Introduction
Metastasis remains the primary cause of mortality in
patients with colorectal cancer (CRC) (1). The metastatic cascade is a complex,
multi-step process initiated by the detachment of cancer cells from
the primary tumor, a step heavily regulated by cell-matrix adhesion
molecules (2). Alterations in the
expression of integrins and their interactions with the
extracellular matrix (ECM) are fundamental to this process,
allowing tumor cells to acquire invasive properties (3). Consequently, there is an urgent need
to identify novel therapeutic targets that can inhibit the
proliferation, invasion and migration of CRC cells. The advent of
high-throughput technologies and public biological databases has
accelerated research into the genetic and epigenetic alterations
driving CRC, revealing that specific gene mutations are pivotal to
its pathogenesis (4). In-depth
investigation of these aberrantly expressed genes is crucial for
understanding CRC biology and developing effective strategies for
early diagnosis and precision treatment.
The authors' research group previously identified
Integrin Beta 4 (ITGB4) as a gene of significant diagnostic and
therapeutic potential in CRC, leveraging advanced technologies such
as mass cytometry (CyTOF), gene chips and protein arrays. This
discovery led to a national invention patent (Patent Number: ZL
2017 1 1148747.1) (5). ITGB4, a
member of the integrin family, is predominantly expressed on the
basal surface of epithelial cells and forms hemidesmosomes by
dimerizing with integrin alpha 6 (ITGA6) (6). Accumulating evidence indicates that
ITGB4 is overexpressed in various malignancies, including breast,
bladder, cervical, head and neck, lung and pancreatic cancers,
where its expression often correlates with tumor invasion and poor
prognosis (7-14). The authors' prior studies
corroborated these findings in CRC, demonstrating that elevated
ITGB4 levels in both tissues and serum are associated with adverse
clinicopathological features and reduced overall survival (OS)
(15,16). Gene co-expression analysis
suggested that ITGB4's regulatory role in CRC involves multiple
signaling pathways related to cell proliferation, migration and
apoptosis. However, the specific molecular mechanism through which
ITGB4 functions in CRC has not been fully elucidated.
Integrins act as critical biomechanical sensors that
relay signals by connecting the ECM to the intracellular actin
cytoskeleton (17). This linkage
is often mediated by scaffolding proteins such as the
Ezrin-Radixin-Moesin (ERM) family, which crosslink actin filaments
to plasma membrane proteins to regulate cell polarity, adhesion and
migration (18). Given this
functional synergy, it was hypothesized that ITGB4 might
orchestrate CRC progression through specific interactions with
cytoskeletal linkers such as Ezrin (EZR). The present study aimed
to systematically investigate the role of ITGB4 in the initiation
and progression of CRC. Its oncogenic functions were first
validated in vitro and in vivo. Subsequently, through
RNA sequencing (RNA-seq) and a multi-step bioinformatic filtering
process, EZR was identified as a novel downstream target of ITGB4.
It was demonstrated that ITGB4 interacts with and positively
regulates the expression of EZR, thereby activating the
Wnt/β-catenin signaling pathway to promote CRC progression. The
findings of the present study delineate the ITGB4/EZR/Wnt/β-catenin
axis as a critical regulatory network in CRC, offering a potential
new target for precision therapeutic intervention.
Materials and methods
Cell culture
The human CRC cell lines SW480 (cat. no. CCL-228™)
and HCT116 (cat. no. CCL-247™) were purchased from the American
Type Culture Collection. Cells were cultured in Dulbecco's Modified
Eagle's Medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin. Cultures were
maintained in a humidified incubator at 37°C with 5%
CO2. All cells used in experiments were in the
logarithmic growth phase. Cell line authentication was performed
prior to the study, and routine testing confirmed the absence of
mycoplasma contamination.
Small interfering (siRNA), short hairpin
(shRNA), plasmids and transfections
For transient knockdown, siRNAs targeting ITGB4
(siITGB4) and a negative control siRNA (siNC) were synthesized by
Shanghai GenePharma Co., Ltd. For EZR overexpression, a human EZR
open reading frame was cloned into the pcDNA3.1 vector
(pcDNA3.1-EZR), with the empty vector serving as a control
(pcDNA3.1-vector); these were also obtained from Shanghai
GenePharma Co., Ltd. For stable knockdown, shRNAs targeting ITGB4
(shITGB4) and a non-targeting control (shNC) were cloned into a
lentiviral vector by Shanghai GenePharma Co., Ltd. The sequences
for all siRNAs and shRNAs used are listed in Table SI. Transfections were performed
using Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol when cell confluence
reached 80-90%. For experiments performed in 6-well plates, 1.2
μg of plasmid or 50 nM of siRNA was transfected per well.
The transfection was performed at 37°C for 6-8 h. Following
transfection, subsequent experiments were performed after 24 h (for
RNA extraction) or 48 h (for protein extraction).
Generation of stable cell lines
To establish stable ITGB4-knockdown cell lines,
HCT116 cells, cultured to 70% confluence, were transfected with
shITGB4 or shNC vectors. A total of 48 h post-transfection, the
cells were subjected to selection with 600 μg/ml G418
(Neomycin). The selection medium was refreshed every 2-3 days.
After ~1 week, when significant cell death was observed, the G418
concentration was reduced to 300 μg/ml for maintenance.
After 10-14 days of selection, resistant cell colonies were pooled,
expanded, and cultured without G418. The efficiency of stable
knockdown was confirmed by reverse transcription-quantitative PCR
(RT-qPCR) and western blot analysis.
RNA extraction and RT-qPCR
Total RNA was extracted from cells using the
RNA-Easy Kit (Vazyme Biotech Co., Ltd.). cDNA was synthesized from
1 μg of total RNA using the PrimeScript RT Reagent Kit
(Takara Biotechnology Co., Ltd.) according to the manufacturer's
instructions. RT-qPCR was performed on a 4800 Real-Time PCR System
using SYBR Green Master Mix (Vazyme Biotech Co., Ltd.). The thermal
cycling conditions were: 95°C for 5 min, followed by 40 cycles of
95°C for 10 sec and 60°C for 30 sec. Relative gene expression was
calculated using the 2−ΔΔCq method (19), with β-actin serving as the
internal control. Primer sequences are listed in Table SII.
Western blot analysis
Cells were lysed in RIPA buffer (Beijing Solarbio
Science & Technology Co., Ltd.) supplemented with a protease
inhibitor cocktail (PMSF; Beijing Solarbio Science & Technology
Co., Ltd.). Protein concentrations were determined using a BCA
protein assay kit (Beijing Solarbio Science & Technology Co.,
Ltd.). Equal amounts of protein (30 μg) were separated by
10% SDS-PAGE and transferred to polyvinylidene fluoride membranes
(Merck KGaA). The membranes were blocked with 5% non-fat milk in
TBST (0.1% Tween-20) for 1 h at room temperature and then incubated
overnight at 4°C with primary antibodies. The primary antibodies
used were: anti-ITGB4 (1:1,000; cat. no. ab261778; Abcam), anti-EZR
(1:1,000; cat. no. 26056-1-AP; Proteintech Group, Inc.),
anti-β-catenin (1:1,000; cat. no. 51067-2-AP; Proteintech Group,
Inc.), anti-c-Myc (1:1,000; cat. no. 10828-1-AP; Proteintech Group,
Inc.), anti-cyclin D1 (1:500; cat. no. 60186-1-Ig; Proteintech
Group, Inc.) and anti-β-actin (1:1,500; cat. no. TA-09; ZSBG-Bio;
http://www.zsbio.com). After washing, membranes
were incubated with corresponding HRP-conjugated secondary
antibodies (1:2,500; cat. no. A23920; Abbkine Scientific Co.,
Ltd.). Immunoreactive bands were visualized using an Odyssey
scanning system (LI-COR Biosciences). Band intensities were
quantified using ImageJ software (version 1.53k; National
Institutes of Health).
Cell proliferation assays
For the Cell Counting Kit-8 (CCK-8) assay, cells
were seeded into 96-well plates at a density of 1×103
cells/well. At 24, 48, 72 and 96 h, 10 μl of CCK-8 reagent
(Dojindo Laboratories, Inc.) was added to each well, followed by a
2 h incubation at 37°C. The absorbance at 450 nm was measured using
a microplate reader (Promega Corporation). For the colony formation
assay, cells were seeded into 6-well plates at 500 cells/well and
cultured for 10-14 days, with the medium changed every 3 days.
Colonies were fixed with 4% paraformaldehyde (PFA) at room
temperature for 30 min, stained with 0.1% crystal violet at room
temperature for 15 min, and colonies containing >50 cells were
counted.
Cell migration and invasion assays
For the wound healing assay, cells were cultured to
100% confluence in 6-well plates. A scratch was made using a
200-μl pipette tip. After washing with PBS, cells were
cultured in serum-free medium. Images were captured at 0 and 48 h.
The migration rate was calculated based on the change in wound
width. For Transwell assays, 1×105 cells in 200
μl of serum-free medium were seeded into the upper chamber
of a Transwell insert (8.0 μm pore size; Corning, Inc.). The
lower chamber contained 700 μl of DMEM with 10% FBS. For the
invasion assay, the upper chamber was pre-coated with Matrigel (BD
Biosciences) at 37°C for 2 h. After 48 h of incubation,
non-migratory/invasive cells were removed from the upper surface.
Cells on the lower surface were fixed with 4% PFA at room
temperature for 30 min, stained with 0.1% crystal violet at room
temperature for 20 min, and counted in five random fields under a
light microscope.
Apoptosis assay
Cell apoptosis was assessed using the Annexin
V-FITC/PI Apoptosis Detection Kit (Neobioscience Technology Co.,
Ltd.). After 48 h of transfection, cells were harvested, washed
with PBS, and resuspended in binding buffer. Cells were then
stained with Annexin V-FITC and Propidium Iodide (PI) for 15 min in
the dark. The percentage of apoptotic cells was determined by flow
cytometry (BD Biosciences) and analyzed using FlowJo software
(version 10; BD Biosciences).
Animal studies
A total of 12 5-week-old male BALB/c nude mice,
weighing 18-22 g, were housed in a specific pathogen-free (SPF)
facility under a 12/12-h light/dark cycle with free access to food
and water. They were randomly divided into two groups (n= 6 per
group). HCT116 cells (2×106) stably expressing shNC or
shITGB4 were suspended in 100 μl of serum-free DMEM and
injected subcutaneously into the right flank of each mouse. Tumor
growth was monitored every 3 days by measuring the length (L) and
width (W) with calipers. Tumor volume was calculated using the
formula: V=(L × W2)/2. After 21 days, all animals were
euthanized by cervical dislocation after anesthesia with
intraperitoneal injection of 40-50 mg/kg sodium pentobarbital, and
tumors were excised, weighed, and processed for histological
analysis. Tumors were fixed at 4°C overnight in 4% PFA for
subsequent H&E and immunohistochemistry (IHC) staining. The
study protocols were approved by the Experimental Animal Care and
Use Committee and Ethics Committee of The First Hospital of Hebei
Medical University (approval no. 20220395; Shijiazhuang,
China).
RNA sequencing (RNA-seq) and
bioinformatic analysis
Total RNA was extracted from SW480 cells transfected
with siNC or siITGB4 (three biological replicates per group) using
TRIzol™ reagent (cat. no. 5596026CN; Thermo Fisher Scientific,
Inc.). RNA quality and integrity were verified using the Agilent
2100 Bioanalyzer. The type of sequencing was paired-end, with a
nucleotide length of 150 bp, with TruSeq RNA Exome Kit (cat. no.
RS-301-2001; Illumina, Inc.) used for sequencing. Library
construction and sequencing were performed by Novogene Technology
Co., Ltd. on an Illumina NovaSeq 6000 platform. The loading
concentration of the final library was 1.5 ng/μl and was
measured using a Qubit 2.0 Fluorometer. After quality control,
clean reads were aligned to the human reference genome (GRCh38).
Differential expression analysis was performed using DESeq2
(version 1.20.0). Genes with an adjusted P<0.05 and a log2|Fold
Change| >1.0 were considered differentially expressed genes
(DEGs).
IHC
Paraffin-embedded tumor sections (4 μm) were
deparaffinized with xylene, rehydrated using ethanol series, and
subjected to antigen retrieval. Sections were then incubated with
an anti-Ki67 antibody (1:100; cat. no. 27309-1-AP; Proteintech
Group, Inc.) overnight at 4°C. Staining was performed using a
two-step detection kit (ZSBG-Bio) and visualized with DAB. Images
were captured using a light microscope, and the staining intensity
was quantified as the average optical density using ImageJ
software.
Co-immunoprecipitation (Co-IP) and
Co-immunofluorescence (Co-IF)
For Co-IP, SW480 cell lysates were incubated with
anti-ITGB4 antibody (10 μg; Proteintech Group, Inc.),
anti-EZR antibody (10 μg; Proteintech Group, Inc.), or
control IgG (10 μg; Proteintech Group, Inc.) using the
Pierce™ Classic Magnetic IP/Co-IP Kit (cat. no. 88804; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Immunoprecipitated proteins were analyzed by western
blotting. For Co-IF, SW480 cells cultured on glass coverslips were
fixed with 4% PFA at room temperature for 30 min, permeabilized
with 0.2% Triton X-100, and blocked with 2% BSA at room temperature
for 30 min. Cells were then incubated with primary antibodies
against ITGB4 (1:50; cat. no. sc-13543; Santa Cruz Biotechnology,
Inc.) and EZR (1:50; cat. no. 26056-1-AP; Proteintech Group, Inc.)
overnight at 4°C. After washing, cells were incubated with
Cy3-conjugated anti-rabbit and FITC-conjugated anti-mouse secondary
antibodies (cat. nos. A0516 and A0568; Beyotime Institute of
Biotechnology. Nuclei were counterstained with DAPI (1 mg/ml; cat.
no. C1002; Beyotime Institute of Biotechnology). Images were
acquired using a Zeiss laser scanning confocal microscope (Carl
Zeiss AG).
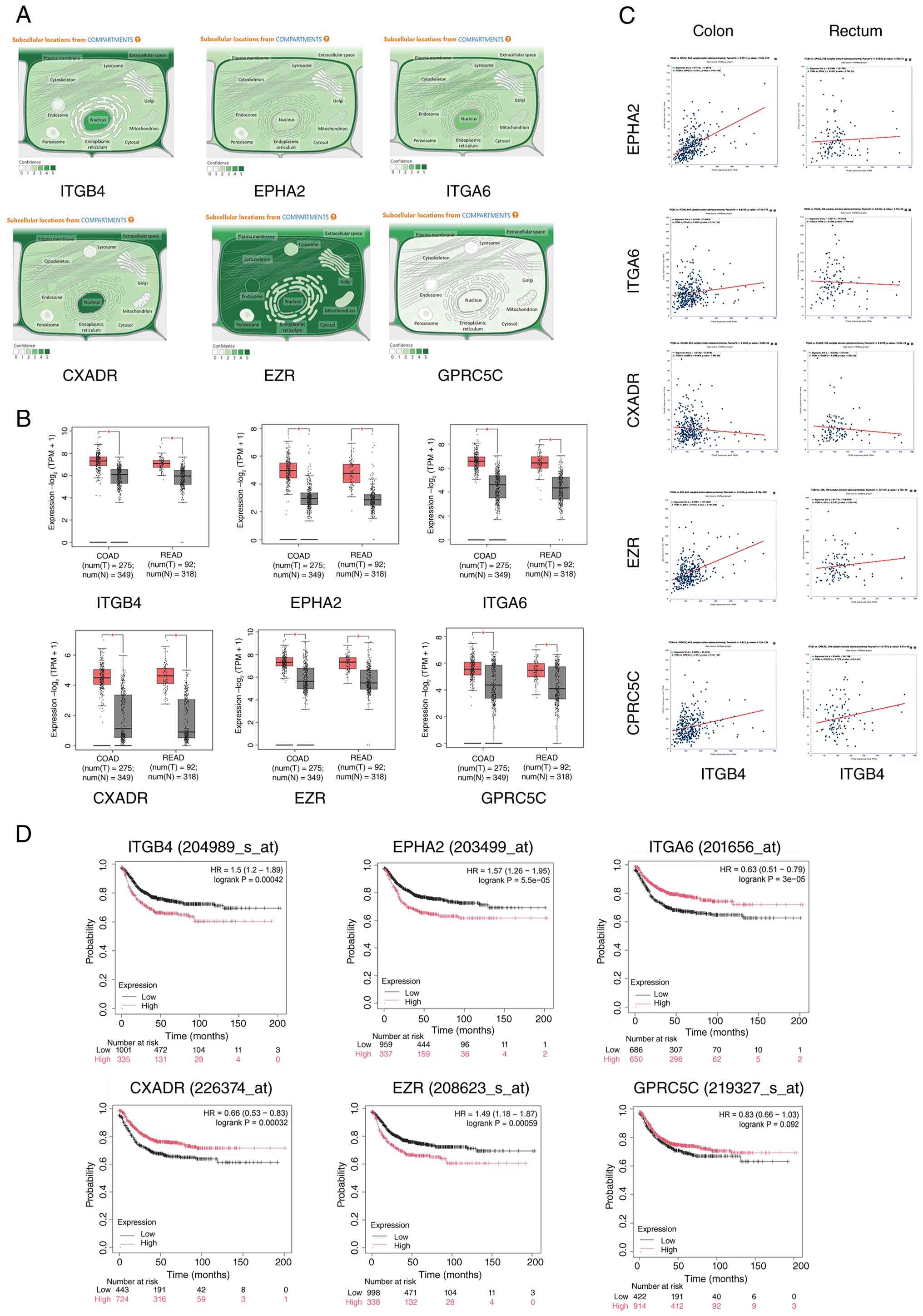
Public database analysis
ITGB4 expression data in CRC and adjacent normal
tissues were obtained from The Cancer Genome Atlas (TCGA) and Gene
Expression Profiling Interactive Analysis 2 (GEPIA2; http://gepia2.cancer-pku.cn/) databases. The TCGA and
GEPIA2 datasets were prioritized due to their large sample sizes,
standardized high-throughput sequencing platforms, and
comprehensive clinical annotations, which provide robust
statistical power for differential expression and survival analyses
compared with smaller, individual datasets. Inclusion criteria
involved datasets containing paired tumor and adjacent normal
tissues with complete follow-up information. Although
clinicopathological variables such as tumor stage and subtype are
available in these datasets, the current analysis focused on
overall differential expression and survival correlations to
identify potential targets, without stratification by specific
parameters. The prognostic value of gene expression was assessed
using the Kaplan-Meier plotter (https://kmplot.com/analysis/) followed by the log-rank
test. Subcellular localization data were retrieved from GeneCards
(https://www.genecards.org/). STRING
database (https://string-db.org/) was used for PPI
interaction network.
Statistical analysis
All experiments were performed with at least three
independent replicates. Data are presented as the mean ± standard
deviation (SD). Statistical analyses were performed using GraphPad
Prism 8.0 (Dotmatics) and SPSS 21.0 (IBM Corp.). Differences
between two groups were analyzed using an unpaired two-tailed
Student's t-test. Comparisons among multiple groups were performed
using one-way or two-way ANOVA followed by Bonferroni post-hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
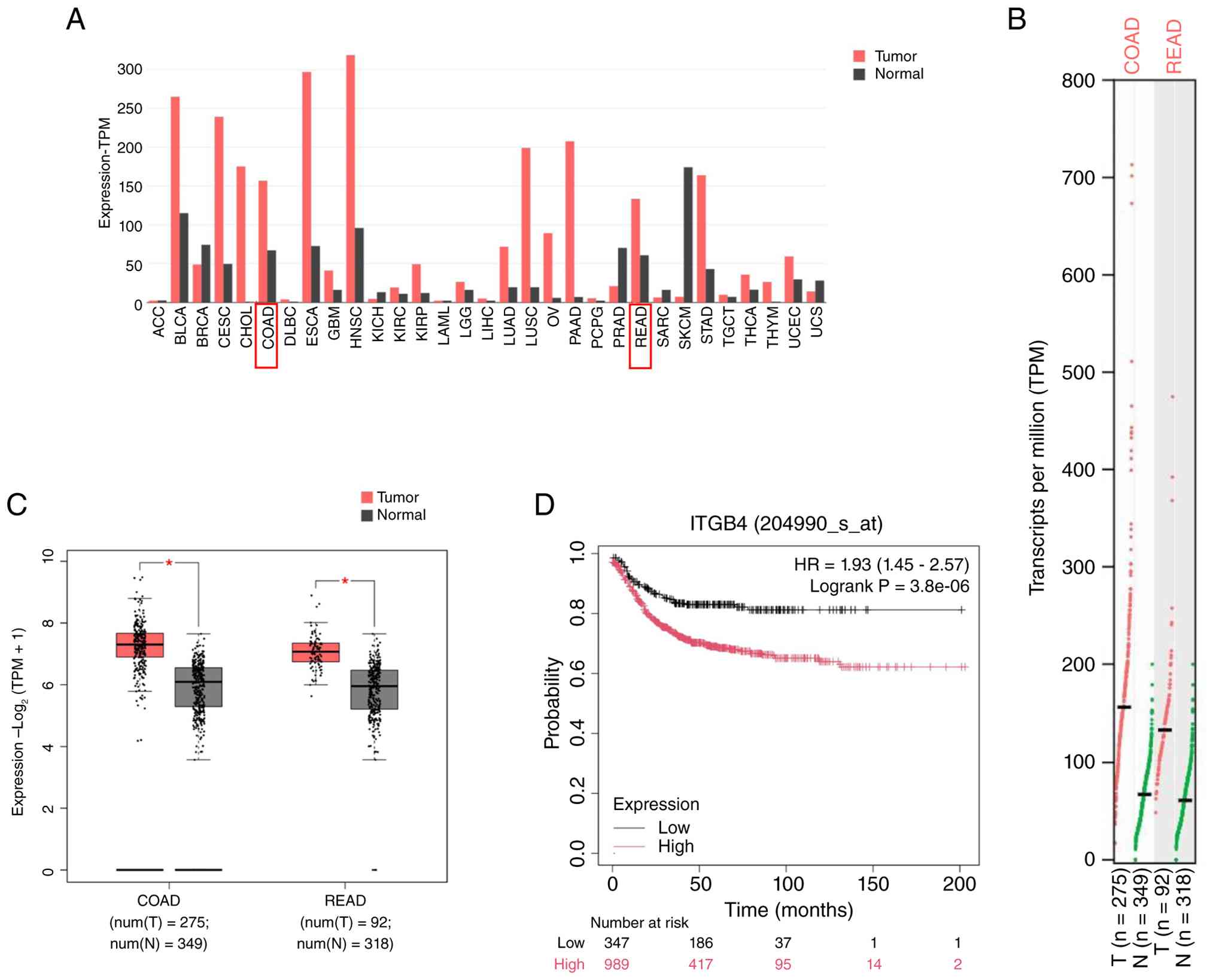
ITGB4 is upregulated in CRC and
correlates with poor prognosis
To corroborate previous findings, ITGB4 expression
was first analyzed using public databases. Analysis of TCGA and
GEPIA2 datasets confirmed that ITGB4 mRNA expression was
significantly higher in CRC tissues compared with adjacent normal
tissues (Fig. 1A-C). Furthermore,
Kaplan-Meier survival analysis revealed that patients with high
ITGB4 expression had significantly poorer OS than those with low
expression (Fig. 1D). These
results are consistent with the authors' prior study (6) and collectively underscore that ITGB4
is an oncogenic factor in CRC, and its overexpression is associated
with unfavorable patient outcomes, highlighting its potential as a
prognostic biomarker.
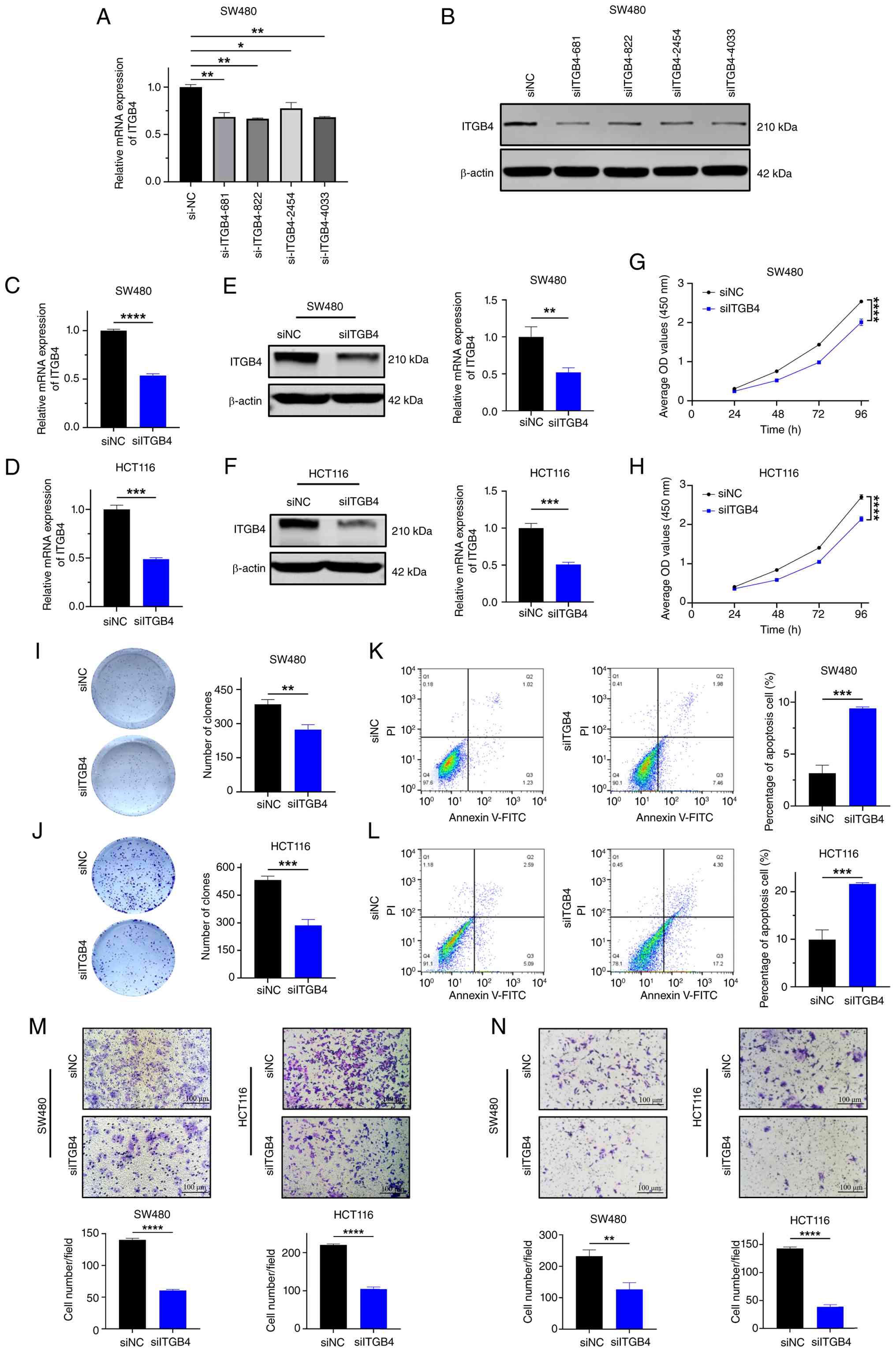
Downregulation of ITGB4 suppresses
malignant phenotypes of CRC cells in vitro
To investigate the biological function of ITGB4 in
CRC, its expression in SW480 and HCT116 cells was first silenced
using siRNAs. Four different siRNA sequences were tested, and the
siITGB4-681 sequence, which demonstrated the most potent knockdown
efficiency at both mRNA and protein levels, was selected for
subsequent experiments (Fig. 2A and
B). Transient transfection with siITGB4-681 effectively reduced
ITGB4 mRNA and protein expression in both cell lines (Fig. 2C-F). Functional assays revealed
that ITGB4 knockdown significantly inhibited cell proliferation, as
determined by CCK-8 assays (Fig. 2G
and H), and suppressed clonogenic ability, as shown by colony
formation assays (Fig. 2I and J).
Moreover, flow cytometric analysis indicated that ITGB4 silencing
significantly increased the proportion of apoptotic cells (Fig. 2K and L). Transwell assays
demonstrated a significant reduction in both the migratory and
invasive capacities of CRC cells upon ITGB4 knockdown (Fig. 2M and N). Collectively, these in
vitro results suggest that ITGB4 plays a critical role in
promoting CRC cell proliferation, survival, migration and
invasion.
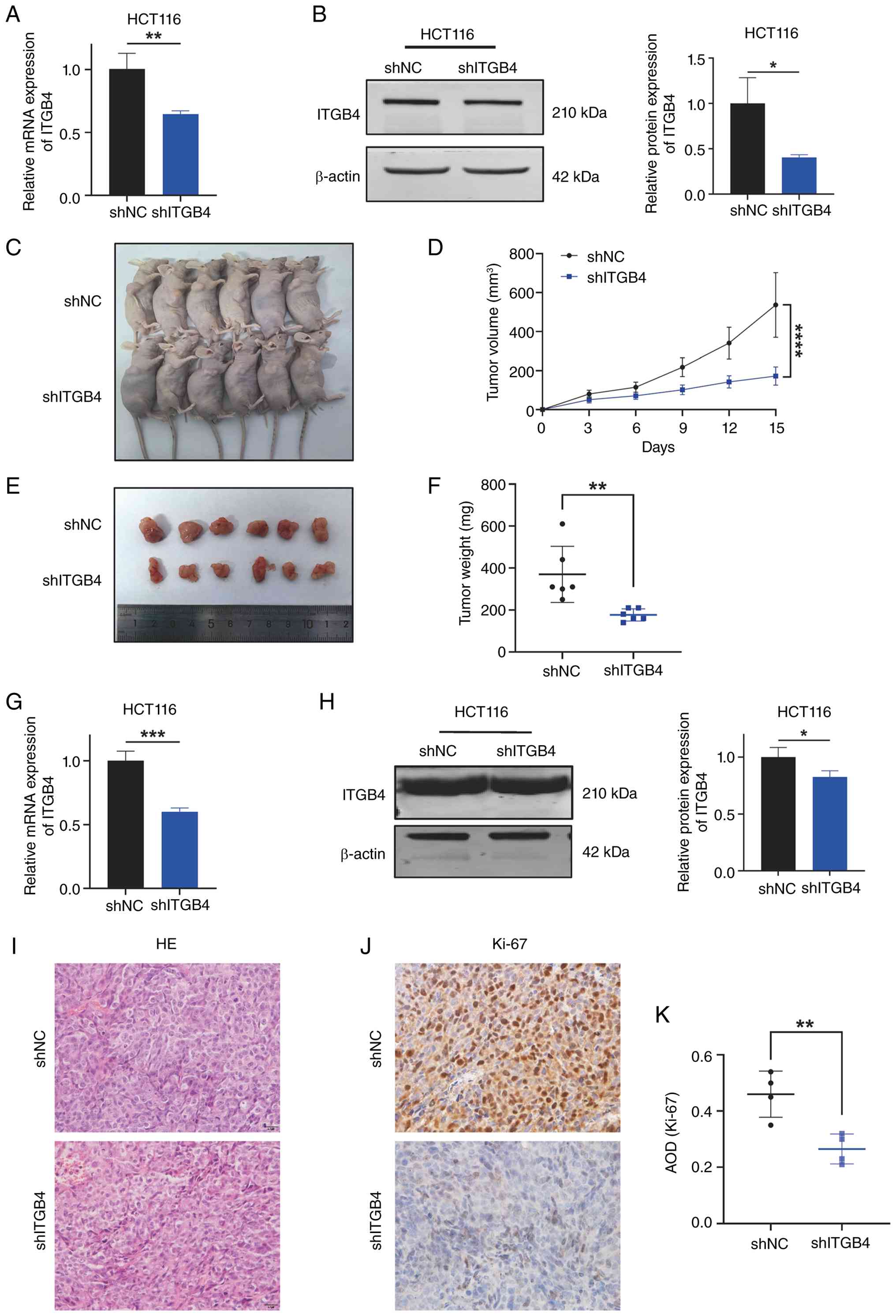
Knockdown of ITGB4 inhibits CRC growth in
vivo
To extend the in vitro findings, the role of
ITGB4 in tumor growth was assessed using a xenograft model. An
HCT116 cell line with stable ITGB4 knockdown (shITGB4) was
established, confirming sustained suppression of ITGB4 expression
(Fig. 3A and B). These cells,
along with control cells (shNC), were subcutaneously injected into
nude mice. Tumor growth was monitored over 21 days. The tumors
formed by shITGB4 cells grew significantly slower and were
substantially smaller and lighter at the end of the experiment
compared with tumors from the shNC group (Fig. 3C-F). The reduced ITGB4 expression
in the xenograft tumors was verified by RT-qPCR and western
blotting (Fig. 3G and H). H&E
staining confirmed the CRC tissue morphology in the tumors
(Fig. 3I). Furthermore, IHC
staining for the proliferation marker Ki-67 showed significantly
lower positivity in the shITGB4 group, indicating reduced cell
proliferation (Fig. 3J and K).
These in vivo data provide strong evidence that ITGB4 is
essential for promoting CRC tumorigenesis.
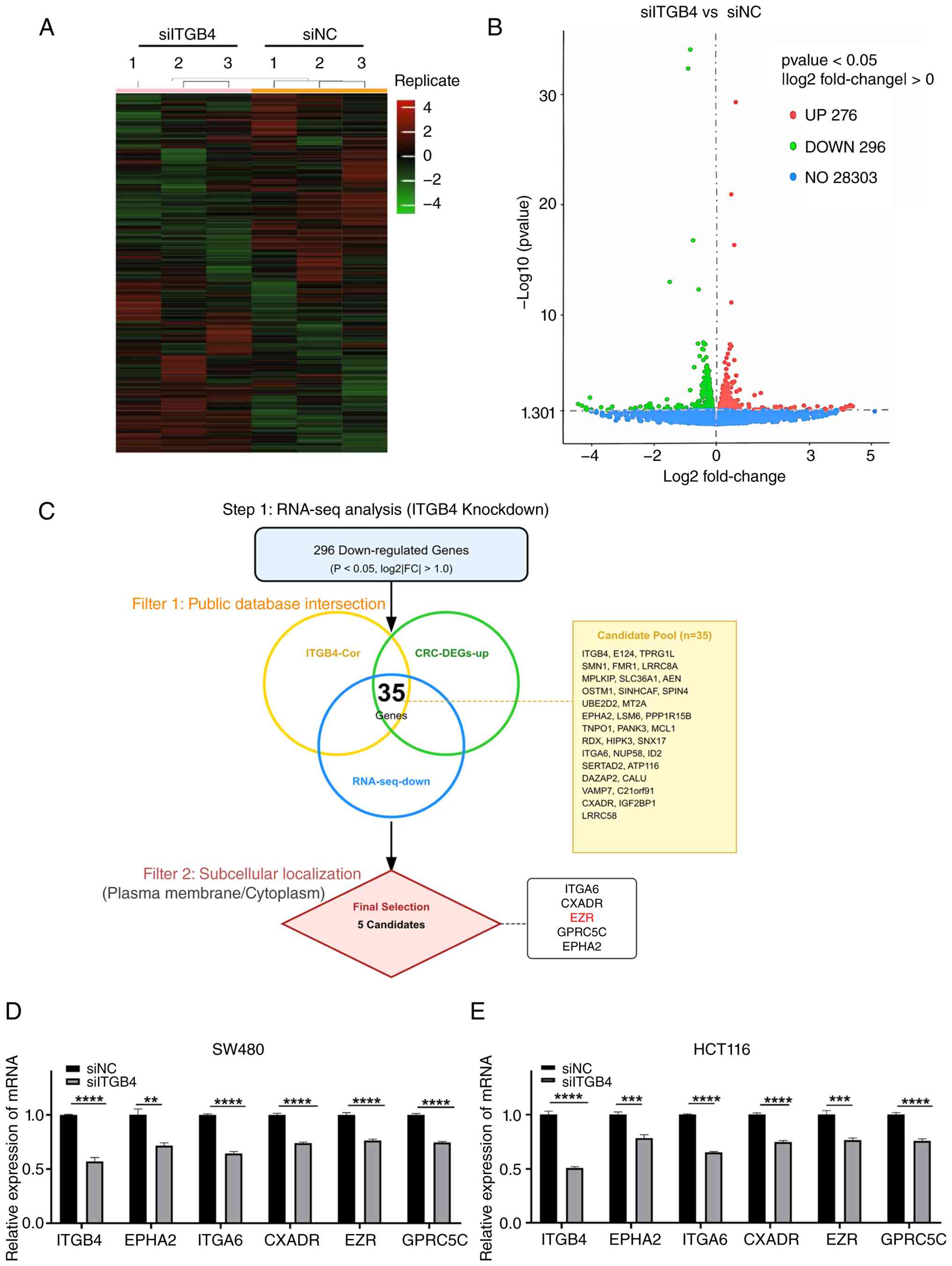
Identification of EZR as a downstream
target of ITGB4 in CRC
To uncover the molecular mechanisms downstream of
ITGB4, RNA-seq was performed on SW480 cells following ITGB4
knockdown. This analysis identified 572 DEGs, of which 296 were
downregulated and 276 were upregulated (Fig. 4A and B). To pinpoint key
downstream effectors, a stringent, multi-step filtering strategy
was devised. First, focus was addressed on the 296 downregulated
genes from our RNA-seq data. This list was then intersected with
two externally derived gene sets: (i) Genes known to be
co-expressed with ITGB4 in CRC patient cohorts (from the GEPIA
database) and (ii) genes that are significantly upregulated in CRC
tissues compared with normal tissues (from the GEPIA2 database).
This intersection yielded a refined list of 35 candidate genes. To
further narrow the selection, genes whose protein products share a
subcellular localization with ITGB4 (that is, plasma membrane or
cytoplasm, based on GeneCards data), were prioritized. This
criterion was selected because ITGB4 is a transmembrane receptor;
therefore, its immediate downstream signal transducers are likely
to be physically proximal, residing at the membrane or in the
sub-membrane cortical cytoplasm. This systematic process identified
five high-confidence candidate target genes: ITGA6, CXADR, EZR,
GPRC5C and EPHA2 (Fig. 4C). The
regulatory relationship was validated by confirming that knockdown
of ITGB4 led to a significant decrease in the mRNA levels of all
five candidate genes in both SW480 and HCT116 cells (Fig. 4D and E). Among the candidates,
EZR, CXADR and ITGA6 showed predominantly plasma membrane
localization, similar to ITGB4, whereas GPRC5C showed mixed
localization. This spatial proximity is a prerequisite for direct
protein-protein interaction (PPI). Among these candidates, EZR was
selected for further investigation based on its strong positive
correlation with ITGB4 expression in CRC patient samples (Fig. 5C), its consistent upregulation in
CRC tissues (Fig. 5B), its shared
subcellular localization (Fig.
5A) and its strong association with poor patient prognosis
(Fig. 5D), which mirrored the
prognostic significance of ITGB4.
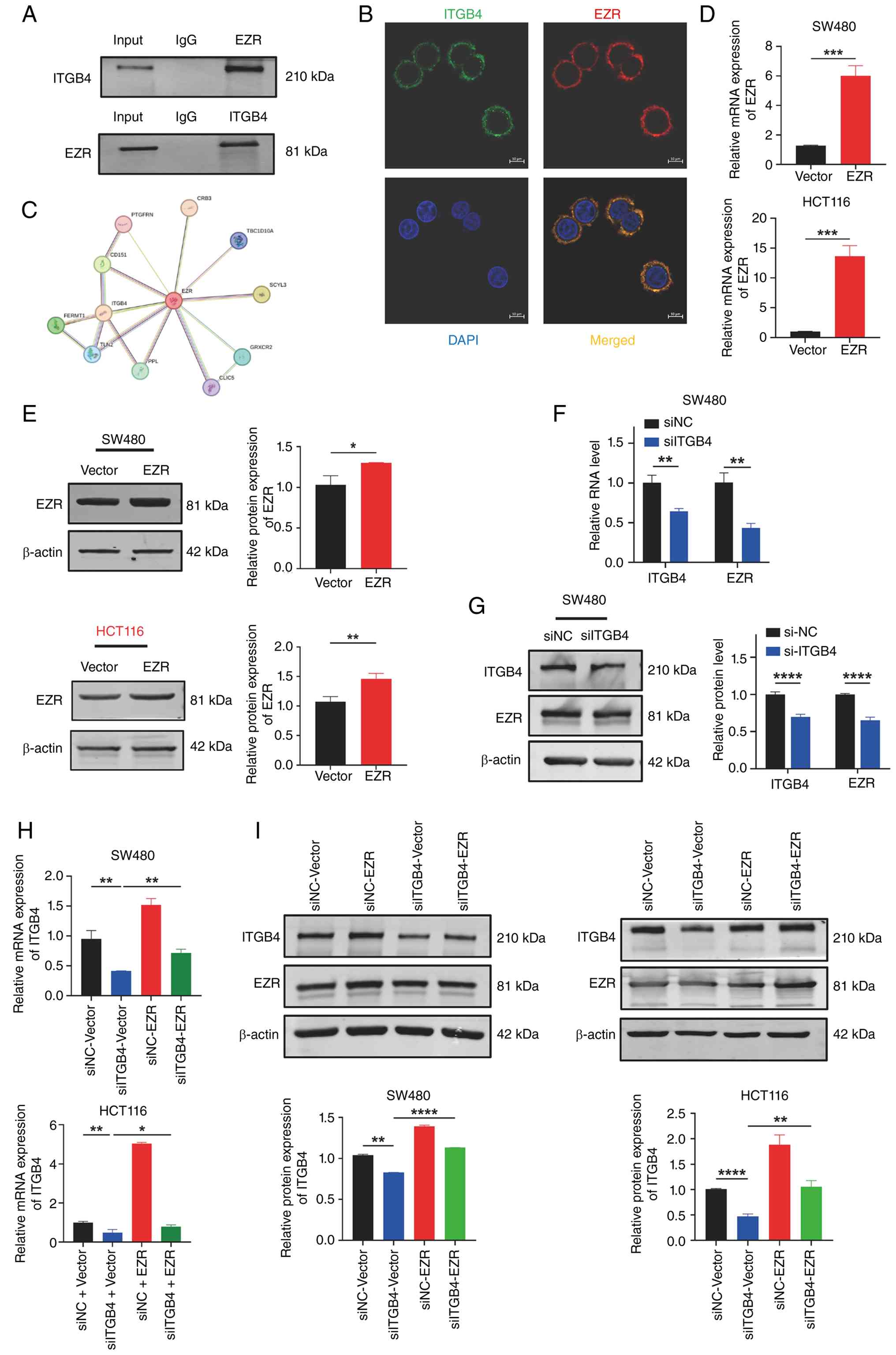
ITGB4 interacts with and positively
regulates the expression of EZR
Given the strong correlational evidence, a potential
physical interaction between ITGB4 and EZR was next investigated.
Co-IP assays in SW480 cells demonstrated that endogenous ITGB4
could be immunoprecipitated with an anti-EZR antibody, and
conversely, EZR was pulled down with an anti-ITGB4 antibody,
confirming that the two proteins exist in the same complex
(Fig. 6A). Co-IF staining further
revealed that ITGB4 and EZR extensively co-localize at the plasma
membrane of CRC cells (Fig. 6B).
This interaction is also supported by PPI network analysis from the
STRING database (Fig. 6C). It was
then confirmed that ITGB4 regulates EZR expression. As shown in
Fig. 6F and G, knockdown of ITGB4
in SW480 cells resulted in a significant reduction of both EZR mRNA
and protein levels. To assess the functional hierarchy, rescue
experiments were performed. After establishing an efficient EZR
overexpression system (Fig. 6D and
E), cells were co-transfected with siITGB4 and the EZR
overexpression plasmid. Interestingly, overexpression of EZR
partially restored the suppressed levels of ITGB4 mRNA and protein
induced by siITGB4 (Fig. 6H and
I). Given that EZR functions as a downstream effector that
activates Wnt/β-catenin signaling, this concomitant increase in
ITGB4 suggests the potential existence of a positive feedback loop.
It was hypothesized that hyperactive Wnt/β-catenin signaling may
transcriptionally upregulate ITGB4 to further sustain the malignant
progression. Collectively, these data establish that ITGB4
interacts with EZR and positively regulates its expression at both
transcriptional and post-transcriptional levels.
ITGB4 promotes CRC progression by
regulating EZR to activate the Wnt/β-catenin signaling pathway
To determine the signaling pathway through which the
ITGB4/EZR axis functions, Kyoto Encyclopedia of Genes and Genomes
pathway enrichment analysis was performed on the DEGs from the
current RNA-seq data. The analysis revealed a significant
enrichment in the Wnt/β-catenin signaling pathway (Fig. 7A). To validate this, key proteins
of the pathway were examined by western blotting. ITGB4 knockdown
in HCT116 cells led to a marked decrease in the levels of active
β-catenin and its downstream transcriptional targets, c-Myc and
Cyclin D1. Crucially, this downregulation was reversed by the
ectopic overexpression of EZR (Fig.
7B). This indicates that ITGB4 activates the Wnt/β-catenin
pathway in an EZR-dependent manner. Finally, functional rescue
experiments were conducted to confirm that EZR mediates the
oncogenic effects of ITGB4. As revealed in Fig. 7C-K, while ITGB4 knockdown
suppressed CRC cell proliferation, migration and invasion,
concomitant overexpression of EZR significantly rescued these
phenotypes. Taken together, these results demonstrate that ITGB4
promotes CRC progression by regulating EZR, which in turn leads to
the activation of the Wnt/β-catenin signaling cascade.
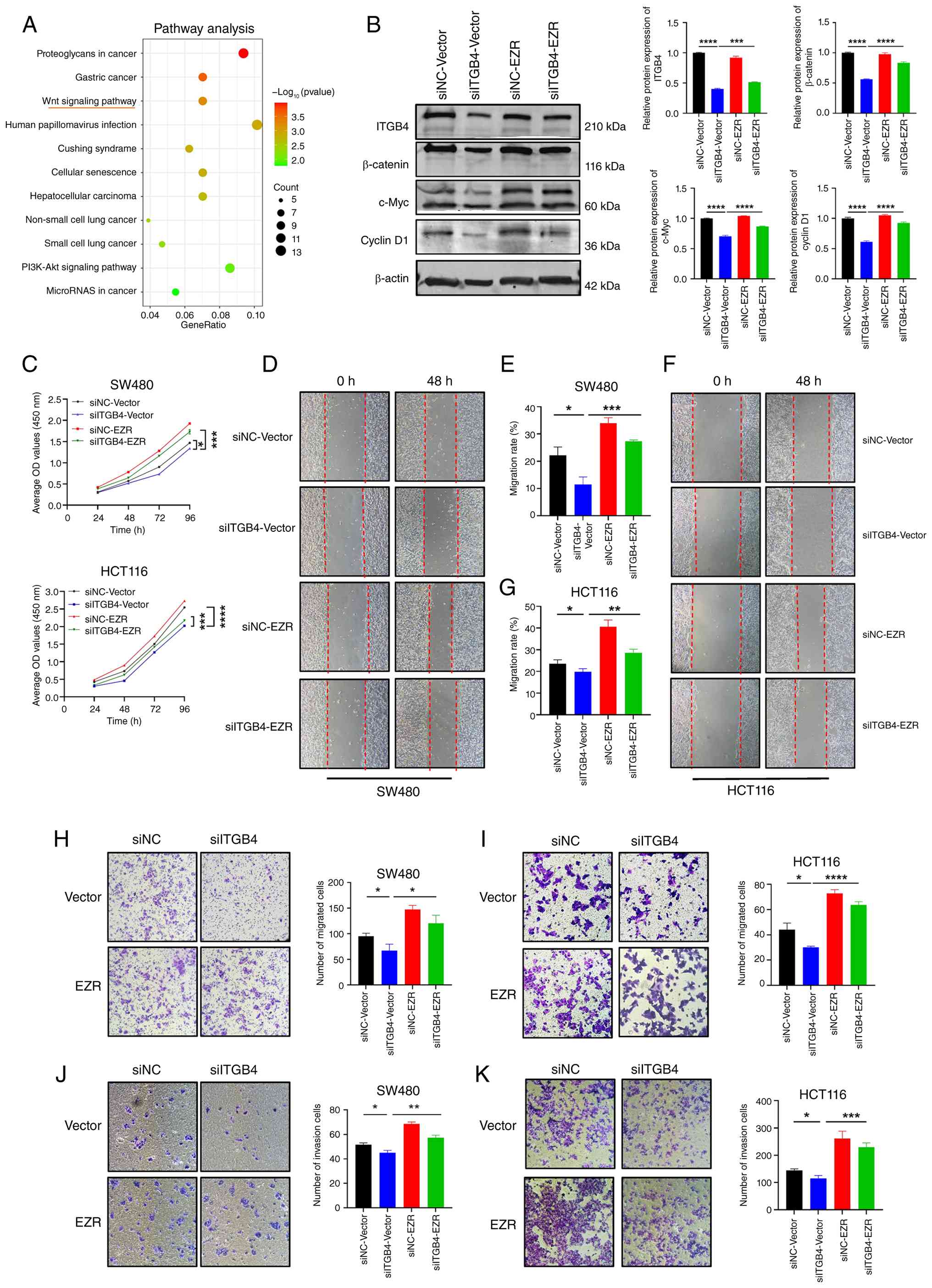 | Figure 7ITGB4 exerts its oncogenic function
through the EZR-mediated activation of the Wnt/β-catenin signaling
pathway. (A) Kyoto Encyclopedia of Genes and Genomes pathway
enrichment analysis of differentially expressed genes following
ITGB4 knockdown, highlighting the Wnt signaling pathway. (B)
Western blot analysis and corresponding quantification showing that
ITGB4 knockdown suppresses the expression of β-catenin, c-Myc and
Cyclin D1, and this effect is rescued by EZR overexpression in
HCT116 cells. (C) Cell Counting Kit-8 assays demonstrating that EZR
overexpression rescues the proliferation defect caused by ITGB4
knockdown in SW480 and HCT116 cells. (D-G) Wound healing assays
showing that EZR overexpression restores the migratory capacity
inhibited by ITGB4 knockdown (magnification, ×200). (H and I)
Transwell migration assays confirming the rescue of cell migration
by EZR overexpression (crystal violet staining; scale bar, 100
μm). (J and K) Transwell invasion assays showing that EZR
overexpression rescues the invasive potential suppressed by ITGB4
knockdown (crystal violet staining; scale bar, 100 μm). Data
are presented as the mean ± SD. *P<0.05,
**P<0.01, ***P<0.001 and
****P<0.0001. ITGB4, integrin beta 4; EZR, Ezrin;
si-, small interfering; NC, negative control. |
Discussion
In the present study, a novel molecular mechanism
underlying the pro-tumorigenic role of ITGB4 in CRC was elucidated.
It was confirmed that ITGB4 is significantly overexpressed in CRC
tissues and that its high expression is associated with poor
patient prognosis. Through a series of in vitro and in
vivo experiments, it was demonstrated that ITGB4 is critical
for promoting CRC cell proliferation, migration, invasion and
survival. Mechanistically, EZR was identified as a key downstream
effector of ITGB4 for the first time. The current data reveal that
ITGB4 interacts with and positively regulates EZR expression, which
subsequently activates the Wnt/β-catenin signaling pathway, a
well-established driver of CRC. These findings highlight the
ITGB4/EZR/Wnt/β-catenin axis as a crucial signaling cascade in CRC
progression and suggest its potential as a therapeutic target.
ITGB4, a transmembrane receptor, plays a pivotal
role in cell-matrix adhesion and signal transduction (6). Its aberrant expression has been
linked to the progression of various cancers (9-14).
Consistent with these studies and the authors' previous findings
(16), the present study
solidifies the oncogenic function of ITGB4 in CRC. The functional
experiments clearly demonstrated that silencing ITGB4 potently
inhibits key malignant phenotypes, including proliferation and
invasion, both in cultured cells and in a xenograft tumor model.
These results strongly support ITGB4's role as a driver of CRC and
reinforce its value as a potential therapeutic target.
A key innovation of the present study is the
identification of EZR as a downstream target regulated by ITGB4.
EZR is a member of the ERM protein family that links the plasma
membrane to the actin cytoskeleton, thereby regulating cell
adhesion, migration and signaling (20). EZR is known to be overexpressed in
numerous malignancies, including CRC, where its expression
correlates with advanced stage, metastasis and poor prognosis
(21-25). While both ITGB4 and EZR have been
independently implicated in cancer, a direct regulatory link
between them in the context of CRC has not been previously
described. The present study provides compelling evidence for this
connection through Co-IP and Co-IF experiments demonstrating their
physical interaction and co-localization. The co-localization of
ITGB4 and EZR at the plasma membrane is biologically significant.
As a transmembrane receptor, ITGB4 requires membrane-proximal
effectors to transmit extracellular signals. EZR, serving as a
cross-linker between the plasma membrane and the actin
cytoskeleton, is ideally positioned to transduce ITGB4-mediated
signals to the intracellular machinery. Furthermore, it was
identified that ITGB4 expression level directly influences EZR mRNA
and protein levels, suggesting that ITGB4 may regulate EZR through
transcriptional or post-transcriptional mechanisms. While the
precise mode of this regulation (for example, via downstream
transcription factors or effects on mRNA stability) warrants
further investigation, the present results firmly place EZR as a
downstream mediator of ITGB4's oncogenic signaling.
The activation of the Wnt/β-catenin signaling
pathway is a hallmark event in the majority of CRC, driving
uncontrolled cell proliferation and survival (26,27). The pathway culminates in the
nuclear translocation of β-catenin, which acts as a co-activator
for TCF/LEF transcription factors to induce the expression of
target genes such as c-Myc and Cyclin D1 (28). The present study connects the
ITGB4/EZR axis to this critical pathway. It was demonstrated that
ITGB4 knockdown deactivates the Wnt/β-catenin pathway and that this
effect is mediated by EZR, as EZR overexpression could restore
pathway activity. This finding is particularly significant as it
provides a mechanistic explanation for how an upstream cell
adhesion molecule such as ITGB4 can influence a core intracellular
oncogenic signaling cascade. The ability of EZR to rescue the
malignant phenotypes suppressed by ITGB4 knockdown further
solidifies the functional importance of this newly identified axis.
Furthermore, the current data serendipitously revealed that EZR
overexpression could partially restore ITGB4 levels. Since
β-catenin acts as a prominent transcription factor, it was
hypothesized that ITGB4 could be a transcriptional target of the
Wnt/β-catenin pathway. This potential positive feedback loop
(ITGB4/EZR/Wnt/β-catenin/ITGB4) may continuously fuel CRC
progression, which remains an intriguing hypothesis that warrants
dedicated exploration and direct transcriptional validation in
future studies.
Mechanistically, EZR may activate Wnt signaling
through several potential pathways. A previous study in
osteosarcoma indicated that EZR can physically interact with
β-catenin and facilitate its nuclear translocation (29). Alternatively, EZR may disrupt the
E-cadherin/β-catenin complex at the cell membrane, releasing a pool
of β-catenin that acts as a signaling molecule rather than an
adhesion component. Furthermore, EZR phosphorylation is a key
regulatory event that modulates multiple downstream signaling
cascades, including those that converge on β-catenin stability
(30). The present findings in
CRC are consistent with these models, suggesting that EZR acts as a
critical signal transducer.
While other integrins, such as Beta 1, have been
reported to modulate Wnt signaling via focal adhesion kinase, the
ITGB4-mediated regulation appears distinct. Unlike Beta 1, ITGB4
primarily forms hemidesmosomes. The current data suggests that in
cancer cells, ITGB4 signaling is re-wired away from stable adhesion
towards pro-migratory signaling via EZR, a mechanism that may be
unique to the structural properties of the Beta 4 subunit.
Nevertheless, the present study has certain
limitations. First, while a regulatory relationship was
demonstrated, the precise molecular details of how ITGB4 modulates
EZR expression require further exploration. Future studies could
investigate whether ITGB4 signaling affects transcription factors
that bind to the EZR promoter or influences the stability of EZR
mRNA. Second, the clinical correlation between ITGB4 and EZR
expression should be validated in a larger cohort of CRC patient
samples to strengthen its clinical relevance. Additionally, while
in vitro assays strongly support a role for ITGB4 in
migration and invasion, the subcutaneous xenograft model used in
the present study primarily assesses tumor growth. Future studies
utilizing orthotopic implantation or tail vein injection models are
necessary to fully validate the metastatic potential of the
ITGB4/EZR axis in vivo. Despite these limitations, the
present study provides a robust foundation for future research.
In conclusion, the present study identifies ITGB4 as
a critical oncogene in CRC that promotes tumor progression by
upregulating its downstream target EZR, leading to the activation
of the Wnt/β-catenin signaling pathway. This newly defined
ITGB4/EZR/Wnt/β-catenin axis offers novel insights into the
molecular pathogenesis of CRC and presents ITGB4 as a promising
biomarker for prognosis and a potential target for therapeutic
intervention.
Supplementary Data
Availability of data and materials
The data generated in the present study may be found
in the Gene Expression Omnibus under accession number GSE326923 or
at the following URL: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE326923.
The data generated in the present study may be requested from the
corresponding author.
Authors' contributions
WY, JiaW and TL conceived and designed the study.
JinW, YS and MX performed the experiments, analyzed the data, and
wrote the manuscript. SH, KL and JJ provided technical support and
study materials. XM and HL contributed to data collection and
assembly. All authors read and approved the final version of the
manuscript. JinW and YS confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
All animal procedures were approved by the
Experimental Animal Care and Use Committee and the Ethics Committee
of The First Hospital of Hebei Medical University (approval no.
20220395; Shijiazhuang, China). The study was carried out in
compliance with the ARRIVE guidelines and all methods were
performed in accordance with the relevant guidelines and
regulations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
CRC
|
colorectal cancer
|
|
ITGB4
|
integrin beta 4
|
|
EZR
|
ezrin
|
|
CCK-8
|
Cell Counting Kit-8
|
|
RNA-seq
|
RNA sequencing
|
|
Co-IP
|
co-immunoprecipitation
|
|
Co-IF
|
co-immunofluorescence
|
|
DMEM
|
Dulbecco's Modified Eagle's Medium
|
|
FBS
|
fetal bovine serum
|
|
siRNA
|
small interfering RNA
|
|
shRNA
|
short hairpin RNA
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
IHC
|
immunohistochemistry
|
|
DEGs
|
differentially expressed genes
|
|
TCGA
|
The Cancer Genome Atlas
|
Acknowledgements
Not applicable.
Funding
The present study was supported by the Hebei Provincial
Government-funded Provincial Medical Excellent Talent Project
(grant nos. ZF2023025, ZF2024134, ZF2025045, ZF2025048, ZF2025051,
LS202208 and LS202212), the Hebei Natural Science Foundation (grant
nos. H2022206292 and H2024206140), the Key R&D Program of Hebei
(grant nos. 223777103D and 223777113D), the Hebei County General
Hospital Appropriate Health Technology Promotion Project (grant no.
20220018), the Prevention and treatment of geriatric diseases by
Hebei Provincial Department of Finance (grant nos. LNB202202,
LNB201809 and LNB201909), the Spark Scientific Research Project of
the First Hospital of Hebei Medical University (grant nos. XH202312
and XH201805), the Hebei Medical Applicable Technology Tracking
Project (grant no. G2019035) and other projects of Hebei (grant
nos. 1387 and SGH201501).
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249.
2021.PubMed/NCBI
|
|
2
|
Biller LH and Schrag D: Diagnosis and
treatment of metastatic colorectal cancer: A review. JAMA.
325:669–685. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Winkler J, Abisoye-Ogunniyan A, Metcalf KJ
and Werb Z: Concepts of extracellular matrix remodelling in tumour
progression and metastasis. Nat Commun. 11:51202020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Janiszewska M, Primi MC and Izard T: Cell
adhesion in cancer: Beyond the migration of single cells. J Biol
Chem. 295:2495–2505. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Han S, Jiang X, Sun XF, Zhang H, Li C,
Zhao Z and Yu W: Application value of CyTOF 2 mass cytometer
technology at single-cell level in human gastric cancer cells. Exp
Cell Res. 384:1115682019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Borradori L and Sonnenberg A: Structure
and function of hemidesmosomes: More than simple adhesion
complexes. J Invest Dermatol. 112:411–418. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang W, Zhang B, Vu T, Yuan G, Zhang B,
Chen X, Manne U and Datta PK: Molecular characterization of
pro-metastatic functions of β4-integrin in colorectal cancer.
Oncotarget. 8:92333–92345. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong F, Lu HP, Chen G, Dang YW, Li GS,
Chen XY, Qin YY, Yao YX, Zhang XG, Liang Y, et al: The clinical
significance and potential molecular mechanism of integrin subunit
beta 4 in laryngeal squamous cell carcinoma. Pathol Res Pract.
216:1527852020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar
|
|
10
|
Boelens MC, van den Berg A, Vogelzang I,
Wesseling J, Postma DS, Timens W and Groen HJ: Differential
expression and distribution of epithelial adhesion molecules in
non-small cell lung cancer and normal bronchus. J Clin Pathol.
60:608–614. 2007. View Article : Google Scholar
|
|
11
|
Chung J, Bachelder RE, Lipscomb EA, Shaw
LM and Mercurio AM: Integrin (alpha 6 beta 4) regulation of eIF-4E
activity and VEGF translation: A survival mechanism for carcinoma
cells. J Cell Biol. 158:165–174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Damhofer H, Medema JP, Veenstra VL, Badea
L, Popescu I, Roelink H and Bijlsma MF: Assessment of the stromal
contribution to Sonic Hedgehog-dependent pancreatic adenocarcinoma.
Mol Oncol. 7:1031–1042. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gan L, Meng J, Xu M, Liu M, Qi Y, Tan C,
Wang Y, Zhang P, Weng W, Sheng W, et al: Extracellular matrix
protein 1 promotes cell metastasis and glucose metabolism by
inducing integrin β4/FAK/SOX2/HIF-1α signaling pathway in gastric
cancer. Oncogene. 37:744–755. 2018. View Article : Google Scholar
|
|
14
|
Ni H, Dydensborg AB, Herring FE, Basora N,
Gagné D, Vachon PH and Beaulieu JF: Upregulation of a functional
form of the beta4 integrin subunit in colorectal cancers correlates
with c-Myc expression. Oncogene. 24:6820–6829. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang X, Wang J, Wang M, Xuan M, Han S, Li
C, Li M, Sun XF, Yu W and Zhao Z: ITGB4 as a novel serum diagnosis
biomarker and potential therapeutic target for colorectal cancer.
Cancer Med. 10:6823–6834. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li M, Jiang X, Wang G, Zhai C, Liu Y, Li
H, Zhang Y, Yu W and Zhao Z: ITGB4 is a novel prognostic factor in
colon cancer. J Cancer. 10:5223–5233. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kechagia JZ, Ivaska J and Roca-Cusachs P:
Integrins as biomechanical sensors of the microenvironment. Nat Rev
Mol Cell Biol. 20:457–473. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
García-Ortiz A and Serrador JM: ERM
proteins at the crossroad of leukocyte polarization, migration and
intercellular adhesion. Int J Mol Sci. 21:15022020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Bretscher A: Fimbrin is a cytoskeletal
protein that crosslinks F-actin in vitro. Proc Natl Acad Sci USA.
78:6849–6853. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lugini L, Lozupone F, Matarrese P, Funaro
C, Luciani F, Malorni W, Rivoltini L, Castelli C, Tinari A, Piris
A, et al: Potent phagocytic activity discriminates metastatic and
primary human malignant melanomas: A key role of ezrin. Lab Invest.
83:1555–1567. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim C, Shin E, Hong S, Chon HJ, Kim HR,
Ahn JR, Hong MH, Yang WI, Roh JK and Rha SY: Clinical value of
ezrin expression in primary osteosarcoma. Cancer Res Treat.
41:138–144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Krishnan K, Bruce B, Hewitt S, Thomas D,
Khanna C and Helman LJ: Ezrin mediates growth and survival in
Ewing's sarcoma through the AKT/mTOR, but not the MAPK, signaling
pathway. Clin Exp Metastasis. 23:227–236. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Q, Ye Z, Zhang Y and Liao F: The
expression and clinical significance of Ezrin and MMP-9 in
colorectal cancer tissue. Altern Ther Health Med. Jun 28–2024.Epub
ahead of print.
|
|
25
|
Liang F, Wang Y, Shi L and Zhang J:
Association of Ezrin expression with the progression and prognosis
of gastrointestinal cancer: A meta-analysis. Oncotarget.
8:93186–93195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tejeda-Muñoz N and Mei KC: Wnt signaling
in cell adhesion, development, and colon cancer. IUBMB Life.
76:383–396. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao H, Ming T, Tang S, Ren S, Yang H, Liu
M, Tao Q and Xu H: Wnt signaling in colorectal cancer: Pathogenic
role and therapeutic target. Mol Cancer. 21:1442022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang J, Li J, Yao W, Wang W, Shi B, Yuan
F, Dong J and Zhang H: FOXC1 Negatively Regulates DKK1 expression
to promote gastric cancer cell proliferation through activation of
wnt signaling pathway. Front Cell Dev Biol. 9:6626242021.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Wang J, Long H, Lin W, Wang H, Chen
Y, Yuan Q and Li X: circCDYL2, overexpressed in highly migratory
colorectal cancer cells, promotes migration by binding to Ezrin.
Front Oncol. 11:7160732021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song Y, Ma X, Zhang M, Wang M, Wang G, Ye
Y and Xia W: Ezrin mediates invasion and metastasis in
tumorigenesis: A review. Front Cell Dev Biol. 8:5888012020.
View Article : Google Scholar : PubMed/NCBI
|