Introduction
Bladder cancer (BC) is one of the most common types
of cancer worldwide and the most prevalent malignancy of the
urinary tract (1). BC accounts for
~200,000 deaths and 500,000 newly diagnosed cases worldwide
(2). Apart from age and geography,
the risk varies between sexes and is notably affected by exposure
to various carcinogens, with cigarette smoke being the most common
(1). Until recently, treatment for
BC, for several years, was limited to surgery and chemotherapy or
immunotherapy (3). Although these
therapies have certain beneficial effects in the initial stages of
treatment, BC patients are prone to relapse and metastasis in the
later stages, leading to treatment failure and patient death
(4). Therefore, it is of great
significance to search for more effective molecular-targeted
therapies for BC.
Pyrroline-5-carboxylate reductase 1 (PYCR1) is a
major enzyme involved in proline production in the mitochondria,
which not only participates in the metabolism of amino acids but is
also associated with energy metabolism and mitochondrial function
and plays a pivotal role in cell proliferation and apoptosis
(5,6). A previous study demonstrated the
potential role of PYCR1 in facilitating the progression of BC, and
PYCR1 may be utilized as a promising and attractive anticancer
target for BC therapy (7). The
transformation from mitochondrial oxidative phosphorylation to
aerobic glycolysis in metabolism is called the Warburg effect, and
the cancer progression attributed to aerobic glycolysis is often
related to the activation of oncogenes or the loss of tumor
suppressor genes (8). Aerobic
glycolysis is a scientifically identified hallmark of the
metabolism of cancer cells and targeting it may provide possible
drug-target cancer therapy strategies (9). Aerobic glycolysis participates in the
proliferation, migration, and invasion of hepatocellular carcinoma
(HCC) cells (10). EGFR is a
tyrosine kinase receptor that functions in the pathways controlling
aberrant and normal cell growth (11). EGFR expression is dysregulated in
various types of tumors, such as melanoma, colorectal cancer,
non-small cell lung cancer, and breast cancer (12-15).
The PI3K/AKT pathway is frequently activated in human cancers,
inducing cell malignant transformation and tumor angiogenesis and
apoptosis (16). The EGFR/PI3K/AKT
pathway stimulates aerobic glycolysis of cancer cells by regulating
the key enzymes of aerobic glycolysis, glucose transporter type 1
(GLUT1) (17,18). The PI3K/AKT axis is also correlated
with PYCR1 expression, and the mRNA levels of AKT1 and PIK3CB are
positively related to PYCR1 in gastric cancer tissues (19). However, whether PYCR1 regulates the
EGFR/PI3K/AKT pathway and aerobic glycolysis in BC remains unknown.
Small interfering RNAs (siRNAs) are widely used to knock down the
posttranscriptional expression of genes through complementarity to
mRNAs in the cytoplasm (20).
Various polymer-based nonviral gene vectors have been designed to
deliver siRNAs into the cytoplasm in an efficient manner due to the
tailored advantages of polymeric biomaterials, and siRNAs have
potential as cancer therapeutics (21-23).
Exosomes (Exos) are the smallest type of extracellular vesicles
(40-100 nm in diameter), consisting of a lipid bilayer surface
structure and containing various essential biomolecules including
DNA, lipid, protein, and RNA, acting as indispensable cell
communication mediators (24).
Exos have the characteristics of good biocompatibility and low
toxicity and immunogenicity and play significant roles in the
development and metastasis of cancers (25,26).
Therefore, delivery of siRNAs targeting PYCR1 (si-PYCR1) through
Exos for BC treatment may theoretically be of clinical value.
However, there are no reports, to the best of our knowledge, on the
mechanism of bone marrow mesenchymal stem cell (BMSC)-derived Exos
(BMSC-Exos) with PYCR1 expression knocked down on the aerobic
glycolysis and growth of BC cells. The aim of this study was to
elucidate the mechanism of PYCR1 in BC and to establish a
therapeutic modality using BMSC-Exos vector containing si-PYCR1 for
the management of BC, serving as a reference for elucidating the
pathogenesis of BC and developing novel targeted therapies for the
management of BC.
Materials and methods
Ethics statement
All procedures were authorized by the Academic
Ethics Committee of Hunan Provincial People's Hospital, The First
Affiliated Hospital of Hunan Normal University (approval no.
2021-071) and were strictly implemented in accordance with the
National Laboratory Animal Guide (27). Laboratory procedures were performed
in such a manner as to reduce the pain and discomfort caused to
mice. The need for approval for the use of primary human BMSCs was
waived by the Ethics Committee of Hunan Provincial People's
Hospital, The First Affiliated Hospital of Hunan Normal
University.
Database analysis of PCYR1 expression in
BC
The StarBase database (starbase.sysu.edu.cn/panCancer.ph) was used to
determine PCYR1 expression in BC as follows: Select 'Gene
Differential Expression', search 'PCYR1 (ENSG00000183010)', and
select 'Bladder Urothelial Carcinoma'. The results were downloaded
from the database.
Cell culture
Normal bladder epithelial cell SV-HUC-1 and BC cell
lines T24, 5637, RT4, and TCCSUP (ATCC) were cultured in the
RPMI-1640 medium (all from Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS, 100 U/ml penicillin, and 100
µg/ml streptomycin in a humidified incubator (Eppendorf)
supplied with 5% CO2 air at 37°C. Cells in the
logarithmic growth phase were collected and PCYR1 levels in cells
were determined by reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) and western blotting.
Extraction of BMSC-Exos
Human BMSCs (cat. no. 7500, ScienCell Research
Laboratories, Inc.) were cultured and subcultured according to the
manufacturer's instructions. After 5 subcultures, Exos in the BMSC
supernatant culture medium were extracted by ultracentrifugation.
BMSCs were cultured to 60% confluence, and the medium was replaced
with Exo-free serum (EXO-FBS-50A-1, SBI). When the cell density
reached 85%, the supernatant was centrifuged at 4°C at 2,000 g for
15 min. The collected supernatant was then centrifuged at 4°C at
16,500 g for 30 min, filtered with 0.22-µm filter membranes
to remove particles >200 nm, and centrifuged twice at 4°C at
100,000 g for 70 min, and the Exos in the lower layer were
collected, resuspended in 100 µl PBS, and stored at -80°C.
The supernatant of BMSCs treated with GW4869, an inhibitor of
exosome secretion (10 µM, cat. no. HY-19363,
MedChemExpress), for 24 h was used as the negative control (NC) of
Exos (28).
Preparation of Exo-si-PYCR1 using the
electroporation method
Cy3-si-PYCR1 (orange fluorescent label) was
synthesized by Shanghai GenePharma Co., Ltd. The extracted Exos and
Cy3-si-PYCR1 were added to the electroporation cuvettes (Bio-Rad
Laboratories, Inc., Gene Pulser Xcell electroporation system) at
the mass ratio of 5:1 (500 ng:100 ng) and were electroplated for 2
sec at 400 V and 125 µF. Subsequently, the mixture was
centrifuged at 135,000 g and 4°C for 70 min to obtain Exo-si-PYCR1,
resuspended in 100 µl PBS and stored at -80°C.
Identification of Exos and
Exo-si-PYCR1
The morphological structure of Exos and Exo-si-PYCR1
was observed using a transmission electron microscope (TEM) (FEI).
The particle size distribution and ζ potential of Exos and
Exo-si-PYCR1 were measured using a Malvern Laser Particle Sizer
(DLS, Zetasizer Nano ZS90, Malvern Instruments). The positive
expression of surface marker proteins CD9, CD81, CD63, and TSG101
(Abcam) in Exos and Exo-si-PYCR1 was detected by western blotting.
The content changes of free si-PYCR1 in Exos, free si-PYCR1, and
Exo-si-PYCR1 after electroporation were determined by agarose gel
electrophoresis to determine whether si-PYCR1 was loaded into
Exos.
Observation of Exo-si-PYCR1 uptake by BC
cells using laser confocal microscopy
T24 cells (2×105) were seeded in the
confocal Petri dishes and used for the control group, si-PYCR1
group, and Exo-si-PYCR1 group. The control cells were not treated,
cells in the si-PYCR1 group were transfected with free Cy3-si-PYCR1
(100 ng) using Lipofectamine® 2000 reagent (cat. no.
11668-019; Invitrogen; Thermo Fisher Scientific, Inc.), and cells
in the Exo-si-PYCR1 group were transfected with Exo-si-PYCR1 with
Cy3-si-PYCR1 (100 ng). After 24 h of transfection, cells were
incubated with immunostaining fixative (cat. no. P0098, Beyotime
Institute of Biotechnology) for 10 min and washed with
immunostaining detergent (cat. no. P0106, Beyotime Institute of
Biotechnology) for 5 min x 4 times. Then, to the confocal Petri
dishes, 200 µl green-fluorescent probe was added
(Actin-Tracker Green, cat. no. C1033, Beyotime Institute of
Biotechnology) diluted with immunofluorescence staining secondary
antibody diluent (1:200, cat. no. P0108, Beyotime Institute of
Biotechnology) and incubated at 37°C for 30 min. After staining,
the cells were washed with detergent for 5 min x 4 times, incubated
with DAPI working solution for 10 min, and washed with PBS 4 times;
the uptake rate (%) of Cy3-si-PYCR1 by T24 cells was observed under
a laser confocal microscope (magnification, ×200, Leica GmbH).
Cell grouping and treatment
T24 cells and RT4 cells were seeded in 6-well
plates, cultured until 80% confluence, and divided into the
following groups based on different treatments: i) blank group, T24
cells or RT4 cells normally cultured for 48 h; ii) si-PYCR1 group
and si-NC group, T24 cells transfected with the si-PYCR1 or the NC,
respectively; iii) oe-PYCR1 group and oe-NC group, RT4 cells
transfected with the oe-PYCR1 or the NC, respectively; iv) oe-PYCR1
+ Vehicle group and oe-PYCR1 + CL-387785 group, RT4 cells treated
with EGFR inhibitor CL-387785 (1 µM, cat. no. HY-10325,
MedChemExpress) or solvent (0.1% DMSO) for 2 h and transfected with
oe-PYCR1. The dosage and method of use of CL-387785 were determined
according prior to the study (data not shown); v) Exo group, T24
cells were cultured with Exos (100 ng/well) for 48 h; and vi)
Exo-si-NC group and Exo-si-PYCR1 group, T24 cells were cultured
with Exo-si-NC or Exo-si-PYCR1 (containing si-NC or si-PYCR1 100
ng/well) for 48 h. si-PYCR1, oe-PYCR1, and corresponding NCs used
in transfection were synthesized by Shanghai GenePharma CO., Ltd.
The sequences of si-RNAs were: si-PYCR1 forward, 5′-UGC UAU CAA CGC
UGU GG-3′ and reverse, 5′-CCA CAG CGU UGA UAG CA-3′; and si-NC
forward, 5′-AAU UCU CCG AAC GUG UAC GU-3′ reverse, 5′-ACG UAC ACG
UUC GGA GAA UU-3′. pcDNA3.1 was used as the vector of oe-PYCR1, and
oe-NC corresponded to the empty plasmid. After 48 h of
transfection, PYCR1 levels were determined by RT-qPCR and western
blotting.
MTT assay
Cell viability was detected using an MTT assay.
Briefly, 2,000 T24 or RT4 cells were seeded in 96-well plates.
After 48 h of culture, cells were incubated with 20 µl MTT
solution (0.5%, cat. no. M1025, Beijing Solarbio Science &
Technology Co., Ltd.) at 37°C for 4 h, and then MTT solution was
removed. Next, 100 µl DMSO was added to cells, and the
plates were shaken at a constant speed for 10 min to fully dissolve
the formazan crystals. The optical density value at 490 nm was
measured using a microplate reader (Thermo Fisher Scientific,
Inc.).
Transwell assays
After 48 h of transfection, the cells in each group
were collected and made into cell suspensions. The invasive ability
of BC cells was assessed using Transwell chambers (cat. no. 3413,
Corning, Inc.). Matrigel matrix adhesive was spread (60 µl,
Corning) on the bottom of the apical chamber, 100 µl cell
suspension was added to the chamber, with a cell count of
approximately 1×105, and 700 µl RPMI-1640 medium
comprising 10% fetal bovine serum was added to the basolateral
chamber. Following 48 h of incubation, the non-invasive cells in
the apical chamber of the Transwell chamber were wiped off with a
cotton swab, cells were fixed with 4% paraformaldehyde at 37°C for
30 min, and stained with 0.1% crystal violet at 37°C for 10 min.
The number of cells passing through the membrane pores was observed
under a microscope (200×, Olympus Corporation) in three random
fields for each well; the number of invasive cells in the different
groups was counted.
Wound-healing assay
The migratory ability of BC cells was assessed using
a wound-healing assay. After 48 h of transfection, T24 or RT4 cells
were seeded at 5×104 cells/well in 96-well plates and
cultured in an. Once a confluent monolayer had formed, a scratch
was created using a 200-µl sterile pipette gun head by
vertically marking a line under an inverted microscope (Olympus
Corporation) and imaged at 0 and 24 h. The cell migration rate (%)
was calculated by measuring the width of the wound.
Determination of glucose uptake, lactate,
and ATP production levels
The levels of aerobic glycolysis-related indicators,
glucose uptake, lactate production, and ATP production were
determined using the colorimetric method. The glucose uptake
colorimetric assay kit (cat. no. 36503, AAT Bioquest), lactate
detection kit (cat. no. KTB1100-1, Abbkine Scientific Co., Ltd.),
and ATP detection kit (cat. no. YT6278, Beijing YITA Biotechnology)
were used according to the instructions of the kits.
Co-immunoprecipitation (Co-IP)
T24 cells in the exponential phase were collected
and total protein was extract using weak RIPA lysate (P0013D,
Beyotime, Shanghai, China), and protein concentration was
determined using BCA reagent kit (P0012, Beyotime). The 200
µg of total protein was taken and 1 µg PYCR1 antibody
(cat. no. ab102601, Abcam) was added and incubated overnight at
4°C. Then, 10 µl pre-treated protein A (cat. no. YJ101,
Epizyme Biomedical Technology) agarose bead was added to the
solution and incubated for 2 h at 4°C to allow coupling of the
antibody with protein A agarose bead. Following
immunoprecipitation, the mixture was centrifuged for 3 min at 4°C
and 200 × g to sink the agarose bead to the bottom of the tube. The
supernatant was removed carefully, and the agarose bead was washed
3 times with 1 ml lysis buffer before examining the binding of
PYCR1 and EGFR protein using western blotting. IgG antibody (cat.
no. ab6715, Abcam) was used as a negative control, and protein
lysis buffer without immunoprecipitation was used as a positive
control (Input).
Xenograft tumor models
The 6-week-old male BALB/c nude mice (Cavens
Laboratory Animal) were used to establish the xenograft tumor
model. T24 cells have a high tumor formation rate in nude mice. T24
cells in the logarithmic growth phase were detached into a
single-cell suspension and subcutaneously injected into the armpits
of mice (2×106 cells per mouse). Drug intervention was
performed when the subcutaneous tumor grew to ~100 mm3
(tumor volume=long × width2 × 1/2). The mice with tumor
formations were randomly assigned to one of four groups (n=6 per
group): NC group, BMSC-Exo group, BMSC-Exo-si-NC group, and
BMSC-Exo-si-PYCR group, in which mice were administered 200
µl normal saline, BMSC-Exo, BMSC-Exo-si-NC, and
BMSC-Exo-si-PYCR1, respectively, by tail vein injection on days 0,
3, 6, and 9, respectively. The physical conditions and behavior of
mice were observed daily during the medication period. The weight
of mice was measured on days 0, 2, 4, 6, 8, and 12, respectively.
The volume of the xenografted tumor was measured once a week. The
tumor volume was measured for the last time in the 5th week (the
35th day) (maximum tumor long diameter: 15.0 mm). Then, mice were
euthanized by intraperitoneal injection of 1% pentobarbital sodium
(200 mg/kg). The tumors were removed for observation and imaging,
weight measurement, and immunohistochemical staining.
RT-qPCR
BC cells or tumor tissues of nude mice were
collected. Total RNA was extracted using the TRIzol®
reagent (cat. no. 15596018; Invitrogen; Thermo Fisher Scientific,
Inc.) and reverse transcribed into cDNA using the RNA RT kit
according to the manufacturer's protocol (cat. no. 4387406;
Invitrogen; Thermo Fisher Scientific, Inc.). qPCR was performed
using the ABI Prism 7300 system using the following amplification
temperature protocol: Pre-denaturation, 95°C for 30 sec; followed
by 40 cycles of denaturation at 95°C for 5 sec, annealing at 60°C
for 30 sec, and extension at 72°C for 30 sec. The relative gene
expression was analyzed using the 2−ΔΔCq method
(29), with β-Actin as the
internal reference control. The primers were synthesized by Sangon
Biotech Co., Ltd. The sequences were as follows: PYCR1:F: 5′-GGC
TGC CCA CAA GAT AAT GGC-3′; R: 5′-CAA TGG AGC TGA TGG TGA CGC-3′
and β-actin: F: 5′-AAT GAG CTG CGT GTG GCT-3′; R: 5′-TAG CAC AGC
CTG GAT AGC AA-3′.
Western blotting
Cells were collected and lysed using RIPA lysis
buffer (cat. no. P0013B, Beyotime Institute of Biotechnology) to
obtain total proteins from cells or tumor tissues of nude mice.
Protein concentration was determined using a BCA kit. Equal amounts
of proteins protein were loaded on SDS-gels, resolved using 10%
SDS-PAGE, and transferred to PVDF membranes. Membranes were blocked
with 5% skimmed milk for 1 h and incubated overnight with the
following antibodies: PYCR1 (1:1,000, cat. no. ab102601; Abcam),
GLUT1 (1 µg/ml, cat. no. ab115730, Abcam), hexokinase 1
(HK1; 1:1,000, cat. no. ab154839, Abcam), lactic dehydrogenase A
(LDHA; 1:5.000, ab52488, Abcam), EGFR (1:10.000, ab52894, Abcam),
p-EGFR (1:1,000, ab40815, Abcam), PI3K (1:1,000, ab32089, Abcam),
p-PI3K (0.5 µg/ml, ab278545, Abcam), AKT (1:500, ab8805,
Abcam), p-AKT (1:1,000, ab38449, Abcam), and β-Actin (1:1,000,
ab8227, Abcam). After incubation with the primary antibodies, the
membranes were probed with horseradish peroxidaseconjugated
secondary antibody (1:3,000, ab6721, Abcam) for 1 h. Finally,
Signals were visualized using the ChemiDoc MP Imaging System
(Bio-Rad Laboratories, Inc.).
Immunohistochemical staining
Tumor tissues were fixed with 4% paraformaldehyde at
4°C for 24 h and cut into 5-µm thick paraffin-embedded
sections. As previously reported (30), the sections were routinely dewaxed
and hydrated, and subjected to antigen retrieval using sodium
citrate buffer (pH=6.0) in a microwave oven at 100°C for 10 min and
incubated with 3% H2O2 at room temperature
for 10 min to block the activity of endogenous peroxidase. After
washing with PBS, the sections were incubated with the primary
antibody against Ki-67 (1:250, cat. no. ab92742, Abcam) in the dark
at 4°C overnight. After PBS washes, the sections were incubated at
room temperature with the secondary antibody goat anti-rabbit IgG
H&L HRP (1:500, cat. no. ab97051, Abcam) for 30 min at 37°C,
washed with PBS, counterstained with hematoxylin for 30 sec at
37°C, differentiated with 0.1% HCl, rinsed with tap water to return
to blue for 5 min. Subsequently, the sections were dehydrated with
a gradient of alcohol solutions, cleared with xylene, and sealed
with neutral gum. After air drying, the sections were observed and
imaged under a light microscope (×200, Olympus Corporation).
Biosafety investigation
The safety of Exo-si-PYCR1 was investigated by
observing the weight changes of mice in each group during the drug
intervention period (days 0-12). After mice were euthanized by
intraperitoneal injection of 1% pentobarbital sodium (200 mg/kg),
the major organs (heart, liver, spleen, lung, and kidney) of mice
were collected for pathological HE staining to further investigate
the effects of Exo-si-PYCR1 on these organs.
Statistical analysis
GraphPad Prism version 8.01 (GraphPad Software Inc.)
was used to perform the statistical analysis. A Shapiro-Wilk test
was used to evaluate the distribution of data. In vitro
experiments were performed three times in each group, and 6 nude
mice were included in each group for the in vivo
experiments. The normally distributed data are presented as the
mean ± SD. An independent samples Student's t-test was used for
comparisons between 2 groups and a one-way ANOVA followed by
Tukey's post hoc test was used to compare differences between
multiple groups. P<0.05 was considered to indicate a
statistically significant difference. a statistically significant
difference.
Results
PYCR1 expression is upregulated in BC
cells and it promotes cell growth
PYCR1 expression is upregulated in various types of
cancer and it is involved in the regulation of BC cell behaviors
(31,32). Using the StarBase database, it was
found that PYCR1 levels were upregulated in BC (Fig. 1A, P<0.05). Next, PYCR1 levels in
SV-HUC-1 normal human bladder epithelial immortalized cells and
four BC cells (T24, 5637, RT4, and TCCSUP) were determined by
RT-qPCR and western blot assay. PYCR1 was found to be upregulated
in the four BC cell lines, in which T24 cells exhibited the highest
level and RT4 cells exhibited the lowest levels (Fig. 1B and C, all P<0.05). To
investigate the effects of PYCR1 on BC cells, interference plasmids
were transfected into T24 cells to knock down PYCR1, and
overexpression plasmids were transfected into RT4 cells to increase
PYCR1 levels. RT-qPCR and western blot assays showed that PYCR1 was
successfully knocked down in T24 cells and overexpressed in RT4
cells (Fig. 1D and E, all
P<0.001). MTT assay showed that T24 cell viability was decreased
following PYCR1 knockdown and increased following PYCR1
overexpression (Fig. 1F, all
P<0.001). Transwell wound-healing assays showed that PYCR1
knockdown reduced T24 cell invasion and migration, whereas PYCR1
overexpression stimulated RT4 cell invasion and migration (Fig. 1G and H, all P<0.001). These
results confirmed that PYCR1 was highly expressed in BC where it
facilitated BC cell proliferation, invasion, and migration.
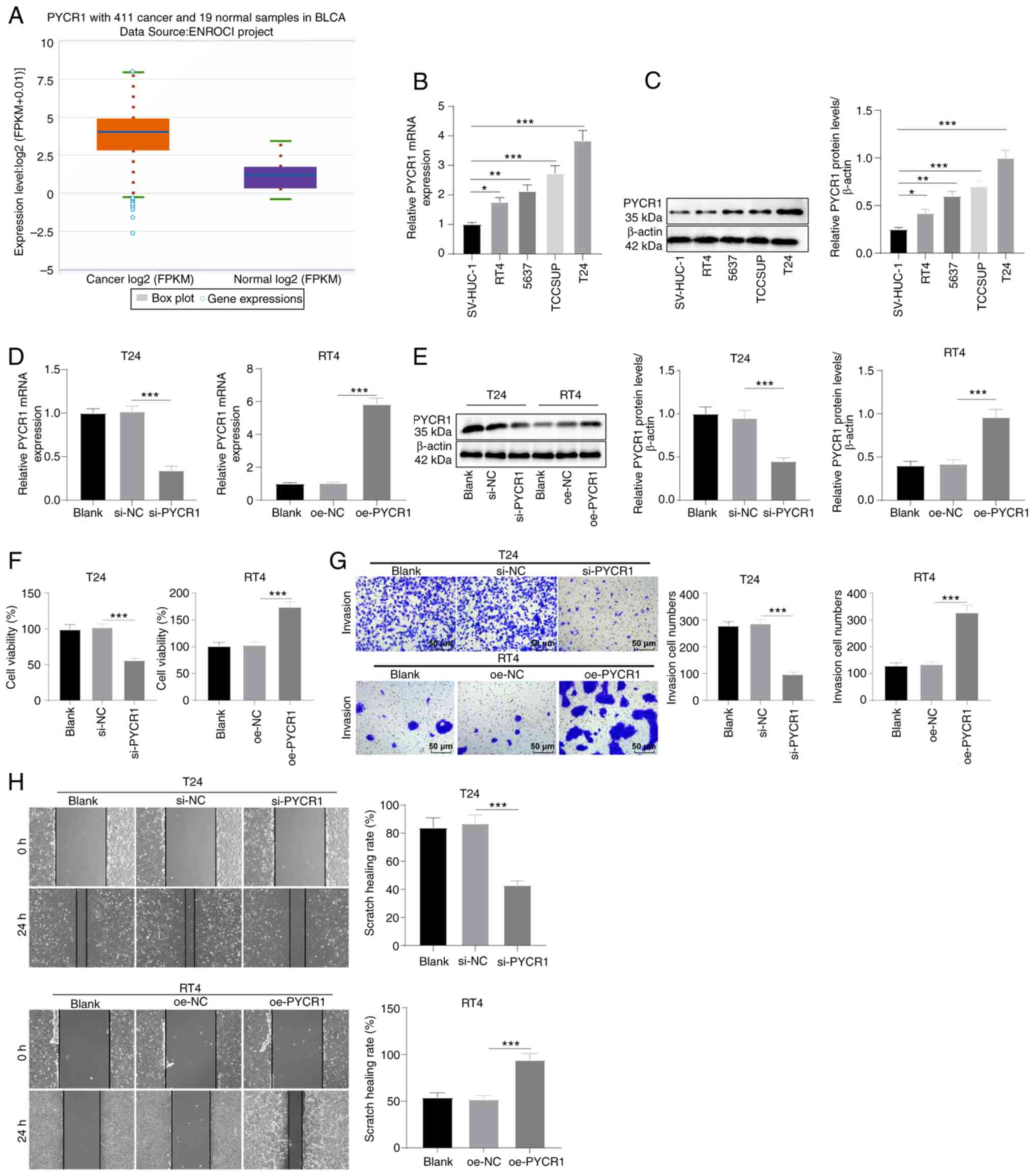 | Figure 1PYCR1 expression is upregulated in BC
cells and it promotes cell proliferation, invasion, and migration.
(A) Elevated expression of PYCR1 in BC was found by StarBase. (B
and C) RT-qPCR and western blot elicited raised mRNA and protein
expression levels of PYCR1 in BC cells RT4, 5637, TCCSUP, and T24
relative to normal bladder epithelial cells SV-HUC-1. (D and E)
RT-qPCR and western blot manifested repressed mRNA and protein
expression levels of PYCR1 in T24 cells after PYCR1 knockout and
facilitated levels in RT4 cells after PYCR1 overexpression. (F) MTT
assay detected decreased T24 viability and increased RT4 viability.
(G) Transwell assay detected reduced T24 invasion and promoted RT4
invasion. (H) Wound-healing assay detected repressed T24 migration
and enhanced RT4 migration. Data are presented as the mean ± SD of
three repeats. A one-way ANOVA followed by a Tukey's test was used
to compare the data. *P<0.05, **P<0.01,
***P<0.001. BC, bladder cancer; PYCR1,
pyrroline-5-carboxylate reductase 1; FPKM, Fragments Per Kilobase
of exon model per Million mapped fragments; oe, overexpression. |
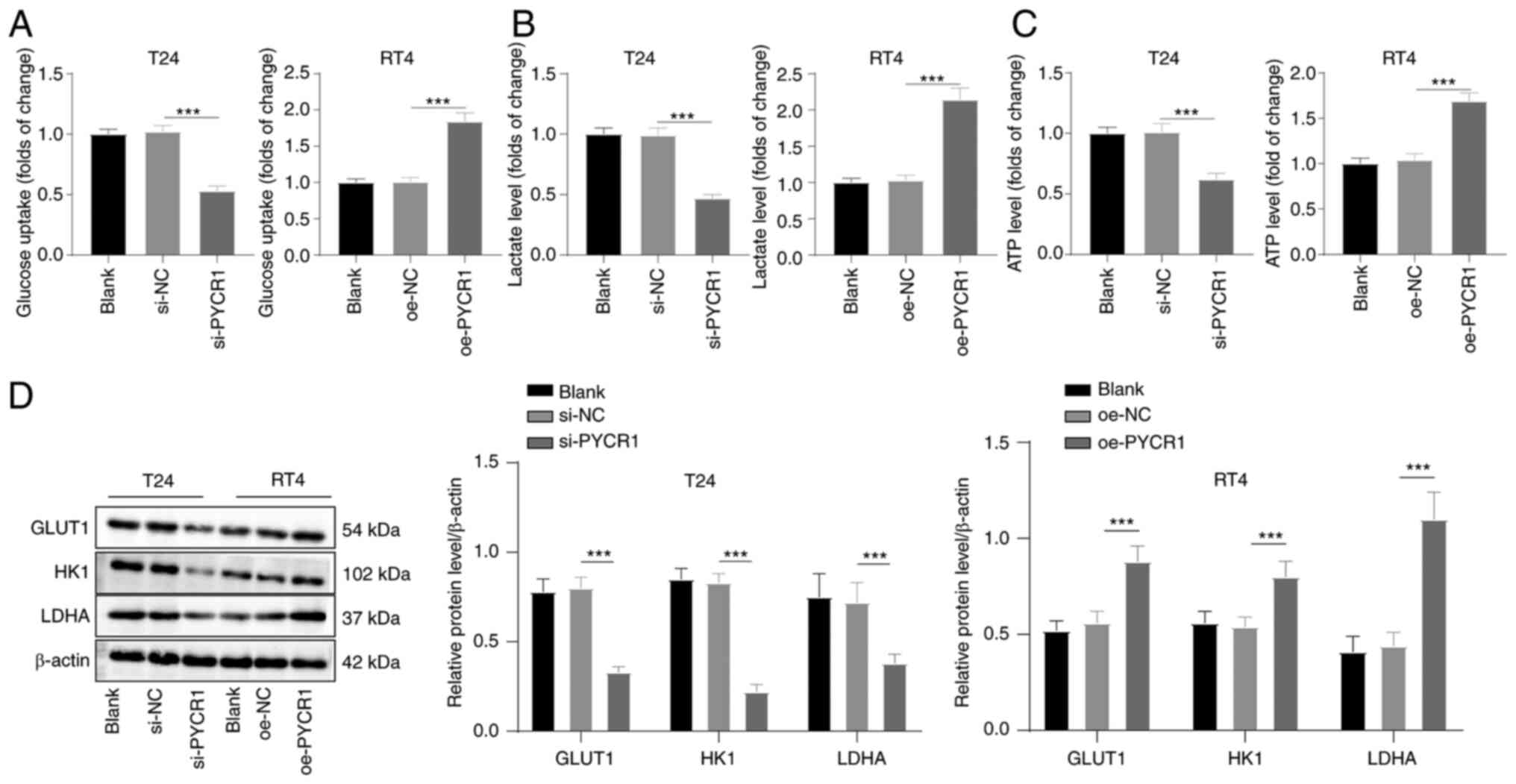
PYCR1 promotes aerobic glycolysis in BC
cells
Aerobic glycolysis is important in regulating tumor
cell behaviors (33,34). It was speculated that PYCR1 may
regulate aerobic glycolysis in BC. The levels of glucose uptake,
lactate production, and ATP production in cells after 48 h of
transfection were assessed. The levels of glucose uptake, lactate
production, and ATP production in T24 cells with PYCR1 expression
knocked down were lower, whereas they were increased in RT4 cells
overexpressing PYCR1 (Fig. 2A-C,
all P<0.001). Furthermore, the protein expression levels of
aerobic glycolysis-related enzymes GLUT1, HK1, and LDHA were
assessed by western blotting. PYCR1 knockdown reduced the protein
expression levels of GLUT1, HK1, and LDHA, while PYCR1
overexpression increased the expression levels of these proteins
(Fig. 2D, all P<0.001). These
results suggested that PYCR1 promoted aerobic glycolysis in BC.
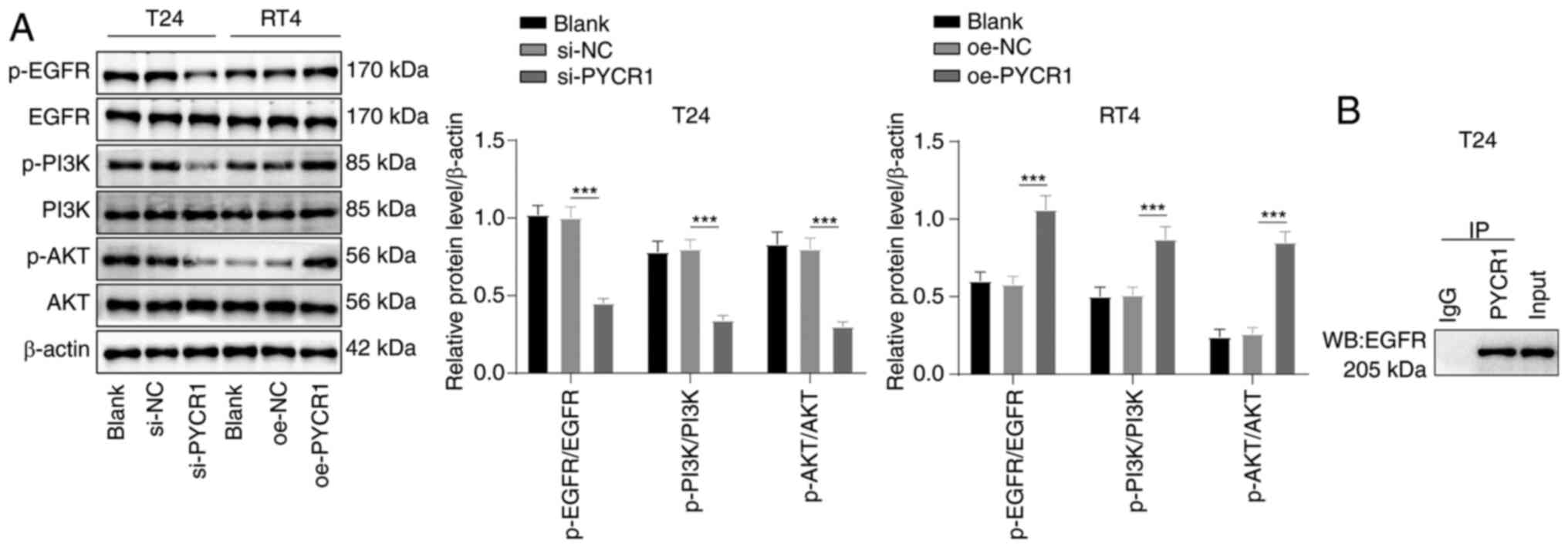
PYCR1 promotes activation of the
EGFR/PI3K/AKT pathway in BC cells
The EGFR/PI3K/AKT pathway is closely associated with
BC cell proliferation, invasion, and metastasis (35) and is implicated in the regulation
of aerobic glycolysis (17,18).
It was speculated that PYCR1 regulated the EGFR/PI3K/AKT pathway.
Western blot assay demonstrated that the EGFR/PI3K/AKT pathway was
inhibited, and the p-EGFR/EGFR, p-PI3K/PI3K, and p-AKT/AKT levels
were repressed in T24 cells following PYCR1 knockdown, whereas this
pathway was activated in RT4 cells following PYCR1 overexpression
(Fig. 3A, all P<0.001). Thus,
PYCR1 activated the EGFR/PI3K/AKT pathway. To probe the mechanism
by which PYCR1 regulated the EGFR/PI3K/AKT pathway, the
protein-protein interactions between PYCR1 and EGFR were examined
by Co-IP, which revealed that endogenous PYCR1 interacted with EGFR
protein in T24 cells, suggesting that PYCR1 could interact with
EGFR (Fig. 3B); that is, PYCR1
activated the PI3K/AKT pathway by binding to EGFR.
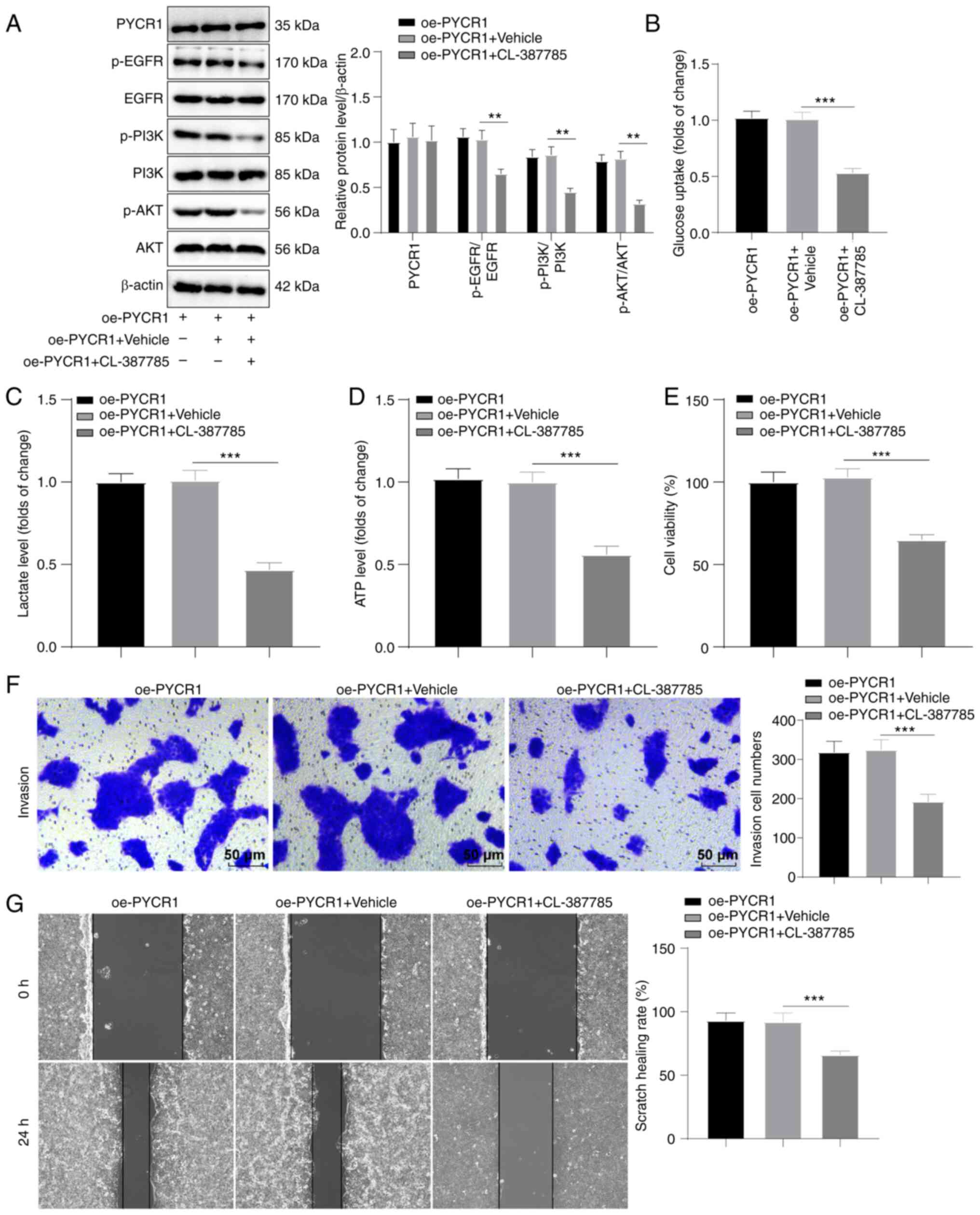
EGFR pathway inhibitor abrogates the
effects of PYCR1 on aerobic glycolysis and BC cell proliferation,
invasion, and migration
To confirm that PYCR1 regulated aerobic glycolysis
and BC cell growth through the EGFR/PI3K/AKT pathway, RT4 cells
overexpressing PYCR1 were treated with the pathway inhibitor
CL-387785. Western blotting showed that the EGFR/PI3K/AKT pathway
was inhibited by CL-387785, while PYCR1 protein expression was not
altered by CL-387785 (Fig. 4A, all
P<0.01). This result confirmed that EGFR was downstream of
PYCR1. Glucose uptake, lactate production, and ATP production
levels were also repressed by CL-387785 treatment (Fig. 4B-D, all P<0.001), and RT4 cell
proliferation, invasion, and migration were also suppressed
(Fig. 4E-G; all P<0.001).
Overall, CL-387785 abrogated the promoting effects of PYCR1
overexpression on aerobic glycolysis and BC cell proliferation,
invasion, and migration.
Identification of BMSC-derived Exos and
loading of si-PYCR1
MSC-shuttled Exos have a good biological affinity,
low toxicity, and low immunogenicity and can be used as carriers to
load si-RNA and other drugs to target cancer (20). BMSC-secreted Exos were isolated by
differential centrifugation and loaded with si-PYCR1 by
electroporation. TEM revealed that Exos were present, visible as
the circular saccular body with a double-layer membrane structure
and loading si-PYCR1 had no obvious effect on their morphology
(Fig. 5A). The particle diameter
of the Exos was 54.8±9.6 nm, and the particle size of Exo-si-PYCR1
after loading si-PYCR1 was not changed significantly (Fig. 5B). The potential of Exos was
−8.67±0.21 mV, while that of Exo-si-PYCR1 was −14.32±0.18 mV, which
indicated that the negatively charged si-PYCR1 was loaded into Exos
(Fig. 5C, P<0.01). The Exo
surface markers CD9, CD81, CD63, and TSG101 in Exos and
Exo-si-PYCR1 were positively expressed (Fig. 5D). Agarose gel electrophoresis
found that the levels of free si-PYCR1 in Exo-si-PYCR1 were lower,
which indicated that si-PYCR1 was loaded into Exos (Fig. 5E). These results indicated that
BMSC-shuttled Exos loaded with si-PYCR1 had been successfully
obtained.
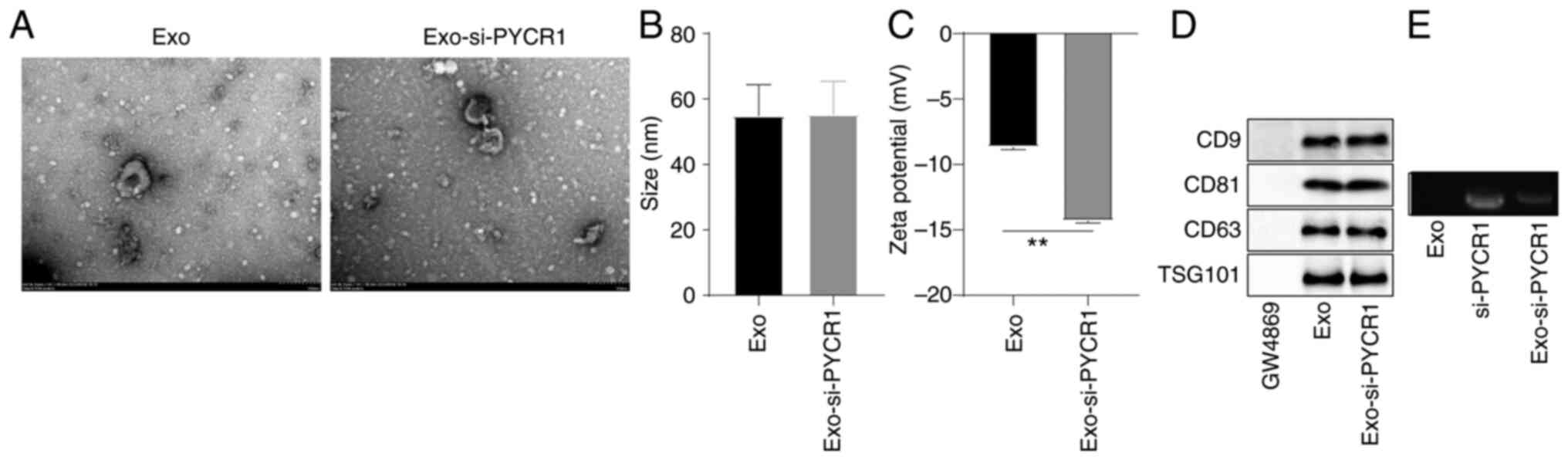 | Figure 5Identification of BMSC-derived Exos
and loading of si-PYCR1. (A) The morphological structure of Exos
and Exo-si-PYCR1 were double-layered membrane vesicle structure
observed using a TEM. (B and C) Particle diameter and potential of
Exo and Exo-si-PYCR1 were determined using the particle size
analyzer and potential analyzer. (D) The protein expression levels
of Exo surface markers CD9, CD63, CD81 and TSG101 were positive, as
determined by western blot. GW4869 is the negative control of Exos.
(E) The levels of free si-PYCR1 in Exos, si-PYCR1, and Exo-si-PYCR1
were measured by agarose gel electrophoresis (Exo-si-PYCR1 was
decreased relative to si-PYCR1). Data are presented as the mean ±
SD of three repeats. A one-way ANOVA followed by a Tukey's test was
used to compare the data. **P<0.01, BMSC, bone
marrow-derived stem cell; BC, bladder cancer; PYCR1,
pyrroline-5-carboxylate reductase 1; Exo, exosome; si, small
interfering; TEM, transmission electron microscopy. |
Exos-si-PYCR1 have stronger inhibitory
effects on aerobic glycolysis and proliferation, invasion and
migration of BC cells
To evaluate the effects of Exo-si-PYCR1 on BC cells,
the proportion of Cy3-labeled Exo-si-PYCR1 entering T24 cells was
observed using a laser confocal microscope. The proportion of
si-PYCR1 plasmid introduced into T24 cells increased relative to
the control group, and the levels of Exo-si-PYCR1 entering the
cells was higher in the si-PYCR1 group (Fig. 6A, all P<0.05). Exo-si-PYCR1
knocked down PYCR1 levels in T24 cells (Fig. 6B and C, all P<0.01). The glucose
uptake, lactate production, and ATP production levels were
suppressed after Exo-si-PYCR1 treatment, and the inhibitory effect
of Exo-si-PYCR1 was more prominent than that of si-PYCR1
transfection (Fig. 6D and F, all
P<0.05). In addition, Exo-si-PYCR1 inhibited T24 cell
proliferation, invasion, and migration, and the inhibitory effect
was stronger than that of si-PYCR1 (Fig. 6G-I, all P<0.05). Collectively,
Exo-si-PYCR1 inhibited aerobic glycolysis and BC cell growth, and
the inhibitory effect of Exo-si-PYCR1 was stronger than that of
si-PYCR1.
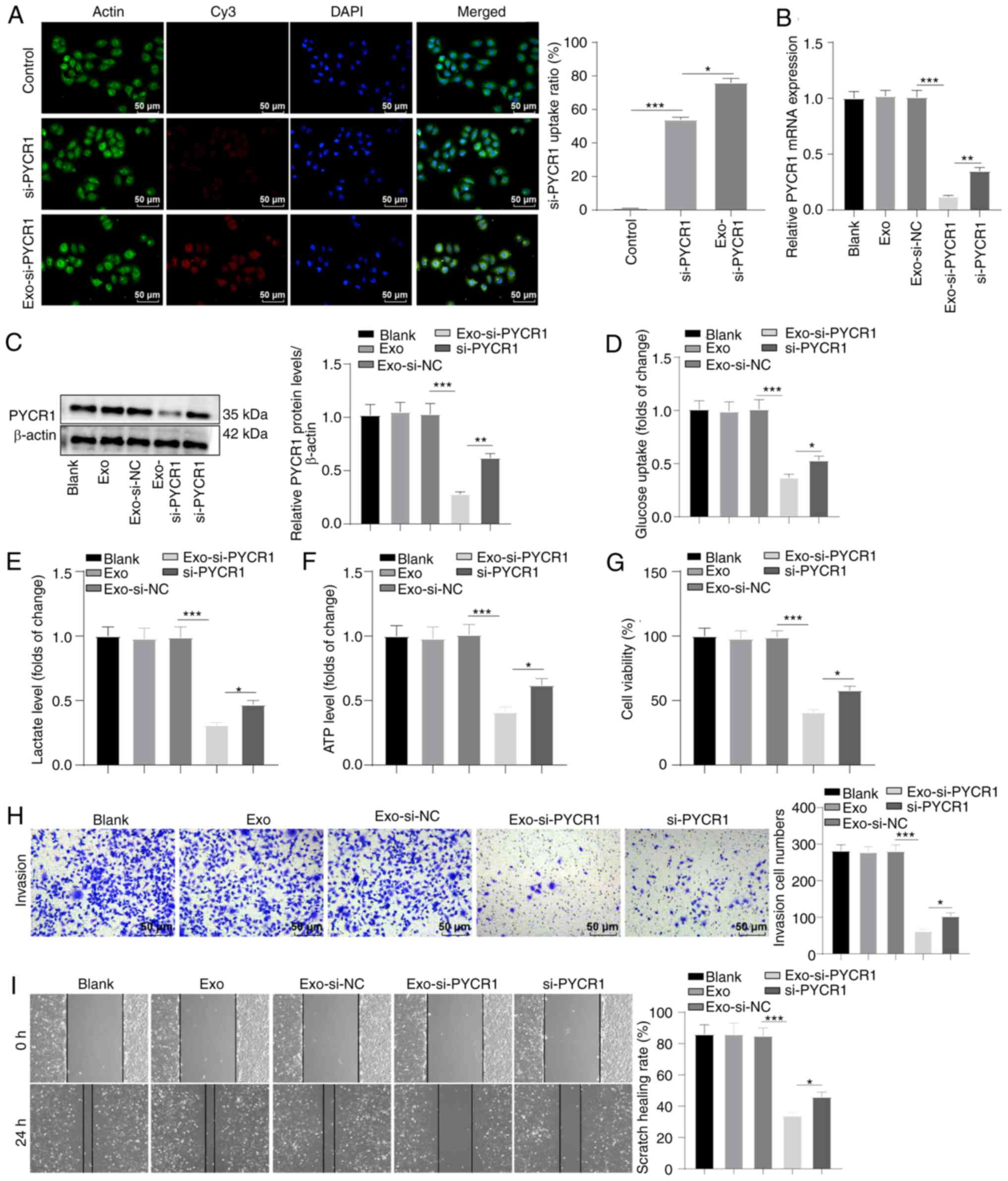 | Figure 6Exo-si-PYCR1 has a stronger
inhibitory effect on aerobic glycolysis and BC cell proliferation,
invasion, and migration. (A) The uptake of Cy3-labeled si-PYCR1 by
T24 cells was observed using laser confocal microscopy. (B and C)
RT-qPCR and western blot revealed reduced mRNA and protein
expression levels of PYCR1 in Exo-si-PYCR1-treated T24 cells. (D-F)
Colorimetry and reagent kit demonstrated diminished levels of
glucose uptake, lactate production, and ATP production in
Exo-si-PYCR1-treated T24 cells. (G) MTT assay detected blocked
proliferation in Exo-si-PYCR1-treated T24 cells. (H) Transwell
assays detected weakened invasion in Exo-si-PYCR1-treated T24
cells. (I) Wound-healing assay detected suppressed migration in
Exo-si-PYCR1-treated T24 cells. Data are presented as the mean ± SD
of three repeats. A one-way ANOVA followed by a Tukey's test was
used to compare the data. *P<0.05,
**P<0.01, ***P<0.001. BC, bladder
cancer; PYCR1, pyrroline-5-carboxylate reductase 1; Exo, exosome;
si, small interfering. |
Exo-si-PYCR1 inhibits growth of BC
xenograft tumors in nude mice
To validate the inhibitory effects of Exo-si-PYCR1
on BC in vivo, T24 cells were injected into nude mice to
establish xenograft tumor models of BC. When the volume of the
xenograft tumors was ~100 mm3, BMSC-Exos,
BMSC-Exo-si-NC, and BMSC-Exo-si-PYCR1 were injected into nude mice
through the tail vein. After BMSC-Exo-si-PYCR1 treatment, the PYCR1
levels in tumor tissues were repressed (Fig. 7A and B, all P<0.001), and
BMSC-Exo-si-PYCR1 inhibited the growth rate, volume, and weight of
xenograft tumors in nude mice (Fig.
7C-E, P<0.001), and the number of Ki67-positive cells that
marked the proliferation of cancer cells in tumor tissues was
reduced (Fig. 7F, P< 0.001),
indicating that BMSC-Exo-si-PYCR1 inhibited BC cell proliferation
in vivo. After the injection of BMSC-Exo-si-PYCR1, any toxic
effects of BMSC-Exo, BMSC-Exo-si-NC, and BMSC-Exo-si-PYCR1 were
evaluated by observing the changes in the body weight of nude mice
and observing any pathological changes in the primary organs
(heart, liver, spleen, lung, and kidney) by HE staining. BMSC-Exo,
BMSC-Exo-si-NC, and BMSC-Exo-si-PYCR1 had no notable effects on the
body weight and the primary organs of nude mice (Fig. 7G-H). These results suggested that
the use of BMSC-Exo-si-PYCR1 may be a feasible method for the
management of BC.
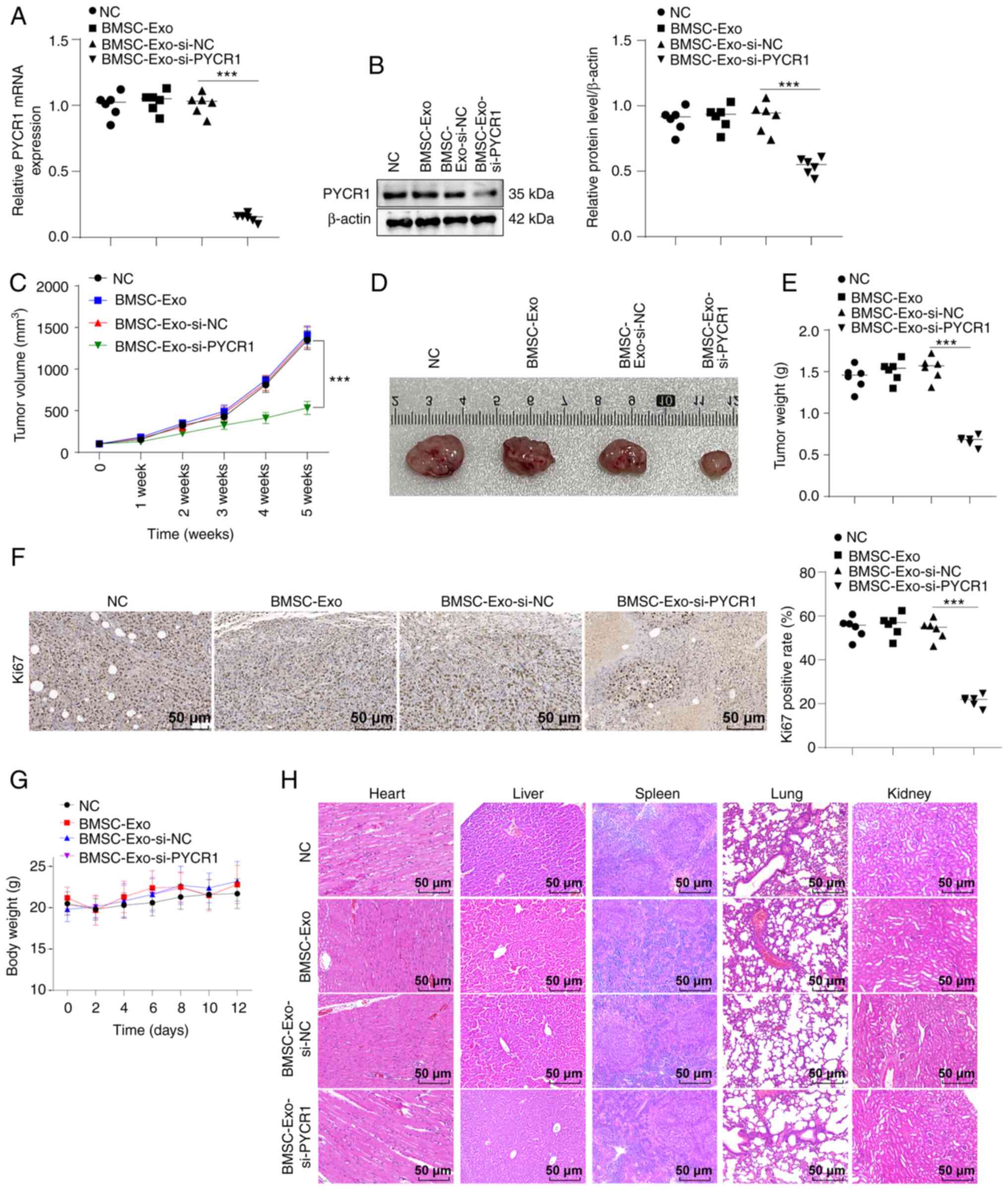 | Figure 7Exo-si-PYCR1 prevents BC xenograft
tumor growth in nude mice and exhibits good biological safety. T24
cells were subcutaneously injected into nude mice to establish a
tumor model of BC. When the tumor volume grew to ~100
mm3, BMSC-Exo, BMSC-Exo-si-NC, or BMSC-Exo-si-PYCR1 was
injected into mice through the tail vein. The NC group was injected
with an equivalent volume of normal saline. (A and B) RT-qPCR and
western blot demonstrated decreased mRNA and protein expression
levels of PYCR1 in tumor tissues of the BMSC-Exo-si-PYCR1 group.
(C) The volume of the xenograft tumor in nude mice of the
BMSC-Exo-si-PYCR1was decreased. (D) The size of the xenograft
tumor. (E) The weight of the xenograft tumor in the
BMSC-Exo-si-PYCR1 group was reduced. (F) The percentage of
Ki67-positive cells in tumor tissues was determined by
immunohistochemistry. (G) The body weight of nude mice 12 days
after administration maintained stability. (H) There were no
obvious changes in cell morphology of the heart, liver, spleen,
lung, and kidney of nude mice analyzed by HE staining. n=6
mice/group. Data are presented as the mean ± SD of three repeats. A
Student's t-test or a one-way ANOVA followed by a Tukey's test was
used to compare the data. ***P<0.001. BC, bladder
cancer; PYCR1, pyrroline-5-carboxylate reductase 1; Exo, exosome;
si, small interfering; BMSC, bone marrow-derived stem cell; Exo,
exosome; NC, negative control; HE, hematoxylin and eosin. |
Discussion
BC is the 4th most prevalent malignancy in men and
also a prevalent malignancy in women; it ranges in severity from
non-invasive and unaggressive tumors that recur in patients, to
long-term invasive surveillance, to invasive and aggressive tumors
with high disease-specific mortality (2). Evidence has revealed that
upregulation of PYCR1 promotes BC development and progression
(36). Exos are important in
cell-cell communication that are natural carriers of biological
cargo and have become a promising platform for cancer treatment
(37). The present study found
that si-PYCR1 loaded into BMSC-derived Exos blocked aerobic
glycolysis and BC growth via regulation of the EGFR/PI3K/AKT
pathway.
The upregulation of PYCR1 contributed to the
progression of various cancers such as gastric cancer,
nasopharyngeal cancer, and lung cancer (19,38,39).
In the present study, it was found that PYCR1 expression was
upregulated in BC cells. In particular, the relative expression
levels of PYCR1 were highest in T24 cells and lowest in RT4 cells.
Therefore, PYCR1 expression was knocked down in T24 cells and
overexpressed in RT4 cells to investigate its effects on BC. T24
cells with PYCR1 expression knocked down exhibited reduced
viability, invasion, and migration, whereas RT4 cells
overexpressing PYCR1 exhibited increased viability, invasion, and
migration. Consistently, PYCR1 expression was elevated in BC
tissues, and PYCR1 knockdown in vivo models resulted in
reduced cell growth, whereas PYCR1 overexpression accelerated cell
proliferation and invasion (7). In
conclusion, high expression of PYCR1 facilitated the proliferation,
invasion, and migration of BC cells.
Aerobic glycolysis participates in the regulation of
the proliferation, invasion, and migration of various tumor cells
including breast cancer, pancreatic cancer, and HCC (9,40,41).
Cancer cells must generate sufficient ATP to satisfy the
requirements of cell proliferation, and they uptake large
quantities of glucose producing high volumes of lactate (42). High glycolytic flux depends on the
upregulation of the glycolysis-associated genes (GLUT1, LDHA, and
HK1), causing excessive production of lactate (43). Accordingly, it was speculated that
PYCR1 might regulate aerobic glycolysis in BC and subsequent
examination evidenced that glucose uptake, lactate production, ATP
production, and GLUT1, HK1, and LDHA protein levels were suppressed
in T24 cells following PYCR1 knockdown, while their levels were
increased in RT4 cells following PYCR1 overexpression.
PYCR1-dependent proline biosynthesis is critical for tumorigenesis
by promoting cell proliferation and linking the proline cycle to
glycolysis (44). PYCR1 knockdown
induced metabolic transition from glycolysis to oxidative
phosphorylation (45). However,
there are no reports on the effects of PYCR1 in aerobic glycolysis
in BC, to the best of our knowledge. Thus, the present study was
the first to show that PYCR1 facilitated aerobic glycolysis in
BC.
Downregulation of ring finger protein 126 inhibited
the metastasis and proliferation of BC cells through the
EGFR/PI3K/AKT pathway (35). The
results showed that the EGFR/PI3K/AKT pathway was inhibited and
p-EGFR/EGFR, p-PI3K/PI3K, and p-AKT/AKT levels were repressed in
T24 cells following PYCR1 knockdown, while the EGFR/PI3K/AKT
pathway was activated in RT4 cells overexpressing PYCR1. Previous
studies have shown that the EGFR/PI3K/AKT pathway facilitated
cancer cell proliferation and suppressed cell apoptosis in various
types of cancer (16,46). PYCR1 has been implicated in the
regulation of the PI3K/AKT/mTOR axis (19). The results of the present study
showed that PYCR1 could interact with EGFR. To further verify the
involvement of the EGFR/PI3K/AKT pathway in aerobic glycolysis and
BC growth, RT4 cells overexpressing PYCR1 were treated with the
pathway inhibitor CL-387785, and the results showed that CL-387785
inhibited glucose uptake, lactate production, ATP production, and
the proliferation, invasion, and migration of RT4 cells, whilst
PYCR1 protein expression remained unchanged. Consistently, the
PI3K/mTOR pathway inhibitor repressed glycolysis in EGFR-mutant
lung adenocarcinoma cells (47).
The EGF-like motif is indispensable for VersicanV1 to facilitate
aerobic glycolysis in HCC cells and the invasion and metastasis
through the activation of the EGFR/PI3K/AKT axis (18). To conclude, CL-387785 abrogated the
effects of PYCR1 overexpression on stimulating aerobic glycolysis
and BC cell growth.
Owing to their nanometer sizes, Exos have been
identified as promising drug delivery tools for the management of
cancer (26). si-PYCR1-loaded
BMSC-Exos were constructed, and it was found that the levels of
Exo-si-PYCR1 entering the cells were elevated. Exos have good
biocompatibility and low immunogenicity and can easily be taken up
by cells (20,26). The results showed that the number
of fluorescent si-PYCR1 plasmids introduced into T24 cells was
larger, and thus the inhibitory rate of Exo-si-PYCR1 on PYCR1
expression was higher and the inhibitory effect on aerobic
glycolysis and malignant behaviors of T24 cells was more prominent.
Exo-si-PYCR1 exerted more prominent effects on suppressing PYCR1
levels, inhibiting glucose uptake, lactate production, and ATP
production, and preventing the proliferation, invasion and
migration of T24 cells than si-PYCR1, illustrating for the first
time that Exo-si-PYCR1 suppressed aerobic glycolysis and BC cell
growth, and the inhibitory effect of Exo-si-PYCR1 was stronger than
that of si-PYCR1. The in vivo results showed that
BMSC-Exo-si-PYCR1 repressed xenograft tumor growth in nude mice and
decreased BC cell proliferation. In addition, BMSC-Exo-si-PYCR1 had
no obvious effect on the primary organs and body weight of nude
mice. In conclusion, loading Exos with si-PYCR1 to knock down PYCR1
in cancerous cells specifically may serve as a feasible method to
treat BC.
In summary, this study showed for the first time
that PYCR1 regulated aerobic glycolysis and BC cell growth via the
EGFR/PI3K/AKT pathway. Furthermore, Exo-si-PYCR1 was constructed, a
nano nucleic acid drug with BMSC-Exos as its carrier and verified
that they inhibited BC progression through in vivo and in
vitro experiments, serving as a reference for the development
of Exo-targeted therapeutic drugs for BC and other types of cancer.
However, there was no further in-depth study on the binding sites
and intermolecular interactions between PYCR1 and EGFR, or the
regulatory effects of PYCR1 on the EGFR/PI3K/AKT pathway in animal
experiments. In addition, the therapeutic efficacy and biosafety of
Exo-si-PYCR1 need to be verified through larger animal experiments.
The regulatory role of PYCR1 in regulating the EGRF/PI3K/AKT
pathway and glycolysis in BC cells was confirmed using both
knockdown and overexpression experiments with RT4 and T24 cells. In
the preliminary experiments, PYCR1 was overexpressed in RT4 cells
and knocked down in T24 cells, and the results showed that PYCR1
overexpression could activate the EGFR/PI3K/AKT pathway. To
validate whether PYCR1 regulates aerobic glycolysis and the growth
of bladder cancer cells through the EGFR/PI3K/AKT pathway, the EGFR
inhibitor CL-387785 was used to treat RT4 cells overexpressing
PYCR1 in rescue experiments. Therefore, RT4 cells were selected for
the functional experiments. It is hypothesized that the difference
between the two cell types will have little impact on the present
experimental results. Moreover, the overall experimental design was
relatively complete. In the future, these limitations will be
addressed. Furthermore, the molecular mechanism of the regulatory
effects of PYCR1/EGFR/PI3K/AKT using animal experiments will be
assessed to further verify the therapeutic effect and biosafety of
Exo-si-PYCR1 and develop and construct additional nanonucleic acid
drugs based on tumor therapeutic drugs and key factors associated
with targeted inhibition of tumor progression through siRNAs to
efficiently target tumors.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
WS is the guarantor of integrity of the entire
study. ZL and WS conceived the study. YWL designed the study and
was involved in data acquisition. YJ performed the literature
research. JL contributed to the manuscript preparation. WS and ZL
wrote and edited the manuscript. HFF and YJ performed the
experiments. HFF, JL and QY performed the data analysis. ZL and YWL
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
All procedures were authorized by the Academic
Ethics Committee of Hunan Provincial People's Hospital, The First
Affiliated Hospital of Hunan Normal University (approval no.
2021-071) and were strictly implemented in accordance with the
National Laboratory Animal Guide. Laboratory procedures were
performed in such a manner as to reduce the pain and discomfort
caused to mice.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
This work was supported by funding from the Hunan Natural
Science Foundation (grant no. 2021JJ70092); Hunan Provincial Health
Commission Project (No. 20200044); the Project of Hunan Provincial
Department of Education (No.20C1174).
References
|
1
|
Dobruch J and Oszczudlowski M: Bladder
cancer: Current challenges and future directions. Medicina
(Kaunas). 57:7492021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lenis AT, Lec PM, Chamie K and Mshs MD:
Bladder cancer: A review. JAMA. 324:1980–1991. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siracusano S, Rizzetto R and Porcaro AB:
Bladder cancer genomics. Urologia. 87:49–56. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Na L, Wang Z, Bai Y, Sun Y, Dong D, Wang W
and Zhao C: WNT7B represses epithelial-mesenchymal transition and
stem-like properties in bladder urothelial carcinoma. Biochim
Biophys Acta Mol Basis Dis. 1868:1662712022. View Article : Google Scholar
|
|
5
|
Hollinshead KER, Munford H, Eales KL,
Bardella C, Li C, Escribano-Gonzalez C, Thakker A, Nonnenmacher Y,
Kluckova K, Jeeves M, et al: Oncogenic IDH1 mutations promote
enhanced proline synthesis through PYCR1 to support the maintenance
of mitochondrial redox homeostasis. Cell Rep. 22:3107–3114. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Westbrook RL, Bridges E, Roberts J,
Escribano-Gonzalez C, Eales KL, Vettore LA, Walker PD,
Vera-Siguenza E, Rana H, Cuozzo F, et al: Proline synthesis through
PYCR1 is required to support cancer cell proliferation and survival
in oxygen-limiting conditions. Cell Rep. 38:1103202022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du S, Sui Y, Ren W, Zhou J and Du C: PYCR1
promotes bladder cancer by affecting the Akt/Wnt/beta-catenin
signaling. J Bioenerg Biomembr. 53:247–258. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
El-Far AH, Al Jaouni SK, Li X and Fu J:
Cancer metabolism control by natural products: Pyruvate kinase M2
targeting therapeutics. Phytother Res. 36:3181–3201. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu Z, Wu J, Zhao Q, Fu S and Jin J:
Emerging roles of aerobic glycolysis in breast cancer. Clin Transl
Oncol. 22:631–646. 2020. View Article : Google Scholar
|
|
10
|
Hua S, Lei L, Deng L, Weng X, Liu C, Qi X,
Wang S, Zhang D, Zou X, Cao C, et al: miR-139-5p inhibits aerobic
glycolysis, cell proliferation, migration, and invasion in
hepatocellular carcinoma via a reciprocal regulatory interaction
with ETS1. Oncogene. 37:1624–1636. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ji H, Li D, Chen L, Shimamura T, Kobayashi
S, McNamara K, Mahmood U, Mitchell A, Sun Y, Al-Hashem R, et al:
The impact of human EGFR kinase domain mutations on lung
tumorigenesis and in vivo sensitivity to EGFR-targeted therapies.
Cancer Cell. 9:485–495. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jamal-Hanjani M, Wilson GA, McGranahan N,
Birkbak NJ, Watkins TBK, Veeriah S, Shafi S, Johnson DH, Mitter R,
Rosenthal R, et al: Tracking the evolution of non-small-cell lung
cancer. N Engl J Med. 376:2109–2121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mertins P, Mani DR, Ruggles KV, Gillette
MA, Clauser KR, Wang P, Wang X, Qiao JW, Cao S, Petralia F, et al:
Proteogenomics connects somatic mutations to signalling in breast
cancer. Nature. 534:55–62. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Srivatsa S, Paul MC, Cardone C, Holcmann
M, Amberg N, Pathria P, Diamanti MA, Linder M, Timelthaler G,
Dienes HP, et al: EGFR in tumor-associated myeloid cells promotes
development of colorectal cancer in mice and associates with
outcomes of patients. Gastroenterology. 153:178–190 e110. 2017.
View Article : Google Scholar
|
|
15
|
Sun C, Wang L, Huang S, Heynen GJJE,
Prahallad A, Robert C, Haanen J, Blank C, Wesseling J, Willems SM,
et al: Reversible and adaptive resistance to BRAF(V600E) inhibition
in melanoma. Nature. 508:118–122. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang B, Zhang Y, Jiang X, Su H, Wang Q,
Wudu M, Jiang J, Ren H, Xu Y, Liu Z and Qiu X: JMJD8 promotes
malignant progression of lung cancer by maintaining EGFR stability
and EGFR/PI3K/AKT pathway activation. J Cancer. 12:976–987. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee JH, Liu R, Li J, Wang Y, Tan L, Li XJ,
Qian X, Zhang C, Xia Y, Xu D, et al: EGFR-phosphorylated platelet
isoform of phosphofructokinase 1 promotes PI3K activation. Mol
Cell. 70:197–210 e197. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhangyuan G, Wang F, Zhang H, Jiang R, Tao
X, Yu D, Jin K, Yu W, Liu Y, Yin Y, et al: VersicanV1 promotes
proliferation and metastasis of hepatocellular carcinoma through
the activation of EGFR-PI3K-AKT pathway. Oncogene. 39:1213–1230.
2020. View Article : Google Scholar
|
|
19
|
Xiao S, Li S, Yuan Z and Zhou L:
Pyrroline-5-carboxylate reductase 1 (PYCR1) upregulation
contributes to gastric cancer progression and indicates poor
survival outcome. Ann Transl Med. 8:9372020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang X, Fu J, Zhong J, Li X, Wang H,
Zhong S, Wei Y, Zhao X, Chen X, Zhou Y, et al: Guanidinylated
cyclic synthetic polypeptides can effectively deliver siRNA by
mimicking the biofunctions of both cell-penetrating peptides and
nuclear localization signal peptides. ACS Macro Lett. 10:767–773.
2021. View Article : Google Scholar
|
|
21
|
Gabrielson NP, Lu H, Yin L, Kim KH and
Cheng J: A cell-penetrating helical polymer for siRNA delivery to
mammalian cells. Mol Ther. 20:1599–1609. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Singh A, Trivedi P and Jain NK: Advances
in siRNA delivery in cancer therapy. Artif Cells Nanomed
Biotechnol. 46:274–283. 2018. View Article : Google Scholar
|
|
23
|
Song Z, Han Z, Lv S, Chen C, Chen L, Yin L
and Cheng J: Synthetic polypeptides: from polymer design to
supramolecular assembly and biomedical application. Chem Soc Rev.
46:6570–6599. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liao W, Du Y, Zhang C, Pan F, Yao Y, Zhang
T and Peng Q: Exosomes: The next generation of endogenous
nanomaterials for advanced drug delivery and therapy. Acta
Biomater. 86:1–14. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang L and Yu D: Exosomes in cancer
development, metastasis, and immunity. Biochim Biophys Acta Rev
Cancer. 1871:455–468. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Y, Liu Q, Zhang X, Huang H, Tang S,
Chai Y, Xu Z, Li M, Chen X, Liu J, et al: Recent advances in
exosome-mediated nucleic acid delivery for cancer therapy. J
Nanobiotechnology. 20:2792022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
National Standard of the P.R.C GB/T
39760-2021 Laboratory animal Guidelines for euthanasia.
|
|
28
|
Song X, Xue Y, Fan S, Hao J and Deng R:
Lipopolysaccharide-activated macrophages regulate the osteogenic
differentiation of bone marrow mesenchymal stem cells through
exosomes. PeerJ. 10:e134422022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Hira VVV, de Jong AL, Ferro K, Khurshed M,
Molenaar RJ and Van Noorden CJF: Comparison of different
methodologies and cryostat versus paraffin sections for chromogenic
immunohistochemistry. Acta Histochem. 121:125–134. 2019. View Article : Google Scholar
|
|
31
|
Cheng C, Song D, Wu Y and Liu B: RAC3
promotes proliferation, migration and invasion via PYCR1/JAK/STAT
signaling in bladder cancer. Front Mol Biosci. 7:2182020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Z, Sun T, Zhang Z, Bi J and Kong C: An
18-gene signature based on glucose metabolism and DNA methylation
improves prognostic prediction for urinary bladder cancer.
Genomics. 113:896–907. 2021. View Article : Google Scholar
|
|
33
|
Cao L, Wu J, Qu X, Sheng J, Cui M, Liu S,
Huang X, Xiang Y, Li B, Zhang X and Cui R: Glycometabolic
rearrangements-aerobic glycolysis in pancreatic cancer: Causes,
characteristics and clinical applications. J Exp Clin Cancer Res.
39:2672020. View Article : Google Scholar
|
|
34
|
Gentric G, Mieulet V and Mechta-Grigoriou
F: Heterogeneity in cancer metabolism: New concepts in an old
field. Antioxid Redox Signal. 26:462–485. 2017. View Article : Google Scholar :
|
|
35
|
Xu H, Ju L, Xiong Y, Yu M, Zhou F, Qian K,
Wang G, Xiao Y and Wang X: E3 ubiquitin ligase RNF126 affects
bladder cancer progression through regulation of PTEN stability.
Cell Death Dis. 12:2392021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Song W, Yang K, Luo J, Gao Z and Gao Y:
Dysregulation of USP18/FTO/PYCR1 signaling network promotes bladder
cancer development and progression. Aging (Albany NY).
13:3909–3925. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Arrighetti N, Corbo C, Evangelopoulos M,
Pasto A, Zuco V and Tasciotti E: Exosome-like nanovectors for drug
delivery in cancer. Curr Med Chem. 26:6132–6148. 2019. View Article : Google Scholar :
|
|
38
|
Li Z and Zhou X, Huang J, Xu Z, Xing C,
Yang J and Zhou X: MicroRNA hsa-miR-150-5p inhibits nasopharyngeal
carcinogenesis by suppressing PYCR1 (pyrroline-5-carboxylate
reductase 1). Bioengineered. 12:9766–9778. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sang S, Zhang C and Shan J:
Pyrroline-5-carboxylate reductase 1 accelerates the migration and
invasion of nonsmall cell lung cancer in vitro. Cancer Biother
Radiopharm. 34:380–387. 2019.PubMed/NCBI
|
|
40
|
Feng J, Li J, Wu L, Yu Q, Ji J, Wu J, Dai
W and Guo C: Emerging roles and the regulation of aerobic
glycolysis in hepatocellular carcinoma. J Exp Clin Cancer Res.
39:1262020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu Q, Qin Y, Ji S, Xu W, Liu W, Sun Q,
Zhang Z, Liu M, Ni Q, Yu X and Xu X: UHRF1 promotes aerobic
glycolysis and proliferation via suppression of SIRT4 in pancreatic
cancer. Cancer Lett. 452:226–236. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
de la Cruz-Lopez KG, Castro-Munoz LJ,
Reyes-Hernandez DO, Garcia-Carranca A and Manzo-Merino J: Lactate
in the regulation of tumor microenvironment and therapeutic
approaches. Front Oncol. 9:11432019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Massari F, Ciccarese C, Santoni M,
Iacovelli R, Mazzucchelli R, Piva F, Scarpelli M, Berardi R,
Tortora G, Lopez-Beltran A, et al: Metabolic phenotype of bladder
cancer. Cancer Treat Rev. 45:46–57. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu W, Hancock CN, Fischer JW, Harman M
and Phang JM: Proline biosynthesis augments tumor cell growth and
aerobic glycolysis: Involvement of pyridine nucleotides. Sci Rep.
5:172062015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tejedor G, Contreras-Lopez R, Barthelaix
A, Ruiz M, Noël D, Ceuninck FD, Pastoureau P, Luz-Crawford P,
Jorgensen C and Djouad F: Pyrroline-5-carboxylate reductase 1
directs the cartilage protective and regenerative potential of
murphy roths large mouse mesenchymal stem cells. Front Cell Dev
Biol. 9:6047562021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang Y, Wang C, Fu Z, Zhang S and Chen J:
miR-30b-5p inhibits proliferation, invasion, and migration of
papillary thyroid cancer by targeting GALNT7 via the EGFR/PI3K/AKT
pathway. Cancer Cell Int. 21:6182021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Makinoshima H, Takita M, Saruwatari K,
Umemura S, Obata Y, Ishii G, Matsumoto S, Sugiyama E, Ochiai A, Abe
R, et al: Signaling through the phosphatidylinositol 3-kinase
(PI3K)/mammalian target of rapamycin (mTOR) axis is responsible for
aerobic glycolysis mediated by glucose transporter in epidermal
growth factor receptor (EGFR)-mutated lung adenocarcinoma. J Biol
Chem. 290:17495–17504. 2015. View Article : Google Scholar : PubMed/NCBI
|