu H, Zhao K, Zeng H, Li Z, Chen K, Zhang
Z, Li E and Wu Z: N6-methyladenosine (m6A) methyltransferase WTAP
accelerates the warburg effect of gastric cancer through regulating
HK2 stability. Biomed Pharmacother. 133:1110752021. View Article : Google Scholar
|
|
22
|
Du Y, Wei N, Ma R, Jiang S and Song D: A
miR-210-3p regulon that controls the warburg effect by modulating
HIF-1α and p53 activity in triple-negative breast cancer. Cell
Death Dis. 11:7312020. View Article : Google Scholar
|
|
23
|
Jing Z, Liu Q, He X, Jia Z, Xu Z, Yang B
and Liu P: NCAPD3 enhances warburg effect through c-myc and E2F1
and promotes the occurrence and progression of colorectal cancer. J
Exp Clin Cancer Res. 41:1982022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Ganapathy-Kanniappan S and Geschwind JF:
Tumor glycolysis as a target for cancer therapy: Progress and
prospects. Mol Cancer. 12:1522013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dayton TL, Jacks T and Vander Heiden MG:
PKM2, cancer metabolism, and the road ahead. EMBO Rep.
17:1721–1730. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu S, Guo Y, Zhang X, Liu H, Yin M, Chen
X and Peng C: Pyruvate kinase M2 (PKM2) in cancer and cancer
therapeutics. Cancer Lett. 503:240–248. 2021. View Article : Google Scholar
|
|
29
|
Zhang Z, Deng X, Liu Y, Liu Y, Sun L and
Chen F: PKM2, function and expression and regulation. Cell Biosci.
9:522019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zahra K, Dey T, Ashish, Mishra SP and
Pandey U: Pyruvate kinase M2 and cancer: The role of PKM2 in
promoting tumorigenesis. Front Oncol. 10:1592020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He X, Du S, Lei T, Li X, Liu Y, Wang H,
Tong R and Wang Y: PKM2 in carcinogenesis and oncotherapy.
Oncotarget. 8:110656–110670. 2017. View Article : Google Scholar
|
|
32
|
Ikeda K and Inoue S: TRIM proteins as RING
finger E3 ubiquitin ligases. Adv Exp Med Biol. 770:27–37. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Anastasiou D, Yu Y, Israelsen WJ, Jiang
JK, Boxer MB, Hong BS, Tempel W, Dimov S, Shen M, Jha A, et al:
Pyruvate kinase M2 activators promote tetramer formation and
suppress tumorigenesis. Nat Chem Biol. 8:839–847. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Popovic D, Vucic D and Dikic I:
Ubiquitination in disease pathogenesis and treatment. Nat Med.
20:1242–1253. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cockram PE, Kist M and Prakash S:
Ubiquitination in the regulation of inflammatory cell death and
cancer. Cell Death Differ. 28:591–605. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zou T and Lin Z: The involvement of
ubiquitination machinery in cell cycle regulation and cancer
progression. Int J Mol Sci. 22:57542021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Senft D, Qi J and Ronai ZA: Ubiquitin
ligases in oncogenic transformation and cancer therapy. Nat Rev
Cancer. 18:69–88. 2018. View Article : Google Scholar :
|
|
38
|
Jia D, Wei L, Guo W, Zha R, Bao M, Chen Z,
Zhao Y, Ge C, Zhao F, Chen T, et al: Genome-wide copy number
analyses identified novel cancer genes in hepatocellular carcinoma.
Hepatology. 54:1227–1236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang J, Xu Z and Yu B, Xu J and Yu B:
Tripartite motif containing 35 contributes to the proliferation,
migration, and invasion of lung cancer cells in vitro and in vivo.
Biosci Rep. Apr 30–2020.Epub ahead of print.
|
|
40
|
Tan X, Cao F, Tang F, Lu C, Yu Q, Feng S,
Yang Z, Chen S, He X, He J, et al: Suppression of DLBCL Progression
by the E3 ligase Trim35 is mediated by CLOCK degradation and NK
cell infiltration. J Immunol Res. 24:99958692021.
|
|
41
|
Liberti MV and Locasale JW: The warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu Z and Hunter T: Metabolic kinases
moonlighting as protein kinases. Trends Biochem Sci. 43:301–310.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Iqbal MA, Gupta V, Gopinath P, Mazurek S
and Bamezai RN: Pyruvate kinase M2 and cancer: An updated
assessment. FEBS Lett. 588:2685–2692. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li Z, Yang P and Li Z: The multifaceted
regulation and functions of PKM2 in tumor progression. Biochim
Biophys Acta. 1846:285–296. 2014.PubMed/NCBI
|
|
45
|
Wong N, Ojo D, Yan J and Tang D: PKM2
contributes to cancer metabolism. Cancer Lett. 356:184–191. 2015.
View Article : Google Scholar
|
|
46
|
Zheng M, Xing W, Liu Y, Li M and Zhou H:
Tetramerization of SATB1 is essential for regulating of gene
expression. Mol Cell Biochem. 430:171–178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
French AC, Luscher B and Litchfield DW:
Development of a stabilized form of the regulatory CK2beta subunit
that inhibits cell proliferation. J Biol Chem. 282:29667–29677.
2007. View Article : Google Scholar : PubMed/NCBI
|