Introduction
According to a 2024 report by The International
Agency for Research on Cancer, gastric cancer (GC) ranked as the
fifth most commonly diagnosed malignancy globally in 2022,
accounting for ~968,000 new cases (4.9% of total cancer cases) and
660,000 deaths (6.8% of total cancer mortality) (1,2).
This positions GC as a major public health challenge requiring
urgent attention worldwide. Although improvements in healthcare and
social development have contributed to a gradual reduction in the
overall burden of GC, countries with large populations such as
China and India continue to experience a high incidence of
early-stage GC (1,2). The incidence of GC exhibits distinct
geographical variation, with age-standardized rates in Asia, Latin
America and Europe being notably higher than those in other regions
(3), indicating substantial
disease risk. The high morbidity and mortality rates associated
with GC are largely attributed to limitations in early diagnosis,
with the majority of patients diagnosed at locally advanced or
metastatic stages, posing a severe threat to both survival and
quality of life (4). In recent
years, the emergence of novel targeted agents and immunotherapies
has partially enhanced clinical management strategies. However,
their efficacy is influenced by factors including patient
pathology, disease stage, clinical symptoms and performance status
(5). The tumor microenvironment
(TME), comprising cellular components such as cancer-associated
fibroblasts and endothelial cells, as well as non-cellular
components including cytokines, chemokines and the extracellular
matrix (ECM), plays a pivotal role in tumor progression; it
promotes ECM remodeling, epithelial-mesenchymal transition and
sustained tumor proliferation, while also fostering an
immunosuppressive milieu. Consequently, a considerable proportion
of patients exhibit a poor response to immunotherapy and are prone
to developing treatment resistance (6). Therefore, enhancing GC prevention
and treatment capabilities, along with continuously refining
clinical strategies, represents a critical and pressing challenge
in the fields of gastroenterology and oncology worldwide.
Tumor tissues exhibit metabolic pathways distinct
from those in normal tissues. Normal cells in the quiescent phase
maintain basal metabolic activity with limited uptake of nutrients
such as glucose and amino acids to support essential life
activities, and during proliferation moderately upregulate nutrient
transporters to facilitate orderly synthesis of genetic materials
in a tightly regulated manner. However, tumor cells display
markedly enhanced nutrient uptake to sustain their high energy and
biosynthetic demands (7). This
metabolic reprogramming in the TME is characterized by dysregulated
metabolic fluxes and serves as a critical basis for tumor
proliferation, metastasis, invasion and therapy resistance,
particularly in digestive system cancer types (8). In the 1920s, Professor Otto Warburg
observed that under aerobic conditions, the ATP yield relative to
oxygen consumption in oxidative phosphorylation (OXPHOS) is ~1:7.
By contrast, tumor cells exhibit notable metabolic plasticity
during proliferation and metastasis. They activate cytokines and
metabolism-related signaling pathways, adapt their metabolic
routes, induce mitochondrial abnormalities and remodel the entire
metabolic network. These alterations enhance energy production
efficiency, facilitate immune evasion, acidify the TME and
collectively promote rapid tumor growth and dissemination (9,10).
Despite these insights, current understanding of tumor metabolic
reprogramming remains insufficient to meet the precision
requirements for GC prevention and treatment. Recent advances in
microarray and high-throughput sequencing technologies have
accelerated the recognition of epigenetic modifications (11), initially conceptualized by
Professor Conrad Waddington. These heritable yet reversible
chemical modifications, affecting DNA, histones or RNA, regulate
gene expression without altering the DNA sequence and thereby
influence cellular phenotypes (11).
Over the past two decades, non-coding RNAs (ncRNAs),
once considered transcriptional 'noise', have been demonstrated to
participate in the development and progression of multiple cancer
types through their regulatory modifications (12). ncRNAs represent a diverse class of
RNAs that lack open reading frames and do not encode proteins.
Notably, up to 98% of RNAs in numerous organisms are non-coding,
highlighting their crucial functional roles at the RNA level within
the transcriptome (13). Among
these, circular RNAs (circRNAs) have emerged as key regulatory
ncRNA molecules. They influence the expression of metabolic enzymes
and transporters, encode short peptides in certain cases and
regulate gene expression at transcriptional and
post-transcriptional levels, thereby participating in various
metabolic processes in GC (14).
The present review focuses on circRNAs as potential key regulatory
nodes in metabolic reprogramming, which may reflect tumor metabolic
activity and drug sensitivity. By integrating circRNAs into the
metabolic remodeling network of GC, the present review aims to
contribute to the development of precise targeted therapies for
GC.
Biological characteristics of circRNA
Linear RNAs are processed by splicing out introns,
connecting exons and adding a 5' cap and a 3' poly(A) tail
(Fig. 1A). By contrast, circRNAs
are generated from pre-mRNAs through back-splicing mediated by
cis-acting elements and specific trans-acting factors, resulting in
exonic circRNAs or exon-intron circRNAs when introns are retained.
circular intronic RNA are derived from intronic lariats that escape
debranching and undergo cyclization (15) (Fig.
1B-D). circRNAs lack a 5' m7GpppN cap and a 3'
poly(A) tail, possess a covalently closed non-linear structure, are
resistant to exonuclease digestion and have a longer half-life in
blood, making them more stable for detection in tissues and body
fluids (16). Efficient
identification and enrichment of circRNAs can be achieved through
two principal strategies: i) Enzymatic digestion of linear RNAs
using RNase R, followed by high-throughput sequencing of the
resistant circRNA fraction; and ii) immunoprecipitation or
affinity-based depletion of linear RNAs using antibodies against
the 5' cap (such as anti-m7G) or oligo(dT) probes
targeting the poly(A) tail (17,18). Notably, certain circRNAs exhibit
higher abundance than their linear mRNA counterparts derived from
the same parental gene (19).
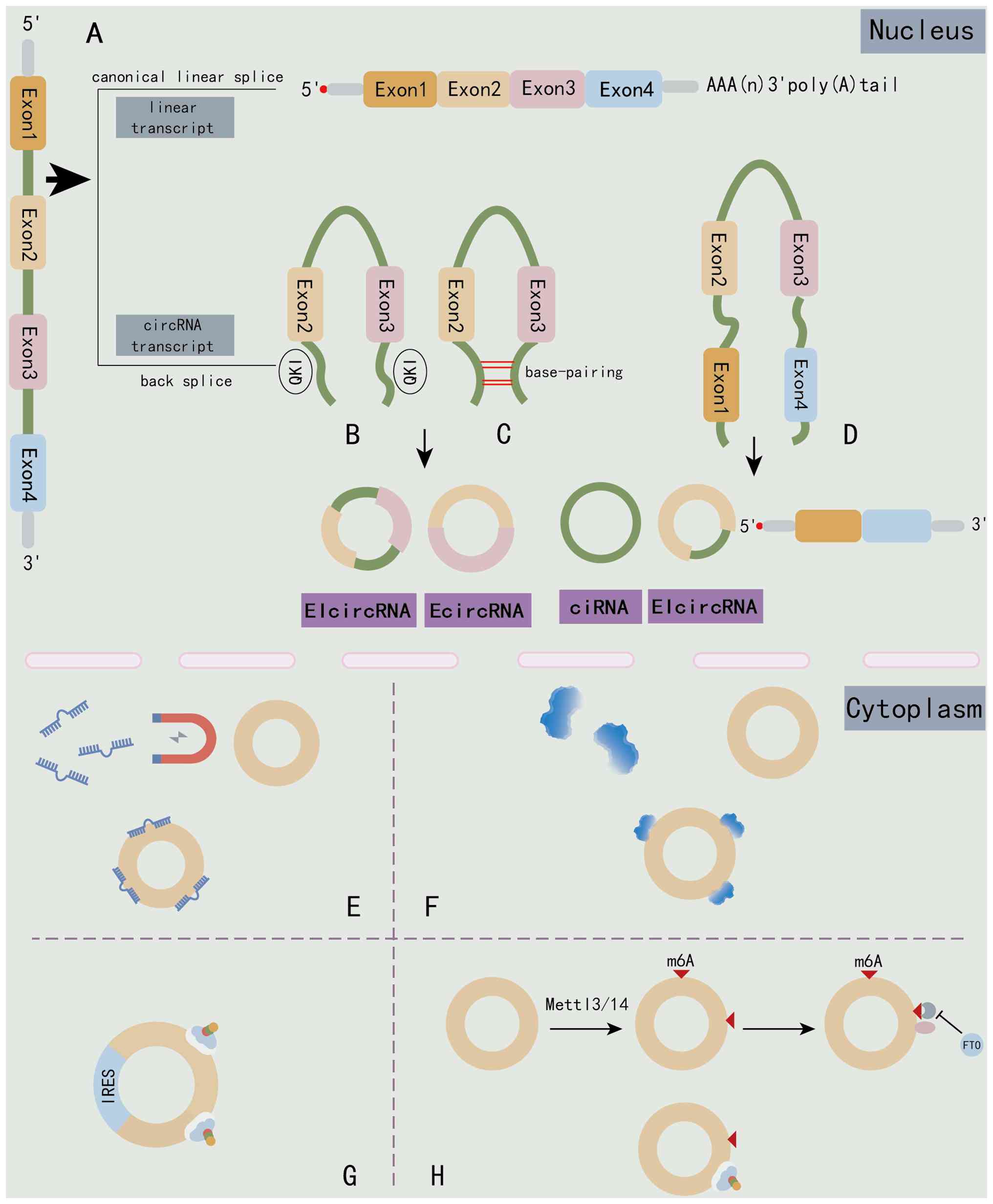 | Figure 1Schematic illustration of circRNA
biogenesis and functions in the nucleus and cytoplasm. (A)
Canonical linear splicing of precursor mRNA produces a linear
transcript containing a 5' cap and 3' poly(A) tail, whereas
back-splicing generates circRNA molecules. (B) EIciRNAs: Formed
when back-splicing retains an intron between the circularizing
exons, mediated by QKI and facilitated by base-pairing
interactions. (C) EcircRNAs: Formed by back-splicing of exons (such
as Exon2 and Exon3) with the introns removed. This results in a
circular molecule composed entirely of exons. (D) ciRNAs: Generated
from introns that escape debranching and form a circle (shown here
as Exon1-derived ciRNA). The diagram also illustrates an EIciRNA
variant comprising Exon1 directly spliced to Exon3/4. circRNAs (E)
acting as miRNA sponges, (F) interacting with RBPs, (G) serving as
templates for cap-independent translation via IRESs and (H)
undergoing m6A-dependent translation mediated by
Mettl3/14 and eIF4G2. circRNA, circular RNA; EIcircRNAs,
exon-intron circRNAs; EcircRNAs, exonic circRNAs; ciRNAs, intronic
circRNAs; RBPs, RNA-binding proteins; IRESs, internal ribosome
entry sites; m6A, N6-methyladenosine; Mettl,
methyltransferase-like; QKI, quaking; FTO, complex and fat mass and
obesity-associated protein. Created in Adobe Illustrator. |
The biogenesis of circRNAs is facilitated by
structural motifs in pre-mRNA introns. Alu repeat elements, which
are enriched in intronic regions, contain inverted complementary
sequences that enable the formation of intramolecular
double-stranded RNA (dsRNA) structures. These structures bring
distal splice sites into spatial proximity, creating a favorable
configuration for the spliceosome to perform backsplicing (20,21). Additionally, specific RNA-binding
proteins (RBPs) recognize cis-elements in pre-mRNAs and promote
circRNA circularization (22).
Quaking (QKI), a member of the STAR family of KH domain-containing
proteins, dynamically regulates ~1/3 of all circRNAs (23). The N-terminal Qua1 domain of QKI
mediates dimerization, enabling the protein to recognize and bind
specific Quaking response elements within introns. This facilitates
the alignment of a downstream 5' splice site with an upstream 3'
splice site, driving efficient and specific circRNA biogenesis via
backsplicing (24).
Adenosine deaminase acting on RNA 1 (ADAR1), which
catalyzes adenosine-to-inosine editing, also plays a critical role
in circRNA formation, particularly near regions with inverted
complementary matches (25).
Notably, ADAR1 knockdown in 293 cells leads to upregulation of
circRNA levels (26). This
phenomenon may be attributed to ADAR1-mediated destabilization of
intramolecular dsRNA structures. In the absence of editing, stable
dsRNA bridges bring the upstream 3' and downstream 5' splice sites
into proximity, facilitating backsplicing. ADAR1 expression
disrupts these dsRNA interactions, preventing spliceosome
recognition and efficient circRNA production.
circRNAs can participate in metabolic regulation by
sponging microRNAs (miRNAs/miRs) (Fig. 1E), interacting with RBPs (Fig. 1F) and driving cap-independent
translation through internal ribosome entry sites (IRESs) (Fig. 1G) or N6-methyladenosine
(m6A) modification co-regulated by the
methyltransferase-like (Mettl)3)/Mettl14 complex and fat mass and
obesity-associated protein (FTO) (Fig. 1H).
Metabolic reprogramming in GC
Glycolysis
Under oxygen-sufficient conditions, normal cells
primarily satisfy their energy requirements through the sequential
biological processes of 'glucose uptake → glycolysis → pyruvate
entry into the tricarboxylic acid (TCA) cycle → OXPHOS'. By
contrast, tumor cells preferentially utilize aerobic glycolysis,
consuming large amounts of glucose and converting pyruvate into
lactate, yielding a net gain of only two ATP molecules per glucose
molecule (27). This metabolic
shift is partly driven by mitochondrial reprogramming, wherein
tumor cells maintain TCA cycle activity to support biosynthetic
precursor synthesis, partially compensating for the reduced ATP
yield from OXPHOS (28).
Glycolytic reprogramming occurs predominantly within
the cytosol. In highly migratory tumor cells, glycolytic enzymes
are recruited to the cell cortex, where they propagate as
self-organizing waves along the membrane/cortex and interact with
actin filaments (29). Key
regulatory enzymes, including hexokinase (HK), pyruvate
dehydrogenase kinase (PDK), phosphofructokinase-1 (PFK1) and the
pyruvate kinase M2 (PKM2) isoform, orchestrate a series of
catalytic reactions (30). These
protein-protein interactions among metabolic enzymes lead to
altered levels of glucose-metabolizing proteins and glycolytic
intermediates such as fructose, glyceraldehyde and pyruvate. Most
pyruvate is subsequently converted into lactate, which is exported
extracellularly via monocarboxylate transporters (MCTs).
Extracellular lactate accumulation further drives histone lysine
lactylation, influencing transcriptional regulation,
post-transcriptional modifications and post-translational processes
in tumor cells (31). By
accelerating glycolytic flux, tumor cells rapidly generate energy,
thereby promoting proliferation, invasion, metastasis and therapy
resistance, a mechanism critically involved in GC pathogenesis
(32,33). Consequently, targeting key
glycolytic enzymes represents a promising strategy to suppress the
Warburg effect and impede GC progression (Fig. 2A).
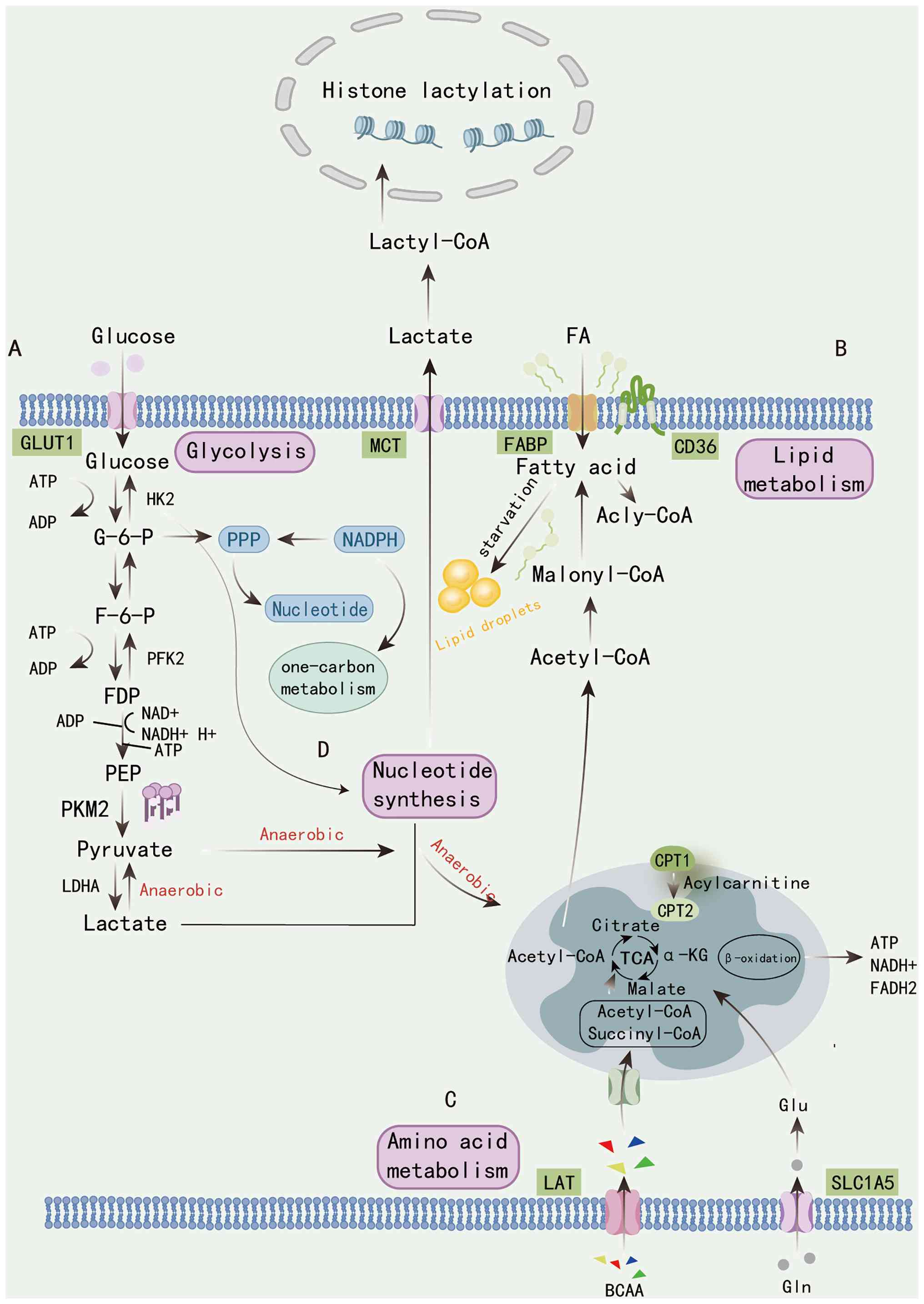 | Figure 2Overview of the metabolic pathways
linking glycolysis, lipid metabolism, amino acid metabolism and
nucleotide metabolism. (A) Glucose uptake via GLUT1 triggers
glycolysis, producing pyruvate and lactate through the sequential
actions of HK2, PFK2, PKM2 and LDHA. Under anaerobic conditions,
lactate is exported and can re-enter cells via MCTs, serving as a
substrate for lactyl-CoA formation that drives histone lactylation.
(B) FAs imported through CD36 and FABP undergo activation to
acyl-CoA, conversion to malonyl-CoA and oxidation via mitochondrial
CPT1/2, producing ATP, NADH and FADH2 through the TCA
cycle. Lipid droplets act as energy reservoirs during starvation.
(C) Amino acids, including BCAAs and glutamine transported by LAT
and SLC1A5, feed into the TCA cycle to replenish intermediates such
as α-KG. (D) Parallel to glycolysis, the PPP generates NADPH and
ribose intermediates to support nucleotide synthesis and one-carbon
metabolism. Together, these integrated metabolic routes coordinate
carbon flux and epigenetic regulation through lactate-derived
histone modifications. HK2, hexokinase 2; PFK2,
phosphofructokinase-2; PKM2, pyruvate kinase M2; MCTs,
monocarboxylate transporters; FA, fatty acid; CPT, carnitine
palmitoyltransferase; TCA, tricarboxylic acid; PPP, pentose
phosphate pathway; LAT, L-type amino acid transporter; SLC1A5,
solute carrier family 1 member 5; α-KG, α-ketoglutarate; GLUT1,
glucose transporter 1; LDHA, lactate dehydrogenase A; FABP, fatty
acid-binding protein; BCAAs, branched-chain amino acids; G-6-P,
glucose-6-phosphate; F-6-P, fructose-6-phosphate; FDP,
fructose-1,6-bisphosphate; PEP, phosphoenolpyruvate; Glu,
glutamate; Gln, glutamine. Created in Adobe Illustrator. |
The HK family comprises four isoforms (HK1-4), each
encoded by distinct genes and exhibiting unique functional and
distribution profiles. Among these, HK2 is upregulated in most
tumor cells, including GC cells, and serves as the key
rate-limiting enzyme catalyzing the first committed step of
glycolysis (34). HK2 transfers a
phosphate group from ATP to glucose, generating glucose-6-phosphate
(G6P), which provides a critical metabolic intermediate for
downstream reactions (35). PFK1,
a major regulatory node in the glycolytic pathway, acts as another
essential rate-limiting enzyme that directs G6P flux into
glycolysis; it catalyzes the conversion of fructose-6-phosphate to
fructose-1,6-bisphosphate. Under conditions of high energy demand
or hypoxia, PFK1 activity is enhanced, accelerating G6P consumption
and favoring glycolytic ATP production (36). The PDK family also consists of
four isoforms (PDK1-4). Elevated expression of PDK isoforms in GC
cells promotes S-phase entry and serves as an independent
prognostic factor in patients with GC (37). PDK plays a decisive role in
directing pyruvate metabolism, where it phosphorylates and
inactivates pyruvate dehydrogenase, thereby preventing the
conversion of pyruvate to acetyl-CoA and entry into the TCA cycle.
This suppression of mitochondrial OXPHOS shifts metabolism toward
anaerobic glycolysis, leading to lactate accumulation.
Concomitantly, altered histone acetylation patterns and suppressed
programmed cell death-ligand 1 (PD-L1) expression contribute to
immune evasion, further enhancing the pro-tumorigenic effects in GC
(38). Pyruvate kinase, the
terminal rate-limiting enzyme of glycolysis, transfers a
high-energy phosphate group from phosphoenolpyruvate to ADP,
generating pyruvate. The metabolic fate of pyruvate is closely
linked to the oligomeric state of the predominant tumor isoform
PKM2. In its tetrameric form, PKM2 exhibits high catalytic
activity, facilitating pyruvate conversion to acetyl-CoA and
supporting sustained energy production via OXPHOS. By contrast, the
dimeric form of PKM2 displays reduced activity, leading to the
accumulation of glycolytic intermediates such as phosphoribosyl
pyrophosphate and 3-phosphoglycerate. This metabolic configuration
prioritizes biosynthetic precursor supply, thereby supporting rapid
tumor proliferation and metastasis (39,40).
Lipid metabolism
Fatty acids (FAs), essential components of various
cellular structures, play critical roles in biological processes
such as membrane biogenesis and energy provision (41). Mitochondrial β-oxidation serves as
the core pathway for FA breakdown and energy production. However,
long-chain FAs cannot freely traverse the mitochondrial membrane.
Instead, they are first converted to acyl-CoA in the cytosol and
then transported into the mitochondrial matrix via the carnitine
palmitoyltransferase (CPT) system located on the inner
mitochondrial membrane (42).
Subsequently, through repeated cycles of dehydrogenation, hydration
and thiolysis, FAs are degraded to generate acetyl-CoA, NADH and
NADPH, which enter the TCA cycle to produce ATP (42). To rapidly adapt to hypoxic and
nutrient-deprived microenvironments, tumor cells reprogram lipid
metabolism by modulating FA uptake, synthesis, storage and
β-oxidation. They utilize lipids for membrane construction, to
support progression, and upregulate factors such as CD36 in
regulatory T cells and CD8+ T cells. This enhances lipid
uptake and alters immune cell recruitment, activation and function,
thereby facilitating immune evasion and accelerating tumor
proliferation and metastasis (43,44) (Fig.
2B).
The transport of long-chain FAs into cells is a
critical step in lipid metabolic reprogramming, primarily mediated
by transport proteins. The transmembrane glycoprotein CD36,
expressed on various cells within the TME, including endothelial,
stromal and immune cells, facilitates long-chain FA uptake via
endocytosis (45). Internalized
FAs can be directed to lipid droplets for storage or undergo
β-oxidation. This process involves activation of the Lck/Yes novel
tyrosine kinase, which phosphorylates and inactivates zinc finger
DHHC-type palmitoyltransferase 5 at tyrosine 91. Subsequently,
acyl-protein thioesterase 1 mediates CD36 depalmitoylation,
initiating a new cycle of CD36-mediated endocytosis and FA
transport. This dynamic palmitoylation-depalmitoylation cycle of
CD36 forms a closed loop crucial for remodeling the tumor lipidome
and generating oncogenic lipid species, thereby promoting tumor
growth and progression (46).
Furthermore, FAs can upregulate O-GlcNAcylation, which in
turn modifies CD36 and enhances its transcriptional expression.
This positive feedback loop increases FA uptake in GC cells,
accelerates metastasis and is closely associated with poor
prognosis in patients with GC (47). While transporter-dependent
pathways are important, they are not the sole mechanism for
long-chain FA uptake. The acidic TME can neutralize negatively
charged FAs, promoting their protonation and passive diffusion into
cells. This alternative pathway further drives FA accumulation and
lipid droplet formation, supporting cancer cell metabolism and
rapid proliferation (48).
The CPT system, a key regulator of FA β-oxidation,
is central to lipid metabolic reprogramming. It consists of two
isoforms: i) CPT1, located on the outer mitochondrial membrane and
serving as the primary rate-limiting enzyme for FA translocation
into mitochondria (49); and ii)
CPT2, residing on the inner mitochondrial membrane, which
facilitates the entry of FAs into the mitochondrial matrix for
β-oxidation (50). In GC, CPT1A
is highly expressed and interacts with S100A10, promoting its
succinylation at lysine 47 (51).
CPT1A expression also correlates with CD44, a key regulator of
cancer metastasis. CD44 activates intercellular signaling pathways
that target CPT1A to modulate FA oxidation (FAO), thereby enhancing
the invasive and metastatic potential of GC cells (51,52). Similarly, CPT2 is notably
upregulated in GC cells (53).
Nuclear factor of activated T cells c3, a member of the NFAT
transcription factor family, can be activated by reactive oxygen
species (ROS) and translocate to the nucleus. There, it binds to
the CPT2 promoter to enhance its transcription, elevating
β-oxidation levels. This increase in FA catabolism raises the
NADPH/NADP+ ratio, protecting GC cells from ROS-induced
damage and maintaining cellular homeostasis (53).
Amino acid metabolism
Through metabolic reprogramming of amino acid
networks, tumor cells continuously adapt to survival pressures.
Glutamine (Gln), the most abundant free amino acid in systemic
circulation, plays a particularly critical role. As tumor cells are
unable to synthesize Gln de novo, they compete with normal
cells for exogenous Gln, developing a metabolic dependency to
sustain proliferation and metastasis (54). Given its hydrophilic nature, Gln
requires specific solute carriers, such as solute carrier family 1
member 5 (SLC1A5; also known as ASCT2), to traverse the plasma
membrane. Once intracellular, Gln is sequentially deamidated by
glutaminase (GLS) and glutamate dehydrogenase to produce glutamate
(Glu), which is further converted to α-ketoglutarate (α-KG). This
metabolite replenishes the TCA cycle, provides biosynthetic
precursors, enhances mitochondrial activity and ultimately fuels
cancer proliferation (55-57).
Due to their rapid energy consumption, tumor cells
frequently face glucose starvation in the TME. To address this,
they reconfigure intrinsic metabolic pathways to maintain NADPH
homeostasis, thereby meeting the high energy demands and preserving
the NADPH/NADP+ balance (58). In GC, upregulation of malic enzyme
1 (ME1) helps replenish intracellular NADPH pools and maintain
redox homeostasis. Conversely, ME1 knockdown depletes NADPH, lowers
the glutathione (GSH)/GSH disulfide ratio, elevates ROS,
exacerbates oxidative stress and promotes GC cell death (58,59). A portion of the Glu generated from
Gln in mitochondria is transported back to the cytosol, where it
serves as a substrate for GSH synthesis. Glu-cysteine ligase
catalyzes the conjugation of Glu and cysteine to form
γ-glutamylcysteine, which is subsequently combined with glycine by
GSH synthase to produce GSH. This pathway assists tumor cells in
countering oxidative stress (60-62) and modulates host immune
surveillance and suppression. When Gln is extensively depleted by
tumors, impaired GSH synthesis triggers Fas/CD95-mediated
apoptosis. However, concurrent upregulation of PD-L1 expression can
suppress CD8+ T cell antitumor activity. Thus,
simultaneously targeting Gln deprivation and PD-L1 activity, by
blocking PD-L1/PD-1 interactions, presents a promising clinical
strategy to alleviate immunosuppression in the TME (63). The oncogenic transcription factor
c-Myc further supports Gln metabolism by binding to E-box sequences
in target gene promoters. This upregulates GLS expression, enhances
Gln uptake and catabolism, supplies TCA cycle intermediates,
promotes nucleotide synthesis and sustains the malignant
proliferative phenotype of tumors (64,65) (Fig.
2C).
In addition to Gln, other amino acids contribute to
metabolic remodeling in GC, enabling tumor cells to compete for
energy resources and support disease progression. Branched-chain
amino acids (BCAAs), such as leucine, isoleucine and valine, are
non-polar aliphatic amino acids characterized by carboxyl groups,
amino groups and isopropyl side chains. They can be directly
incorporated into proteins or catabolized into various metabolites
to fuel related metabolic pathways (66). BCAAs enter cells via L-type amino
acid transporters (LATs) and are shuttled into mitochondria by
SLC25A44. Within mitochondria, they are transaminated to
branched-chain α-keto acids (BCKAs), which subsequently transfer
amino groups to α-KG. BCKAs are further metabolized to acetyl-CoA
and succinyl-CoA, while generating Glu. The resulting metabolites
ultimately enter the TCA cycle to support energy production
(67).
In addition, an inter-amino acid compensatory
mechanism exists. Under Gln-deficient conditions, elevated
expression of the PPM1K protein promotes BCAA catabolism to sustain
tumor cell survival (68).
Previous studies suggest that elevated circulating BCAA levels may
be associated with several human cancer types (69,70). However, a large-scale prospective
cohort study by Yu et al (71) reached a contrasting conclusion,
indicating that higher BCAA levels correlate with a reduced risk of
GC, particularly among elderly populations. Given this dual role of
BCAAs in tumor development, further investigation is needed to
determine whether these context-dependent effects stem from
distinct components within BCAA catabolic pathways.
Nucleotide metabolism
Nucleotides, composed of a nucleoside and a
phosphate group, are essential for DNA and RNA synthesis, enzyme
regulation, substrate activation and various metabolic processes in
cells. Their synthesis is energy-intensive and requires carbon and
nitrogen donors for support (72). This process relies on the
cytosolic one-carbon (1C) metabolic pathway. To meet the demands of
proliferation and metastasis, tumor cells upregulate the expression
of enzymes related to 1C metabolism, altering their metabolic
patterns to support the de novo synthesis of purines and
pyrimidines (73). Pyrimidines
and purines are the core structural components of nucleotides, and
nucleotide synthesis involves two pathways: i) De novo
synthesis; and ii) salvage pathways (74). In the de novo synthesis
process, precursor molecules generated by various metabolic
pathways are utilized. For example, Gln is deaminated by GLS to
produce Glu, which is then converted by glutamate dehydrogenase,
releasing ammonia in the process. In glucose metabolism, the
pentose phosphate pathway produces G6P, which then undergoes
dehydrogenation, decarboxylation and isomerization reactions to
form ribose-5-phosphate. Together, these intermediates provide
essential raw materials for nucleotide synthesis.
The salvage pathway allows cells to rapidly generate
nucleoside monophosphates from free nucleobases or nucleosides
through phosphorylation or phosphoribosyl transfer reactions. This
process avoids base wastage, is relatively energy-efficient and
helps replenish the nucleotide pool (75). Normal cells primarily rely on the
salvage pathway for nucleotide synthesis. However, the aggressive
behavior of tumor cells depends on higher nucleotide synthesis and
metabolic activity, requiring faster and greater acquisition of
nucleotides to support rapid proliferation, drug resistance, immune
evasion and metastasis. Therefore, tumor cells mainly utilize the
de novo synthesis pathway for nucleotide production
(76). U2AF homology motif kinase
1 (UHMK1) is a nuclear-localized serine/threonine kinase and acts
as an effective oncogenic factor. In GC, UHMK1 expression is
upregulated; it can phosphorylate nuclear receptor coactivator 3 at
serine 1062 and threonine 1067, promoting its interaction with
activating transcription factor 4 (ATF4) and inducing the nuclear
translocation of ATF4. This activates transcriptional activity,
mediates oligomeric nucleotide synthesis pathways and
simultaneously enhances the de novo purine synthesis
pathway, providing a material basis for GC cell proliferation and
driving GC progression (77,78) (Fig.
2D).
Cells can secrete nucleotides and nucleosides to
initiate paracrine and autocrine signaling, thereby driving
cellular metabolic networks, regulating immune responses and
influencing the development and progression of tumor cells
(79). Due to genomic mutations,
the expression level of major histocompatibility complex (MHC)
class I polypeptide-related sequence A (MICA) increases in tumors.
MICA binds to the natural killer group 2 member D receptor, thereby
triggering NK cell-mediated immune surveillance. This process is
associated with the high levels of nucleotide metabolism observed
in tumor cells (80,81). A study found that certain
nucleotide metabolism-related genes are associated with levels of
immune cell infiltration (82).
Patients with GC exhibiting high nucleotide metabolism have poorer
overall survival, disease-specific survival, disease-free interval
and progression-free interval. Additionally, these patients show
increased expression of immunosuppressive factors and MHC molecules
(82).
Adenosine can receive a phosphate group under the
catalysis of adenosine kinase, which is an important pathway for
nucleotide synthesis. Conversely, nucleotides can be
dephosphorylated by phosphatases to generate adenosine; this
bidirectional conversion is a key node in intracellular purine
metabolism (83). Adenosine
receptors are present on the surface of various cell types
including human dendritic cells (DCs), which mainly express A1 and
A3 receptors (84). Adenosine can
mediate the migration of DCs to lymphoid organs, promote antigen
presentation and T cell activation, thereby driving immune
regulation (85,86). Additionally, regulatory B cells
can regulate their own function through adenosine signaling derived
from ATP enzymatic degradation, maintaining an immunosuppressive
cell phenotype and inhibiting T cell activation, thus participating
in the regulation of adaptive immunity. Excessive adenosine
signaling can impede immune surveillance and thereby promote tumor
progression (84).
Core mechanisms of circRNA in regulating
metabolic reprogramming in GC
Non-coding mechanism: Acting as a
'molecular sponge' to competitively bind miRNA
Early transcriptome studies mainly focused on the
competing endogenous RNA (ceRNA) interactions between mRNA and long
ncRNA. With advances in high-throughput sequencing and RNA
interaction analysis technologies, it was discovered that circRNAs
retain only the circularized regions of exons, introns or
exon-intron sequences, and naturally lack a 5' cap and 3' poly(A)
tail. This makes them more stable within the ceRNA network and
enables them to have specific regulatory functions (87). circRNAs can bind to miRNA through
sequence-complementary miRNA response elements, competitively
inhibiting the degradation or translational repression of target
mRNAs by miRNAs. Therefore, circRNAs regulate the expression level
of target gene transcripts and are extensively involved in various
cellular biological processes (88).
circDNMT1 can act as a molecular sponge for
miR-576-3p, thereby relieving its inhibitory effect on the
hypoxia-inducible factor 1-α (HIF-1α) 3' untranslated region
(3'UTR). This accelerates glucose uptake in GC cells, increases
lactate production, suppresses pyruvate and ATP generation and
promotes tumor progression (89).
Similarly, inhibition of circSLAMF6 can enhance the targeted
regulation of myosin heavy chain 9 (MYH9) by miR-204-5p, thereby
weakening the glycolytic capacity of GC cells under hypoxia and
further suppressing tumor metastasis and invasion (90). As shown in Table I, numerous circRNAs have been
reported to participate in GC glycolysis via the ceRNA network in
previous years. The majority of circRNAs linked to GC are
upregulated and act as miRNA sponges to abolish the inhibition of
key glycolytic enzymes or transcription factors, consequently
enhancing glucose uptake and lactate production as well as
promoting GC proliferation, migration and invasion.
 | Table IThe potential mechanism of circRNA in
regulating metabolic reprogramming of gastric cancer. |
Table I
The potential mechanism of circRNA in
regulating metabolic reprogramming of gastric cancer.
A, ceRNA
|
|---|
| Authors, year | circRNAs | Expression | Target | Metabolic
mechanism | Effect | Function | (Refs.) |
|---|
| Zhou et al,
2022 | circ_0006089 | Up |
miR-361-3p/TGFB1 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, HK2↑ | Proliferation,
migration | (175) |
| Jiang et al,
2022 | circ_0067514 | Down |
miR-654-3p/LATS2 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, HK2↑, LDHA↑ | Proliferation,
invasion | (176) |
| Shen et al,
2022 | circZNF131 | Up |
miR-186-5p/PFKFB2 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑, GLUT1↑ | Migration,
invasion | (177) |
| Qian et al,
2025 | circ_0001756 | Up |
miR-185-3p/PGK1 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑, LDHA↑, GLUT1↑ | Proliferation,
migration, invasion | (100) |
| Yang et al,
2024 | circFLNA | Up | miR-1200/SOX5 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑ | Proliferation | (178) |
| Wang et al,
2021 | circBFAR | Up |
miR-513a-3p/HK2 | Glycolysis | Lactate↑, ECAR↑,
OCR↓, HK2↑ | Proliferation | (179) |
| Zheng et al,
2023 | circRPS19 | Up |
miR-125a-5p/USP7 | Glycolysis | Glucose
absorption↑, lactate↑, ECAR↑, OCR↓, HK2↑ | Proliferation | (180) |
| Chen et al,
2020 | circ_0032821 | Up |
miR-1236-3p/HMGB1 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, ECAR↑, OCR↓ | Proliferation,
migration, invasion | (181) |
| Liu et al,
2022 | circ_0009910 | Up |
miR-361-3p/SNRPA | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑, PKM2↑ | Proliferation,
migration, invasion | (182) |
| Liu et al,
2020 | circ-NRIP1 | Up |
miR-186-5p/MYH9 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, HK2↑, PKM2↑ | Proliferation,
migration | (169) |
| Zheng et al,
2024 | circKIAA1797 | Up | miR-4429/PBX3 | Glycolysis | Glucose
absorption↑, lactate↑, | Proliferation,
migration, invasion | (183) |
| Zhao et al,
2021 | circATP2B1 | Up |
miR-326-3p/miR-330-5p/PKM2 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, GLUT1↑, GLUT3↑, LDHA↑, PKM2↑ | Proliferation | (184) |
| Chen et al,
2021 | circC6orf132 | Up |
miR-873-5p/PRKAA1 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, HK2↑, GLUT1↑ | Proliferation,
migration, invasion | (185) |
| Yang et al,
2021 | circUBE2Q2 | Up |
miR-370-3p/STAT3 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, ECAR↑, HK2↑, PFK↑ | Proliferation,
migration, invasion | (186) |
| Liu et al,
2020 | circ-MAT2B | Up |
miR-515-5p/HIF-1α | Glycolysis | Glucose
absorption↑, lactate↑, HIF-1α↑ | Proliferation | (187) |
| Fang et al,
2020 | circSLAMF6 | Up |
miR-204-5p/MYH9 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑ | Migration,
invasion | (90) |
| Ou et al,
2025 | circ_0043256 | Up |
miR-593-5p/RRM2 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, HIF-1α↑, HK2↑, ENO1↑, LDHA↑, GLUT1↑,
PKM2↑ | Proliferation,
migration | (188) |
| Lu et al,
2021 | circVPS33B | Up |
miR-873-5p/HNRNPK | Glycolysis | Glucose
absorption↑, lactate↑, ECAR↑, OCR↓ | Proliferation,
migration, invasion | (189) |
| Qu et al,
2020 | circFLNA | Up | miR-646/PFKFB2 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, PFKFB2↑ | Proliferation,
migration | (190) |
| Lan et al,
2024 | circPRDM5 | Down |
miR-485-3p/GCNT4 | Glycolysis | ECAR↑, HK2↑ | Proliferation,
migration, invasion | (191) |
| Li et al,
2022 | circDNMT1 | Up |
miR-576-3p/HIF-1α | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, ECAR↑, OCR↓ | Proliferation,
migration, invasion | (89) |
| Lu et al,
2023 | circ_0000419 | Down | miR-300/RGMB | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑, GLUT1↑ | Migration,
invasion | (192) |
| Wu et al,
2021 | circ-RNF111 | Up |
miR-876-3p/KLF12 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑ | Migration,
invasion | (193) |
| Ji et al,
2022 | circ_000059 | Up | miR-1179/ANXA4 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑ | Proliferation,
migration, invasion | (194) |
| Xu et al,
2020 | circNRIP1 | Up |
miR-138-5p/HIF-1α | Glycolysis | Glucose
absorption↑, lactate↑, G6P↑, HIF-1α↑ | 5-FU
resistance | (170) |
| Pu et al,
2020 | circCUL3 | Up |
miR-515-5p/STAT3/HK2 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, ECAR↑ | Proliferation | (195) |
| Wang et al,
2023 | circ_0024107 | Up |
miR-5572/6855-5p/CPT1A | Lipid
metabolism | CPT1A↑, β oxidation
rate↑ | Migration,
invasion | (92) |
| Ye et al,
2020 | circB3GNTL1 | Up | miR-598 | Amino acid
metabolism | Gln↑, Glu↑,
α-KG↑ | Proliferation | (94) |
| Li et al,
2022 | circAKT3 | Up |
miR-515-5p/SLC1A5 | Amino acid
metabolism | Gln↑, SLC1A5↑ | Proliferation,
survival | (93) |
| Li et al,
2020 | circSFMBT2 | Up | miR-665 | Amino acid
metabolism | Gln↑, Glu↑,
α-KG↑ | Migration,
invasion | (95) |
|
| B, Encoding
peptide |
|
| Jiang et al,
2024 | circ_0008035 | Up | EXT1/PKM2 | Glycolysis | PKM2↑ | Proliferation | (113) |
| Lu et al,
2025 | circUBE2G1 | Down | ENO1 | Glycolysis | Glucose
absorption↑, lactate↑, ATP↑, pyruvate↑, ENO1↑ | Proliferation,
migration, invasion | (114) |
|
| C, RBP |
|
| Qian et al,
2025 | circ_0001756 | Up | PTBP1/PGK1 | Glycolysis | Glucose
absorption↑, lactate↑, HK2↑, LDHA↑, GLUT1↑ | Proliferation,
migration | (100) |
| Lin et al,
2025 | circTFRC | Up | SCD1 | Lipid
metabolism | SCD1↑ | Proliferation,
migration | (102) |
| Lin et al,
2024 | circ-CDK13 | Up | EIF4A3 | Amino acid
metabolism | Methionine↑ | DDP resistance | (105) |
|
| D, m6A
modification |
|
| Wu et al,
2024 | circFAM192A | Up | FTO | Amino acid
metabolism | FTO↑, SLC7A5↑ | Proliferation | (128) |
GC cells can recruit mesenchymal stem cells (MSCs)
by secreting microvesicles. Under the combined induction of immune
cells and Helicobacter pylori, these MSCs are transformed
into GC-MSCs, which retain mesenchymal lineage and stem cell
properties, possess multipotent differentiation potential and
strongly promote GC cell proliferation, migration and angiogenesis
(91). circ_0024107 derived from
GC-MSCs can act as a molecular sponge for miR-5572 and miR-6855-5p,
negatively regulating their expression, upregulating CPT1A mRNA and
protein expression, participating in FA transport and activating
FAO, thus promoting GC proliferation and metastasis (92). Additionally, circRNAs can enhance
the expression of amino acid metabolism-related transporters by
sponging miRNAs through sequence complementarity. A study has shown
that circAKT3 targets miR-515-5p, relieving its suppression of
SLC1A5 by binding to the SLC1A5 3'UTR, thereby increasing the
expression of the SLC1A5 transporter and enhancing Gln uptake in GC
cells, which in turn promotes malignant proliferation (93). circB3GNTL1 and circSFMBT2 can also
participate in amino acid metabolism in GC by sponging miR-598 and
miR-665, respectively (94,95). Although direct research focusing
on circRNA-mediated regulation of nucleotide metabolic
reprogramming in GC via the ceRNA network is limited, a study has
found that hsa_circ_0008434 in GC can sponge miR-6838-5p, relieving
its inhibition of ubiquitin-specific peptidase 9X (USP9X),
resulting in USP9X protein accumulation and promoting GC cell
proliferation, migration and invasion (96). USP9X, through multiple synergistic
mechanisms, regulates the stability of Gln synthetase, increases
nucleotide availability, supports nucleotide synthesis and stemness
maintenance in tumor cells, and participates in tumor growth and
proliferation (97).
Non-coding mechanism: Acting as a
'protein decoy' by binding to the functional domains of
proteins
The binding of circRNAs to RBPs relies on the
complementary pairing or spatial recognition between specific
binding sites within circRNA sequences and the RNA-binding domains
of RBPs. This interaction forms distinct circRNA-RBP complexes,
which regulate the activity, conformational changes and subcellular
localization of target proteins, thereby modulating circRNA
functions in tumor cells (98).
Additionally, circRNAs can regulate the expression of key metabolic
dependency enzymes or transporters and participate in multiple
signaling pathways to directly or indirectly influence metabolic
reprogramming in GC cells.
Polypyrimidine tract-binding protein 1 (PTBP1), a
shuttling protein between the cytoplasm and nucleus with distinct
functions in each compartment, is involved in tumor proliferation
and invasion (99). circ_0001756
binds to PTBP1, facilitating its interaction with phosphoglycerate
kinase 1 (PGK1) via the RRM1 domain. This interaction enhances the
stability of PGK1 mRNA, promotes its cytoplasmic expression and
participates in the first step of ATP production through the
glycolytic pathway, thereby contributing to GC initiation and
progression (100). Stearoyl-CoA
desaturase 1 (SCD1), a key rate-limiting enzyme that desaturates
saturated FAs to produce monounsaturated FAs, controls lipid
metabolic flux. Aberrant transcriptional regulation and epigenetic
modifications of SCD1 induce abnormal lipid accumulation, driving
tumor progression (101).
circTFRC directly binds to SCD1 mRNA, recruits ELAV-like RNA
binding protein 1 and interacts with the 3'UTR of SCD1 mRNA. This
protects SCD1 from nuclease degradation, enhances its stability and
sustains SCD1 mRNA translation. Consequently, this promotes
oncogenic lipid metabolic remodeling, thereby accelerating GC
progression (102).
Rapidly dividing tumor cells exhibit methionine
(Met) dependence due to their high demand for Met in processes such
as histone modification, nuclear division and GSH synthesis
(103,104). Met deprivation induces DNA
damage in tumor cells, leading to proliferation arrest. A study has
shown that eukaryotic initiation factor 4A3 (EIF4A3) binds to and
interacts with the flanking sequences downstream of circ-CDK13. Met
degradation downregulates EIF4A3 expression, restricting circ-CDK13
circularization, inhibiting GC cell proliferative activity,
increasing intracellular cisplatin concentrations and enhancing
chemosensitivity (105). The
oncogenic transcription factor myelocytomatosis oncogene (MYC) has
been extensively studied in metabolic contexts, with its expression
consistently elevated across all stages of intestinal-type GC
development (106,107). MYC provides sugar backbones for
purine and pyrimidine biosynthesis and activates phosphoribosyl
pyrophosphate synthetase 2 (PRPS2) via eukaryotic translation
initiation factor 4E. Through specific cis-regulatory elements
within its 5'UTR, PRPS2 couples protein and nucleotide biosynthesis
(108), enabling tumor cells to
reprogram nucleotide metabolic pathways to adapt to
nutrient-deprived environments. Research has confirmed that
circ-TNPO3 binds to the KH domain at the C-terminus of insulin-like
growth factor 2 mRNA-binding protein 3 (IGF2BP3), impairing
IGF2BP3-mediated stabilization of MYC mRNA and thereby inhibiting
GC proliferation and metastasis (109). Additionally, circ-hnRNPU can
also directly bind to non-POU domain-containing octamer-binding
protein (NONO), inducing the cytoplasmic accumulation of NONO,
thereby abolishing the activating effect of nuclear NONO on c-Myc
and downregulating c-Myc expression (110). c-Myc is well established as a
transcriptional activator of PRPS2, a rate-limiting enzyme in de
novo purine and pyrimidine synthesis (108). Whether circ-hnRNPU-mediated
c-Myc downregulation translates into altered nucleotide metabolic
flux in GC cells, however, has not been investigated. Direct
evidence linking circ-hnRNPU to changes in nucleotide synthesis
rates, PRPS2 activity, or intracellular metabolite levels is
currently absent, and this putative relationship awaits
experimental validation.
Coding mechanism: Encoding peptide
Conventional translation initiation typically
involves the recognition of the 5' cap by the small ribosomal
subunit and its associated initiation factors. However, circRNAs
lack this structure and cannot initiate translation through the
canonical pathway. IRESs, which are structured RNA elements capable
of directly recruiting the small ribosomal subunit and initiation
factors, enable a subset of circRNAs to undergo cap-independent
translation, yielding functional short peptides (111).
Emerging evidence indicates that peptides encoded by
endogenous circRNAs play potential roles in tumorigenesis (112). For example, circ_0008035 encodes
a novel peptide, EXT1-219aa, which suppresses EXT1 phosphorylation.
This promotes the nuclear localization of PKM2 and reduces its
EXT1-dependent glycosylation, thereby enhancing pyruvate metabolism
and accelerating glycolysis in GC cells (113). By contrast, circUBE2G1 can
encode circ_UBE2G1-99aa, which binds to enolase 1, thereby
promoting glycolysis in GC (114). Similarly, circFNDC3B may encode
a 267-amino-acid peptide that facilitates c-Myc protein degradation
via the proteasomal pathway, inhibiting GC cell proliferation and
migration (115). These findings
underscore the importance of c-Myc in amino acid metabolic
remodeling and suggest that circFNDC3B may participate in GC
progression through this metabolic axis. c-Myc serves as a key
transcriptional driver of PRPS2 in the de novo synthesis
pathways of purines and pyrimidines (108). Whether the negative regulatory
mechanism of FNDC3B-267aa on c-Myc (115) establishes a link between this
peptide and nucleotide metabolism remains to be determined, as
direct evidence demonstrating that FNDC3B-267aa alters PRPS2
activity, nucleotide precursor levels or pyrimidine/purine pools in
GC cells is currently lacking.
To date, studies on circRNA-encoded micropeptides
regulating metabolic reprogramming in GC remain relatively limited,
with most focusing on glycolytic pathways. However, research on
other solid tumors, such as glioblastoma and colorectal cancer, has
revealed that circRNA-derived micropeptides influence malignant
progression by modulating key processes such as nucleotide
metabolism and FAO (112,116).
Given the high conservation of these metabolic pathways in cancer,
it is plausible that GC proliferation similarly depends on such
mechanisms. For example, a novel 127-amino-acid micropeptide
encoded by circSpdyA binds to the ER catalytic domain of FA
synthase, activating de novo FA synthesis, enhancing FAO and
upregulating CPT1A activity. This promotes the uptake of
accumulated FAs to supply membrane biosynthesis and support tumor
cell proliferation and metastasis in breast cancer (117). Another micropeptide, E2F1-99aa,
is encoded by circE2F1, which contains an IRES and a 297-nucleotide
open reading frame. E2F1-99aa mimics the DP-binding domain of E2F1,
interacts with SPIB and competitively inhibits E2F1-SPIB binding.
This intervention downregulates nucleotide biosynthetic gene
expression, reduces purine and pyrimidine synthesis and
consequently suppresses neuroblastoma progression (118). Future research should focus on
identifying GC-specific circRNA-encoded micropeptides and
validating whether their functions align with these conserved
metabolic pathways.
Non-coding and coding mechanisms:
m6A modification
m6A modification of circRNAs is regulated
by methyltransferases, demethylases and reader proteins, primarily
occurring at the N6 position of adenine residues
(119). The covalently closed
circular structure of circRNAs restricts the spatial distribution
of modification sites, facilitating the formation of an
'aggregation effect' of m6A modifications. This effect
enhances the recruitment efficiency of binding proteins and results
in a more concentrated distribution of modification sites within
the exonic sequences of circularized regions (120). Notably, m6A
modification of circRNAs primarily reinforces and expands their
non-coding functional repertoire; it enables the subcellular
trafficking and localization of circRNAs between the nucleus and
cytoplasm and regulates processes such as RNA processing,
degradation, nuclear export and translation (121). By modulating the abundance and
functional duration of circRNAs, m6A modification serves
as one of the critical pathways mediating circRNA degradation
(122), thereby further
interfering with the metabolic homeostasis of tumor cells and
contributing to tumor progression (123). However, in rare, specialized
contexts, m6A modification can endow circRNAs with
coding potential, whereby the m6A reader protein YTHDF3
recognizes m6A-modified circRNAs, recruits the
translation initiation factor eIF4G2, and mediates ribosome
assembly on circRNAs. By providing an IRES, this process drives the
translation of circRNAs into short peptides (124). Notably, a single m6A
modification is sufficient to initiate circRNA translation
(125). In such cases, certain
circRNA molecules temporarily acquire coding capacity upon
m6A modification, which contributes to the complex
regulatory networks underlying tumor initiation and
progression.
Abnormal glycolytic flux is a key hallmark of
bioenergetic metabolism in GC. circRNAs can regulate glycolytic
pathways through epigenetic remodeling, while dynamic
m6A epitranscriptomic modifications modulate the
expression of glycolytic enzymes via
methyltransferase/demethylase-mediated RNA methylation cycles.
These interconnected mechanisms collectively maintain the metabolic
plasticity of GC cells (126).
FTO, an RNA demethylase with specific m6A demethylation
activity (127), binds to
circFAM192A at specific sites and removes its m6A
modifications to prevent degradation. Subsequently, circFAM192A
interacts with the membrane-localized leucine transporter SLC7A5 to
enhance its stability, increasing the membrane localization of
SLC7A5 for leucine transport. Elevated leucine uptake thereby
promotes GC cell proliferation (128). FTO can also specifically remove
the m6A modification of circ_0112136 to maintain its
stability, thereby activating the PI3K/AKT/mTOR pathway. Notably,
mTORC1 is a critical signaling molecule that activates the
rate-limiting enzyme of pyrimidine synthesis (129,130). It should be emphasized that
whether the FTO/circ_0112136 axis regulates pyrimidine
biosynthesis, carbamoyl-phosphate synthetase 2, aspartate
transcarbamylase and dihydroorotase (CAD) activity or nucleotide
levels in GC cells via the mTORC1 signaling pathway warrants
further investigation. This speculation is derived solely from the
established function of mTORC1 in nucleotide metabolism.
Although the most abundant epitranscriptomic
modification of RNA, direct evidence for m6A-modified
circRNAs regulating metabolic remodeling pathways in GC remains
limited. However, findings from other tumor types provide valuable
insights. For example, in hepatocellular carcinoma,
m6A-modified circRAPGEF1 competitively binds to IGF2BP3
via its KH domain, leading to the degradation of certain enzymes in
aspartate metabolism. Aspartate activates CAD, the rate-limiting
enzyme in de novo pyrimidine synthesis, thereby enhancing
tumor cell stemness (131). The
'm6A-circRNA-metabolic reprogramming' axis represents a
conserved regulatory module across tumors, which strengthens the
credibility of a similar mechanism operating in GC. Table I systematically summarizes the
molecular mechanisms of circRNAs in the metabolic reprogramming of
GC.
Evidence for circRNAs as potential
biomarkers
Patients with early-stage GC typically present with
no specific clinical manifestations, but may exhibit non-specific
symptoms such as mild epigastric pain, abdominal distension and
acid reflux (132,133). These symptoms are often
overlooked, leading to diagnostic delays. Consequently, most
patients are diagnosed at an advanced stage when they first seek
medical attention, which notably increases the difficulty of
treatment. Therefore, early diagnosis is a critical strategy to
improve patient survival rates. In certain high-incidence
countries, screening and intervention measures have alleviated part
of the disease burden. However, challenges such as low early
diagnosis rates, limited treatment options due to high tumor
heterogeneity and insufficient sensitivity of disease monitoring
indicators still persist (1).
Thus, the identification of novel biomarkers, development of
reproducible detection methods, detection of preclinical disease
and implementation of large-scale population screening are crucial
for early diagnosis and improving clinical outcomes (134).
At present, endoscopic biopsy combined with
histopathological examination is the gold standard for the
diagnosis of GC, with endoscopic ultrasonography providing
specificity for tumor staging (135). However, these methods have
certain limitations, including being highly invasive, having poor
patient compliance and relatively high operational costs.
Conventional tumor markers such as carcinoembryonic antigen (CEA)
and carbohydrate antigen 19-9 (CA19-9) are not specific to GC and
have insufficient sensitivity and specificity (136). Therefore, developing
non-invasive biomarkers with high sensitivity and accuracy that can
dynamically reflect disease activity has become a key focus in GC
clinical research (137). Due to
their covalently closed circular structure, circRNAs are highly
stable in body fluids (such as serum, plasma and gastric juice),
exhibit high tissue specificity and show expression patterns
correlated with tumor progression. They are also closely related to
tumor metabolic reprogramming and have been recognized as novel
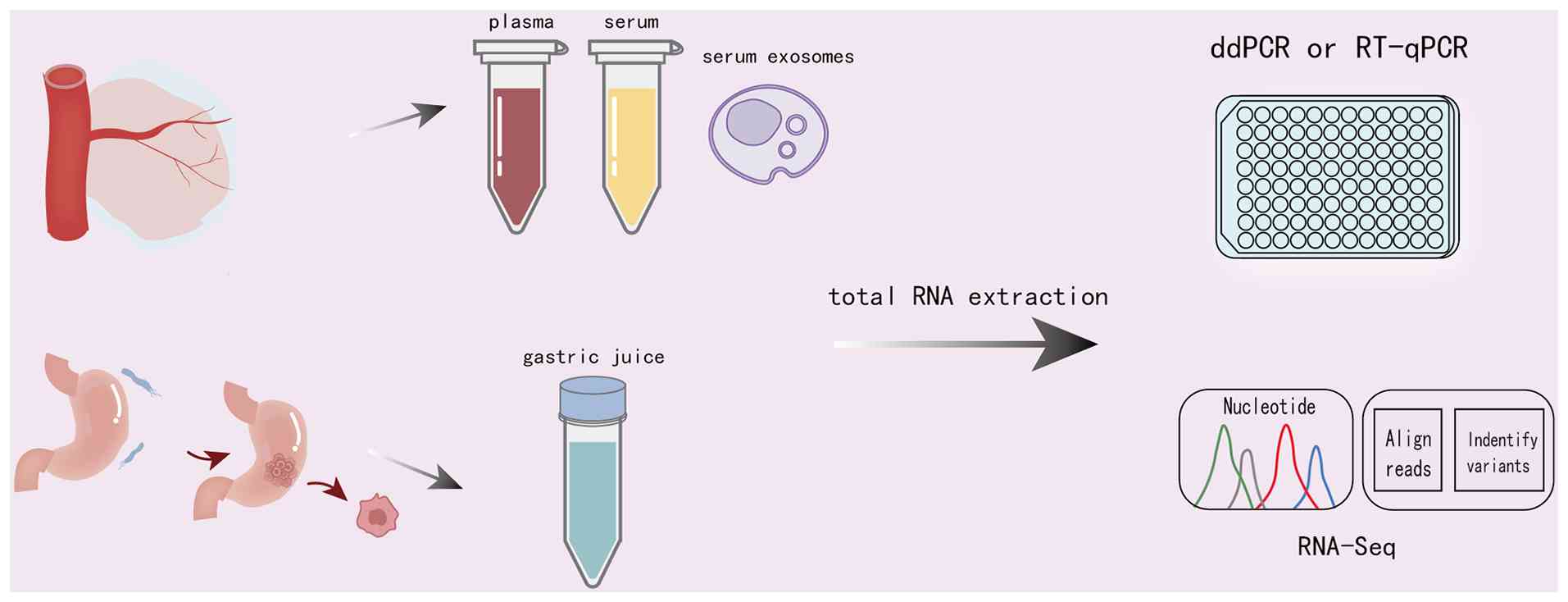
biomarkers (138). Currently,
techniques such as droplet digital polymerase chain reaction,
reverse transcription quantitative PCR or RNA sequencing (RNA-seq)
are commonly used for detection (Fig.
3). A study has confirmed that circRNAs in the body fluids of
patients with GC display differential expression, which may be
associated with the anatomical location of the tumor (139). These findings provide a
molecular 'yardstick' for the clinical diagnosis, screening and
treatment of GC. Table II
summarizes the potential diagnostic value of circRNAs from
different sample sources.
| Table IIResearch on circRNAs from different
sample sources as clinical diagnostic biomarkers. |
Table II
Research on circRNAs from different
sample sources as clinical diagnostic biomarkers.
| First author,
year | circRNAs | Samples | Expression
alteration | AUC | SEN (%), SPE
(%) | Function | (Refs.) |
|---|
| Li et al,
2018 |
hsa_circ_0001017 | Plasma | Down | 0.871 | 79.4, 81.1 | Diagnosis,
prognosis | (140) |
| Li et al,
2018 |
hsa_circ_0061276 | Plasma | Down | 0.764 | 90.3, 51.7 | Diagnosis,
prognosis | (140) |
| Huang et al,
2017 |
hsa_circ_0000745 | Plasma | Down | 0.683 | 85.5, 45 | Diagnosis, tumor
differentiation | (143) |
| Chen et al,
2017 |
hsa_circ_0000190 | Plasma | Down | 0.60 | 41.4, 87.5 | Screening | (144) |
| Zhao et al,
2018 |
hsa_circ_0000181 | Plasma | Down | 0.582 | 99.0, 20.6 | Tumor
differentiation | (145) |
| Ma et al,
2022 |
hsa_circ_0003195 | Plasma | Down | 0.695 | 46.9, 94.6 | Screening,
prognosis | (146) |
| Ma et al,
2021 | circPTPN22 | Plasma | Up | 0.857 | 78, 84 | Diagnosis | (147) |
| Li et al,
2017 |
hsa_circ_0001649 | Serum | Down | 0.834 | 71.1, 81.6 | Screening | (150) |
| Fang et al,
2024 |
hsa_circ_0000231 | Serum | Down | 0.781 | 86.73, 57.81 | Diagnosis
monitoring, prognosis | (151) |
| Ma et al,
2024 | circHAS2 | Serum | Up | 0.845 | 74, 83 | Screening,
prognosis | (152) |
| Yuan et al,
2023 |
hsa_circ_0000702 | Serum | Down | 0.745 | 82.69, 48.44 | Diagnosis,
monitoring | (153) |
| Zang et al,
2024 |
hsa_circ_0050547 | Serum exosomes | Up | 0.90 | 68.42, 100 | Diagnosis | (154) |
| Huang et al,
2023 |
hsa_circ_000200 | Serum exosomes | Up | 0.7092 | 98, 44 | Diagnosis | (155) |
| Shao et al,
2017 |
hsa_circ_0014717 | Gastric juice | ns | / | / | Screening | (157) |
| Shao et al,
2024 |
hsa_circ_0005927 | Plasma | Up | 0.623 | 52.38, 76.19 | Screening | (148) |
| Shao et al,
2024 |
hsa_circ_0005927 | Gastric juice | ns | / | / | / | (148) |
| Shao et al,
2020 |
hsa_circ_0065149 | Plasma | Down | 0.640 | 48.7, 90.2 | Screening,
prognosis | (149) |
| Shao et al,
2020 |
hsa_circ_0065149 | Gastric juice | ns | / | / | / | (149) |
Plasma and serum
Plasma and serum are clinically accessible body
fluid samples, and research on circRNA biomarkers in these fluids
is relatively advanced. On the one hand, plasma circRNAs can be
used for early detection and to distinguish between healthy
individuals and patients with GC. For example, the combined
detection of circulating hsa_circ_0001017 and hsa_circ_0061276 is
closely associated with major clinicopathological factors of GC,
showing a sensitivity and specificity of 84.7 and 96.6%,
respectively, which demonstrates notable clinical value for GC
diagnosis and prognosis assessment (140). On the other hand, plasma
circRNAs can differentiate between different clinical stages of
patients with GC. For instance, the expression level of plasma
exosomal circLPAR1 in patients with GC is notably decreased in the
advanced stages (stages IIb and III), while it is elevated in the
early and intermediate stages (stages I and IIa) or after surgery.
Moreover, the diagnostic performance of plasma exosomal circLPAR1
is superior to that of CEA and CA19-9, with notably improved
diagnostic efficiency, demonstrating good sensitivity and
specificity, making it a potential biomarker for GC diagnosis and
staging (141). Additionally,
studies have isolated exosomes from the plasma of patients with GC
and healthy individuals and used high-throughput RNA-seq to analyze
circRNA expression in circulating exosomes. The authors found that
hsa_circ_0093425, hsa_circ_0007247 and hsa_circ_0029780 were
significantly upregulated, while hsa_circ_0139983,
hsa_circ_0001989, hsa_circ_0011115 and hsa_circ_0019618 were
significantly downregulated in patients with GC, indicating that
plasma exosomal circRNAs exhibit differential expression in GC
(142). In addition,
plasma-derived hsa_circ_0000745 (143), hsa_circ_0000190 (144), hsa_circ_0000181 (145), hsa_circ_0003195 (146), circPTPN22 (147), hsa_circ_0005927 (148) and hsa_circ_0065149 (149), as well as serum-derived
hsa_circ_0001649 (150),
hsa_circ_0000231 (151),
circHAS2 (152),
hsa_circ_0000702 (153),
hsa_circ_0050547 (154) and
hsa_circ_000200 (155), also
play important roles in the screening, diagnosis and prognosis of
GC.
Gastric juice
Gastric juice is directly secreted by gastric cells
and more accurately reflects local gastric lesions compared with
serum and plasma; it is not cleared by the liver and, in theory,
may lack ribonucleoprotein and lipoprotein complexes, thereby
reducing the risk of ncRNA degradation by nucleases, proteases and
the acidic environment of gastric juice. This makes gastric juice a
potentially ideal source of biomarkers for the dynamic monitoring
of GC screening, diagnosis, treatment efficacy and recurrence
(156). Shao et al
(157) studied circRNA
expression changes in gastric juice and confirmed that
hsa_circ_0014717 can stably exist in human gastric juice. By
comparing healthy individuals, patients with gastric ulcer,
patients with chronic atrophic gastritis and patients with GC, they
found that hsa_circ_0014717 expression was significantly
downregulated in the chronic atrophic gastritis group, but
increased in the GC group, a trend similar to that observed for
hsa_circ_0005927 in gastric juice (148). This suggests a possible link to
tumor cell secretion mechanisms and changes in the TME.
Additionally, hsa_circ_0065149 was shown to stably exist in gastric
juice, but its expression did not notably differ among the four
groups. Furthermore, the expression trend of hsa_circ_0065149 in
gastric juice was not consistent with that in GC tissues or plasma
exosomes, suggesting that this may be related to small sample size
or selective changes in circRNA sources (149). Therefore, gastric juice circRNA
holds unique value for early evaluation of GC treatment efficacy
and is especially suitable for patients unable to tolerate repeated
endoscopic examinations.
Challenges in circRNA and metabolic
reprogramming research
Current research on circRNA biomarkers
predominantly focuses on tissue samples, with a smaller number of
studies investigating their presence in body fluids such as serum,
plasma and exosomes. The majority of these are single-center,
retrospective studies with limited statistical power, which
currently precludes their translation into routine clinical testing
(158). Some scholars have
performed independent validation using serum samples from both
training and validation sets, including patients with GC and
control subjects (159).
However, the cut-off value was mainly determined based on the
training set, lacking real-world prospective cohort studies.
Furthermore, the diagnostic efficacy of a single circRNA is
limited, requiring evaluation using multi-molecule panels (160). In addition, the individual
physiological status of the patient is associated with circRNA
expression levels in body fluids, which may lead to fluctuations in
the accuracy and sensitivity of circRNAs as biomarkers (161). Although circRNA-based biomarker
prediction systems remain in the exploratory stage, multiple
hospitals have completed the clinical registration of prospective,
multicenter studies investigating circRNA, indicating that this
field is advancing. Efforts are being made to overcome the 'noise'
caused by individual heterogeneity through technical optimization,
multicenter validation and large-sample verification.
There is also a lack of uniform standards for the
identification and validation of circRNAs. The back-splice site is
critical for circRNA identification. However, variations exist
among research teams in raw materials such as cell types and
platforms, as well as in analytical pipelines and methods including
RNA isolation, linear RNA depletion and library preparation
(162). Furthermore, circRNA
abundance in body fluids is affected by multiple pre-analytical
factors, including centrifugation time and temperature, freeze-thaw
cycles and serum clotting time (163). In addition, the detection and
analysis of circRNAs require specialized high-precision instruments
to generate millions of sequencing reads, making it difficult to
extend to routine clinical settings in the short term (164).
Meanwhile, cell lines used in laboratory studies
differ metabolically and functionally from their in vivo
counterparts. Animal models, while useful, are limited by
species-specific differences and cannot fully recapitulate human
disease pathology. Therefore, validating circRNA findings in human
cohorts is essential to strengthen their clinical relevance
(165).
Research on metabolic reprogramming has often
lacked visual evidence directly linking it to clinical disease
manifestations. However, the emergence of 18F-FDG PET/CT
imaging has repositioned metabolic reprogramming at the forefront
of oncology research. This technique assesses tumor metabolic
activity by measuring the accumulation of 18F-FDG, a
glucose analog, within key molecules involved in the metabolic
reprogramming of cancer cells (166). A growing body of evidence
indicates that aberrantly expressed circRNAs contribute to GC
progression by modulating the activity and expression of metabolic
enzymes, thereby driving metabolic remodeling. This influence on
cellular energy supply and biosynthetic capacity positions circRNAs
as critical regulators of GC proliferation, invasion, metastasis,
drug resistance and immune evasion. While studies focusing on the
glycolytic axis are relatively abundant, further investigation is
needed to elucidate the roles of circRNAs in other metabolic
pathways, such as amino acid and nucleotide metabolism, within the
context of GC metabolic reprogramming. Such an imbalance in
research focus not only reflects that glycolytic indicators are
relatively easy to observe but also reveals that current findings
are largely limited to easily verifiable stages, which does not
allow the inference that circRNAs participate in the regulation of
the entire metabolic network in GC.
Moreover, studies investigating circRNA-mediated
regulation of nucleotide synthesis or catabolism in GC are
characterized by substantial limitations, most notably the absence
of direct quantification of nucleotide intermediates, de
novo purine/pyrimidine synthesis flux or the activities of
rate-limiting enzymes. By contrast to the conventional assay kit
approaches widely employed in glycolysis research, methodologies
for nucleotide metabolic analysis are technically demanding and
lack accessible phenotypic endpoints. These challenges likely
represent major contributing factors to the current dearth of
research in this field.
The mechanisms by which circRNAs regulate metabolic
reprogramming in GC are not limited to linear pathways. Recent
studies have demonstrated that circRNAs influence GC remodeling
through multiple strategies, including direct interaction with
metabolic enzymes (114),
modulation of epigenetic modifications (167) and participation in various
signaling pathways (168). For
example, a single circRNA can regulate multiple signaling circuits.
circ-NRIP1 competitively sequesters miR-186-5p, thereby relieving
its suppression of MYH9 and promoting GC cell proliferation,
migration and glycolysis (169).
Independently, it also acts as a sponge for miR-138-5p, modulating
HIF-1α-dependent glycolysis and sensitizing GC cells to
hypoxia-induced 5-fluorouracil resistance (170). This suggests that the circRNA
regulatory network is context-dependent, yet the mechanisms
underlying the functional allocation of circRNAs across distinct
signaling pathways under specific cellular backgrounds or
microenvironmental conditions remain elusive.
Furthermore, metabolic reprogramming in GC exhibits
notable heterogeneity. Different cellular subpopulations within the
same tumor, such as cancer stem cells and differentiated tumor
cells, utilize distinct metabolic pathways, which dynamically
evolve during disease progression (171). There are substantial differences
between model cell lines and human tumors. Nutrient composition,
oxygen tension and cell-cell interactions in in vitro
culture systems tend to exaggerate specific metabolic phenotypes;
meanwhile, animal models are limited by interspecies differences.
Current research predominantly relies on bulk RNA-seq of mixed cell
populations, which provides averaged gene expression profiles, or
single-cell RNA-seq of individual cell lines, which may be subject
to limited genetic drift and fail to fully represent intratumoral
diversity (172,173). Both approaches face challenges
in comprehensively capturing this metabolic heterogeneity. Although
notable hurdles remain in elucidating circRNA-mediated metabolic
reprogramming, spanning mechanistic validation, technological
limitations and clinical translation, the field is poised for
advancement. The deepening integration of multi-omics technologies,
the initiation of multi-center prospective clinical studies and
continued optimization of targeted delivery systems (such as
antisense oligonucleotides, small interfering RNAs and CRISPR/Cas
systems) collectively strengthen the potential of circRNAs to
provide critical insights for GC screening, diagnosis, therapeutic
monitoring and detection of recurrence (174).
Conclusion
GC exhibits high heterogeneity and its pathogenesis
involves complex metabolic remodeling networks. circRNAs, as
stable, conserved and functionally versatile members of the ncRNA
family, are no longer regarded as biological 'dark matter'.
Instead, they have emerged as key regulators of metabolic
reprogramming in GC. Through diverse coding and non-coding
mechanisms, circRNAs indirectly modulate target genes involved in
glucose, lipid, amino acid and nucleotide metabolism. These
findings highlight the interplay between epigenetic regulation and
metabolic phenotypes, underscoring the potential of circRNAs as
biomarkers and therapeutic targets. However, the pronounced
heterogeneity of GC poses notable challenges for clinical
management. The expression profiles of circRNAs and their selective
roles in metabolic regulation across different molecular subtypes,
such as HER2-positive, EBV-positive and microsatellite
instability-high GC remain poorly characterized. Moreover, current
research has largely focused on the impact of circRNAs on
individual metabolic pathways, such as glycolysis, while systematic
investigations into the mechanisms by which circRNAs coordinately
regulate interconnected metabolic processes in GC are still
lacking. Integrative analysis using metabolomics and single-cell
sequencing remains limited, making it difficult to capture the
spatiotemporal dynamics of circRNA expression and metabolite
concentrations in GC. This gap hinders the identification of
critical regulatory networks. Future studies should prioritize
multi-omics approaches to construct comprehensive molecular maps of
circRNA-mediated metabolic reprogramming, thereby providing a
theoretical foundation and technical support for precision
metabolic therapy in GC. In summary, targeting circRNA-regulated
metabolic reprogramming holds substantial promise for advancing
both basic research and clinical applications in GC.
Availability of data and materials
Not applicable.
Authors' contributions
XL conceived and designed the review. TW wrote the
first draft. JM, SZ, WG, YY and XZ participated in writing the
manuscript. HL edited, reviewed and supervised the manuscript and
acquired funding. All authors contributed to the manuscript. All
authors read and approved the final version of the manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science
Foundation of China (grant no. 82274442).
References
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Li X, Wang J, Ma Y, Wang S, Yu X, Niu K,
Yan P, Wu D, Song J, Kou Y, et al: Burden and trends of early-onset
gastric cancer in the 11 BRICS countries (2025 expansion):
1990-2021 with projections to 2035. BMC Cancer. 25:14782025.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Luzko I, Moreira L and Bornschein J:
Screening for and surveillance of premalignant conditions of the
stomach. Best Pract Res Clin Gastroenterol. 75:1019782025.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ko MT, Fung A, Kumar A, McArdle A and
Alexandre L: Post-endoscopy upper gastrointestinal cancer: Emerging
data and opportunities to improve early detection. Best Pract Res
Clin Gastroenterol. 75:1020032025. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo D, Zhou J, Ruan S, Zhang B, Zhu H, Que
Y, Ying S, Li X, Hu Y and Song Z: Overcoming immunotherapy
resistance in gastric cancer: insights into mechanisms and emerging
strategies. Cell Death Dis. 16:752025. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun Q, Li S, Lou J, Wang X and Xu X:
Recent advances in tumour microenvironment impact immunotherapy
resistance in gastric cancer. Crit Rev Oncol Hematol.
215:1048372025. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vander Heiden MG, Cantley LC and Hompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schiliro C and Firestein BL: Mechanisms of
metabolic reprogramming in cancer cells supporting enhanced growth
and proliferation. Cells. 10:10562021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cai H, Zhang F, Xu F and Yang C: Metabolic
reprogramming and therapeutic targeting in non-small cell lung
cancer: Emerging insights beyond the Warburg effect. Front Oncol.
15:15642262025. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Marei HE: Epigenetic regulators in cancer
therapy and progression. NPJ Precis Oncol. 9:2062025. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Toden S, Zumwalt TJ and Goel A: Non-coding
RNAs and potential therapeutic targeting in cancer. Biochim Biophys
Acta Rev Cancer. 1875:1884912021. View Article : Google Scholar
|
|
13
|
Saw PE, Xu X, Chen J and Song E:
Non-coding RNAs: The new central dogma of cancer biology. Sci China
Life Sci. 64:22–50. 2021. View Article : Google Scholar
|
|
14
|
Xie S, Chang Y, Jin H, Yang F, Xu Y, Yan
X, Lin A, Shu Q and Zhou T: Non-coding RNAs in gastric cancer.
Cancer Lett. 493:55–70. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Sun D, Pu W, Wang J and Peng Y:
Circular RNAs in cancer: Biogenesis, function, and clinical
significance. Trends Cancer. 6:319–336. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia L, Song M, Sun M, Wang F and Yang C:
Circular RNAs as biomarkers for cancer. Adv Exp Med Biol.
1087:171–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pandey PR, Munk R, Kundu G, De S,
Abdelmohsen K and Gorospe M: Methods for analysis of circular RNAs.
Wiley Interdiscip Rev RNA. 11:e15662020. View Article : Google Scholar
|
|
18
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He AT, Liu J, Li F and Yang BB: Targeting
circular RNAs as a therapeutic approach: Current strategies and
challenges. Signal Transduct Target Ther. 6:1852021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee K, Ku J, Ku D and Kim Y: Inverted Alu
repeats: Friends or foes in the human transcriptome. Exp Mol Med.
56:1250–1262. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar :
|
|
22
|
Wang S, Sun Z, Lei Z and Zhang H:
RNA-binding proteins and cancer metastasis. Semin Cancer Biol.
86:748–768. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Galarneau A and Richard S: Target RNA
motif and target mRNAs of the Quaking STAR protein. Nat Struct Mol
Biol. 12:691–698. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hong H, An O, Chan THM, Ng VHE, Kwok HS,
Lin JS, Qi L, Han J, Tay DJT, Tang SJ, et al: Bidirectional
regulation of adenosine-to-inosine (A-to-I) RNA editing by DEAH box
helicase 9 (DHX9) in cancer. Nucleic Acids Res. 46:7953–7969. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ivanov A, Memczak S, Wyler E, Torti F,
Porath HT, Orejuela MR, Piechotta M, Levanon EY, Landthaler M,
Dieterich C and Rajewsky N: Analysis of intron sequences reveals
hallmarks of circular RNA biogenesis in animals. Cell Rep.
10:170–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Warburg O: On respiratory impairment in
cancer cells. Science. 124:269–270. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ciccarone F and Ciriolo MR: Reprogrammed
mitochondria: A central hub of cancer cell metabolism. Biochem Soc
Trans. 52:1305–1315. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhan H, Pal DS, Borleis J, Deng Y, Long Y,
Janetopoulos C, Huang CH and Devreotes PN: Self-organizing
glycolytic waves tune cellular metabolic states and fuel cancer
progression. Nat Commun. 16:55632025. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang H, Liu S, Fu S, Zhao Q, Wang Y, Yuan
Y and Zhang C: Novel insight into the Warburg effect: Sweet
temptation. Crit Rev Oncol Hematol. 214:1048442025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qu J, Li P and Sun Z: Histone lactylation
regulates cancer progression by reshaping the tumor
microenvironment. Front Immunol. 14:12843442023. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pavlova NN, Zhu J and Thompson CB: The
hallmarks of cancer metabolism: Still emerging. Cell Metab.
34:355–377. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barba I, Carrillo-Bosch L and Seoane J:
Targeting the Warburg effect in cancer: Where do we stand? Int J
Mol Sci. 25:31422024. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Q, Li M, Chen C, Xu L, Fu Y, Xu J,
Shu C, Wang B, Wang Z, Chen C, et al: Glucose homeostasis controls
N-acetyltransferase 10-mediated ac4C modification of HK2 to drive
gastric tumorigenesis. Theranostics. 15:2428–2450. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bian X, Jiang H, Meng Y, Li YP, Fang J and
Lu Z: Regulation of gene expression by glycolytic and gluconeogenic
enzymes. Trends Cell Biol. 32:786–799. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Campos M and Albrecht LV: Hitting the
sweet spot: How glucose metabolism is orchestrated in space and
time by phosphofructokinase-1. Cancers (Basel). 16:162023.
View Article : Google Scholar
|
|
37
|
Zhang Z, Han S, Ouyang S, Zeng Z, Liu Z,
Sun J and Kang W: PDK4 constitutes a novel prognostic biomarker and
therapeutic target in gastric cancer. Diagnostics (Basel).
12:11012022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang R, Mao G, Tang Y, Li C, Gao Y, Nie
W, Song T, Liu S, Zhang P, Tao K and Li W: Inhibition of glycolysis
enhances the efficacy of immunotherapy via PDK-mediated
upregulation of PD-L1. Cancer Immunol Immunother. 73:1512024.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhu S, Guo Y, Zhang X, Liu H, Yin M, Chen
X and Peng C: Pyruvate kinase M2 (PKM2) in cancer and cancer
therapeutics. Cancer Lett. 503:240–248. 2021. View Article : Google Scholar
|
|
40
|
Zhang Z, Deng X, Liu Y, Liu Y, Sun L and
Chen F: PKM2, function and expression and regulation. Cell Biosci.
9:522019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Röhrig F and Schulze A: The multifaceted
roles of fatty acid synthesis in cancer. Nat Rev Cancer.
16:732–749. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Houten SM and Wanders RJ: A general
introduction to the biochemistry of mitochondrial fatty acid
β-oxidation. J Inherit Metab Dis. 33:469–477. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jin HR, Wang J, Wang ZJ, Xi MJ, Xia BH,
Deng K and Yang JL: Lipid metabolic reprogramming in tumor
microenvironment: From mechanisms to therapeutics. J Hematol Oncol.
16:1032023. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li C, Zhang L, Qiu Z, Deng W and Wang W:
Key molecules of fatty acid metabolism in gastric cancer.
Biomolecules. 12:7062022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Guerrero-Rodríguez SL, Mata-Cruz C,
Pérez-Tapia SM and Velasco-Velázquez MA: Role of CD36 in cancer
progression, stemness, and targeting. Front Cell Dev Biol.
10:10790762022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hao J, Wang J, Guo H, Zhao Y, Sun H, Li Y,
Lai X, Zhao N, Wang X, Xie C, et al: CD36 facilitates fatty acid
uptake by dynamic palmitoylation-regulated endocytosis. Nat Commun.
11:47652020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jiang M, Wu N, Xu B, Chu Y, Li X, Su S,
Chen D, Li W, Shi Y, Gao X, et al: Fatty acid-induced CD36
expression via O-GlcNAcylation drives gastric cancer metastasis.
Theranostics. 9:5359–5373. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Severin M, Hansen RK, Rolver MG, Hels T,
Maeda K, Pardo LA and Pedersen SF: Tumor acidosis supports cancer
cell lipid uptake via a rapid transporter-independent mechanism. J
Cell Sci. 138:jcs2636882025. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Schlaepfer IR and Joshi M: CPT1A-mediated
fat oxidation, mechanisms, and therapeutic potential.
Endocrinology. 161:bqz0462020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pereyra AS, Rajan A, Ferreira CR and Ellis
JM: Loss of muscle carnitine palmitoyltransferase 2 prevents
diet-induced obesity and insulin resistance despite long-chain
acylcarnitine accumulation. Cell Rep. 33:1083742020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang C, Zhang C, Li X, Shen J, Xu Y, Shi
H, Mu X, Pan J, Zhao T, Li M, et al: CPT1A-mediated succinylation
of S100A10 increases human gastric cancer invasion. J Cell Mol Med.
23:293–305. 2019. View Article : Google Scholar :
|
|
52
|
Wang M, Yu W, Cao X, Gu H, Huang J, Wu C,
Wang L, Sha X, Shen B, Wang T, et al: Exosomal CD44 transmits lymph
node metastatic capacity between gastric cancer cells via
YAP-CPT1A-mediated FAO reprogramming. Front Oncol. 12:8601752022.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wang Y, Lu JH, Wang F, Wang YN, He MM, Wu
QN, Lu YX, Yu HE, Chen ZH, Zhao Q, et al: Inhibition of fatty acid
catabolism augments the efficacy of oxaliplatin-based chemotherapy
in gastrointestinal cancers. Cancer Lett. 473:74–89. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ma G, Zhang Z, Li P, Zhang Z, Zeng M,
Liang Z, Li D, Wang L, Chen Y, Liang Y and Niu H: Reprogramming of
glutamine metabolism and its impact on immune response in the tumor
microenvironment. Cell Commun Signal. 20:1142022. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lieu EL, Nguyen T, Rhyne S and Kim J:
Amino acids in cancer. Exp Mol Med. 52:15–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang B, Pei J, Xu S, Liu J and Yu J: A
glutamine tug-of-war between cancer and immune cells: Recent
advances in unraveling the ongoing battle. J Exp Clin Cancer Res.
43:742024. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zou W, Han Z, Wang Z and Liu Q: Targeting
glutamine metabolism as a potential target for cancer treatment. J
Exp Clin Cancer Res. 44:1802025. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ying M, You D, Zhu X, Cai L, Zeng S and Hu
X: Lactate and glutamine support NADPH generation in cancer cells
under glucose deprived conditions. Redox Biol. 46:1020652021.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lu YX, Ju HQ, Liu ZX, Chen DL, Wang Y,
Zhao Q, Wu QN, Zeng ZL, Qiu HB, Hu PS, et al: ME1 regulates NADPH
homeostasis to promote gastric cancer growth and metastasis. Cancer
Res. 78:1972–1985. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu JC, Jiang ZM and Li DM: Glutamine: A
precursor of glutathione and its effect on liver. World J
Gastroenterol. 5:143–146. 1999. View Article : Google Scholar
|
|
61
|
Jiang B, Zhang J, Zhao G, Liu M, Hu J, Lin
F, Wang J, Zhao W, Ma H, Zhang C, et al: Filamentous GLS1 promotes
ROS-induced apoptosis upon glutamine deprivation via insufficient
asparagine synthesis. Mol Cell. 82:1821–1835.e6. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jaune-Pons E and Vasseur S: Role of amino
acids in regulation of ROS balance in cancer. Arch Biochem Biophys.
689:1084382020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Byun JK, Park M, Lee S, Yun JW, Lee J, Kim
JS, Cho SJ, Jeon HJ, Lee IK, Choi YK and Park KG: Inhibition of
glutamine utilization synergizes with immune checkpoint inhibitor
to promote antitumor immunity. Mol Cell. 80:592–606.e8. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Caforio M, Sorino C, Iacovelli S,
Fanciulli M, Locatelli F and Folgiero V: Recent advances in
searching c-Myc transcriptional cofactors during tumorigenesis. J
Exp Clin Cancer Res. 37:2392018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gao P, Tchernyshyov I, Chang TC, Lee YS,
Kita K, Ochi T, Zeller KI, De Marzo AM, Van Eyk JE, Mendell JT and
Dang CV: c-Myc suppression of miR-23a/b enhances mitochondrial
glutaminase expression and glutamine metabolism. Nature.
458:762–765. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Xu E, Ji B, Jin K and Chen Y:
Branched-chain amino acids catabolism and cancer progression: Focus
on therapeutic interventions. Front Oncol. 13:12206382023.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Peng H, Wang Y and Luo W: Multifaceted
role of branched-chain amino acid metabolism in cancer. Oncogene.
39:6747–6756. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yang D, Liu H, Cai Y, Lu K, Zhong X, Xing
S, Song W, Zhang Y, Ye L, Zhu X, et al: Branched-chain amino acid
catabolism breaks glutamine addiction to sustain hepatocellular
carcinoma progression. Cell Rep. 41:1116912022. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang L and Han J: Branched-chain amino
acid transaminase 1 (BCAT1) promotes the growth of breast cancer
cells through improving mTOR-mediated mitochondrial biogenesis and
function. Biochem Biophys Res Commun. 486:224–231. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kami K, Fujimori T, Sato H, Sato M,
Yamamoto H, Ohashi Y, Sugiyama N, Ishihama Y, Onozuka H, Ochiai A,
et al: Metabolomic profiling of lung and prostate tumor tissues by
capillary electrophoresis time-of-flight mass spectrometry.
Metabolomics. 9:444–453. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yu L, Bao S, Zhu F, Xu Y, Liu Y, Jiang R,
Yang C, Cao F, Chen W and Li P: Association between branched-chain
amino acid levels and gastric cancer risk: Large-scale prospective
cohort study. Front Nutr. 11:14798002024. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lane AN and Fan TWM: Regulation of
mammalian nucleotide metabolism and biosynthesis. Nucleic Acids
Res. 43:2466–2485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Feng Q, Zhang W, Peng Y, Zhao C, Ren J and
Qu X: Precise targeting one-carbon metabolism for potent cancer
therapy and metastasis suppression. Small. 21:e046312025.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Malvi P, Bugide S, Dutta R, Reddi KK,
Edwards YJK, Singh K, Gupta R and Wajapeyee N: PDE7A inhibition
suppresses triple-negative breast cancer by attenuating de novo
pyrimidine biosynthesis. Cell Rep Med. 6:1023562025. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Mullen NJ and Singh PK: Nucleotide
metabolism: A pan-cancer metabolic dependency. Nat Rev Cancer.
23:275–294. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tran DH, Kim D, Kesavan R, Brown H, Dey T,
Soflaee MH, Vu HS, Tasdogan A, Guo J, Bezwada D, et al: De novo and
salvage purine synthesis pathways across tissues and tumors. Cell.
187:3602–3618.e20. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yoon BK, Kim H, Oh TG, Oh SK, Jo S, Kim M,
Chun KH, Hwang N, Lee S, Jin S, et al: PHGDH preserves one-carbon
cycle to confer metabolic plasticity in chemoresistant gastric
cancer during nutrient stress. Proc Natl Acad Sci USA.
120:e22178261202023. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Feng X, Ma D, Zhao J, Song Y, Zhu Y, Zhou
Q, Ma F, Liu X, Zhong M, Liu Y, et al: UHMK1 promotes gastric
cancer progression through reprogramming nucleotide metabolism.
EMBO J. 39:e1025412020. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Shi DD, Savani MR, Abdullah KG and
McBrayer SK: Emerging roles of nucleotide metabolism in cancer.
Trends Cancer. 9:624–635. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Møller SH, Mellergaard M, Madsen M,
Bermejo AV, Jepsen SD, Hansen MH, Høgh RI, Aldana BI, Desler C,
Rasmussen LJ, et al: Cytoplasmic citrate flux modulates the immune
stimulatory NKG2D ligand MICA in cancer cells. Front Immunol.
11:19682020. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Ferrari de Andrade L, Tay RE, Pan D, Luoma
AM, Ito Y, Badrinath S, Tsoucas D, Franz B, May KF Jr, Harvey CJ,
et al: Antibody-mediated inhibition of MICA and MICB shedding
promotes NK cell-driven tumor immunity. Science. 359:1537–1542.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Chen S and Wang Z: Integration of
mult-omics and nucleotide metabolism reprogramming signature
analysis reveals gastric cancer immunological and prognostic
features. Cancer Cell Int. 24:2122024. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Losenkova K, Zuccarini M, Karikoski M,
Laurila J, Boison D, Jalkanen S and Yegutkin GG:
Compartmentalization of adenosine metabolism in cancer cells and
its modulation during acute hypoxia. J Cell Sci. 133:jcs2414632020.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Antonioli L, Fornai M, Blandizzi C, Pacher
P and Haskó G: Adenosine signaling and the immune system: When a
lot could be too much. Immunol Lett. 205:9–15. 2019. View Article : Google Scholar
|
|
85
|
Haskó G, Linden J, Cronstein B and Pacher
P: Adenosine receptors: Therapeutic aspects for inflammatory and
immune diseases. Nat Rev Drug Discov. 7:759–770. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wang M, Zhao X, Hou S, Wu Z and Yin HY:
Purinergic receptors in dendritic cells. J Inflamm Res.
18:13423–13432. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Pisignano G, Michael DC, Visal TH, Pirlog
R, Ladomery M and Calin GA: Going circular: History, present, and
future of circRNAs in cancer. Oncogene. 42:2783–2800. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Shahpari M, Hashemi M, Younesirad T,
Hasanzadeh A, Mosanne MM and Ahmadifard M: The functional roles of
competitive endogenous RNA (ceRNA) networks in apoptosis in human
cancers: The circRNA/miRNA/mRNA regulatory axis and cell signaling
pathways. Heliyon. 10:e370892024. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Li H, Cao B, Zhao R, Li T, Xu X, Cui H,
Deng H, Gao J and Wei B: circDNMT1 promotes malignant progression
of gastric cancer through targeting miR-576-3p/hypoxia inducible
factor-1 alpha axis. Front Oncol. 12:8171922022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Fang X, Bai Y, Zhang L and Ding S:
Silencing circSLAMF6 represses cell glycolysis, migration, and
invasion by regulating the miR-204-5p/MYH9 axis in gastric cancer
under hypoxia. Biosci Rep. 40:BSR202012752020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Shen J and Zhu W: Research advances in the
role of gastric cancer-derived mesenchymal stem cells in tumor
progression (review). Int J Mol Med. 47:455–462. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wang L, Wu C, Xu J, Gong Z, Cao X, Huang
J, Dong H, Zhu W, Huang F, Zhou C and Wang M: GC-MSC-derived
circ_0024107 promotes gastric cancer cell lymphatic metastasis via
fatty acid oxidation metabolic reprogramming mediated by the
miR-5572/6855-5p/CPT1A axis. Oncol Rep. 50:1382023. View Article : Google Scholar
|
|
93
|
Li F, Zhang L and Sun Q: CircAKT3 promotes
cell proliferation, survival and glutamine metabolism of gastric
cancer by activating SLC1A5 expression via targeting miR-515-5p.
Histol Histopathol. 37:227–241. 2022.
|
|
94
|
Ye B, Yu S, Wang J and Ren Y: CircB3GNTL1
and miR-598 regulation effects on proliferation, apoptosis, and
glutaminolysis in gastric cancer cells. Cell Mol Biol
(Noisy-le-grand). 66:18–23. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Li J, Zhang G and Wu G: Effect of paeonol
on proliferation, apoptosis, migration, invasion and glutamine of
gastric cancer cells via circSFMBT2/miR-665 axis. Cell Mol Biol
(Noisy-le-grand). 66:33–40. 2020. View Article : Google Scholar
|
|
96
|
Xu X, Wang S, Wang H, Pan C, Yang W and Yu
J: Hsa_circ_0008434 regulates USP9X expression by sponging
miR-6838-5p to promote gastric cancer growth, migration and
invasion. BMC Cancer. 21:12892021. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kim GW, Cha M, Ong HTM, Yoo J, Jeon YH,
Lee SW, Oh SY, Kang MJ, Kim Y and Kwon SH: HDAC6 and USP9X control
glutamine metabolism by stabilizing GS to promote glioblastoma
tumorigenesis. Adv Sci (Weinh). 12:e25015532025. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Li L, Wei C, Xie Y, Su Y, Liu C, Qiu G,
Liu W, Liang Y, Zhao X, Huang D and Wu D: Expanded insights into
the mechanisms of RNA-binding protein regulation of circRNA
generation and function in cancer biology and therapy. Genes Dis.
12:1013832024. View Article : Google Scholar
|
|
99
|
Ji C, Zhu J, Hou X, Zhou C, Zhao J, Zheng
X and Tang Y: PTBP1 and cancer: From RNA regulation to therapeutic
potential. J Cell Mol Med. 29:e706752025. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Qian L, Wang L, Chen H, Wang S, Hou Y, Xu
L, Xia Y, Xu M and Huang X: Hsa_circ_0001756 drives gastric cancer
glycolysis by increasing the expression and stability of PGK1 mRNA.
Front Immunol. 16:15112472025. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Sun Q, Xing X, Wang H, Wan K, Fan R, Liu
C, Wang Y, Wu W, Wang Y and Wang R: SCD1 is the critical signaling
hub to mediate metabolic diseases: Mechanism and the development of
its inhibitors. Biomed Pharmacother. 170:1155862024. View Article : Google Scholar
|
|
102
|
Lin Z, Zhong C, Shi M, Long Q, Jing L, Yu
Y, Chou J, Chen M, Lan M and Long F: Circular RNA TFRC/SCD1 mRNA
interaction regulates ferroptosis and metastasis in gastric cancer.
Cell Death Dis. 16:4362025. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Cellarier E, Durando X, Vasson MP, Farges
MC, Demiden A, Maurizis JC, Madelmont JC and Chollet P: Methionine
dependency and cancer treatment. Cancer Treat Rev. 29:489–499.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Li F, Liu P, Mi W, Li L, Anderson NM,
Lesner NP, Burrows M, Plesset J, Majer A, Wang G, et al: Blocking
methionine catabolism induces senescence and confers vulnerability
to GSK3 inhibition in liver cancer. Nat Cancer. 5:131–146. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Xin L, Zou YH, Liu CX, Lu H, Fan LJ, Xu
HS, Zhou Q, Liu J, Yue ZQ and Gan JH: Methionine restriction
promotes cisplatin sensitivity of gastric cancer resistant cells by
down-regulating circ-CDK13 level. Exp Cell Res. 443:1143152024.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Xu AG, Li SG, Liu JH and Gan AH: Function
of apoptosis and expression of the proteins Bcl-2, p53 and C-myc in
the development of gastric cancer. World J Gastroenterol.
7:403–406. 2001. View Article : Google Scholar
|
|
107
|
Calcagno DQ, Leal MF, Assumpcao PP, Smith
MAC and Burbano RR: MYC and gastric adenocarcinoma carcinogenesis.
World J Gastroenterol. 14:5962–5968. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Cunningham JT, Moreno MV, Lodi A, Ronen SM
and Ruggero D: Protein and nucleotide biosynthesis are coupled by a
single rate-limiting enzyme, PRPS2, to drive cancer. Cell.
157:1088–1103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Yu T, Ran L, Zhao H, Yin P, Li W, Lin J,
Mao H, Cai D, Ma Q, Pan X, et al: Circular RNA circ-TNPO3
suppresses metastasis of GC by acting as a protein decoy for
IGF2BP3 to regulate the expression of MYC and SNAIL. Mol Ther
Nucleic Acids. 26:649–664. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Li H, Jiao W, Song J, Wang J, Chen G, Li
D, Wang X, Bao B, Du X, Cheng Y, et al: circ-hnRNPU inhibits
NONO-mediated c-Myc transactivation and mRNA stabilization
essential for glycosylation and cancer progression. J Exp Clin
Cancer Res. 42:3132023. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Lei M, Zheng G, Ning Q, Zheng J and Dong
D: Translation and functional roles of circular RNAs in human
cancer. Mol Cancer. 19:302020. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhang M, Zhao K, Xu X, Yang Y, Yan S, Wei
P, Liu H, Xu J, Xiao F, Zhou H, et al: A peptide encoded by
circular form of LINC-PINT suppresses oncogenic transcriptional
elongation in glioblastoma. Nat Commun. 9:44752018. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Jiang R, Li P, Meng E, Cheng X, Wu X and
Wu H: Hsa_Circ_0008035 drives immune evasion of gastric cancer via
promoting EXT1-mediated nuclear translocation of PKM2. Transl
Oncol. 48:1020042024. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Lu L, Guo G, Guo J, Li H, Chen K, Chen Y,
Li Q, Li Q, Diao Y, Sun M, et al: A novel protein encoded by
circUBE2G1 suppresses glycolysis in gastric cancer through binding
to ENO1. Cell Death Discov. 11:3502025. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Liu YY, Zhang YY, Ran LY, Huang B, Ren JW,
Ma Q, Pan XJ, Yang FF, Liang C, Wang XL, et al: A novel protein
FNDC3B-267aa encoded by circ0003692 inhibits gastric cancer
metastasis via promoting proteasomal degradation of c-Myc. J Transl
Med. 22:5072024. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Xiong L, Liu HS, Zhou C, Yang X, Huang L,
Jie HQ, Zeng ZW, Zheng XB, Li WX, Liu ZZ, et al: A novel protein
encoded by circINSIG1 reprograms cholesterol metabolism by
promoting the ubiquitin-dependent degradation of INSIG1 in
colorectal cancer. Mol Cancer. 22:722023. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Gao X, Sun Z, Liu X, Luo J, Liang X, Wang
H, Zhou J, Yang C, Wang T and Li J: 127aa encoded by circSpdyA
promotes FA synthesis and NK cell repression in breast cancers.
Cell Death Differ. 32:416–433. 2025. View Article : Google Scholar :
|
|
118
|
Wang J, Wang X, Yang C, Li Q, Li D, Du X,
Cheng Y, Tian M, Zheng L and Tong Q: circE2F1-encoded peptide
inhibits circadian machinery essential for nucleotide biosynthesis
and tumor progression via repressing SPIB/E2F1 axis. Int J Biol
Macromol. 280:1356982024. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Hu H, Tang J, Wang H, Guo X, Tu C and Li
Z: The crosstalk between alternative splicing and circular RNA in
cancer: Pathogenic insights and therapeutic implications. Cell Mol
Biol Lett. 29:1422024. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Zhou C, Molinie B, Daneshvar K, Pondick
JV, Wang J, Van Wittenberghe N, Xing Y, Giallourakis CC and Mullen
AC: Genome-wide maps of m6A circRNAs identify widespread and
cell-type-specific methylation patterns that are distinct from
mRNAs. Cell Rep. 20:2262–2276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Yi Q, Liao Y and Sun W, Li J, Yang D,
Shang H and Sun W: m6A modification of non-coding RNA: Mechanisms,
functions and potential values in human diseases (review). Int J
Mol Med. 56:1642025. View Article : Google Scholar
|
|
122
|
An Y and Duan H: The role of m6A RNA
methylation in cancer metabolism. Mol Cancer. 21:142022. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Wang T, Kong S, Tao M and Ju S: The
potential role of RNA N6-methyladenosine in cancer progression. Mol
Cancer. 19:882020. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Yang Y, Fan X, Mao M, Song X, Wu P, Zhang
Y, Jin Y, Yang Y, Chen LL, Wang Y, et al: Extensive translation of
circular RNAs driven by N6-methyladenosine. Cell Res.
27:626–641. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zeng K, Peng J, Xing Y, Zhang L, Zeng P,
Li W, Zhang W, Pan Z, Zhou C and Lin J: A positive feedback circuit
driven by m6A-modified circular RNA facilitates
colorectal cancer liver metastasis. Mol Cancer. 22:2022023.
View Article : Google Scholar
|
|
126
|
Shan Z and Liu Y: Harnessing glycolysis in
gastric cancer: Molecular targets, therapeutic strategies, and
clinical horizons. Front Immunol. 16:16289372025. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang
Y, Yi C, Lindahl T, Pan T, Yang YG and He C: N6-methyladenosine in
nuclear RNA is a major substrate of the obesity-associated FTO. Nat
Chem Biol. 7:885–887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Wu X, Fang Y, Gu Y, Shen H, Xu Y, Xu T,
Shi R, Xu D, Zhang J, Leng K, et al: Fat mass and
obesity-associated protein (FTO) mediated m6A
modification of circFAM192A promoted gastric cancer proliferation
by suppressing SLC7A5 decay. Mol Biomed. 5:112024. View Article : Google Scholar
|
|
129
|
Liu J and Fang X: Regulation of
hsa_circ_0112136 by m6A demethylase FTO can enhance the malignancy
of gastric cancer via the regulation of the PI3K/AKT/mTOR pathway.
Biotechnol Appl Biochem. 71:1316–1328. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Villa E, Ali ES, Sahu U and Ben-Sahra I:
Cancer cells tune the signaling pathways to empower de novo
synthesis of nucleotides. Cancers (Basel). 11:6882019. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Shi J, Mui S, Yan Y, Liu S, Wen K, He C,
Li H, Liao H, Tao M, Wen J, et al: m6A-modified
circRAPGEF1 interaction with IGF2BP3 promotes hepatocellular
carcinoma progression via reprogramming aspartate metabolism. Adv
Sci (Weinh). 12:e038512025. View Article : Google Scholar
|
|
132
|
Necula L, Matei L, Dragu D, Neagu AI,
Mambet C, Nedeianu S, Bleotu C, Diaconu CC and Chivu-Economescu M:
Recent advances in gastric cancer early diagnosis. World J
Gastroenterol. 25:2029–2044. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Ott JJ, Ullrich A and Miller AB: The
importance of early symptom recognition in the context of early
detection and cancer survival. Eur J Cancer. 45:2743–2748. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Hazelton WD and Luebeck EG:
Biomarker-based early cancer detection: Is it achievable? Sci
Transl Med. 3:109fs92011. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Jacobson BC, Bhatt A, Greer KB, Lee LS,
Park WG, Sauer BG and Shami VM: ACG clinical guideline: Diagnosis
and management of gastrointestinal subepithelial lesions. Am J
Gastroenterol. 118:46–58. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Feng F, Tian Y, Xu G, Liu Z, Liu S, Zheng
G, Guo M, Lian X, Fan D and Zhang H: Diagnostic and prognostic
value of CEA, CA19-9, AFP and CA125 for early gastric cancer. BMC
Cancer. 17:7372017. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Smyth EC, Nilsson M, Grabsch HI, van
Grieken NC and Lordick F: Gastric cancer. Lancet. 396:635–648.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Zhang HD, Jiang LH, Sun DW, Hou JC and Ji
ZL: CircRNA: A novel type of biomarker for cancer. Breast Cancer.
25:1–7. 2018. View Article : Google Scholar
|
|
139
|
Reis-das-Mercês L, Vinasco-Sandoval T,
Pompeu R, Ramos AC, Anaissi AKM, Demachki S, de Assumpção PP, Vidal
AF, Ribeiro-Dos-Santos  and Magalhães L: CircRNAs as potential
blood biomarkers and key elements in regulatory networks in gastric
cancer. Int J Mol Sci. 23:6502022. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Li T, Shao Y, Fu L, Xie Y, Zhu L, Sun W,
Yu R, Xiao B and Guo J: Plasma circular RNA profiling of patients
with gastric cancer and their droplet digital RT-PCR detection. J
Mol Med (Berl). 96:85–96. 2018. View Article : Google Scholar
|
|
141
|
Yang X, Xia J, Peng C and Cai W:
Expression of plasma exosomal circLPAR1 in patients with gastric
cancer and its clinical application value. Am J Cancer Res.
13:4269–4276. 2023.PubMed/NCBI
|
|
142
|
Rao M, Zhu Y, Qi L, Hu F and Gao P:
Circular RNA profiling in plasma exosomes from patients with
gastric cancer. Oncol Lett. 20:2199–2208. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Huang M, He YR, Liang LC, Huang Q and Zhu
ZQ: Circular RNA hsa_circ_0000745 may serve as a diagnostic marker
for gastric cancer. World J Gastroenterol. 23:6330–6338. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Chen S, Li T, Zhao Q, Xiao B and Guo J:
Using circular RNA hsa_circ_0000190 as a new biomarker in the
diagnosis of gastric cancer. Clin Chim Acta. 466:167–171. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Zhao Q, Chen S, Li T, Xiao B and Zhang X:
Clinical values of circular RNA 0000181 in the screening of gastric
cancer. J Clin Lab Anal. 32:e223332018. View Article : Google Scholar
|
|
146
|
Ma Y, Li Z, Ma D, Guo J and Sun W:
Hsa_circ_0003195 as a biomarker for diagnosis and prognosis of
gastric cancer. Int J Clin Oncol. 27:354–361. 2022. View Article : Google Scholar
|
|
147
|
Ma S, Kong S, Gu X, Xu Y, Tao M, Shen L,
Shen X and Ju S: As a biomarker for gastric cancer, circPTPN22
regulates the progression of gastric cancer through the EMT
pathway. Cancer Cell Int. 21:442021. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Shao Y, Yu X, Hu M, Yan J, Miao M, Ye G
and Guo J: Acting mechanism and clinical significance of
hsa_circ_0005927 in the invasion and metastasis of gastric cancer.
J Cancer. 15:4081–4094. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Shao Y, Tao X, Lu R, Zhang H, Ge J, Xiao
B, Ye G and Guo J: Hsa_circ_0065149 is an indicator for early
gastric cancer screening and prognosis prediction. Pathol Oncol
Res. 26:1475–1482. 2020. View Article : Google Scholar
|
|
150
|
Li WH, Song YC, Zhang H, Zhou ZJ, Xie X,
Zeng QN, Guo K, Wang T, Xia P and Chang DM: Decreased expression of
Hsa_circ_00001649 in gastric cancer and its clinical significance.
Dis Markers. 2017:45876982017. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Fang R, Yuan W, Mao C, Cao J, Chen H, Shi
X and Cong H: Human circular RNA hsa_circ_0000231 clinical
diagnostic effectiveness as a new tumor marker in gastric cancer.
Cancer Rep (Hoboken). 7:e20812024.PubMed/NCBI
|
|
152
|
Ma S, Yao Y, Xu Y, Zou M, Zhou M,
Abudushalamu G, Chen Y, Cai S, Zhang C and Wu G: Comprehensive
evaluation of serum circHAS2 as a novel diagnostic and prognostic
biomarker for gastric cancer. Mol Carcinog. 63:94–105. 2024.
View Article : Google Scholar
|
|
153
|
Yuan W, Fang R, Mao C, Chen H, Tai B and
Cong H: Serum circular RNA hsa_circ_0000702 as a novel biomarker
for diagnosis of gastric cancer. J Clin Lab Anal. 37:e248422023.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Zang X, Wang R, Wang Z, Qiu S, Zhang F,
Zhou L, Shen Y, Qian H, Xu W and Jiang J: Exosomal circ50547 as a
potential marker and promotor of gastric cancer progression via
miR-217/HNF1B axis. Transl Oncol. 45:1019692024. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Huang XJ, Wang Y, Wang HT, Liang ZF, Ji C,
Li XX, Zhang LL, Ji RB, Xu WR, Jin JH and Qian H: Exosomal
hsa_circ_000200 as a potential biomarker and metastasis enhancer of
gastric cancer via miR-4659a/b-3p/HBEGF axis. Cancer Cell Int.
23:1512023. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Gareev I, Ahmad A, Wang J, Beilerli A,
Ilyasova T, Sufianov A and Beylerli O: Gastric juice non-coding
RNAs as potential biomarkers for gastric cancer. Front Physiol.
14:11795822023. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Shao Y, Li J, Lu R, Li T, Yang Y, Xiao B
and Guo J: Global circular RNA expression profile of human gastric
cancer and its clinical significance. Cancer Med. 6:1173–1180.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Arnaiz E, Sole C, Manterola L,
Iparraguirre L, Otaegui D and Lawrie CH: CircRNAs and cancer:
Biomarkers and master regulators. Semin Cancer Biol. 58:90–99.
2019. View Article : Google Scholar
|
|
159
|
Roy S, Kanda M, Nomura S, Zhu Z, Toiyama
Y, Taketomi A, Goldenring J, Baba H, Kodera Y and Goel A:
Diagnostic efficacy of circular RNAs as noninvasive, liquid biopsy
biomarkers for early detection of gastric cancer. Mol Cancer.
21:422022. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Xu J, Liu Y, Zhang J, Yang S, Li T, Huang
H, Liu Q, Wang H, Cao L, An Z, et al: Single-droplet dual-target
quantification of circRNA biomarkers for colorectal cancer
screening. Adv Sci (Weinh). 12:e061592025. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Yang L, Wilusz JE and Chen LL: Biogenesis
and regulatory roles of circular RNAs. Annu Rev Cell Dev Biol.
38:263–289. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Szabo L and Salzman J: Detecting circular
RNAs: Bioinformatic and experimental challenges. Nat Rev Genet.
17:679–692. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Porwal O, Visht S and Muhammed RA:
Circular RNAs as biomarkers and targets in ovarian cancer. Clin
Chim Acta. 582:1208022026. View Article : Google Scholar
|
|
164
|
Guria A, Sharma P, Srikakulam N, Baby A,
Natesan S and Pandi G: Cost-effective transcriptome-wide profiling
of circular RNAs by the improved-tdMDA-NGS method. Front Mol
Biosci. 9:8863662022. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Saadh MJ, Ahmed HH, Kareem RA, Bishoyi AK,
Roopashree R, Shit D, Arya R, Joshi KK, Sameer HN, Yaseen A, et al:
The hidden messengers: Tumor microenvironment-derived exosomal
ceRNAs in gastric cancer progression. Pathol Res Pract.
269:1559052025. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Ferda J, Ferdová E, Vítovec M, Glanc D and
Mírka H: The imaging of the hypoxic microenvironment in tumorous
tissue using PET/CT and PET/MRI. Eur J Radiol. 154:1104582022.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Zandieh MA, Farahani MH, Rajabi R, Avval
ST, Karimi K, Rahmanian P, Razzazan M, Javanshir S, Mirzaei S,
Paskeh MDA, et al: Epigenetic regulation of autophagy by non-coding
RNAs in gastrointestinal tumors: Biological functions and
therapeutic perspectives. Pharmacol Res. 187:1065822023. View Article : Google Scholar
|
|
168
|
Barati T, Mirzaei Z, Abarghooi Kahaki F
and Afkhami H: Regulatory roles of ncRNAs in gastric cancer
progression: A comprehensive insight into key signaling pathways.
Med Oncol. 42:3092025. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Liu Y, Jiang Y, Xu L, Qu C, Zhang L, Xiao
X, Chen W, Li K, Liang Q and Wu H: circ-NRIP1 promotes glycolysis
and tumor progression by regulating miR-186-5p/MYH9 axis in gastric
cancer. Cancer Manag Res. 12:5945–5956. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Xu G, Li M, Wu J, Qin C, Tao Y and He H:
Circular RNA circNRIP1 sponges microRNA-138-5p to maintain
hypoxia-induced resistance to 5-fluorouracil through
HIF-1α-dependent glucose metabolism in gastric carcinoma. Cancer
Manag Res. 12:2789–2802. 2020. View Article : Google Scholar
|
|
171
|
Cordani M, Dando I, Ambrosini G and
González-Menéndez P: Signaling, cancer cell plasticity, and
intratumor heterogeneity. Cell Commun Signal. 22:2552024.
View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Golestanifar A, Zakeri M, Gohari-Lasaki S,
Khedri H and Saberiyan M: Combined bulk and single-cell
transcriptomic analysis reveals cell-type-specific inflammatory
crosstalk in pancreatic cancer. Clin Exp Med. 25:2632025.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Li Y, Xu Q, Wu D and Chen G: Exploring
additional valuable information from single-cell RNA-Seq data.
Front Cell Dev Biol. 8:5930072020. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Aquino-Jarquin G: CircRNA knockdown based
on antisense strategies. Drug Discov Today. 29:1040662024.
View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Zhou Y, Zhang Q, Liao B, Qiu X, Hu S and
Xu Q: circ_0006089 promotes gastric cancer growth, metastasis,
glycolysis, and angiogenesis by regulating miR-361-3p/TGFB1. Cancer
Sci. 113:2044–2055. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Jiang SF and Li RR: hsa_circ_0067514
suppresses gastric cancer progression and glycolysis via
miR-654-3p/LATS2 axis. Neoplasma. 69:1079–1091. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Shen X, Zhu X, Hu P, Ji T, Qin Y and Zhu
J: Knockdown circZNF131 inhibits cell progression and glycolysis in
gastric cancer through miR-186-5p/PFKFB2 axis. Biochem Genet.
60:1567–1584. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Yang X, Li D, Sun Y, Yi L, Chen Q and Lai
Y: CircFLNA facilitates gastric cancer cell proliferation and
glycolysis via regulating SOX5 by sponging miR-1200. Arab J
Gastroenterol. 25:369–377. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Wang Y, Cao B, Zhao R, Li H, Wei B and Dai
G: Knockdown of circBFAR inhibits proliferation and glycolysis in
gastric cancer by sponging miR-513a-3p/hexokinase 2 axis. Biochem
Biophys Res Commun. 560:80–86. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Zheng X, Shao J, Qian J and Liu S:
circRPS19 affects HK2-mediated aerobic glycolysis and cell
viability via the miR-125a-5p/USP7 pathway in gastric cancer. Int J
Oncol. 63:982023. View Article : Google Scholar
|
|
181
|
Chen L, Chi K, Xiang H and Yang Y:
Circ_0032821 facilitates gastric cancer cell proliferation,
migration, invasion and glycolysis by regulating MiR-1236-3p/HMGB1
axis. Cancer Manag Res. 12:9965–9976. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Liu J, Li J, Su Y, Ma Z, Yu S and He Y:
Circ_0009910 serves as miR-361-3p sponge to promote the
proliferation, metastasis, and glycolysis of gastric cancer via
regulating SNRPA. Biochem Genet. 60:1809–1824. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Zheng X, Xiao H, Liu X, Huang T and Deng
C: Exosomal circKIAA1797 regulates cell progression and glycolysis
by targeting miR-4429/PBX3 pathway in gastric cancer. Biochem
Genet. 62:1762–1778. 2024. View Article : Google Scholar
|
|
184
|
Zhao X, Tian Z and Liu L: circATP2B1
promotes aerobic glycolysis in gastric cancer cells through
regulation of the miR-326 gene cluster. Front Oncol. 11:6286242021.
View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Chen W and Ji Y: CircC6orf132 facilitates
proliferation, migration, invasion, and glycolysis of gastric
cancer cells under hypoxia by acting on the miR-873-5p/PRKAA1 axis.
Front Genet. 12:6363922021. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Yang J, Zhang X, Cao J, Xu P, Chen Z, Wang
S, Li B, Zhang L, Xie L, Fang L and Xu Z: Circular RNA UBE2Q2
promotes malignant progression of gastric cancer by regulating
signal transducer and activator of transcription 3-mediated
autophagy and glycolysis. Cell Death Dis. 12:9102021. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Liu J, Liu H, Zeng Q, Xu P, Liu M and Yang
N: Circular RNA circ-MAT2B facilitates glycolysis and growth of
gastric cancer through regulating the miR-515-5p/HIF-1α axis.
Cancer Cell Int. 20:1712020. View Article : Google Scholar
|
|
188
|
Ou W, Tan R, Zhai J, Sun L, Quan Z, Huang
X, Xu F, Xu Q and Zhou C: Silencing circ_0043256 inhibited
CoCl2-induced proliferation, migration, and aerobic glycolysis in
gastric cancer cells. Sci Rep. 15:1712025. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Lu Y, Cheng J, Cai W, Zhuo H, Wu G and Cai
J: Inhibition of circRNA circVPS33B reduces warburg effect and
tumor growth through regulating the miR-873-5p/HNRNPK axis in
infiltrative gastric cancer. Onco Targets Ther. 14:3095–3108. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Qu J, Yang J, Chen M, Wei R and Tian J:
CircFLNA acts as a sponge of miR-646 to facilitate the
proliferation, metastasis, glycolysis, and apoptosis inhibition of
gastric cancer by targeting PFKFB2. Cancer Manag Res. 12:8093–8103.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Lan ZZ, Sun FH, Chen C, Niu L, Shi JD and
Zhang WY: CircPRDM5 inhibits the proliferation, migration,
invasion, and glucose metabolism of gastric cancer cells by
reducing GCNT4 expression in a miR-485-3p-dependent manner.
Kaohsiung J Med Sci. 40:231–243. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Lu J, Zhou Y, Chen Z, Jiang H, Li J and
Dou G: Circ_0000419 acts as a tumor suppressor in gastric cancer
development via regulating miR-300/RGMB axis. Int J Clin Oncol.
28:1475–1485. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Wu G, Zhang A, Yang Y and Wu D:
Circ-RNF111 aggravates the malignancy of gastric cancer through
miR-876-3p-dependent regulation of KLF12. World J Surg Oncol.
19:2592021. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Ji Z, Diao W and Shang J: Circular RNA
circ_0000592 elevates ANXA4 expression via sponging miR-1179 to
facilitate tumor progression in gastric cancer. Anticancer Drugs.
33:e644–e654. 2022. View Article : Google Scholar
|
|
195
|
Pu Z, Xu M, Yuan X, Xie H and Zhao J:
Circular RNA circCUL3 accelerates the warburg effect progression of
gastric cancer through regulating the STAT3/HK2 axis. Mol Ther
Nucleic Acids. 22:310–318. 2020. View Article : Google Scholar : PubMed/NCBI
|