Introduction
Cancer remains a leading cause of mortality
worldwide, with the World Health Organization reporting nearly 10
million deaths in 2020, accounting for ~1 in 6 fatalities globally
(1). Despite the efficacy of
conventional therapies in markedly reducing tumor burden,
recurrence and metastasis frequently occur, ultimately resulting in
treatment failure. A major factor contributing to this failure is
the inadequate consideration or neglect of the tumor
microenvironment (TME) in antitumor strategies (2). The TME comprises various immune cell
populations, cancer-associated fibroblasts (CAFs), endothelial
cells, pericytes and a diverse array of other tissue-resident cell
types. Historically regarded as passive bystanders in
tumorigenesis, these host cells are now recognized as critical
players in cancer pathogenesis. The cellular composition and
functional states of the TME vary markedly depending on the organ
of tumor origin, intrinsic characteristics of cancer cells, tumor
stage and patient-specific factors (3). Among TME components, CAFs are
pivotal in cancer initiation and progression, owing to their
multifaceted roles in extracellular matrix (ECM) remodeling,
maintenance of stemness properties, angiogenesis, modulation of
tumor metabolism and immune responses, as well as their promotion
of cancer cell proliferation, migration, invasion and therapeutic
resistance. CAFs represent a highly heterogeneous stromal
population, engaging in complex and intricate crosstalk with cancer
cells mediated by an array of signaling pathways, including
transforming growth factor-β (TGF-β), phosphatidylinositol 3-kinase
(PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR),
mitogen-activated protein kinase (MAPK), Wnt, Janus kinase
(JAK)/signal transducer and activator of transcription (STAT),
epidermal growth factor receptor (EGFR), Hippo and nuclear factor
κB (NF-κB) pathways. These signaling cascades give rise to distinct
CAF phenotypes during tumor progression and represent promising
targets for anticancer therapy (4).
Furthermore, fibroblasts display phenotypic
plasticity influenced by their microenvironmental context.
Activated fibroblasts within tumors are termed CAFs (5,6).
Distinct from normal fibroblasts, CAFs exhibit differential
expression of markers such as α-smooth muscle actin (α-SMA),
fibroblast activation protein (FAP), fibroblast-specific protein 1
and platelet-derived growth factor receptor (PDGFR) (7). In addition to these, proteins
including collagen type XI α1 chain (COL11A1),
microfibril-associated protein 5 and asporin are predominantly
expressed in CAFs (8,9). The concept of intratumoral CAF
heterogeneity recognizes the existence of distinct CAF subsets,
such as inflammatory CAFs characterized by low α-SMA and high
interleukin-6 (IL-6) expression, contrasting with TGF-β-dependent
myofibroblastic CAFs exhibiting high α-SMA levels (10).
A novel therapeutic approach targeting CAFs is the
'stromal reprogramming' strategy, which aims to convert
tumor-promoting CAFs into tumor-inhibitory phenotypes by reducing
matrix stiffness (11). Despite
recent advances, several molecular mechanisms underpinning CAF
biology remain to be fully elucidated, posing challenges to the
development of personalized anticancer therapies targeting the
stroma. Accordingly, strategies that concurrently address both the
tumor stroma and cancer cells have garnered considerable attention
in the context of precision oncology. Collectively, CAFs constitute
a predominant cellular component within the TME and play a pivotal
role in regulating tumor biological behavior (12). However, most current tumor
therapies primarily target the proliferative tumor cells and exert
limited effects on CAFs. This therapeutic gap partially explains
why reductions in tumor volume do not invariably translate into
improved patient survival outcomes.
Due to their diverse origins and complex activation
mechanisms, CAFs are recognized as critical modulators of
tumorigenesis, tumor progression and therapeutic resistance
(13). Consequently, specific
interventions targeting CAFs hold promise for enhancing anticancer
efficacy and their combination with conventional therapies may
potentially achieve curative results. The biological functions of
CAFs are orchestrated by various intracellular and extracellular
factors, notably signaling pathways intimately involved in cancer
progression, which present viable targets for therapeutic
intervention. Currently, signaling cascades including FAP,
PI3K/AKT/mTOR, JAK/STAT, NF-κB and TGF-β, as well as pathways
regulating ferroptosis, apoptosis and autophagy, are extensively
investigated for mediating the crosstalk between CAFs and cancer
cells, thereby serving as potential targets for cancer therapy
(14).
Of particular interest, the induction of
ferroptosis, apoptosis and autophagy in CAFs has recently emerged
as a novel mechanistic approach, demonstrating promising
therapeutic potential (15).
Integrating conventional treatments such as chemotherapy or
radiotherapy with CAFs-targeted agents may result in improved
inhibition of tumor initiation, growth, progression, recurrence and
metastasis. The present review summarized the current landscape of
small-molecule compounds that suppress CAF activity, focusing on
inhibitors of the PI3K/AKT/mTOR, JAK/STAT, TGF-β, ferroptosis and
autophagy pathways, as well as activators of apoptotic signaling.
These insights may provide a valuable foundation for the
development of enhanced therapeutic strategies in cancer
management.
Signaling pathway regulators
CAFs, a major component of the TME, have been found
to be involved in various cellular processes. Their biological
activity is mediated by multiple intracellular and extracellular
factors, particularly cancer-associated signaling cascades, which
may serve as targets for anticancer therapy. Among these signaling
pathways, PI3K/AKT/mTOR, JAK/STAT, NF-κB and TGF-β (14) have been extensively studied for
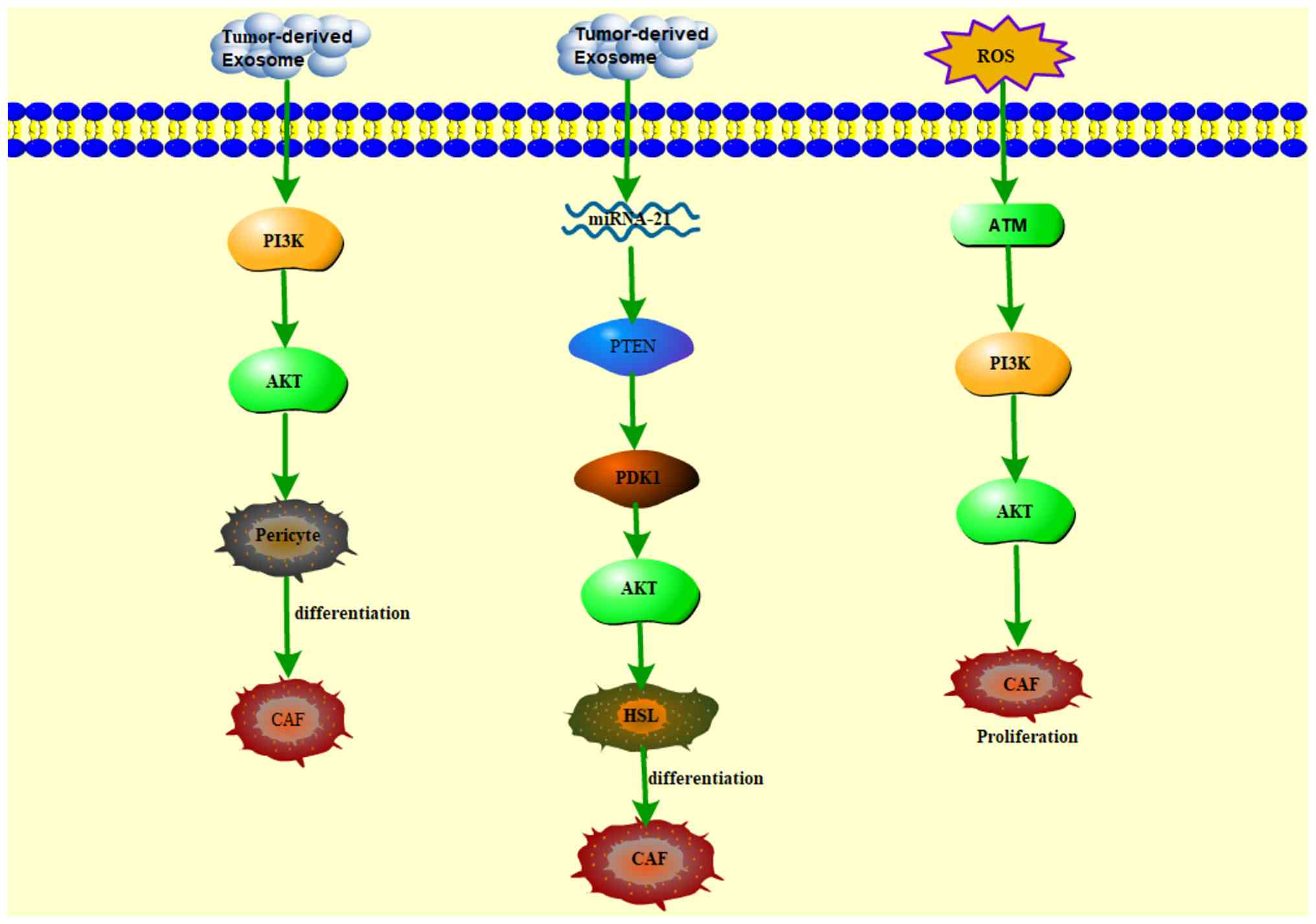
the development of new drugs targeting CAFs (Fig. 1).
 | Figure 1Small molecule agents inhibit the
proliferation, differentiation and activation of CAFs by inhibiting
PI3K/AKT/mTOR signaling pathways, TGF-β pathway and NF-κB. CAFs,
cancer-associated fibroblasts; PI3K, phosphatidylinositol 3-kinase;
AKT, protein kinase B; mTOR, mechanistic target of rapamycin;
TGF-β, transforming growth factor β; NF-κB, nuclear factor κB; ATM,
ataxia telangiectasia-mutated genataxia telangiectasia-mutated;
PIP3, phosphatidylinositol 3 trisphosphate; PDK1, pyruvate
dehydrogenase kinase 1; SMAD, mothers against DPP homolog; TLR4,
Toll-like receptors 4; MyD88, myeloid differentiation primary
response 88; IL-6, interleukin-6. |
PI3K/AKT/mTOR signaling pathway
inhibitors
The PI3K/AKT/mTOR signaling pathway is intimately
involved in multiple facets of cancer biology, including the
proliferation, differentiation, growth, apoptosis and migration of
CAFs (16). PI3K consists of a
regulatory subunit (p85) and a catalytic subunit (p110). Upon
engagement with growth factor receptors such as EGFR, PI3K
undergoes conformational changes that lead to the activation of
Akt. Activated Akt subsequently phosphorylates downstream
substrates, including apoptosis-related proteins Bad and Caspase-9,
thereby modulating cellular processes such as proliferation,
differentiation, apoptosis and migration. The mTOR, a key
downstream effector of the PI3K/Akt pathway, is activated when Akt
directly phosphorylates its Ser1448 residue. Activation of mTOR and
its downstream signaling cascades regulates the translation of
specific proteins essential for cell proliferation and
transformation (17). The
PI3K/AKT pathway has a critical role in CAF proliferation (Fig. 2), which is often driven by
oxidative stress; a fundamental contributor to aberrant CAF
expansion. Ataxia-telangiectasia mutated protein kinase, a pivotal
redox sensor, is induced by PI3K/AKT pathway activation, thereby
promoting the abnormal proliferation of breast CAFs (18). Furthermore, accumulating evidence
indicates that the PI3K/AKT signaling axis facilitates the
differentiation of various precursor cell types into CAFs. For
instance, exosomes derived from gastric cancer cells induce the
transdifferentiation of pericytes into CAFs via activation of the
PI3K/AKT pathway (19).
Additionally, hepatocellular carcinoma (HCC)-derived exosomal micro
(mi) RNA-21 transforms hepatic stellate cells into CAFs through the
Akt signaling pathway (20).
Additionally, osteosarcoma cells stimulate the differentiation of
mesenchymal stem cells into CAFs through Akt-dependent pathways,
evidenced by increased expression of the CAF marker α-SMA (21). Given these roles, targeting
components of the PI3K/AKT pathway represents a promising
therapeutic strategy to inhibit the function of CAFs. Multiple
studies have demonstrated that inhibitors targeting the
PI3K/AKT/mTOR axis, whether administered alone or in combination
with chemotherapeutic agents, effectively suppress CAF activity and
consequently impede tumor progression (Table I) (22-35).
 | Table ISmall-molecule compounds inhibiting
CAF-induced cancer through suppressing PI3K/AKT/mTOR signaling
pathways. |
Table I
Small-molecule compounds inhibiting
CAF-induced cancer through suppressing PI3K/AKT/mTOR signaling
pathways.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| Zhang, 2015 | Dovitinib with
LY294002 or RAD001 | PI3K/AKT/mTOR | Breast cancer | Preclinical | Inhibited CAF
invasion | (22) |
| Li, 2023 | Rocuronium | PI3K/AKT/mTOR | Esophageal
cancer | Preclinical | Inhibited CAF
secretion of the pro-tumor cytokine CXCL12 | (23) |
| Lu and Zhang,
2023 | Oxaliplatin | CXCR3/PI3K/AKT | Colorectal
cancer | Preclinical | Reduced cancer cell
proliferation | (24) |
| Chen, 2023 | Biejiajian
pill | PI3K/AKT | Hepatocellular
carcinoma | Preclinical | Reduced expression
of VEGF-A and HGF by CAF | (25) |
| Li, 2020 | Cisplatin | AKT | Lung cancer | Preclinical | Attenuated CAF
metastasis-promoting effects | (26) |
| Che, 2018 | Tiplaxtinin | AKT | Esophageal squamous
cell carcinoma | Preclinical | Inhibited tumor
growth and improved CAF-induced tumor microenvironment | (27) |
| Zhao, 2018 | Gold
nanoparticles | AKT | Colorectal
cancer | Preclinical | Enhanced cisplatin
delivery and decompression of cancer vessels | (28) |
| Zhu, 2021 | Capmatinib | AKT | NSCLC | Preclinical | Reduced production
of CAF | (29) |
| Heits, 2016 | Mycophenolic acid
and Everolimus | mTOR; AKT | Cholangiocellular
carcinoma | Preclinical | Reduced migration
and invasive activity of CAF | (30) |
| Zhang, 2022 | Regorafenib | AKT | Gastrointestinal
cancer | Preclinical | Induced apoptosis
in gastrointestinal CAF | (31) |
| Tang, 2024 | Anlotinib | AKT | NSCLC | Preclinical | Promoted CAF
apoptosis | (32) |
| Al-Ansariand
Aboussekhra, 2014 | Caffeine | AKT | Breast cancer | Preclinical | Induced sustained
inactivation of breast CAF | (33) |
| Fan, 2017 |
Dihydromyricetin | AKT | Lung cancer | Preclinical | Inhibited
proliferation of lung CAF | (34) |
| Yadav, 2020 | MSI-N1014 | mTOR | Colorectal
cancer | Preclinical | Reduced conversion
of cancer cells to CAF | (35) |
For example, the combination treatment of dovitinib
with the PI3K/AKT/mTOR signaling inhibitors LY294002 or RAD001
resulted in additive inhibition of CAF invasion, demonstrating
therapeutic activity against breast cancer metastasis (22). Rocuronium bromide, a
non-depolarizing neuromuscular blocker, has been shown to inhibit
the PI3K/AKT/mTOR signaling pathway, thereby blocking CAFs and
attenuating esophageal cancer progression (23). Oxaliplatin, a classic
third-generation platinum-based chemotherapeutic agent, inhibits
xenograft tumor growth in mouse xenograft models by targeting the
C-X-C chemokine motif receptor 3 (CXCR3)/PI3K/AKT axis secreted by
CAFs (24).
The classic Chinese medicine formula Biejiajian pill
has been found to inhibit HCC progression by suppressing PI3K and
AKT phosphorylation in a mouse model of diethylnitrosamine/carbon
tetrachloride-induced HCC (25).
Inhibition of AKT phosphorylation is a key strategy to target CAFs.
For instance, cisplatin, a commonly used anticancer drug, has been
shown to reduce the ability of CAFs to promote lung cancer cell
migration and invasion by inhibiting the AKT signaling pathway both
in tumor cell models and in BALB/c nude mice tumor models (26). Cisplatin inadvertently induces
CAFs to secrete plasminogen activator inhibitor-1 (PAI-1) and this
paracrine cue fosters tumor progression while diminishing cisplatin
response. Targeting PAI-1 with Tiplaxtinin inhibits AKT activation
in CAFs and enhances cisplatin efficacy, producing in vitro
and in vivo synergy (27).
Similarly, gold nanoparticles can enhance the
effects of cisplatin by decreasing the density of CAFs through the
Akt signaling pathway and reducing collagen I production in
colorectal cancer (CRC) xenograft mice (28). Furthermore, the combination of
camatinib and osimertinib markedly enhances tumor suppression and
decreases CAF abundance in patient-derived xenograft models via AKT
pathway inhibition in non-small cell lung cancer (NSCLC) (29). Mesalazine and the mTOR inhibitor
everolimus, originally developed as immunosuppressants to prevent
organ rejection, have recently been found to exert
anti-proliferative effects on cholangiocarcinoma cells by blocking
CAF-activated AKT signaling (30). Regorafenib, a multikinase
inhibitor, has been shown to inhibit proliferation and induce
apoptosis in CAFs in vitro, with its mechanism linked to the
inhibition of AKT phosphorylation (31).
In NSCLC models, anlotinib was demonstrated to
modulate the TME by inhibiting the AKT pathway and promoting CAF
apoptosis, thereby enhancing therapeutic efficacy against NSCLC
(32). Furthermore, bioactive
compounds isolated from commonly consumed beverages exhibit
CAF-inhibitory effects via AKT suppression. For instance, caffeine
(1,3,7-trimethylxanthine), the most widely consumed psychoactive
substance worldwide, inhibits CAF migration and invasion through
phosphatase and tensin homolog-dependent inactivation of Akt and
Erk1/2 signaling, effectively preventing breast tumor growth and
recurrence in a safe manner (33). Dihydromyricetin, a flavonoid that
is the main active ingredient in Garcinia cambogia, has been
found to inhibit the proliferative potential of CAFs by targeting
Akt activation (34).
Additionally, researchers synthesized a drug targeting mTOR,
tetracyclic heterocyclic azathiophenone MSI-N1014, which was able
to inhibit the transformation of CAFs by decreasing mTOR
expression, thus markedly reducing migratory ability, tumor-balloon
generation and resistance to 5-fluorouracil (35).
In conclusion, current studies on regulating CAFs by
inhibiting the PI3K/AKT/mTOR pathway primarily focus on suppressing
the functions and activities of CAFs through this pathway. These
include inhibiting CAF proliferation, promoting CAF apoptosis and
reducing the secretion of pro-tumorigenic factors by CAFs. Most
approaches utilize small molecule inhibitors or genetic
interventions, indicating the critical role of the PI3K/AKT/mTOR
signaling in modulating CAF activity. However, given the broad
involvement of this pathway in diverse cellular processes, highly
selective interventions targeting CAFs specifically are still
lacking and treatment often carries significant systemic side
effects. Overall, among the strategies targeting CAFs via the
PI3K/AKT/mTOR pathway, the application of small molecule inhibitors
is the most mature and has demonstrated considerable efficacy in
suppressing CAF function. Nevertheless, these approaches
predominantly focus on direct depletion or inactivation of CAFs,
with relatively limited emphasis on stromal reprogramming. A subset
of studies aiming for selective blockade of specific PI3K isoforms
or downstream effectors has shown promising improvements in CAF
selectivity, suggesting potential advantages. Furthermore,
strategies that induce stromal remodeling through modulation of
this pathway remain in the early stages of investigation; combining
pathway inhibition with stromal reprogramming could yield more
durable therapeutic outcomes in the future. In terms of clinical
translation, most research remains at the preclinical stage,
lacking comprehensive clinical validation and safety evaluation.
Future efforts should prioritize the development of highly
selective PI3K/AKT/mTOR modulators in conjunction with multimodal
treatment strategies to optimize CAF-targeted therapies, ultimately
achieving more effective antitumor responses with reduced
off-target toxicity.
JAK-STAT signaling pathway
inhibitors
The JAK-STAT signaling pathway consists of three
components: The tyrosine kinase-associated receptor that receives
the signal, the tyrosine kinase JAK that transmits the signal and
the transcription factor STAT that produces the effect. When
various cytokines or growth factors bind to receptors, they
phosphorylate and activate JAK. JAK then phosphorylates tyrosine
residues on downstream target proteins and recruits and
phosphorylates the transcription factor STAT (36). STAT forms dimers and enters the
nucleus, where it binds to target genes and regulates their
transcription. This process influences cell proliferation,
differentiation and apoptosis (37). The JAK/STAT signaling pathway is
constitutively activated in CAFs. For instance, CAFs can promote
endometrial cancer growth by inducing the IL-6-activated STAT-3
pathway (38).
Similarly, in prostate cancer, CAF-derived IL-6 can
attenuate p53 activity by activating the JAK/STAT signaling
pathway, thereby protecting cancer cells from chemotherapy
(39). Furthermore, in pancreatic
ductal adenocarcinoma (PDAC), activation of the JAK/STAT signaling
pathway induced by IL-1 also produces inflammatory CAFs, which
promote tumor growth (40). In
summary, CAF-induced cytokines activate the JAK/STAT signaling
pathway and the activated JAK/STAT signaling pathway in turn
produces more CAFs, ultimately promoting tumorigenesis and
progression. Therefore, inhibition of the JAK2/STAT3 signaling
pathway may be a crucial strategy to suppress cancer stemness,
plasticity and intercellular signaling between CAFs and cancer
cells (Table II) (41-49).
| Table IISmall-molecule compounds inhibiting
CAF-induced cancer through suppressing JAK-STAT signaling
pathways. |
Table II
Small-molecule compounds inhibiting
CAF-induced cancer through suppressing JAK-STAT signaling
pathways.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| Wang, 2023 | Bufalin | STAT3 | Colorectal
cancer | Preclinical | Reversed
CAF-mediated cancer invasion and metastasis | (41) |
| Ochi, 2022 | Tranilast | STAT3 | Non-small cell lung
cancer | Preclinical | Reversed CAF
resistance to molecularly targeted drugs | (42) |
| Al-Jomah, 2021 | Tocilizumab | STAT3 | Breast cancer | Preclinical | Inhibited
proliferation, migration and invasive capacity of activated breast
CAF | (43) |
| Ham, 2022 | Curcumin | JAK/STAT3 | Gastric cancer | Preclinical | Counteracted
CAF-induced chemotherapy resistance | (44) |
| Cao, 2020 | Oroxylin A | STAT3 | Breast cancer | Preclinical | Prevented the
activation of CAF | (45) |
| Suh,2018 | Resveratrol | STAT3 | Breast cancer | Preclinical | Inhibited
CAF-induced migration, invasion and self-renewal activity of breast
cancer cells | (46) |
| Tsang, 2013 | Berberine | STAT3 | Nasopharyngeal
carcinoma | Preclinical | Regulated the
CAF-induced inflammatory tumor microenvironment | (47) |
| Zhang, 2023 | Lentinus
edodes polysaccharide | JAK2/STAT3 | Prostate
cancer | Preclinical | Inhibited
CAF-induced drug resistance | (48) |
| Lee, 2020 | β-Carotene | STAT3 | Colorectal
cancer | Preclinical | Prevented the
activation of CAFs | (49) |
Bufalin, the main active monomer of the clinical
drug cinobufacini, has garnered increasing attention for its
antitumor activity in various cancers. Research has shown that
bufalin can reverse CAF-mediated CRC metastasis by inhibiting the
STAT3 signaling pathway (41).
Trinilast, an anti-allergic drug that inhibits cytokine release in
various inflammatory cells, was recently found to reverse
CAF-mediated resistance of NSCLC cells to molecularly targeted
drugs both in vitro and in vivo when co-administered
with molecularly targeted therapy. This effect is related to the
inhibition of CAF-induced upregulation of phosphorylated STAT by
trinilast (42). Tocilizumab, a
recombinant humanized monoclonal antibody approved by the Food and
Drug Administration (FDA) for treating various immune disorders,
normalizes active breast CAFs and inhibits their paracrine
pro-oncogenic effects by blocking the STAT3 pathway (43).
Curcumin, extracted from the root of the herb
turmeric, has been found to inhibit CAF-mediated activation of the
JAK/STAT3 signaling pathway, thereby overcoming chemoresistance
(44). Additionally, Oroxylin A
and resveratrol, both extracted from herbs, have been shown to
prevent CAF activation by inhibiting STAT3 phosphorylation
(45,46). Similarly, berberine, another
compound extracted from Chinese herbs, effectively inhibited
CAF-induced STAT3 activation, thereby suppressing the
tumorigenicity and growth of nasopharyngeal carcinoma cells
(45). A novel mushroom
polysaccharide, MPSSS, extracted from food, was shown to be an
adjuvant treatment for prostate cancer by inhibiting the JAK2/STAT3
pathway in CAFs (48).
β-Carotene, isolated from carrots, inhibited CAF activation by
suppressing the IL-6/STAT3 signaling pathway, thus demonstrating
its therapeutic potential in cancer progression and metastasis
(49).
Current research targeting CAFs through inhibition
of the JAK-STAT signaling pathway not only focuses on suppressing
CAF activity and function but also addresses overcoming CAF-driven
drug resistance and modulating tumor stromal remodeling.
Representative agents include Oroxylin A, Resveratrol and
Berberine; however, these interventions generally lack high
specificity toward CAFs, limiting therapeutic precision and
potentially causing off-target effects. Most studies remain at the
preclinical stage, providing foundational insights for future
clinical translation. Among these approaches, small molecule
natural compounds are the most mature, demonstrating effectiveness
in inhibiting CAF activation and reversing resistance mechanisms.
Importantly, some evidence indicates that such treatments do not
merely deplete CAF populations but also induce functional
reprogramming of the tumor stroma, broadening therapeutic
possibilities. From a translational perspective, these strategies
are still in their infancy, highlighting the need to develop more
selective JAK-STAT modulators targeting CAFs and prioritize
therapies promoting stromal remodeling rather than simple
elimination of CAFs, thereby facilitating the safe and effective
transition of preclinical findings into clinical applications.
TGF-β signaling pathway inhibitors
The TGF-β signaling pathway is a superfamily of
structurally related multifunctional cytokines, including TGF-β
proteins, activins, inhibins, bone morphogenetic proteins (BMPs)
and growth and differentiation factors. TGF-β signaling is closely
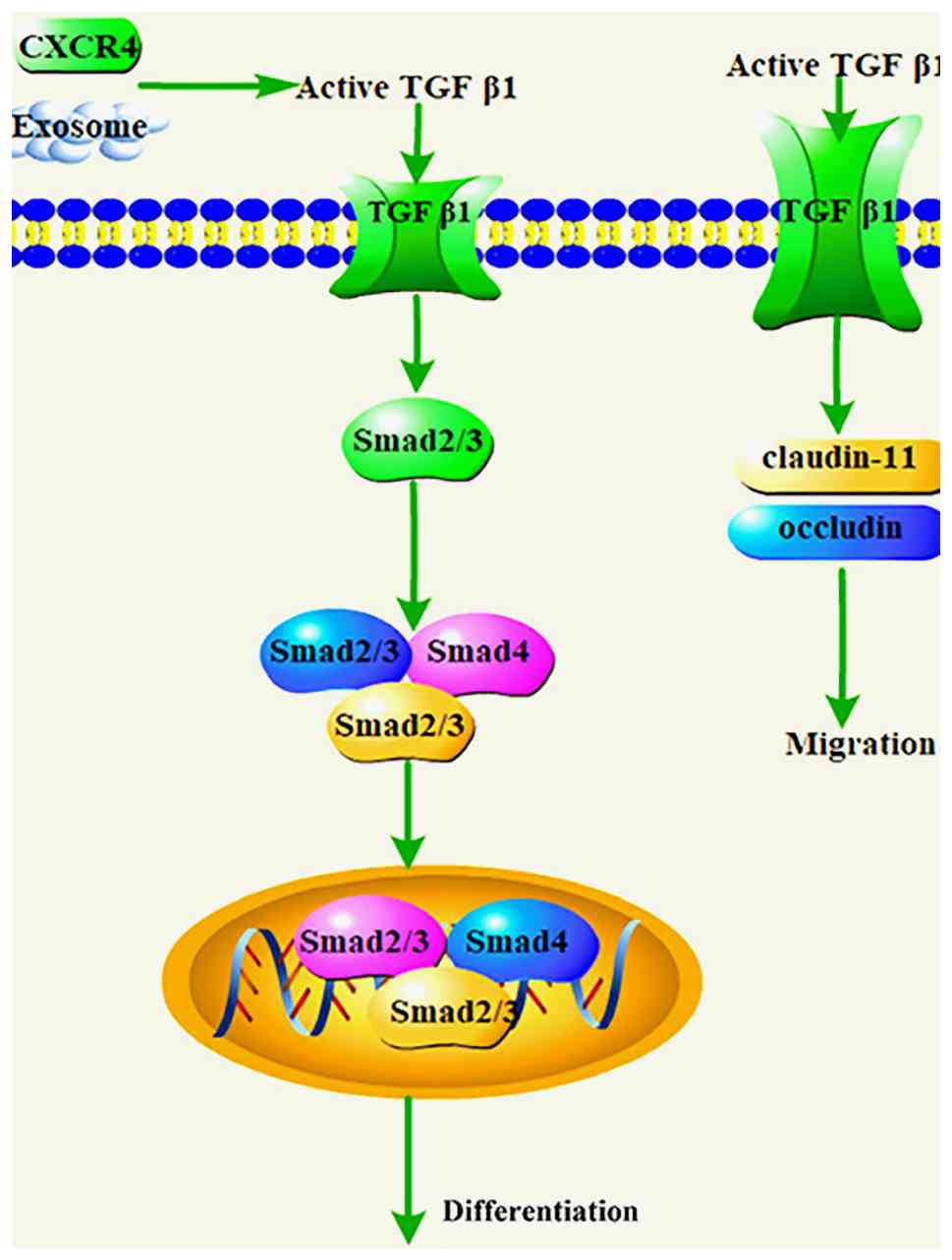
associated with CAF activation or transformation (Fig. 3). For example, bladder cancer
cell-derived exosomes have been found to induce the differentiation
of fibroblasts into CAFs via TGF-β signaling (50). Similarly, hepatic stellate cells
can differentiate into CAFs through the activation of the TGF-β1
signaling pathway, promoting liver metastasis in colon cancer
(51). In ovarian cancer, COL11A1
promotes tumor formation and CAF activation through the activation
of the TGF-β3 signaling pathway (52). Additionally, the prostate cancer
microenvironment can recruit and differentiate mesenchymal stromal
cells into CAFs via TGF-β1 (53).
Activation of TGF-β signaling is also associated with CAF
migration. Evidence suggests that TGF-β signaling activation
promotes the collective migration of CAFs by overexpressing tight
junction-associated proteins claudin-11 and occluding (54). Furthermore, the CAF-mediated TGF-β
pathway promotes cancer progression by regulating cancer cell
proliferation, migration, invasion and metastasis (55). Overall, TGF-β signaling and CAFs
complement each other: TGF-β signaling influences CAF activity,
while CAFs promote cancer progression through TGF-β signaling.
Therefore, targeted inhibition of the TGF-β signaling pathway may
be an effective strategy to suppress cancer progression and CAFs
activation (Table III)
(56-79).
| Table IIISmall-molecule compounds inhibiting
CAF-induced cancer through suppressing TGF-β signaling
pathways. |
Table III
Small-molecule compounds inhibiting
CAF-induced cancer through suppressing TGF-β signaling
pathways.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| Luongand Cukierman,
2022 | Eribulin | TGF-β | Pancreatic
cancer | Preclinical | Normalized
pancreatic CAFs | (56) |
| Lv, 2018 | HA-PTX/MATT-LTSLs
HNPs | TGF-β1 | Breast cancer | Preclinical | Blocked CAF
activation | (57) |
| Zhang, 2018 | LY2157299
monohydrate | TGF-β1 | Ovarian cancer | Preclinical | Blocked CAF
activation | (58) |
| Liu, 2016 | LY2157299 | TGF-β1 | Breast cancer | Preclinical | Inhibited the
formation of CAFs | (59) |
| Wang, 2024 | Sulfosuccinimidyl
oleate sodium | TGF-β1 | Hepatocellular
carcinoma | Preclinical | Inhibited CAF
proliferation | (60) |
| Wawro, 2019 | Aspirin and
ibuprofen | TGF-βs | Colon cancer | Preclinical | Inhibited the
formation of CAFs | (61) |
| Shao, 2020 | Metformin | TGF-β | Breast cancer | Preclinical | Inhibited tumor
invasion | (62) |
| Coleman, 2016 | Digoxin | TGF-β | Others | Preclinical | Inhibited
differentiation of CAFs | (63) |
| Takai, 2016 | Pirfenidone | TGF-β | Breast cancer | Preclinical | Delayed
transdifferentiation of fibroblasts to CAFs | (64) |
| Mediavilla-Varela,
2016 | Pirfenidone | TGF-β1 | Non-small cell lung
cancer | Preclinical | Killed CAFs | (65) |
| Mazzocca, 2010 | LY2109761 | TGF-β | Hepatocellular
carcinoma | Preclinical | Inhibited CAF
proliferation | (66) |
| Wang, 2022 | Halofuginone | TGF-β | Oral squamous cell
carcinoma | Preclinical | Inhibited CAF
proliferation | (67) |
| Gabasa, 2017 | Nintedanib | TGF-β1 | Lung
adenocarcinoma | Preclinical | Blocked CAF
activation | (68) |
| Shangguan,
2012 | Bone morphogenetic
protein and activin membrane-bound inhibitor | TGF-β | Others | Preclinical | Blocked
differentiation of human mesenchymal stem cells to CAFs | (69) |
| Yao, 2019 | Artemisinin
derivatives | TGF-β | Breast cancer | Preclinical | Blocked CAF
activation | (70) |
| Milián, 2022 |
Δ9-Tetrahydrocannabinol and
Cannabidiol | TGF-β | Lung cancer | Preclinical | Inhibited CAF
proliferation | (71) |
| Melegová, 2022 | Aesculus
hippocastanum L. extract | TGF-β1 | Basal cell
carcinoma; squamous cell carcinoma | Preclinical | Improved
CAF-induced tumor microenvironment | (72) |
| Buhrmann, 2014 | Curcumin | TGF-β | Colon cancer | Preclinical | Improved
CAF-induced tumor microenvironment | (73) |
| Liang, 2023 | Chelerythrine
chloride | TGF-β2 | Colon cancer | Preclinical | Blocked CAF
activation | (74) |
| Jalilian, 2023 | Curcumin | TGF-β | Breast cancer | Preclinical | Inhibited the CAF
phenotype | (75) |
| Ting, 2016 | Silibinin | TGF-β2 | Prostate
cancer | Preclinical | Blocked CAF
activation | (76) |
| Ting, 2015 | Silibinin | TGF-β2 | Prostate
cancer | Preclinical | Prevented prostate
cancer cell-mediated differentiation of naive fibroblasts into
CAFs | (77) |
| Jin, 2023 | Baicalein-loaded
mPEG-PLGA nanoparticles | TGF-β | Breast cancer | Preclinical | Blocked CAF
activation | (78) |
| Pang, 2024 | The TGF-β receptor
I inhibitor SB525334 (SB) and docetaxel micelles | TGF-β | Pancreatic
cancer | Preclinical | Suppresses the
activity of myofibroblastic CAFs | (79) |
Eribulin is a well-tolerated anti-microtubule drug
used to treat various tumors. It has been found to 'normalize' CAF
function in pancreatic CAFs by a mechanism similar to blocking the
TGF-β-induced pathway, suggesting that Eribulin could be used as a
CAFs/stromal standardizing drug (56). Preclinical studies reported a
self-assembled nanoplatform of hyaluronic acid (HA)-paclitaxel
(PTX) (HA-PTX) prodrugs and Marimastat (MATT)-loaded
thermosensitive liposomes (LTSLs; MATT-LTSLs) for the treatment of
metastatic cancers by downregulating TGF-β expression and blocking
CAF activation (57).
Similarly, the TGF-βR1 inhibitor Galunisertib
(LY2157299) monohydrate inhibited tumor growth in ovarian cancer by
blocking fibroblast activation through downregulation of TGF-β
expression (58). Of note, TGF-β1
induced the formation of CAF phenotypes in starved NIH3T3
fibroblasts and xenografted Balb/c mice, thereby promoting breast
cancer tumor growth. By contrast, blockade of TGF-β1 induced these
effects following the administration of the TGF-β type I receptor
kinase inhibitor LY-2157299 (59).
Sulfosuccinimidyl oleate sodium, a CD3 inhibitor,
was found to have a significant negative effect on the
proliferative and migratory capacity of CAFs and further
mechanistic studies were found to be associated with a reduction in
the levels of related activation genes (α-SMA, FAP and waveform
protein) and cytokines (IL-6, TGF-β and VEGF-α) in nude mice
orthotopically implanted with CAFs and HCC cells (60).
The combination of non-steroidal anti-inflammatory
drugs (aspirin and ibuprofen) with vincristine effectively
inhibited the secretion of TGF-β and IL-6 from CAFs in CRC, thereby
reducing chemoresistance to vincristine (61).
Metformin (Met), a first-line drug for diabetes
treatment, can disrupt tumor-stromal crosstalk by blocking TGF-β
signaling in breast cancer cells (62). Additionally, the cardiac glycoside
analog digoxin is a particularly potent CAF blocker, capable of
inhibiting TGF-β-induced fibronectin expression at low nanomolar
concentrations without causing cytotoxicity (63).
Pirfenidone, an antifibrotic agent and TGF-β
antagonist, synergizes with adriamycin to inhibit triple-negative
breast cancer by inducing apoptosis in CAFs cells and suppressing
CAF proliferation through inhibiting TGF-β expression (64). Furthermore, pirfenidone induces
apoptosis in lung CAF cells by inhibiting TGF-β1 (65). Another TGF-β receptor inhibitor,
LY2109761, was found to disrupt the crosstalk between cancer cells
and CAFs by inhibiting TGF-β expression, leading to a significant
reduction in HCC growth and dissemination (66).
Meanwhile, Halofuginone, an old antiparasitic drug
for poultry, has been found to inhibit the TGF-β/Smad2/3 signaling
pathway, thereby attenuating the promotional effect of CAFs on the
migration and invasion of oral squamous cell carcinoma (OSCC) cells
(67). Similarly, Nintedanib, a
clinically approved multikinase receptor inhibitor, has been found
to inhibit TGF-β1-induced expression of a panel of pro-fibrotic
activation markers in adenocarcinoma CAFs (68). Additionally, some investigators
have used lentiviral vectors encoding BMPs and activin
membrane-bound inhibitor to inhibit TGF-β/Smad signaling in human
BM-MSCs, thereby preventing them from differentiating into CAFs
induced by the TME and the ensuing effects on preneoplastic lesions
(69).
Active ingredients from natural products can also
target CAFs by modulating the TGF-β signaling pathway. For example,
artemisinin and its derivatives have been found to inhibit TGF-β
signaling, thereby inactivating cancer-associated fibroblasts and
inhibiting cancer metastasis (70). Additionally, tetrahydrocannabinol
and cannabidiol, either alone or in combination, inhibit
TGF-β-induced interactions between CAFs and cancer cells (71). Furthermore, chestnut extract
combined with TGF-β1 has shown synergistic effects on the presence
of polymerized α-SMA stress fibers, particularly in CAFs (72).
In CRC, the natural herbal compounds curcumin and
chelerythrine chloride have been found to modulate the
TGF-β2/Smad2/3 axis, effectively inhibiting cancer cell invasion
and migration by intervening in CAF activity (73,74). In breast cancer, curcumin
effectively inhibited the CAF phenotype by a mechanism associated
with a significant reduction in prostaglandin E2 and TGF-β
production (75).
Silibinin, a flavonoid isolated from the fruit of
the chrysanthemum plant Silybum marianum, has recently been
shown to target CAF-mediated prostate cancer aggressiveness by
inhibiting the expression of the TGF-β2-related pathway (76,77).
Despite the considerable therapeutic potential of
these natural compounds, their low bioavailability remains a
significant limitation. Consequently, the development of
nanomedicines to enhance bioavailability has garnered increasing
attention. Furthermore, fibroblasts within the TME undergo
phenotypic and functional changes, typically driven by interactions
with tumor cells; these transformed cells are known as CAFs.
Therefore, inhibiting the activation of CAFs can effectively
regulate tumor progression at its early stages. For example,
mPEG-poly(lactic-co-glycolic acid) (PLGA) nanoparticles loaded with
baicalein have been developed, showing high drug loading capacity,
stability, biocompatibility and low toxicity. In breast cancer
models, this formulation inhibits TGF-β/Smad and TGF-β/MAPK
signaling pathways, reduces CAF activation and ultimately enhances
chemotherapy efficacy (78).
In pancreatic cancer treatment, a two-stage combined
therapy has been introduced, comprising the TGF-β receptor I
inhibitor SB525334 and docetaxel micelles. This approach decreases
TGF-β secretion and suppresses the activity of myofibroblastic
(my)CAFs, identified by α-SMA and FAPα expression, thereby
improving therapeutic outcomes (79).
Overall, research on modulating CAFs via inhibition
of the TGF-β signaling pathway has not only focused on suppressing
the pro-tumorigenic functions and activity of CAFs but also on
disrupting tumor-stroma interactions, as exemplified by agents such
as Met and pirfenidone. Within the TME, fibroblasts undergo
phenotypic and functional transitions typically driven by
interactions with tumor cells, resulting in the emergence of CAFs.
Notably, targeting the TGF-β pathway can reprogram CAFs, as
demonstrated by curcumin, and prevent the differentiation of naïve
fibroblasts into CAF phenotypes using compounds such as
Nano-baicalein, LY364947 and silibinin, thereby inhibiting tumor
initiation at early stages.
Although strategies targeting CAF markers such as
FAP have been explored, they do not directly intervene in the key
signaling pathways, such as TGF-β, that have been extensively
summarized, leaving the precise characteristics of TGF-β-targeted
CAF modulation insufficiently understood. Due to the specificity of
stromal-targeted therapies, efficacy evaluations have predominantly
relied on preclinical models.
In summary, among the various approaches targeting
the TGF-β pathway, those involving small molecule natural compounds
to reprogram CAF function or prevent fibroblast-to-CAF
differentiation appear most mature and selective toward CAFs,
offering the advantage of functional stromal remodeling rather than
mere depletion. These strategies demonstrate promising prospects
for clinical translation, although further studies are needed to
optimize specificity and confirm efficacy in human settings.
NF-κB signaling pathway inhibitors
The NF-κB signaling pathway consists of receptor and
receptor proximal signaling interface proteins, the inhibitor of
NF-κB (IκB) kinase complex, IκB protein and NF-κB dimer. When cells
are subjected to various intracellular and extracellular stimuli,
IκB kinase is activated, leading to the phosphorylation and
ubiquitination of the IκB protein. This process results in the
degradation of the IκB protein and the release of the NF-κB dimer,
which is further activated by various post-translational
modifications (80).
The classical NF-κB signaling pathway is closely
associated with various aspects of cancer biology, including CAF
progression, secretory phenotype and immunosuppressive functions.
Activation of the NF-κB signaling pathway promotes tumor growth,
migration and invasion. Evidence suggests that NF-κB signaling
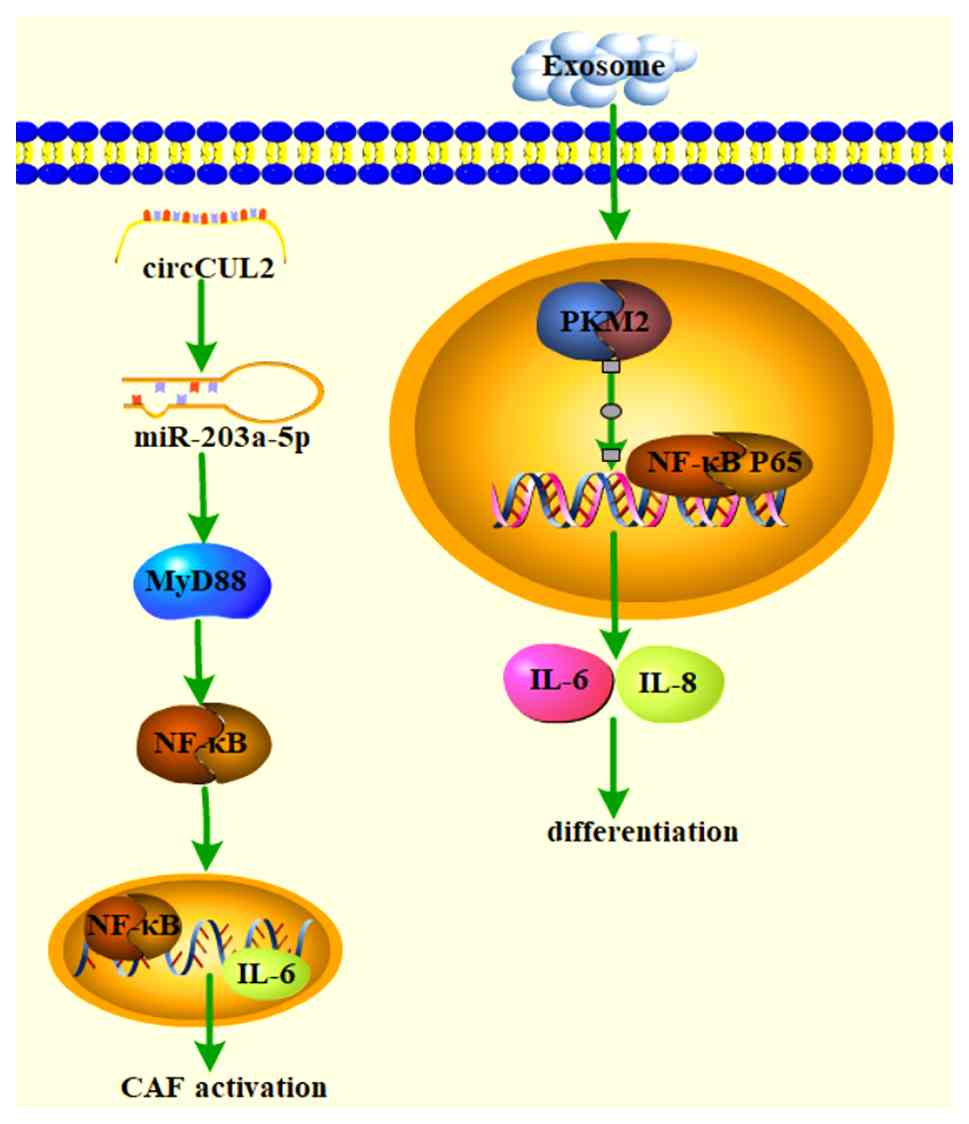
activation facilitates CAF activation (Fig. 4). For instance, circular RNA
circCUL2 (hsa_circ_0000234) is specifically expressed in CAFs and
in pancreatic ductal adenocarcinoma; circCUL2 induces an
inflammatory CAF phenotype through the activation of the MYD88
innate immune signal transduction adaptor (MyD88)-dependent NF-κB
signaling pathway, establishing a distinct fibroblast ecological
niche for cancer progression (81).
Exosomes from gastric cancer have been found to
promote the differentiation of MSCs into CAFs by acetylating the
NF-κB P65 signaling pathway, thereby inducing aberrant metabolic
and inflammatory activation (82). CD146, a cell membrane protein, has
been associated with various human cancers. A study found that
CD146 knockdown promoted CAF activation by potentially inducing the
production of pro-tumorigenic factors through the regulation of
NF-κB activity (83). Eukaryotic
translation initiation factor 4A3 (EIF4A3), a core component of the
exon junction complex, has a crucial role in pre-mRNA splicing.
Evidence suggests that EIF4A3 stabilizes the expression of long
non-coding RNA AGAP2-AS1, activates CAFs through MyD88/NF-κB
signaling and thus exacerbates lung cancer, revealing a new
regulatory axis in lung cancer (84).
In summary, activation of the NF-κB signaling
pathway facilitates the activation or transformation of CAFs and
subsequently promotes tumor progression. Conversely, inhibition of
the NF-κB pathway can eliminate CAFs and increase drug sensitivity.
Genes in the NF-κB signaling pathway may represent potential cancer
therapeutic targets (Table IV)
(85-93).
| Table IVSmall-molecule compounds inhibiting
CAF-induced cancer through suppressing NF-κB signaling
pathways. |
Table IV
Small-molecule compounds inhibiting
CAF-induced cancer through suppressing NF-κB signaling
pathways.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| De Sanctis,
2023 | Chloroquine and
hydroxychloroquine | NF-κB | Lung cancer | Preclinical | Improved tumor
microenvironment | (85) |
| Liu, 2021 | Melatonin | NF-κB | Gastric cancer | Preclinical | Improved
CAF-induced tumor microenvironment | (86) |
| Lou, 2024 | Repertaxin | NF-κB | Gastric cancer | Preclinical | Improved
CAF-induced tumor microenvironment | (87) |
| Jin, 2023 | Bepotastine | NF-κB | Ovarian cancer | Preclinical | Improved
CAF-induced tumor microenvironment | (88) |
| Li, 2022 | Jianpi Jiedu
Recipe | NF-κB | Colorectal
cancer | Preclinical | Blocked CAF
activation | (89) |
| Xu, 2019 | Lentinus
edodes polysaccharides | NF-κB | Prostate
cancer | Preclinical | Impaired
immunosuppressive function of CAFs | (90) |
| Ma, 2019 | Ligustilide | NF-κB | Others | Preclinical | Blocked CAF
activation | (91) |
| Chen, 2024 | Bruceine D | NF-κB | TNBC | Preclinical | Inhibited
CAF-promoted TNBC transfer | (92) |
| Pan, 2024 | Scutellaria
barbata D. Don and Scleromitrion diffusum (Willd.)
R.J.Wang | NF-κB | TNBC | Preclinical | Blocked CAF
activation | (93) |
NF-κB inhibitors can be used alone or in combination
with chemotherapeutic agents to treat cancer and prevent
recurrence. For example, chloroquine (CQ) and hydroxychloroquine
(HCQ) are known antimalarials that have been successful in treating
autoimmune and oncologic diseases. CQ and HCQ normalize the
tumor-associated vascular system and stimulate cancer-associated
fibroblasts through mechanisms related to the regulation of the
Toll-like receptor 9 (TLR9)/NF-κB pathway (85). Melatonin has been reported to
indirectly reduce the proliferation and invasion of gastric cancer
cells by affecting cancer-associated fibroblasts, with the
mechanism involving the regulation of the NF-κB pathway (86). Although REPERTAXIN is an IL-8
receptor (CXCR1/2) inhibitor, it effectively attenuates the
protection of CAFs against CD8+ T cytotoxicity-resistant cancer
cells by a mechanism that may be related to reduced levels of
phosphorylated (p)-P38, p-JNK and p-NF-κB (87). In addition, bepotastine, an
approved H1 antihistamine, inhibited the senescence-associated
secretory phenotype (SASP) of CAFs induced by poly (ADP-ribose)
polymerase inhibitors (PARPis) at clinical serum concentrations. It
was further demonstrated that bepotastine attenuated PARPi
resistance in fibroblast-promoted tumors in three-dimensional
organotypic cultures and homologous recombination-deficient
patient-derived xenograft models. Mechanistically, bepotastine
inhibits PARPi-triggered SASP by inhibiting NF-κB signaling
independent of histamine H1 receptors (88).
Additionally, Jianpi Jiedu Recipe (JPJDR), a classic
traditional Chinese medicine prescription, can prevent CRC liver
metastasis by blocking CAF activation through the regulation of the
integrin β-like 1 (ITGBL1)-TNF-α-induced protein 3 (TNFAIP3)-NF-κB
signaling pathway, providing experimental evidence for its clinical
application in preventing and treating CRC metastasis (89). Furthermore, the aforementioned
polysaccharide MPSSS extracted from shiitake mushrooms can alter
the function of prostate CAFs by activating the TLR4-NF-κB pathway,
providing a new strategy for comprehensive tumor treatment
(90). Similarly, ligustilide can
modulate the immunosuppressive function of CAFs by affecting the
TLR4-NF-κB pathway, thereby restoring T-cell proliferation
previously inhibited by CAF supernatant (91).
CAFs have been suggested to be an important factor
in inducing early metastasis and postoperative recurrence tendency
in triple-negative breast cancer (TNBC) due to its characteristics
of inducing dysfunction and promoting tumor metastasis. Chen et
al (92) found that Bruceine
D, an active compound derived from the Chinese herb Brucea
javanica, inhibited CAFs-promoted TNBC metastasis by
suppressing Notch1-Jagged1/NF-κB (p65) signaling. In addition, the
herb is commonly used in clinical practice against Scutellaria
barbata D. Don and Scleromitrion diffusum (Willd.) R.J.
Wang for its anti-tumor properties. Using a combination of
bioinformatics and in vitro and in vivo experiments,
researchers have found that the herb inhibits the progression of
TNBC through inhibition of NF-κB activation triggered by
CAFs-derived IL-6 in S. barbata and S. diffusum
(93).
Overall, research exploring the regulation of CAFs
through inhibition of the NF-κB signaling pathway has extended
beyond merely suppressing the pro-tumorigenic activities and
activation of CAFs, as seen with agents such as JPJDR and
Ligustilide, to also enhancing chemosensitivity with compounds such
as Bepotastine. Additionally, CAF-induced immunosuppression remains
a major barrier to effective cancer therapy and studies have
demonstrated that targeting the NF-κB pathway with drugs such as
MPSSS and Repertaxin can effectively mitigate immune resistance,
thus opening new avenues to potentiate immunotherapeutic outcomes.
Although therapies directed at CAF surface markers have been
explored, they frequently bypass critical intracellular signaling
cascades, including NF-κB, that govern CAF behavior, leaving the
precise characteristics of NF-κB-targeted CAF interventions
insufficiently defined. Furthermore, due to the complexity and
context-dependent nature of stromal-targeted treatments, current
evaluations of therapeutic efficacy primarily rely on preclinical
models.
In summary, the most advanced and selective
approaches targeting the NF-κB pathway in CAFs combine inhibition
of CAF activation with the reversal of immunosuppressive functions,
favoring stromal reprogramming over outright depletion. These
strategies hold substantial promise for clinical translation,
contingent upon further refinement to ensure specificity and
durable therapeutic effects in patients.
FAP signaling inhibitors
FAP, a membrane-bound glycoprotein, is upregulated
in CAFs but not in normal fibroblasts (94). FAP is positive in >90% of human
cancers, making it a promising universal target for solid tumors
(95). A phase I clinical study
found that sibrotuzumab can inhibit tumor progression by
specifically targeting and inhibiting CAFs in patients with
FAP-positive cancers (96).
Another clinical trial with iodine 131-labeled monoclonal antibody
F19 (131I-mAbF19) demonstrated inhibition of tumor metastasis by
recognizing FAP-specific depletion of CAFs (97).
Activated CAFs and their secreted collagen
contribute to a dense fibrotic stroma that impedes drug penetration
and characterizes pancreatic ductal adenocarcinoma (PDAC) as an
immune-desert tumor. In a clinical study on PDAC, combination
treatment with paricalcitol and hydroxychloroquine alongside
gemcitabine reduced the number of fibroblasts expressing Ki67, FAP
and SMA, while concurrently decreasing autophagy-related
transcripts (98), indicating
that this regimen enhances chemosensitivity and improves
therapeutic outcomes in PDAC. Similarly, preclinical studies have
demonstrated that in vivo administration of FAP-targeted
chimeric antigen receptor macrophages (FAP-CAR-M), engineered to
eliminate activated CAFs marked by FAP, markedly reduced markers of
activated CAFs (FAP), collagen volume fraction and Col1a1 secretion
in murine models. These findings suggest that FAP-CAR-M represent a
potential therapeutic strategy to overcome the fibrotic barrier and
thereby potentiate chemotherapy and immunotherapy against PDAC
(99).
Additionally, in mouse models of epithelial-derived
solid tumors, preclinical studies have found that PT630 can target
FAP to reduce the content of CAFs and blood vessel density in
tumors, thereby inhibiting stromatogenesis, angiogenesis and ECM
remodeling, ultimately suppressing tumor growth (100). AMD3100, a CXCL12 receptor
chemokine (C-X-C motif) receptor 4 inhibitor removes FAP-expressing
CAFs to immunocontrol tumor growth in PDA-bearing mice, revealing
the anti-tumor effects of immunotherapeutic antibodies (101). Natural active ingredients have
great potential in targeting FAP and thus inhibiting CAFs.
Oxidized omphalosine (Om) is an alkaloid compound
and one of the active components of bitter ginseng in Chinese
medicine. Om effectively inhibits the activation of CAFs and
promotes T-cell infiltration across CAFs by downregulating the
expression of FAP and α-SMA (102). This intervention may lead to
promising therapeutic directions for the treatment of TNBC.
Trigonelline (TGN), an alkaloid found in medicinal
plants such as Coffea spp. and Trigonella
foenum-graecum (fenugreek), has exhibited notable anticancer
properties across various malignancies. A recent study revealed
that TGN reduces fibroblast-to-CAFs conversion by downregulating
α-SMA and FAP expression in bladder cancer, indicating its ability
to normalize the TME. These results suggest that TGN has potential
to alleviate chemoresistance and thereby enhance chemotherapeutic
efficacy (103).
Similarly, rosmarinic acid (RA) treatment markedly
attenuated CAF activation markers and reversed EMT-related
alterations in cancer cells, resulting in reduced tumor growth in
CAF-enriched xenograft models. The combination of RA with gefitinib
exhibited enhanced antitumor effects compared with gefitinib
monotherapy (104). Echinocandin
A (Ech A), a marine substance isolated from sea urchins, is a
strong antioxidant. A study has reported that Ech A attenuates
CAFs-induced lung cancer cell migration, which is accompanied by a
decrease in the expression levels of CAF markers, waveform proteins
and FAP (105).
In addition, nanomaterial-based drug delivery
systems are particularly promising for cancer therapy. Leveraging
the self-assembly properties of amphiphilic peptides, researchers
developed a smart transformer-like drug delivery system based on
cleavable amphiphilic peptides that specifically respond to FAP-α
expressed on CAFs in the TME, achieving enhanced drug delivery and
promising antitumor effects (106). In addition, a CAF-associated
ITGB1-inactivating peptide-rich membrane nanodelivery system
(designated PMNP-D) has been shown to target both CAFs and tumor
cells to enhance chemotherapy by promoting drug infusion. After
prolonged blood circulation and active targeting of the tumor site,
PMNP-D triggers the release of FNIII14 in response to CAFs
overexpressing FAP-α, which binds to ITGB1 and inhibits the
biological function of CAFs to produce stromal matrix, thereby
loosening the condensed stromal structure and enhancing the
permeability of nanotherapeutic agents in the tumor (107). In breast cancer treatment, a
novel polymeric nanoparticle system targeting CAFs has been
developed by encapsulating synthetic 8-O-methylfusarubin (OMF)
within nanoparticles modified with anti-FAP antibodies
(OMF@NPs-anti-FAP). This formulation showed markedly enhanced
cytotoxicity in 3D spheroid models. Compared with
low-FAP-expressing cells such as MCF-10A, HDFa and PC-B-142 CAFs,
the high-FAP-expressing PC-B-132 CAF cells exhibited markedly
higher levels of cell death, as confirmed by flow cytometry. These
findings suggest that the developed high-specificity OMF-loaded
polymeric nanoparticle system may serve as a promising nanoplatform
for improved breast cancer therapy (108). Similarly, in breast cancer, a
biomimetic nanodrug system (FAP-C NPs) was engineered by fusing 4T1
cell-derived extracellular vesicles with FAP single-chain antibody
fragments to form a biomimetic shell around calcipotriol-loaded
PLGA nanoparticles. Experimental evidence demonstrated that FAP-C
NPs could revert activated CAFs to a quiescent state, thereby
neutralizing their pro-tumor functions, inhibiting cancer cell
stemness, promoting dendritic cell maturation and alleviating the
immunosuppressive effects of CAFs on lymphocytes. Furthermore, when
combined with radiotherapy, this biomimetic nanosystem inhibited
CAF activation, enhanced radiosensitivity and elicited a robust
antitumor immune response, evidenced by a twofold increase in
cytotoxic T-cell infiltration in the TME, ultimately suppressing
tumor growth effectively. These results highlight the great
potential of FAP-C NPs as a targeted therapeutic strategy in breast
cancer treatment (109).
Meanwhile, phototherapeutic techniques have been
used to target CAFs in tumors. Investigators developed a novel
near-infrared photoimmunotherapy technique focusing on FAP, which
specifically induced rapid cell death in vitro and in
vivo mouse xenograft models with no adverse effects (110). Notably, the combination of
photodynamic therapy and nanoparticles could substantially improve
the ability to eradicate CAFs and induce broad-spectrum anticancer
effects. Researchers used ferritin, a compact protein cage of
nanoparticles, as a photosensitizing carrier and coupled
FAP-specific single-chain variable fragments to the surface of
ferritin to create a nanoparticle-based photoimmunotherapy
(nano-PIT). This method effectively eliminated CAFs from tumors by
light irradiation enabled by nano-PIT while causing minimal damage
to healthy tissues due to the localized nature of the treatment
(111).
Other researchers developed a photodynamic therapy
method called αFAP-Z@FRT, which is based on ZnF16Pc (a
photosensitizer) loading and FAP-specific single-chain
variable-fragment conjugated apoferritin nanoparticles. This method
is also effective in eradicating CAFs from tumors without causing
systemic toxicity (112). The
aforementioned results imply that the FAP-targeting
polymersomes-based delivery system has the potential to be an
excellent tool for cancer therapy.
FAP represents the most extensively studied and
clinically advanced target among the various strategies aimed at
modulating CAFs. Therapeutic agents targeting FAP not only inhibit
CAF activation, thereby attenuating tumor progression and the
fibroblast-to-CAF transition, as exemplified by compounds such as
RA and Echinocandin A, but also serve as direct targets for drug
delivery systems that selectively suppress CAFs. Noteworthy
examples include transformable peptide nanocarriers, FNIII14
peptide-enriched membrane nanocarriers and polymeric nanoparticles
loaded with 8-O-methylfusarubin, all of which have demonstrated
promising efficacy across diverse cancer models. Among the distinct
molecular pathways implicated in CAF regulation, approaches
centered on FAP inhibition stand out for their maturity,
specificity and capacity to reprogram the tumor stroma rather than
solely deplete fibroblast populations. Consequently, FAP-targeted
interventions exhibit superior selectivity towards CAF populations
and hold significant translational potential, underscoring their
leading role in the clinical advancement of matrix-focused cancer
therapies (Table V) (96-109,112).
| Table VSmall-molecule compounds inhibiting
CAF-induced cancer through suppressing FAP signaling. |
Table V
Small-molecule compounds inhibiting
CAF-induced cancer through suppressing FAP signaling.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| Scott, 2003 | Sibrotuzumab | FAP | NSCLC | Phase I | Depleted CAFs to
suppress tumor progression | (96) |
| Welt, 1994 | Iodine 131-labeled
monoclonal antibody F19 | FAP | Colorectal
carcinomas | Phase I | Inhibited the
function of CAFs to suppress cancer progression | (97) |
| Nagaraju, 2025 | Paricalcitol and
hydroxychloroquine alongside gemcitabine | FAP | Pancreatic ductal
adenocarcinoma | Phase I | Decreases the
number of Ki67, FAP and α-smooth muscle actin-expressing CAFs | (98) |
| Wang, 2025 | FAP-targeted
chimeric antigen receptor macrophages | FAP | Pancreatic
cancer | Preclinical | Reduced markers of
activated CAFs | (99) |
| Santos, 2009 | PT630 | FAP | Lung cancer | Preclinical | Reduced CAF levels
in the tumor | (100) |
| Feig, 2013 | αFAP-PE38 | FAP | Breast cancer | Preclinical | Inhibited the
function of CAFs to suppress cancer progression | (101) |
| Wang, 2023 | Oxymatrine | FAP | Triple negative
breast cancer | Preclinical | Blocked CAF
activation | (102) |
| Kao, 2025 | Trigonelline | FAP | Bladder cancer | Preclinical | Reduced CAF levels
in the tumor | (103) |
| Li, 2025 | Rosmarinic
acid | FAP | NSCLC | Preclinical | Attenuated CAF
activation | (104) |
| Eum, 2024 | Echinochrome A | FAP | Lung cancer | Preclinical | Inhibited
CAF-induced mediated lung cancer cell migration | (105) |
| Ji, 2016 |
Drug-loadedCAP-NPs | FAP | Prostate
cancer | Preclinical | Disrupted the
matrix barrier of the drug | (106) |
| Liu, 2023 | FNIII14
peptide-enriched membrane nanocarrier | FAP | Adenoid cystic
carcinoma | Preclinical | Inhibited CAF
produced substrate | (107) |
| Rodponthukwaji,
2024 |
OMF@NPs-anti-FAP | FAP | Breast cancer | Preclinical | Promoted
CAF-induced apoptosis | (108) |
| Gao, 2025 | FAP-C NPs | FAP | Breast cancer | Preclinical | Inhibited the
activation of CAFs | (109) |
| Zhou, 2021 | αFAP-Z@FRT | FAP | Lung cancer | Preclinical | Stimulated
immunization against CAF | (112) |
Other signaling pathways or target
regulators
Beyond the aforementioned pathways, additional axes,
including TLR4, hypoxia-inducible factor 1α (HIF-1α), Wnt, G
protein-coupled estrogen receptor (GPER) and PDGFR, also shape the
bidirectional crosstalk between CAFs and cancer cells, with
context-specific features that create complementary intervention
points (Table VI) (62,113-123). Collectively, these pathways
regulate immune suppression, (lymph)angiogenesis, metabolic
reprogramming and hormone signaling, thereby influencing tumor
progression.
| Table VISmall-molecule compounds inhibiting
CAF induced cancer through suppressing various signaling
pathways. |
Table VI
Small-molecule compounds inhibiting
CAF induced cancer through suppressing various signaling
pathways.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| Mei, 2020 | Cinnamaldehyde | TLR4 | Prostate
cancer | Preclinical | Inhibited the
inhibitory effect of CAF T cells | (113) |
| Wang, 2021 | Polysaccharides
from Lentinus edodes | TLR4 | Colorectal
cancer | Preclinical | Decreased CAF
secretion VEGF-C | (114) |
| Ma, 2022 | Ligustilide | TLR4 | Prostate
cancer | Preclinical | Downregulated VEGFA
secretion in prostate CAF | (115) |
| Lappano, 2015 | Calixpyrroles | GPER | Breast cancer | Preclinical | Inhibited the
proliferation of CAF | (117) |
| Maggiolini,
2015 | Benzo(b)pyrrolo
(1,2-d)(1,4)oxazin-4-one structure | GPER | Breast cancer | Preclinical | Inhibited the
stimulatory effect of CAF | (118) |
| Shao, 2020 | Metformin | HIF-1α | Breast cancer | Preclinical | Prevented
tumor-stromal crosstalk in breast cancer | (62) |
| Du, 2015 | Curcumin | HIF-1α | Prostate
cancer | Preclinical | Suppressed
CAF-driven prostate cancer invasion | (120) |
| Kinoshita,
2010 | Imatinib | PDGF | Lung cancer | Preclinical | Inhibited CAF
proliferation | (122) |
| Pietras, 2008 | Imatinib | PDGF | Cervical
carcinoma | Preclinical | Inhibited the
matrix-supportive function of CAFs | (123) |
TLR4-centered signaling exemplifies the immune- and
angiogenesis-related dimensions of CAFs function. Cinnamaldehyde
alleviates CAFs-mediated T-cell suppression in a TLR4-dependent
manner, functionally reprogramming CAFs and offering therapeutic
potential (113). In CRC, the
polysaccharide MPSSS reduces lymphangiogenesis by decreasing
CAFs-derived VEGF-C via the TLR4/JNK pathway, suggesting a strategy
for comprehensive treatment (114). Similarly, ligustilide
downregulates VEGFA levels in CAFs through a TLR4-related pathway,
thereby blocking CAFs-driven angiogenesis and highlighting the
anti-angiogenic promise of natural active molecules (115).
GPR30 (also known as GPER), a member of the G
protein-coupled receptor family, mediates estrogen signaling in
various types of normal and malignant cells, leading to specific
gene markers, as well as the migration and proliferation of cancer
cells and CAFs (116).
Investigators synthesized cuprolyl derivatives to act as GPER
antagonists, which effectively inhibited CAFs from stimulating
cancer progression in the TME (117). Additionally, two novel selective
GPER antagonists based on the structure of benzo(b)pyrrolo(1,2-d)
(1,4)oxazin-4-one were synthesized to
inhibit the stimulatory effect of CAFs on cancer progression in
breast cancer (118).
HIF-1α is closely related to CAFs migration and
invasion. In a nude mouse subcutaneous xenograft model,
upregulation of HIF-1α expression promotes CAF migration and
invasion via miR-210, which leads to CRC metastasis and increases
patient mortality (119).
Therefore, inhibition of HIF-1α expression is beneficial in
inhibiting CAF migration and invasion, thus suppressing cancer
progression. HIF-1α signaling in CAFs promotes tumor progression
mainly by regulating glycolytic metabolism. Met, a diabetes drug,
was found to inhibit HIF-1α signaling in CAFs, leading to reduced
breast cancer cell invasion (62). Furthermore, curcumin, an active
ingredient of Chinese medicine, eliminated CAF-induced invasion and
EMT through a mechanism related to its inhibition of monoamine
oxidase A/mTOR/HIF-1α signaling in prostate cancer (120).
PDGF, a disulfide-linked dimer, is a potent mitogen
in various cancer cell types, including glioma, sarcoma, pancreatic
and prostate cancers. A study has shown that PDGF-associated
receptors are expressed by cancer-associated stromal cells,
pericytes and CAFs in human colon cancer (121). Imatinib, a small-molecule
tyrosine kinase inhibitor, has shown promising antitumor activity.
Mechanistic studies revealed that its antitumor activity is
mediated by blocking PDGF signaling, thereby reducing the
proliferation-stimulating effect of CAFs on lung cancer cells
(122). Similarly, in a mouse
model of human cervical cancer, imatinib inhibited the stromal PDGF
receptor, leading to reduced proliferation and angiogenesis of
cervical lesions by inhibiting CAFs-expressed angiogenic factors,
such as fibroblast growth factor 2 (FGF-2) and epithelial cell
growth factor FGF-7 (123).
Selective inducers of cell death
Cell death encompasses multiple, mechanistically
distinct modalities, including apoptosis, necroptosis, ferroptosis,
pyroptosis and necrosis, each defined by characteristic
morphological and biochemical signatures, while autophagy
represents a stress-adaptive catabolic process that can intersect
with and, in certain contexts, contribute to cell death. During
tumor evolution and therapy, cancer cells can undergo diverse forms
of regulated cell death, notably apoptosis, autophagy-associated
cell death and ferroptosis. Beyond classical apoptosis-inducing
agents traditionally used to eliminate tumor cells and, in certain
strategies, CAFs, emerging approaches that suppress CAFs by
triggering autophagy or ferroptosis have shown preclinical efficacy
in curbing recurrence and metastasis. Representative interventions,
targets and outcomes are summarized in Table VII (65,129-149), underscoring the therapeutic
potential of modulating multiple death pathways within the TME.
| Table VIISmall-molecule compounds inhibiting
CAF-induced cancer through regulating ferropotosis, apoptosis and
autophagy. |
Table VII
Small-molecule compounds inhibiting
CAF-induced cancer through regulating ferropotosis, apoptosis and
autophagy.
| First author/s,
year | Name | Target | Type of cancer | Study type | Anticancer
mechanisms | (Refs.) |
|---|
| Li, 2023 | FER-1 | Ferroptosis | Gastric cancer | Preclinical | Inhibited
CAF-derived DACT3-AS1 | (129) |
| Li, 2023 | RSL-3 | Ferroptosis | Oral squamous cell
carcinoma | Preclinical | Inhibited CAFs with
podoplanin positive expression | (129) |
| Hu, 2021 | eNVs-FAP | Ferroptosis | Colon, melanoma,
lung and breast cancer | Preclinical | Inactivated
CAFs | (130) |
| Yao,2023 | Combination of
deferrioxamine and FSTL1 neutralizing antibody | Ferroptosis | Gastric cancer | Preclinical | Inactivated
CAFs | (131) |
| Li, 2020
ferroptosis | Disulfiram/copper
Nasopharyngeal cancer |
Apoptosis/Preclinical | Inactivated
CAFs | | | (135) |
| Lee, 2018 | Bortezomib and
panobinostat | Apoptosis | Breast cancer | Preclinical | Reduced the
viability of CAFs | (136) |
| Mediavilla-Varela,
2016 | Pirfenidone | Apoptosis | Lung cancer | Preclinical | Induced lung CAF
apoptosis | (65) |
| Zeng, 2020 | Curcumin | Apoptosis | Others | Preclinical | Promotion of CAF
apoptosis by ROS-mediated endoplasmic reticulum stress | (137) |
| Han, 2020 | Cinnamaldehyde | Apoptosis | Prostate
cancer | Preclinical | Inhibited CAF
proliferation and promoted CAF apoptosis | (138) |
| Chang, 2018 | Nab-paclitaxel | Apoptosis |
Cholangiocarcinoma | Preclinical | Destroyed CAF | (139) |
| Donthireddy,
2022 | ONP-302
nanoparticles | Apoptosis | Others | Preclinical | Induced CAF
apoptosis | (140) |
| Chen, 2016 | Nanoliposome with
navitoclax | Apoptosis | Liver cancer | Preclinical | Specifically
eradicated CAF | (141) |
| Shen, 2023 | Nanoemulsions (DOX
and siRNA) | Apoptosis | Others | Preclinical | Inhibited CAF
proliferation and promoted CAF apoptosis | (142) |
| Zhao, 2019 | Chloroquine | Autophagy | Liver cancer | Preclinical | Attenuated the
stemness enhanced by CAF | (146) |
|
Martinez-Outschoorn, 2010 | Chloroquine | Autophagy | Breast cancer | Preclinical | Reduced CAF
generation | (147) |
| Han, 2016 | Polysaccharides
from Polygonatum | Autophagy | Prostate
cancer | Preclinical | Inhibited CAF
proliferation | (148) |
| Ferraresi,
2017 | Resveratrol | Autophagy | Lung cancer | Preclinical | Offset CAF
maturity | (149) |
Ferroptosis and CAFs
Ferroptosis is a recently characterized,
iron-dependent form of programmed cell death that is
mechanistically distinct from apoptosis, necrosis and autophagy. It
is driven by divalent iron and ester oxygenases that catalyze
peroxidation of polyunsaturated fatty acids within membrane
phospholipids, leading to lethal lipid-reactive oxygen species
(ROS) accumulation. This lipid peroxidation is normally
counteracted by the glutathione system, with glutathione peroxidase
4 (GPX4) as the core enzyme; decreases in GPX4 activity and
glutathione availability sensitize cells to ferroptosis (124). Within the TME, CAFs modulate
therapy response in part by suppressing ferroptosis, thereby
fostering drug resistance. CAFs-secreted exosomal miR-522, for
example, targets arachidonate 15-lipoxygenase to block lipid-ROS
accumulation and inhibit ferroptosis in cancer cells, ultimately
reducing chemotherapy sensitivity (125). Similarly, CAFs-derived exosomal
miRNAs that target acyl-CoA synthetase long chain family member 4
(ACSL4) inhibit ferroptosis and induce gemcitabine resistance in
pancreatic cancer cells (126).
Beyond exosomal signaling, CAFs can reprogram cysteine metabolism
to boost glutathione synthesis and increase resistance to
ferroptosis in pancreatic cancer (127). These findings collectively
support ferroptosis as a promising axis for CAFs-focused
intervention. Although research on ferroptosis activators is broad,
relatively few small molecules have been designed to target CAFs by
mediating ferroptosis; accordingly, this article summarizes
small-molecule approaches to modulate ferroptosis in the context of
CAFs (Fig. 5).
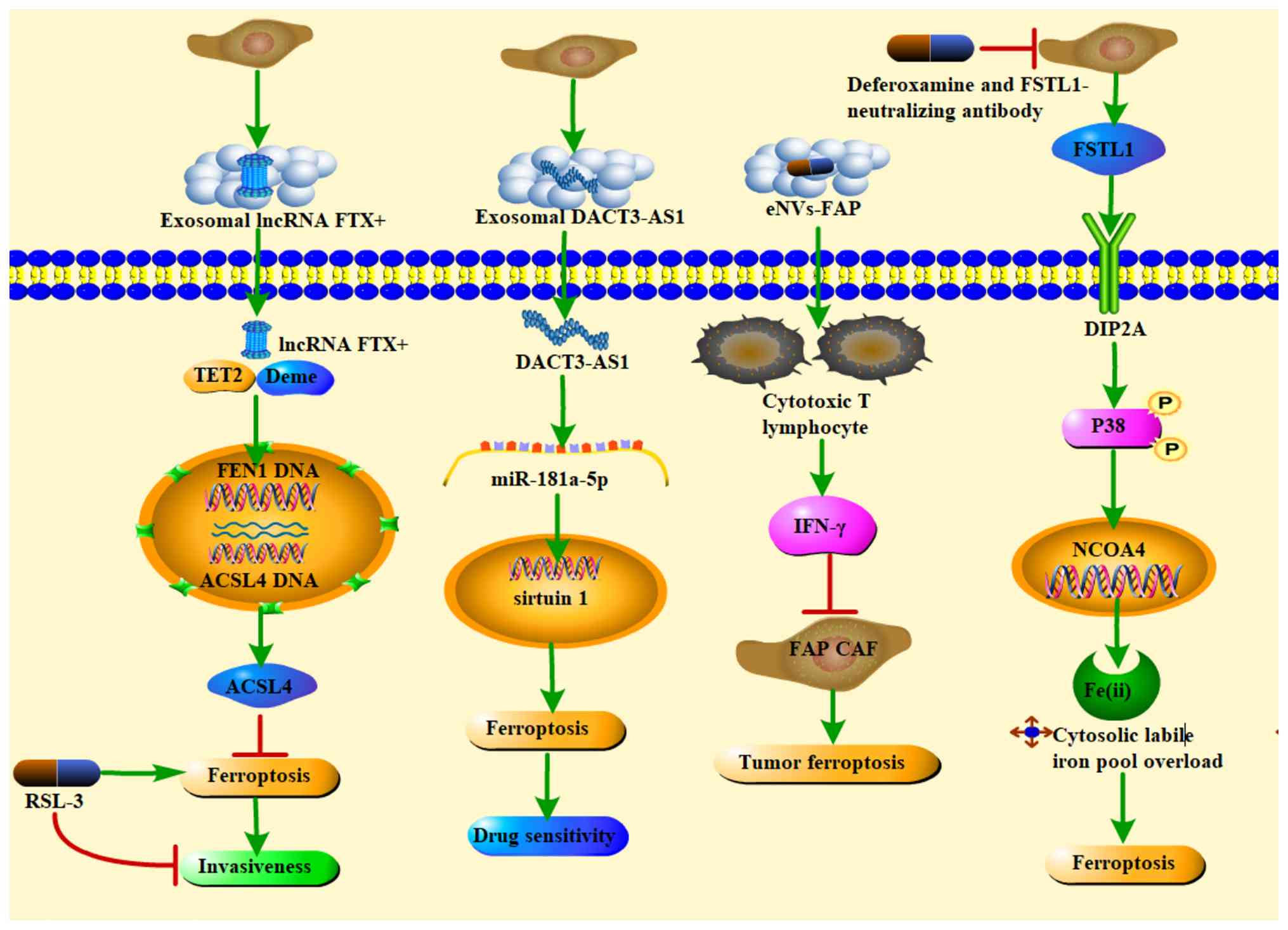 | Figure 5Small molecule compounds mediate
mutual crosstalk between CAFs and ferroptosis to inhibit tumor
progression or enhance chemotherapeutic drug sensitivity. RSL-3
inhibits the tumorigenesis potential of oral squamous cell
carcinoma cells overexpressing FEN1; exosome DACT3-AS1 confers
cancer cell sensitivity to oxaliplatin through miR-181a-5p/sirtuin
1-mediated iron death; cellular immune responses activated by the
NVs-FAP vaccine can promote tumour ferroptosis by releasing IFN-γ
from the CD8+ T lymphocytes (CTLs) and depleting FAP CAFs;
functional targeting of CAFs with a combination of deferoxamine and
FSTL1-neutralising antibody markedly alleviated CAFs-induced iron
death in NK cells and enhanced NK cell cytotoxicity against GC.
CAFs, cancer-associated fibroblasts; miR, microRNA; NVs,
nanovesicles; FAP, fibroblast activation protein; IFN-γ,
interferon-γ; CTLS, CD8+ T lymphocytes; TET2, ten-eleven
translocation 2; Deme, Demeter; FEN1, flap structure-specific
endonuclease 1; ACSL4, long-chain-fatty-acid-CoA ligase 4;
DACT3-AS1, DACT3 antisense RNA 1; IFN, interferon; FSTL1,
follistatin-like 1; DIP2A, disco interacting protein 2 homolog A;
NCOA4, nuclear receptor coactivator 4. |
Importantly, CAF-derived signals are context
dependent and can also sensitize tumors to ferroptosis. CAFs
exosomal disheveled binding antagonist of beta catenin 3 antisense
1 (DACT3-AS1) acts as an inhibitory regulator of malignant
transformation and oxaliplatin resistance by inducing ferroptosis,
characterized by decreased GPX4 and solute carrier family 7 member
11 (SLC7A11) expression, reduced GSH levels and suppression of
oxaliplatin-treated cells; the ferroptosis inhibitor ferrostatin-1
reverses these effects, indicating that DACT3-AS1 engages
ferroptotic mechanisms in oxaliplatin-treated gastric cancer cells
(128). In OSCC, a
podoplanin-positive CAFs subset promotes invasiveness by inhibiting
ferroptosis in tumor cells via the FTX/flap structure-specific
endonuclease 1 (FEN1)/ACSL4 signaling cascade; treatment with the
ferroptosis activator RSL-3 suppresses the tumorigenic potential of
OSCC cells overexpressing FEN1 in vitro and in vivo
(129). Leveraging the
prevalence of FAP overexpression in >90% of human tumors,
exocrine-like nanovesicles (eNVs-FAP) derived from FAP-engineered
tumor cells have been developed as a tumor vaccine that promotes
tumor ferroptosis and inhibits growth by releasing interferon-γ
(IFN-γ) from cytotoxic T lymphocytes while depleting FAP+ CAFs
(130). Ferroptosis also
intersects with antitumor immunity: In gastric cancer, CAFs as a
major immunosuppressive component can induce ferroptosis in natural
killer (NK) cells through iron regulation, impairing their
cytotoxic function and promoting tumor progression and immune
escape. In patient-derived organoid models, functional targeting of
CAFs using the iron chelator deferoxamine combined with a
follistatin like protein 1-neutralizing antibody alleviates
CAFs-induced ferroptosis in NK cells and enhances their
cytotoxicity against gastric cancer (131). Similarly, CAFs-induced
resistance to immunotherapy is closely linked to anoctamin 1
(ANO1)-mediated inhibition of cancer cell ferroptosis; ANO1
upregulates nuclear factor erythroid 2-related factor 2/SLC7A11 via
a PI3K-Akt-dependent pathway, whereas the ferroptosis agonist
erastin reverses ANO1-driven suppression of lipid ROS and
malondialdehyde and mitigates ANO1's malignant effects (132).
Together, these studies delineate a bidirectional
relationship between CAFs and ferroptosis: CAFs can impede
ferroptosis to support drug resistance and immune evasion, yet they
can also be targeted or leveraged to restore ferroptotic
vulnerability and resensitize tumors. Practically, integrating
ferroptosis modulators, whether small molecules such as RSL-3 and
erastin or targeted nanovesicle platforms such as eNVs-FAP, into
CAF-centric strategies offers a route to counteract stromal
protection, enhance chemotherapy efficacy and strengthen antitumor
immunity.
Apoptosis and CAFs
Apoptosis is a genetically programmed, autonomous
and orderly form of cell death that preserves tissue homeostasis by
removing damaged or superfluous cells, thereby enabling improved
adaptation to the microenvironment. Within the TME, CAFs are
pivotal drivers of tumorigenesis, progression and migration;
accordingly, eliminating CAFs or attenuating their tumor-promoting
activity can facilitate tumor immunotherapy (133). It has been reported that CAFs
exhibit tumor-promoting properties by inhibiting apoptosis of tumor
cells; however, evidence also suggests that CAFs may stimulate
apoptosis of tumor cells (134).
These observations position apoptosis as a tractable regulatory
node for modulating CAF activity. Although the available evidence
suggests that CAF apoptosis may be a double-edged sword, wherein
excessive depletion could perturb stromal integrity or provoke
maladaptive remodeling, most current strategies intentionally
promote CAF apoptosis to restrain tumor progression. Pharmacologic
and repurposed agents illustrate this approach. Disulfiram/copper
(DSF/Cu), noted for clinical promise based on its anticancer
activity and safety, induces apoptosis in nasopharyngeal
fibroblasts via an aldehyde dehydrogenase-independent mechanism;
in vivo, DSF/Cu combined with cisplatin (CDDP) is well
tolerated and markedly suppresses nasopharyngeal tumor growth both
in tumor cell models and in 5-8F xenografts (135). Clinically advanced agents such
as bortezomib and panobinostat have likewise emerged as anti-CAF
candidates: They reduce the viability of multiple patient-derived
CAF populations by inducing caspase-3-mediated apoptosis, with
combination therapy outperforming either monotherapy in
vitro and in xenografts of mouse breast cancer cells with mouse
CAFs (136). Pirfenidone, an
antifibrotic drug approved for idiopathic pulmonary fibrosis,
triggers apoptotic death in pulmonary CAFs at high concentrations
and low-dose combinations with cisplatin further increase CAF death
in both co-culture systems and in vivo settings (65). Natural compounds add complementary
mechanisms: Curcumin and cinnamaldehyde, extracted from Chinese
herbs, induce apoptosis and cell-cycle arrest in prostate cancer
CAFs via ROS-mediated endoplasmic reticulum (ER) stress and
intrinsic apoptotic pathways, respectively (137,138).
Nanotechnology has expanded the toolkit for
selectively inducing apoptosis in CAFs while potentially improving
therapeutic indices. Nab-paclitaxel, an effective agent against
intrahepatic cholangiocarcinoma (IH-CCA), more efficiently destroys
CAFs than its prototype paclitaxel in a rat model of
thioacetamide-induced spontaneous desmoplastic IH-CCA (139). Biodegradable, negatively charged
ONP-302 nanoparticles have been used to induce apoptosis in CAFs
within the TME (140). Targeted
delivery systems further enhance specificity: Navitoclax-loaded
nanoliposomes modified with the FH peptide (FH-SSL-Nav), which
binds tenascin C predominantly expressed by CAFs, selectively
engage apoptotic pathways in CAFs (141); similarly, nucleic acid
aptamer-modified PLGA nanoemulsions effectively target CAFs and
induce their apoptosis (142).
Collectively, these data support apoptosis-oriented
modulation of CAFs as a rational strategy to weaken stromal support
for malignancy and to potentiate standard anticancer therapies.
Conceptually, the convergence of repurposed small molecules and
targeted nanocarriers offers a path to enhance selectivity, reduce
off-target toxicity and integrate CAF apoptosis with cytotoxic or
targeted regimens. Given the context dependence of stromal biology,
careful attention to dosing, scheduling and readouts of on-target
activity (e.g., cleaved caspase-3 and ER-stress biomarkers in CAFs,
alongside stromal remodeling endpoints) will be essential to
maximize antitumor benefit while preserving necessary tissue
homeostasis, without altering the fundamental rationale and
evidence base aforementioned.
Autophagy and CAFs
Autophagy is a catabolic process in which
cytoplasmic proteins or organelles are sequestered into
double-membrane vesicles and subsequently fuse with lysosomes to
form autolysosomes that degrade their contents. Emerging evidence
indicates that autophagy contributes to the metabolic and signaling
interplay between tumors and CAFs, with CAFs promoting tumor growth
by supplying nutrients to cancer cells through autophagic activity
(143). Consistent associations
between autophagy and CAF progression have been reported across
breast, and prostate, cancers, supporting autophagy as a tractable
node for modulating CAFs-mediated tumor support (144). Conceptually, therapeutic benefit
can arise from autophagy modulation in either direction, but most
current efforts emphasize inhibition of autophagy within CAFs to
blunt their pro-tumor functions, while in selected contexts
induction of autophagy in cancer or stromal cells can also
counteract CAF-driven malignant behaviors (134).
Among autophagy inhibitors, chloroquine has been
shown to abrogate the stimulatory effects of CAFs on tumor growth
and to suppress the pro-carcinogenic homologous TLR4/NF-κB
signaling pathway in a mouse xenograft model (145). In HCC models, chloroquine also
inhibits CAFs-induced activation of autophagic flux in cancer
cells, leading to light chain (LC)3-II accumulation and increased
LC3 puncta, thereby preventing CAFs from enhancing stem cell-like
properties in these cells (146). In parallel, matrix caveolin-1
(Cav-1) deficiency has emerged as a stromal biomarker of CAFs
associated with poor outcomes in breast cancer and ductal carcinoma
in situ; chloroquine or other autophagy/lysosomal inhibitors
may act as anticancer agents by therapeutically restoring stromal
Cav-1 expression in CAFs (147).
Complementing these inhibitory strategies, several interventions
leverage autophagy induction to disrupt CAF-related tumor support:
Polygonatum polysaccharides extracted from Chinese herbs stimulate
autophagy and inhibit the growth of prostate CAFs, suggesting a
novel approach centered on limiting prostate CAF expansion
(148); the naturally occurring
polyphenol resveratrol counters IL-6-induced migration of ovarian
cancer cells by inducing autophagy in migrating frontier cells,
thereby attenuating invasive and metastatic behavior (149); and resveratrol together with its
analogue 4,4'-dihydroxy-trans-stilbene reduces lung carcinoma tumor
mass through enhanced autophagy, which blocks the phenotypic
transformation of normal fibroblasts into CAFs (150).
Taken together, these findings delineate autophagy
as a context-dependent lever in the tumor-stroma ecosystem:
CAF-driven autophagy can fuel tumor growth, yet judicious
modulation, predominantly inhibition within CAFs and in specific
scenarios induction within cancer or stromal compartments, can
dismantle CAF-mediated support for malignancy. This logic argues
for cell type-specific, endpoint-anchored strategies that track
autophagy dynamics (such as LC3-II accumulation and puncta)
alongside stromal phenotypes such as Cav-1, enabling the selection
of inhibition vs. induction to achieve sustained suppression of
CAF-driven tumor progression without undermining essential tissue
homeostasis.
CAF heterogeneity and therapeutic risks: A
detailed analysis
CAFs exhibit pronounced heterogeneity within the
TME, as demonstrated by the research employing single-cell RNA
sequencing, spatial transcriptomics and surface marker profiling
(151). Distinct CAF
subpopulations with discrete molecular signatures and functional
phenotypes have been identified across various malignancies,
including breast cancer, PDAC and NSCLC. Representative CAF subsets
comprise inflammatory CAFs (iCAFs), characterized by high
expression of pro-inflammatory cytokines such as IL-6 and CXCL1
(138); myCAFs, which express
elevated α-SMA and are enriched in ECM components; and
antigen-presenting CAFs (apCAFs), expressing major
histocompatibility class II molecules capable of modulating local
immune responses (152).
These CAF subsets differ not only in their spatial
distribution but also in the signaling pathways they engage with
and their influence on tumor biology. For instance, myCAFs
predominantly rely on TGF-β signaling to maintain their
contractile, matrix-remodeling phenotype, thereby contributing to
fibrotic stroma formation (153). Conversely, iCAFs exhibit
activation of the JAK/STAT3 pathway and secrete inflammatory
factors that foster an immunosuppressive milieu (154). Importantly, despite the general
tumor-promoting perception of CAFs, emerging evidence indicates
that certain CAF subsets may exert tumor-restraining functions in
specific contexts. For example, myCAFs have been reported to form
physical barriers limiting tumor cell invasion, while apCAFs may
affect anti-tumor immunity through antigen presentation mechanisms
(155).
Given this functional complexity, non-selective
depletion of CAFs poses significant therapeutic risks. In PDAC
mouse models, ablation of α-SMA+ myCAFs paradoxically accelerated
tumor progression and enhanced immune evasion, indicating a
protective role of this subset under certain conditions (156). Broad CAF elimination may disrupt
stromal barriers, facilitating tumor dissemination or triggering
maladaptive remodeling of the TME, thereby exacerbating disease. By
contrast, functional reprogramming strategies, such as inhibition
of TGF-β and IL-6 signaling, have demonstrated potential to convert
pro-tumorigenic CAFs into a more inert or tumor-restraining
phenotype, mitigating adverse effects linked to CAF clearance. This
approach has thus gained increasing attention as a more nuanced
therapeutic avenue (157).
Accordingly, future CAF-targeted interventions must
be founded on precise subtype identification and functional
stratification. Defining subtype-specific signaling pathways and
markers will enable selective targeting that minimizes collateral
damage and improves therapeutic efficacy. Furthermore, advances in
single-cell technologies and mechanistic studies are essential to
elucidate the dynamic regulation of CAF phenotypes, bridging the
gap from indiscriminate ablation toward sophisticated functional
modulation of tumor stroma.
Discussion
The present review synthesized well-characterized
CAF-associated signaling pathways, including FAP, PI3K/AKT/mTOR,
JAK/STAT, NF-κB, TGF-β, ferroptosis, autophagy and apoptosis and
highlighted small-molecule modulators that act on these axes with
the potential to eliminate or functionally disable CAFs, thereby
supporting cancer therapy. Although biologics have progressed
rapidly, small molecules remain essential in clinical practice due
to lower cost and simpler generic manufacturing compared with
monoclonal antibodies (158) and
the present study therefore summarized these agents in Tables I-VII. As most chemotherapeutics induce
varying degrees of toxicity (159-161), an appropriate balance between
anticancer efficacy and adverse effects is necessary when targeting
CAFs; in this context, natural compounds, typically characterized
by lower toxicity and multitarget capacity, offer a promising
anti-CAFs strategy despite the lengthy path from discovery to
clinical use (162-165). Drug repurposing of approved
agents, whose pharmacology and safety are well defined, may also
provide a shortcut that reduces attrition and economic burden.
Notably, FDA-approved drugs with reported anti-CAFs activity
include modulators of PI3K/AKT/mTOR (such as Oxaliplatin, CDDP,
Capmatinib, Regorafenib and Caffeine), JAK/STAT (Tocilizumab),
TGF-β (Eribulin, Aspirin, Ibuprofen, Met, Digoxin, Pirfenidone and
Nintedanib), NF-κB (Chloroquine and Hydroxychloroquine), autophagy
(Chloroquine) and apoptosis (Pirfenidone, Bortezomib, Panobinostat
and DSF/Cu) and several have entered clinical evaluation. In
addition, numerous natural small molecules or prescription
formulations show good tolerance but require further validation of
efficacy and safety, such as PI3K/AKT/mTOR modulators (Biejiajian
pill, Tiplaxtinin, gold nanoparticles, Mycophenolic acid,
Everolimus, Dihydromyricetin and MSI-N1014), JAK/STAT modulators
(Bufalin, Tranilast, Curcumin, Oroxylin A, Resveratrol, Berberine,
Lentinus edodes polysaccharide and β-Carotene), TGF-β modulators
(HA-PTX/MATT-LTSLs HNPs, LY2157299 monohydrate, LY2109761,
Halofuginone, bone morphogenetic protein and activin membrane-bound
inhibitor, Artemisinin derivatives, Δ9-Tetrahydrocannabinol and
Cannabidiol, Aesculus hippocastanum L. extract, Curcumin,
Chelerythrine chloride, Silibinin, Evodiamine and Berberine), NF-κB
modulators (Melatonin, JPJDR, Lentinus edodes polysaccharides,
Ligustilide), FAP modulators (PT630, AMD3100, αFAP-PE38,
drug-loaded CAP-NPs and αFAP-Z@FRT), autophagy modulators
(Glycyrrhizin, Polygonatum polysaccharides and Resveratrol),
apoptosis modulators [Curcumin, Cinnamaldehyde, Nab-paclitaxel,
ONP-302 nanoparticles, nanoliposome with navitoclax, nanoemulsions
carrying doxorubicin (DOX) and small interfering RNA (siRNA)] and
ferroptosis modulators (FER-1, RSL-3 and eNVs-FAP). The present
study also listed CAF-targeting compounds in phase I trials,
including FAP modulators such as Sibrotuzumab and 131I-mAbF19;
notably, Sibrotuzumab was reported to target and block activation
of FAP-positive CAFs to inhibit tumor growth. Most other agents
remain at the preclinical stage yet exhibit anti-CAF activity,
suggesting that dose optimization, rational combinations and
structure-guided derivative development could enhance utility,
while the potential for significant side effects underscores the
need for rigorous dose-finding and toxicity monitoring. Overall,
current evidence indicates that CAF-targeting compounds show
acceptable tolerability in mice or patients and multitarget
strategies spanning several oncogenic pathways may play a
meaningful role in forthcoming clinical studies. The most tractable
path to translation is to deploy these agents as short,
schedule-defined CAF-priming interventions that transiently soften
the matrix and reduce immune exclusion and to pair them with
targeted delivery platforms that concentrate activity within the
stroma; this approach retains the advantages of small molecules
while mitigating risks to wound healing and systemic
homeostasis.
Growing mechanistic evidence supports the use of
small-molecule compounds targeting FAP, PI3K/AKT/mTOR, JAK/STAT,
NF-κB, TGF-β and autophagy to suppress CAF function and several of
these agents are entering clinical evaluation. Nevertheless, most
studies to date have been conducted at cellular and murine levels
and trials explicitly designed to assess CAF-targeting small
molecules with CAF-specific clinical endpoints remain limited.
Small molecules directed at FAP demonstrate notable specificity and
potent inhibitory activity against CAFs, positioning FAP as a
promising clinical target to enable development of anticancer
therapies. As aberrantly activated CAFs engage diverse and
context-dependent signaling pathways, single-agent strategies may
be insufficient; rational combination regimens that concurrently
target multiple pathways are likely to yield superior outcomes.
Importantly, targeting CAFs alone is generally not
adequate to treat cancer, as numerous CAF-focused small molecules
do not directly kill cancer cells; combining CAF-directed agents
with cytotoxic chemotherapy or targeted therapies has the potential
to improve efficacy during tumor development (166). Looking ahead, a coherent
framework emerges from the available data and our interpretation:
CAF-focused small molecules should be integrated into combination
regimens with clear pharmacodynamic anchors, reductions in matrix
stiffness and collagen crosslinking, improved perfusion and
enhanced effector immune-cell infiltration, to demonstrate
on-mechanism stromal remodeling (10); patient selection ought to reflect
CAF heterogeneity by matching drugs to dominant CAF programs such
as FAP-high or TGF-β-driven phenotypes (167); dosing should favor
window-of-opportunity or intermittent schedules to balance efficacy
with the physiological roles of targets; and sequential strategies
that pair brief CAF suppression or apoptosis with subsequent
normalization or reprogramming may stabilize a permissive
microenvironment and reduce rebound once drug pressure is
withdrawn. These directions are supported by the dual realities
that CAFs are plastic and context-dependent and that their
modulation exerts system-level effects on drug delivery and immune
accessibility (168). The open
questions, namely which CAF subsets should be eliminated vs.
functionally suppressed, how to anticipate and prevent adaptive
escape and how to optimally synchronize CAFs priming with
chemotherapy, radiotherapy and immunotherapy, are best answered
through CAF-centric trial designs that embed stromal endpoints and
leverage targeted delivery to widen the therapeutic window without
altering the fundamental clinical rationale aforementioned
(169).
CAFs exhibit significant heterogeneity within the
TME, encompassing diverse subtypes with distinct molecular and
functional characteristics. Advances in single-cell sequencing and
spatial transcriptomics have identified major CAF subsets,
including iCAFs, myCAFs and apCAFs, which differ in spatial
distribution and active signaling pathways. MyCAFs maintain ECM
remodeling largely via TGF-β signaling, whereas iCAFs rely on the
JAK/STAT3 pathway to secrete pro-inflammatory factors that foster
an immunosuppressive milieu (170). Notably, some CAFs subsets may
exert tumor-restraining effects in specific contexts, such as
myCAFs forming physical barriers that limit tumor invasion
(40). Given this complexity,
non-selective depletion of CAFs can inadvertently accelerate tumor
progression and immune evasion, underscoring potential risks of
broad CAF targeting. Therefore, precise identification and
functional modulation of CAFs subtypes are critical for developing
safer and more effective therapeutic strategies, with reprogramming
of CAF phenotypes through pathway-specific interventions emerging
as a promising approach.
In summary, CAFs serve as essential regulators
within the TME, playing critical roles in tumor progression, immune
suppression and therapy resistance. With increasingly comprehensive
insights into their functions and mechanisms, emerging
CAFs-targeted therapeutics, including small molecule inhibitors,
monoclonal antibodies, immune modulators and gene editing
technologies, have shown promise not only in disrupting pathogenic
signaling between CAFs and cancer cells but also in effectively
remodeling the TME to restore immune competence (171). However, despite these
encouraging advances, several challenges remain for the clinical
translation of these strategies. Ensuring safety and efficacy
requires extensive validation through large-scale clinical trials,
given the biological heterogeneity across patient populations that
may affect therapeutic responses. Furthermore, complexities in
manufacturing, quality control and delivery of such agents pose
significant obstacles to widespread clinical application. Future
research must leverage cutting-edge technologies such as
single-cell sequencing, spatial omics and high-throughput screening
to comprehensively characterize the cellular states and dynamic
changes of CAFs across diverse tumor contexts, delineating their
precise functions and regulatory networks. Systematic elucidation
of the complex crosstalk between CAFs and other cellular components
such as immune cells and cancer stem cells will uncover novel
synergistic therapeutic targets and enable personalized precision
medicine approaches. Realistic recapitulation of the in vivo
tumor milieu through advanced in vitro models and
genetically engineered animal systems remains indispensable for
rigorous evaluation of drug safety and efficacy. Furthermore,
exploring combinational regimens of CAF-targeted agents with
existing radiotherapy, chemotherapy and immunotherapy represents a
crucial strategy to overcome resistance and enhance treatment
outcomes. Clinical trial designs must prioritize patient
stratification and biomarker integration to improve response
precision and maximize clinical benefit. Overall, addressing these
clinical translation challenges through the tight integration of
fundamental research and clinical application will accelerate the
development and translation of CAF-targeting drugs, ultimately
providing cancer patients with safer, more effective and tailored
therapeutic options, thereby advancing precision oncology into a
new era.
Availability of data and materials
Not applicable.
Authors' contributions
YW contributed to the drafting of the manuscript
and was a major contributor in writing. XQL designed, conceived and
supervised the study and revised the manuscript. DL conducted the
literature search and analysis. KDL generated the figures. DDZ
generated the tables. Data authentication is not applicable. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
No funding was received.
References
|
1
|
Singh D, Vignat J, Lorenzoni V, Eslahi M,
Ginsburg O, Lauby-Secretan B, Arbyn M, Basu P, Bray F and
Vaccarella S: Global estimates of incidence and mortality of
cervical cancer in 2020: A baseline analysis of the WHO Global
Cervical Cancer Elimination Initiative. Lancet Glob Health.
11:E197–E206. 2023. View Article : Google Scholar :
|
|
2
|
Fang H and Declerck YA: Targeting the
tumor microenvironment: From understanding pathways to effective
clinical trials. Cancer Res. 73:4965–4977. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
de Visser KE and Joyce JA: The evolving
tumor microenvironment: From cancer initiation to metastatic
outgrowth. Cancer Cell. 41:374–403. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rimal R, Desai P, Daware R, Hosseinnejad
A, Prakash J, Lammers T and Singh S: Cancer-associated fibroblasts:
Origin, function, imaging, and therapeutic targeting. Adv Drug
Deliv Rev. 189:1145042022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yoshida GJ, Azuma A, Miura Y and Orimo A:
Activated fibroblast program orchestrates tumor initiation and
progression; molecular mechanisms and the associated therapeutic
strategies. Int J Mol Sci. 20:22562019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
LeBleu VS and Kalluri R: A peek into
cancer-associated fibroblasts: Origins, functions and translational
impact. Dis Model Mech. 11:dmm0294472018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Biffi G and Tuveson DA: Diversity and
biology of cancer-associated fibroblasts. Physiol Rev. 101:147–176.
2021. View Article : Google Scholar
|
|
9
|
Gascard P and Tlsty TD:
Carcinoma-associated fibroblasts: Orchestrating the composition of
malignancy. Genes Dev. 30:1002–1019. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yoshida GJ: Regulation of heterogeneous
cancer-associated fibroblasts: The molecular pathology of activated
signaling pathways. J Exp Clin Cancer Res. 39:1122020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Z, Sun C and Qin Z: Metabolic
reprogramming of cancer-associated fibroblasts and its effect on
cancer cell reprogramming. Theranostics. 11:8322–8336. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fiori ME, Di Franco S, Villanova L, Bianca
P, Stassi G and De Maria R: Cancer-associated fibroblasts as
abettors of tumor progression at the crossroads of EMT and therapy
resistance. Mol Cancer. 18:702019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goulet CR, Champagne A, Bernard G, Vandal
D, Chabaud S, Pouliot F and Bolduc S: Cancer-associated fibroblasts
induce epithelial-mesenchymal transition of bladder cancer cells
through paracrine IL-6 signalling. BMC Cancer. 19:1372019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu F, Yang J, Liu J, Wang Y, Mu J, Zeng Q,
Deng S and Zhou H: Signaling pathways in cancer-associated
fibroblasts and targeted therapy for cancer. Signal Transduct
Target Ther. 6:2182021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ping Q, Yan R, Cheng X, Wang W, Zhong Y,
Hou Z, Shi Y, Wang C and Li R: Cancer-associated fibroblasts:
Overview, progress, challenges, and directions. Cancer Gene Ther.
28:984–999. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Zhang Q, Wu Q, Cui Y, Zhu H, Fang M,
Zhou X, Sun Z and Yu J: Interleukin-22 secreted by
cancer-associated fibroblasts regulates the proliferation and
metastasis of lung cancer cells via the PI3K-Akt-mTOR signaling
pathway. Am J Transl Res. 11:4077–4088. 2019.PubMed/NCBI
|
|
17
|
Yu L, Wei J and Liu P: Attacking the
PI3K/Akt/mTOR signaling pathway for targeted therapeutic treatment
in human cancer. Semin Cancer Biol. 85:69–94. 2022. View Article : Google Scholar
|
|
18
|
Tang S, Hou Y, Zhang H, Tu G, Yang L, Sun
Y, Lang L, Tang X, Du YE, Zhou M, et al: Oxidized ATM promotes
abnormal proliferation of breast CAFs through maintaining
intracellular redox homeostasis and activating the PI3K-AKT,
MEK-ERK, and Wnt-β-catenin signaling pathways. Cell Cycle.
14:1908–1924. 2015. View Article : Google Scholar
|
|
19
|
Ning X, Zhang H, Wang C and Song X:
Exosomes released by gastric cancer cells induce transition of
pericytes into cancer-associated fibroblasts. Med Sci Monit.
24:2350–2359. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Y, Ren H, Dai B, Li J, Shang L, Huang
J and Shi X: Hepatocellular carcinoma-derived exosomal miRNA-21
contributes to tumor progression by converting hepatocyte stellate
cells tocancer-associated fibroblasts. J Exp Clin Cancer Res.
41:3592022. View Article : Google Scholar
|
|
21
|
Wang YM, Wang W and Qiu ED: Osteosarcoma
cells induce differentiation of mesenchymal stem cells into cancer
associated fibroblasts through Notch and Akt signaling pathway. Int
J Clin Exp Pathol. 10:8479–8486. 2017.PubMed/NCBI
|
|
22
|
Zang C, Eucker J, Habbel P, Neumann C,
Schulz CO, Bangemann N, Kissner L, Riess H and Liu H: Targeting
multiple tyrosine kinase receptors with Dovitinib blocks invasion
and the interaction between tumor cells and cancer-associated
fibroblasts in breast cancer. Cell Cycle. 14:1291–1299. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li J, Gu X, Wan G, Wang Y, Chen K, Chen Q
and Lu C: Rocuronium bromide suppresses esophageal cancer via
blocking the secretion of C-X-C motif chemokine ligand 12 from
cancer associated fibroblasts. J Transl Med. 21:2482023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu C and Zhang C: Oxaliplatin inhibits
colorectal cancer progression by inhibiting CXCL11 secreted by
cancer-associated fibroblasts and the CXCR3/PI3K/AKT pathway. Clin
Transl Oncol. 25:160–172. 2023. View Article : Google Scholar
|
|
25
|
Chen W, Yang X, Sun J, Chen Y, Zhao W, He
C, An H, Pang J, Xu W, Wen B, et al: Biejiajian pill inhibits
progression of hepatocellular carcinoma by downregulating PDGFRβ
signaling in cancer-associated fibroblasts. J Ethnopharmacol.
301:1158252023. View Article : Google Scholar
|
|
26
|
Li X, Song Q, Guo X, Wang L, Zhang Q, Cao
L, Ren Y, Wu X, Meng Z and Xu K: The metastasis potential promoting
capacity of cancer-associated fibroblasts was attenuated by
cisplatin via modulating KRT8. Onco Targets Ther. 13:2711–2723.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Che Y, Wang J, Li Y, Lu Z, Huang J, Sun S,
Mao S, Lei Y, Zang R, Sun N and He J: Cisplatin-activated PAI-1
secretion in the cancer-associated fibroblasts with paracrine
effects promoting esophageal squamous cell carcinoma progression
and causing chemoresistance. Cell Death Dis. 9:7592018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao X, Pan J, Li W, Yang W, Qin L and Pan
Y: Gold nanoparticles enhance cisplatin delivery and potentiate
chemotherapy by decompressing colorectal cancer vessels. Int J
Nanomedicine. 13:6207–6221. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu K, Lv Z, Xiong J, Zheng H, Zhang S,
Jin H, Yu L, Li Z, Zhang J, Li C and Liang P: MET inhibitor,
capmatinib overcomes osimertinib resistance via suppression of
MET/Akt/snail signaling in non-small cell lung cancer and decreased
generation of cancer-associated fibroblasts. Aging (Albany NY).
13:6890–6903. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Heits N, Heinze T, Bernsmeier A, Kerber J,
Hauser C, Becker T, Kalthoff H, Egberts JH and Braun F: Influence
of mTOR-inhibitors and mycophenolic acid on human cholangiocellular
carcinoma and cancer associated fibroblasts. BMC Cancer.
16:3222016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang M, Li X, Wu W, Gao J, Han Q, Sun Z
and Zhao RC: Regorafenib induces the apoptosis of gastrointestinal
cancer-associated fibroblasts by inhibiting AKT phosphorylation.
Stem Cells Dev. 31:383–394. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang H, You T, Ge H, Gao J, Wang Y, Bai C,
Sun Z, Han Q and Zhao RC: Anlotinib may enhance the efficacy of
anti-PD1 therapy by inhibiting the AKT pathway and promoting the
apoptosis of CAFs in lung adenocarcinoma. Int Immunopharmacol.
133:1120532024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Al-Ansari MM and Aboussekhra A: Caffeine
mediates sustained inactivation of breast cancer-associated
myofibroblasts via up-regulation of tumor suppressor genes. PLoS
One. 9:e909072014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fan KJ, Yang B, Liu Y, Tian XD and Wang B:
Inhibition of human lung cancer proliferation through targeting
stromal fibroblasts by dihydromyricetin. Mol Med Rep. 16:9758–9762.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yadav VK, Huang YJ, George TA, Wei PL,
Sumitra MR, Ho CL, Chang TH, Wu ATH and Huang HS: Preclinical
evaluation of the novel small-molecule MSI-N1014 for treating
drug-resistant colon cancer via the LGR5/β-catenin/miR-142-3p
network and reducing cancer-associated fibroblast transformation.
Cancers (Basel). 12:15902020. View Article : Google Scholar
|
|
36
|
Harrison DA: The Jak/STAT pathway. Cold
Spring Harb Perspect Biol. 4:a0112052012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Morris R, Kershaw NJ and Babon JJ: The
molecular details of cytokine signaling via the JAK/STAT pathway.
Protein Sci. 27:1984–2009. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Subramaniam KS, Omar IS, Kwong SC, Mohamed
Z, Woo YL, Mat Adenan NA and Chung I: Cancer-associated fibroblasts
promote endometrial cancer growth via activation of
interleukin-6/STAT-3/c-Myc pathway. Am J Cancer Res. 6:200–213.
2016.PubMed/NCBI
|
|
39
|
Cheteh EH, Sarne V, Ceder S, Bianchi J,
Augsten M, Rundqvist H, Egevad L, Östman A and Wiman KG:
Interleukin-6 derived from cancer-associated fibroblasts attenuates
the p53 response to doxorubicin in prostate cancer cells. Cell
Death Discov. 6:422020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Biffi G, Oni TE, Spielman B, Hao Y, Elyada
E, Park Y, Preall J and Tuveson DA: IL1-Induced JAK/STAT Signaling
Is Antagonized by TGFβ to Shape CAF Heterogeneity in Pancreatic
Ductal Adenocarcinoma. Cancer Discov. 9:282–301. 2019. View Article : Google Scholar
|
|
41
|
Wang H, Chen J, Li S, Yang J, Tang D, Wu
W, Yu K, Cao Y, Xu K, Yin P, et al: Bufalin reverses
cancer-associated fibroblast-mediated colorectal cancer metastasis
by inhibiting the STAT3 signaling pathway. Apoptosis. 28:594–606.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ochi K, Suzawa K, Thu YM, Takatsu F,
Tsudaka S, Zhu Y, Nakata K, Takeda T, Shien K, Yamamoto H, et al:
Drug repositioning of tranilast to sensitize a cancer therapy by
targeting cancer-associated fibroblast. Cancer Sci. 113:3428–3436.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Al-Jomah N, Al-Mohanna FH and Aboussekhra
A: Tocilizumab suppresses the pro-carcinogenic effects of breast
cancer-associated fibroblasts through inhibition of the STAT3/AUF1
pathway. Carcinogenesis. 42:1439–1448. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ham IH, Wang L, Lee D, Woo J, Kim TH,
Jeong HY, Oh HJ, Choi KS, Kim TM and Hur H: Curcumin inhibits the
cancer-associated fibroblast-derived chemoresistance of gastric
cancer through the suppression of the JAK/STAT3 signaling pathway.
Int J Oncol. 61:852022. View Article : Google Scholar
|
|
45
|
Cao Y, Cao W, Qiu Y, Zhou Y, Guo Q, Gao Y
and Lu N: Oroxylin A suppresses ACTN1 expression to inactivate
cancer-associated fibroblasts and restrain breast cancer
metastasis. Pharmacol Res. 159:1049812020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Suh J, Kim DH and Surh YJ: Resveratrol
suppresses migration, invasion and stemness of human breast cancer
cells by interfering with tumor-stromal cross-talk. Arch Biochem
Biophys. 643:62–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tsang CM, Cheung YC, Lui VW, Yip YL, Zhang
G, Lin VW, Cheung KC, Feng Y and Tsao SW: Berberine suppresses
tumorigenicity and growth of nasopharyngeal carcinoma cells by
inhibiting STAT3 activation induced by tumor associated
fibroblasts. BMC Cancer. 13:6192013. View Article : Google Scholar
|
|
48
|
Zhang W, Tao N and Bai L: Polysaccharides
from Lentinus edodes prevent acquired drug resistance to docetaxel
in prostate cancer cells by decreasing the TGF-β1 secretion of
cancer-associated fibroblasts. J Nat Med. 77:817–828. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lee NY and Kim Y, Kim YS, Shin JH, Rubin
LP and Kim Y: β-Carotene exerts anti-colon cancer effects by
regulating M2 macrophages and activated fibroblasts. J Nutr
Biochem. 82:1084022020. View Article : Google Scholar
|
|
50
|
Ringuette Goulet C, Bernard G, Tremblay S,
Chabaud S, Bolduc S and Pouliot F: Exosomes induce fibroblast
differentiation into cancer-associated fibroblasts through TGFβ
signaling. Mol Cancer Res. 16:1196–1204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tan HX, Gong WZ, Zhou K, Xiao ZG, Hou FT,
Huang T, Zhang L, Dong HY, Zhang WL, Liu Y, et al: CXCR4/TGF-β1
mediated hepatic stellate cells differentiation into
carcinoma-associated fibroblasts and promoted liver metastasis of
colon cancer. Cancer Biol Ther. 21:258–268. 2020. View Article : Google Scholar
|
|
52
|
Wu YH, Huang YF, Chang TH, Chen CC, Wu PY,
Huang SC and Chou CY: COL11A1 activates cancer-associated
fibroblasts by modulating TGF-β3 through the NF-κB/IGFBP2 axis in
ovarian cancer cells. Oncogene. 40:4503–4519. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Barcellos-de-Souza P, Comito G,
Pons-Segura C, Taddei ML, Gori V, Becherucci V, Bambi F, Margheri
F, Laurenzana A, Del Rosso M and Chiarugi P: Mesenchymal stem cells
are recruited and activated into carcinoma-associated fibroblasts
by prostate cancer microenvironment-derived TGF-β1. Stem Cells.
34:2536–2547. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Karagiannis GS, Schaeffer DF, Cho CK,
Musrap N, Saraon P, Batruch I, Grin A, Mitrovic B, Kirsch R,
Riddell RH and Diamandis EP: Collective migration of
cancer-associated fibroblasts is enhanced by overexpression of
tight junction-associated proteins claudin-11 and occludin. Mol
Oncol. 8:178–195. 2014. View Article : Google Scholar
|
|
55
|
Shi X, Young CD, Zhou H and Wang X:
Transforming growth factor-β signaling in fibrotic diseases and
cancer-associated fibroblasts. Biomolecules. 10:16662020.
View Article : Google Scholar
|
|
56
|
Luong T and Cukierman E: Eribulin
normalizes pancreatic cancer-associated fibroblasts by simulating
selected features of TGFβ inhibition. BMC Cancer. 22:12552022.
View Article : Google Scholar
|
|
57
|
Lv Y, Xu C, Zhao X, Lin C, Yang X, Xin X,
Zhang L, Qin C, Han X, Yang L, et al: Nanoplatform Assembled from a
CD44-Targeted prodrug and smart liposomes for dual targeting of
tumor microenvironment and cancer cells. ACS Nano. 12:1519–1536.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang Q, Hou X, Evans BJ, VanBlaricom JL,
Weroha SJ and Cliby WA: LY2157299 Monohydrate, a TGF-βR1 inhibitor,
suppresses tumor growth and ascites development in ovarian cancer.
Cancers (Basel). 10:2602018. View Article : Google Scholar
|
|
59
|
Liu FL, Mo EP, Yang L, Du J, Wang HS,
Zhang H, Kurihara H, Xu J and Cai SH: Autophagy is involved in
TGF-β1-induced protective mechanisms and formation of
cancer-associated fibroblasts phenotype in tumor microenvironment.
Oncotarget. 7:4122–4141. 2016. View Article : Google Scholar :
|
|
60
|
Wang H, Liu F, Wu X, Zhu G, Tang Z, Qu W,
Zhao Q, Huang R, Tian M, Fang Y, et al: Cancer-associated
fibroblasts contributed to hepatocellular carcinoma recurrence and
metastasis via CD36-mediated fatty-acid metabolic reprogramming.
Exp Cell Res. 435:1139472024. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wawro ME, Sobierajska K, Ciszewski WM and
Niewiarowska J: Nonsteroidal anti-inflammatory drugs prevent
vincristine-dependent cancer-associated fibroblasts formation. Int
J Mol Sci. 20:19412019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Shao S, Zhao L, An G, Zhang L, Jing X, Luo
M, Li W, Meng D, Ning Q, Zhao X and Lei J: Metformin suppresses
HIF-1α expression in cancer-associated fibroblasts to prevent
tumor-stromal cross talk in breast cancer. FASEB J. 34:10860–10870.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Coleman DT, Gray AL, Stephens CA, Scott ML
and Cardelli JA: Repurposed drug screen identifies cardiac
glycosides as inhibitors of TGF-β-induced cancer-associated
fibroblast differentiation. Oncotarget. 7:32200–32209. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Takai K, Le A, Weaver VM and Werb Z:
Targeting the cancer-associated fibroblasts as a treatment in
triple-negative breast cancer. Oncotarget. 7:82889–82901. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mediavilla-Varela M, Boateng K, Noyes D
and Antonia SJ: The anti-fibrotic agent pirfenidone synergizes with
cisplatin in killing tumor cells and cancer-associated fibroblasts.
BMC Cancer. 16:1762016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Mazzocca A, Fransvea E, Dituri F, Lupo L,
Antonaci S and Giannelli G: Down-regulation of connective tissue
growth factor by inhibition of transforming growth factor beta
blocks the tumor-stroma cross-talk and tumor progression in
hepatocellular carcinoma. Hepatology. 51:523–534. 2010. View Article : Google Scholar
|
|
67
|
Wang D, Tian M, Fu Y, Sun Y, Ding L, Zhang
X, Jing Y, Sun G, Ni Y and Song Y: Halofuginone inhibits tumor
migration and invasion by affecting cancer-associated fibroblasts
in oral squamous cell carcinoma. Front Pharmacol. 13:10563372022.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gabasa M, Ikemori R, Hilberg F, Reguart N
and Alcaraz J: Nintedanib selectively inhibits the activation and
tumour-promoting effects of fibroblasts from lung adenocarcinoma
patients. Br J Cancer. 117:1128–1138. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Shangguan L, Ti X, Krause U, Hai B, Zhao
Y, Yang Z and Liu F: Inhibition of TGF-β/Smad signaling by BAMBI
blocks differentiation of human mesenchymal stem cells to
carcinoma-associated fibroblasts and abolishes their protumor
effects. Stem Cells. 30:2810–2819. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yao Y, Guo Q, Cao Y, Qiu Y, Tan R, Yu Z,
Zhou Y and Lu N: Correction to: Artemisinin derivatives inactivate
cancer-associated fibroblasts through suppressing TGF-β signaling
in breast cancer. J Exp Clin Cancer Res. 38:4512019. View Article : Google Scholar
|
|
71
|
Milián L, Monleón-Guinot I, Sancho-Tello
M, Galbis JM, Cremades A, Almenar-Ordaz M, Peñaroja-Martinez J,
Farras R, Martín de Llano JJ, Carda C and Mata M: In vitro effect
of Δ9-tetrahydrocannabinol and cannabidiol on cancer-associated
fibroblasts isolated from lung cancer. Int J Mol Sci. 23:67662022.
View Article : Google Scholar
|
|
72
|
Melegová N, Čoma M, Urban L, Kaňuchová M,
Szabo P, Smetana K Jr, Mučaji P and Gál P: Aesculus hippocastanum
L. extract differently modulates normal human dermal fibroblasts
and cancer-associated fibroblasts from basal/squamous cell
carcinoma. Neoplasma. 69:224–232. 2022. View Article : Google Scholar
|
|
73
|
Buhrmann C, Kraehe P, Lueders C, Shayan P,
Goel A and Shakibaei M: Curcumin suppresses crosstalk between colon
cancer stem cells and stromal fibroblasts in the tumor
microenvironment: Potential role of EMT. PLoS One. 9:e1075142014.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Liang D, Liu L, Zheng Q, Zhao M, Zhang G,
Tang S, Tang J and Chen N: Chelerythrine chloride inhibits the
progression of colorectal cancer by targeting cancer-associated
fibroblasts through intervention with WNT10B/β-catenin and
TGFβ2/Smad2/3 axis. Phytother Res. 37:4674–4689. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Jalilian E, Abolhasani-Zadeh F, Afgar A,
Samoudi A, Zeinalynezhad H and Langroudi L: Neutralizing
tumor-related inflammation and reprogramming of cancer-associated
fibroblasts by Curcumin in breast cancer therapy. Sci Rep.
13:207702023. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ting H, Deep G, Kumar S, Jain AK, Agarwal
C and Agarwal R: Beneficial effects of the naturally occurring
flavonoid silibinin on the prostate cancer microenvironment: Role
of monocyte chemotactic protein-1 and immune cell recruitment.
Carcinogenesis. 37:589–599. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ting HJ, Deep G, Jain AK, Cimic A,
Sirintrapun J, Romero LM, Cramer SD, Agarwal C and Agarwal R:
Silibinin prevents prostate cancer cell-mediated differentiation of
naïve fibroblasts into cancer-associated fibroblast phenotype by
targeting TGF β2. Mol Carcinog. 54:730–741. 2015. View Article : Google Scholar
|
|
78
|
Zheng F, Luo Y, Liu Y, Gao Y, Chen W and
Wei K: Nano-baicalein facilitates chemotherapy in breast cancer by
targeting tumor microenvironment. Int J Pharm. 635:1227782023.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Pang N, Yang Z, Zhang W, Du Y, Zhang L, Li
X, Peng Y and Qi X: Cancer-associated fibroblasts barrier breaking
via TGF-β blockade paved way for docetaxel micelles delivery to
treat pancreatic cancer. Int J Pharm. 665:1247062024. View Article : Google Scholar
|
|
80
|
Oeckinghaus A, Hayden MS and Ghosh S:
Crosstalk in NF-κB signaling pathways. Nat Immunol. 12:695–708.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zheng S, Hu C, Lin H, Li G, Xia R, Zhang
X, Su D, Li Z, Zhou Q and Chen R: circCUL2 induces an inflammatory
CAF phenotype in pancreatic ductal adenocarcinoma via the
activation of the MyD88-dependent NF-κB signaling pathway. J Exp
Clin Cancer Res. 41:712022. View Article : Google Scholar
|
|
82
|
Gu J, Li X, Zhao L, Yang Y, Xue C, Gao Y,
Li J, Han Q, Sun Z, Bai C and Zhao RC: The role of PKM2 nuclear
translocation in the constant activation of the NF-κB signaling
pathway in cancer-associated fibroblasts. Cell Death Dis.
12:2912021. View Article : Google Scholar
|
|
83
|
Zheng B, Ohuchida K, Chijiiwa Y, Zhao M,
Mizuuchi Y, Cui L, Horioka K, Ohtsuka T, Mizumoto K, Oda Y, et al:
CD146 attenuation in cancer-associated fibroblasts promotes
pancreatic cancer progression. Mol Carcinog. 55:1560–1572. 2016.
View Article : Google Scholar
|
|
84
|
Xu Q, Zhao T, Han H, Fan J and Xie W:
EIF4A3 stabilizes the expression of lncRNA AGAP2-AS1 to activate
cancer-associated fibroblasts via MyD88/NF-κb signaling. Thorac
Cancer. 14:450–461. 2023. View Article : Google Scholar :
|
|
85
|
De Sanctis JB, Charris J, Blanco Z,
Ramírez H, Martínez GP and Mijares MR: Molecular mechanisms of
chloroquine and hydroxychloroquine used in cancer therapy.
Anticancer Agents Med Chem. 23:1122–1144. 2023. View Article : Google Scholar
|
|
86
|
Liu D, Shi K, Fu M and Chen F: Melatonin
indirectly decreases gastric cancer cell proliferation and invasion
via effects on cancer-associated fibroblasts. Life Sci.
277:1194972021. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Lou M, Iwatsuki M, Wu X, Zhang W,
Matsumoto C and Baba H: Cancer-associated fibroblast-derived IL-8
Upregulates PD-L1 expression in gastric cancer through the NF-κB
pathway. Ann Surg Oncol. 31:2983–2995. 2024. View Article : Google Scholar
|
|
88
|
Jin P, Li X, Xia Y, Li H, Li X, Yang ZY,
Wang Z, Xu C, Fang T, Zhou D, et al: Bepotastine sensitizes ovarian
cancer to PARP inhibitors through suppressing NF-κB-Triggered SASP
in cancer-associated fibroblasts. Mol Cancer Ther. 22:447–458.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Li R, Zhou J, Wu X, Li H, Pu Y, Liu N, Han
Z, Zhou L, Wang Y, Zhu H, et al: Jianpi Jiedu Recipe inhibits
colorectal cancer liver metastasis via regulating ITGBL1-rich
extracellular vesicles mediated activation of cancer-associated
fibroblasts. Phytomedicine. 100:1540822022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Xu Y, Ma J, Zheng Q, Wang Y, Hu M, Ma F,
Qin Z, Lei N and Tao N: MPSSS impairs the immunosuppressive
function of cancer-associated fibroblasts via the TLR4-NF-κB
pathway. Biosci Rep. 39:BSR201821712019. View Article : Google Scholar
|
|
91
|
Ma J, Xu Y, Zheng Q, Wang Y, Hu M, Ma F,
Long H, Qin Z and Tao N: Ligustilide inhibits the activation of
cancer-associated fibroblasts. Life Sci. 218:58–64. 2019.
View Article : Google Scholar
|
|
92
|
Chen H, Han X, Zhang Y, Wang K, Liu D, Hu
Z and Wang J: Bruceine D suppresses CAF-promoted TNBC metastasis
under TNF-α stimulation by inhibiting Notch1-Jagged1/NF-κB(p65)
signaling. Phytomedicine. 123:1549282024. View Article : Google Scholar
|
|
93
|
Pan Y, Ma T, Chen D, Wang Y, Peng Y, Lu T,
Yin X, Li H, Zhang G and Wang X: Scutellaria barbata D.Don and
Scleromitrion diffusum (Willd.) R.J.Wang inhibits the progression
of triple negative breast cancer though the activation inhibition
of NF-κB triggered by CAFs-derived IL6. J Ethnopharmacol.
335:1186562024. View Article : Google Scholar
|
|
94
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Liao Z, Tan ZW, Zhu P and Tan NS:
Cancer-associated fibroblasts in tumor microenvironment-Accomplices
in tumor malignancy. Cell Immunol. 343:1037292019. View Article : Google Scholar
|
|
96
|
Scott AM, Wiseman G, Welt S, Adjei A, Lee
FT, Hopkins W, Divgi CR, Hanson LH, Mitchell P, Gansen DN, et al: A
Phase I dose-escalation study of sibrotuzumab in patients with
advanced or metastatic fibroblast activation protein-positive
cancer. Clin Cancer Res. 9:1639–1647. 2003.PubMed/NCBI
|
|
97
|
Welt S, Divgi CR, Scott AM, Garin-Chesa P,
Finn RD, Graham M, Carswell EA, Cohen A, Larson SM, Old LJ, et al:
Antibody targeting in metastatic colon cancer: A phase I study of
monoclonal antibody F19 against a cell-surface protein of reactive
tumor stromal fibroblasts. J Clin Oncol. 12:1193–1203. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Nagaraju GP, Saddala MS, Foote JB, Khaliq
AM, Masood A, Golivi Y, Bandi DSR, Sarvesh S, Reddy SP, Switchenko
J, et al: Mechanism of enhancing chemotherapy efficacy in
pancreatic ductal adenocarcinoma with paricalcitol and
hydroxychloroquine. Cell Rep Med. 6:1018812025. View Article : Google Scholar :
|
|
99
|
Wang W, Hu K, Xue J, Chen J, Du X, Zhao T,
Chen Y, Tang X, Xu L, Hao X, et al: In vivo FAP-CAR macrophages
enhance chemotherapy and immunotherapy against pancreatic cancer by
removing the fibrosis barrier. J Control Release. 384:1138882025.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Santos AM, Jung J, Aziz N, Kissil JL and
Puré E: Targeting fibroblast activation protein inhibits tumor
stromagenesis and growth in mice. J Clin Invest. 119:3613–3625.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Feig C, Jones JO, Kraman M, Wells RJ,
Deonarine A, Chan DS, Connell CM, Roberts EW, Zhao Q, Caballero OL,
et al: Targeting CXCL12 from FAP-expressing carcinoma-associated
fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic
cancer. Proc Natl Acad Sci USA. 110:20212–20217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Wang H, Wei L, Mao D, Che X, Ye X, Liu Y
and Chen Y: Combination of oxymatrine (Om) and astragaloside IV
(As) enhances the infiltration and function of TILs in
triple-negative breast cancer (TNBC). Int Immunopharmacol.
125:1110262023. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Kao CC, Shih JW, Huynh HTLK, Chang CH,
Lawal B, Iamsaard S, Azizah N, Ritmaleni R, Lin JK, Huang PY, et
al: Nutraceutical evaluation of trigonelline's therapeutic
potential by targeting bladder cancer stem cells and
cancer-associated fibroblasts via downregulation of TGFβ3/GLI2/YAP1
Signaling Hub. Int J Med Sci. 22:1194–1207. 2025. View Article : Google Scholar
|
|
104
|
Li D, Lv Y, Hu L, Sun A and Sun L:
Rosmarinic acid potentiates gefitinib in lung adenocarcinoma by
modulating interactions between cancer cells and cancer-associated
fibroblasts. Sci Rep. 15:241002025. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Eum DY, Lee C, Tran CS, Lee J, Park SY,
Jeong MS, Jin Y, Shim JW, Lee SR, Koh M, et al: Regulatory role of
Echinochrome A in cancer-associated fibroblast-mediated lung cancer
cell migration. Toxicol Res. 40:409–419. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Ji T, Zhao Y, Ding Y, Wang J, Zhao R, Lang
J, Qin H, Liu X, Shi J, Tao N, et al: Transformable peptide
nanocarriers for expeditious drug release and effective cancer
therapy via cancer-associated fibroblast activation. Angew Chem Int
Ed Engl. 55:1050–1055. 2016. View Article : Google Scholar :
|
|
107
|
Liu Z, Ji P, Liu H, Yu L, Zhang SM, Liu P,
Zhang XZ, Luo GF and Shang Z: FNIII14 peptide-enriched membrane
nanocarrier to disrupt stromal barriers through reversing CAFs for
augmenting drug penetration in tumors. Nano Lett. 23:9963–9971.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Rodponthukwaji K, Thongchot S, Deureh S,
Thongkleang T, Thaweesuvannasak M, Srichan K, Srisawat C, Thuwajit
P, Nguyen KT, Tadpetch K, et al: Development of cancer-associated
fibroblasts-targeting polymeric nanoparticles loaded with
8-O-methylfusarubin for breast cancer treatment. Int J Pharm X.
8:1002942024.PubMed/NCBI
|
|
109
|
Gao C, Jian C, Wang L, Liu Y, Xiong Y, Wu
T and Shi C: FAP-targeting biomimetic nanosystem to restore the
activated cancer-associated fibroblasts to quiescent state for
breast cancer radiotherapy. Int J Pharm. 670:1251902025. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Watanabe S, Noma K, Ohara T, Kashima H,
Sato H, Kato T, Urano S, Katsube R, Hashimoto Y, Tazawa H, et al:
Photoimmunotherapy for cancer-associated fibroblasts targeting
fibroblast activation protein in human esophageal squamous cell
carcinoma. Cancer Biol Ther. 20:1234–1248. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhen Z, Tang W, Wang M, Zhou S, Wang H, Wu
Z, Hao Z, Li Z, Liu L and Xie J: Protein nanocage mediated
fibroblast-activation protein targeted photoimmunotherapy to
enhance cytotoxic T cell infiltration and tumor control. Nano Lett.
17:862–869. 2017. View Article : Google Scholar
|
|
112
|
Zhou S, Zhen Z, Paschall AV, Xue L, Yang
X, Bebin-Blackwell AG, Cao Z, Zhang W, Wang M, Teng Y, et al:
FAP-Targeted photodynamic therapy mediated by ferritin
nanoparticles elicits an immune response against cancer cells and
cancer associated fibroblasts. Adv Funct Mater. 31:20070172021.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Mei J, Ma J, Xu Y, Wang Y, Hu M, Ma F, Qin
Z, Xue R and Tao N: Cinnamaldehyde treatment of prostate
cancer-associated fibroblasts prevents their inhibitory effect on T
cells through toll-like receptor 4. Drug Des Devel Ther.
14:3363–3372. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Wang Y, Zeng Y, Zhu L, Wan J, Lei N, Yao
X, Duan X, Zhang Y, Cheng Y, Tao N and Qin Z: Polysaccharides from
lentinus edodes inhibits lymphangiogenesis via the toll-like
receptor 4/JNK pathway of cancer-associated fibroblasts. Front
Oncol. 10:5476832021. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Ma J, Chen X, Chen Y, Tao N and Qin Z:
Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA
Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via
TLR4. Cancers (Basel). 14:24062022. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Prossnitz ER and Barton M: The
G-protein-coupled estrogen receptor GPER in health and disease. Nat
Rev Endocrinol. 7:715–726. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lappano R, Rosano C, Pisano A, Santolla
MF, De Francesco EM, De Marco P, Dolce V, Ponassi M, Felli L, Cafeo
G, et al: A calixpyrrole derivative acts as an antagonist to GPER,
a G-protein coupled receptor: Mechanisms and models. Dis Model
Mech. 8:1237–1246. 2015.PubMed/NCBI
|
|
118
|
Maggiolini M, Santolla MF, Avino S, Aiello
F, Rosano C, Garofalo A and Grande F: Identification of two
benzopyrroloxazines acting as selective GPER antagonists in breast
cancer cells and cancer-associated fibroblasts. Future Med Chem.
7:437–448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Yang Y, Gu J, Li X, Xue C, Ba L, Gao Y,
Zhou J, Bai C, Sun Z and Zhao RC: HIF-1α promotes the migration and
invasion of cancer-associated fibroblasts by miR-210. Aging Dis.
12:1794–1807. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Du Y, Long Q, Zhang L, Shi Y, Liu X, Li X,
Guan B, Tian Y, Wang X, Li L and He D: Curcumin inhibits
cancer-associated fibroblast-driven prostate cancer invasion
through MAOA/mTOR/HIF-1α signaling. Int J Oncol. 47:2064–2072.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Kitadai Y, Sasaki T, Kuwai T, Nakamura T,
Bucana CD, Hamilton SR and Fidler IJ: Expression of activated
platelet-derived growth factor receptor in stromal cells of human
colon carcinomas is associated with metastatic potential. Int J
Cancer. 119:2567–2574. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Kinoshita K, Nakagawa K, Hamada J, Hida Y,
Tada M, Kondo S and Moriuchi T: Imatinib mesylate inhibits the
proliferation-stimulating effect of human lung cancer-associated
stromal fibroblasts on lung cancer cells. Int J Oncol. 37:869–877.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Pietras K, Pahler J, Bergers G and Hanahan
D: Functions of paracrine PDGF signaling in the proangiogenic tumor
stroma revealed by pharmacological targeting. PLoS Med. 5:e192008.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Tang D, Kang R, Berghe TV, Vandenabeele P
and Kroemer G: The molecular machinery of regulated cell death.
Cell Res. 29:347–364. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhang H, Deng T, Liu R, Ning T, Yang H,
Liu D, Zhang Q, Lin D, Ge S, Bai M, et al: CAF secreted miR-522
suppresses ferroptosis and promotes acquired chemo-resistance in
gastric cancer. Mol Cancer. 19:432020. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Qi R, Bai Y, Li K, Liu N, Xu Y, Dal E,
Wang Y, Lin R, Wang H, Liu Z, et al: Cancer-associated fibroblasts
suppress ferroptosis and induce gemcitabine resistance in
pancreatic cancer cells by secreting exosome-derived
ACSL4-targeting miRNAs. Drug Resist Updat. 68:1009602023.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Zhu Y, Fang S, Fan B, Xu K, Xu L, Wang L,
Zhu L, Chen C, Wu R, Ni J and Wang J: Cancer-associated fibroblasts
reprogram cysteine metabolism to increase tumor resistance to
ferroptosis in pancreatic cancer. Theranostics. 14:1683–1700. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Qu X, Liu B, Wang L, Liu L, Zhao W, Liu C,
Ding J, Zhao S, Xu B, Yu H, et al: Loss of cancer-associated
fibroblast-derived exosomal DACT3-AS1 promotes malignant
transformation and ferroptosis-mediated oxaliplatin resistance in
gastric cancer. Drug Resist Updat. 68:1009362023. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Li Y, Ma Z, Li W, Xu X, Shen P, Zhang SE,
Cheng B and Xia J: PDPN+ CAFs facilitate the motility of
OSCC cells by inhibiting ferroptosis via transferring exosomal
lncRNA FTX. Cell Death Dis. 14:7592023. View Article : Google Scholar
|
|
130
|
Hu S, Ma J, Su C, Chen Y, Shu Y, Qi Z,
Zhang B, Shi G, Zhang Y, Zhang Y, et al: Engineered exosome-like
nanovesicles suppress tumor growth by reprogramming tumor
microenvironment and promoting tumor ferroptosis. Acta Biomater.
135:567–581. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Yao L, Hou J, Wu X, Lu Y, Jin Z, Yu Z, Yu
B, Li J, Yang Z, Li C, et al: Cancer-associated fibroblasts impair
the cytotoxic function of NK cells in gastric cancer by inducing
ferroptosis via iron regulation. Redox Biol. 67:1029232023.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Jiang F, Jia K, Chen Y, Ji C, Chong X, Li
Z, Zhao F, Bai Y, Ge S, Gao J, et al: ANO1-Mediated Inhibition of
Cancer Ferroptosis Confers Immunotherapeutic Resistance through
Recruiting Cancer-Associated Fibroblasts. Adv Sci (Weinh).
10:e23008812023. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Pfeifer E, Burchell JM, Dazzi F, Sarker D
and Beatson R: Apoptosis in the pancreatic cancer tumor
microenvironment-the double-edged sword of cancer-associated
fibroblasts. Cells. 10:16532021. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Chen C, Liu J, Lin X, Xiang A, Ye Q, Guo
J, Rui T, Xu J and Hu S: Crosstalk between cancer-associated
fibroblasts and regulated cell death in tumors: Insights into
apoptosis, autophagy, ferroptosis, and pyroptosis. Cell Death
Discov. 10:1892024. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Li Y, Chen F, Chen J, Chan S, He Y, Liu W
and Zhang G: Disulfiram/copper induces antitumor activity against
both nasopharyngeal cancer cells and cancer-associated fibroblasts
through ROS/MAPK and ferroptosis pathways. Cancers (Basel).
12:1382020. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Lee HM, Lee E, Yeo SY, Shin S, Park HK,
Nam DH and Kim SH: Drug repurposing screening identifies bortezomib
and panobinostat as drugs targeting cancer associated fibroblasts
(CAFs) by synergistic induction of apoptosis. Invest New Drugs.
36:545–560. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Zeng Y, Du Q, Zhang Z, Ma J, Han L, Wang
Y, Yang L, Tao N and Qin Z: Curcumin promotes cancer-associated
fibroblasts apoptosis via ROS-mediated endoplasmic reticulum
stress. Arch Biochem Biophys. 694:1086132020. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Han L, Mei J, Ma J, Wang F, Gu Z, Li J,
Zhang Z, Zeng Y, Lou X, Yao X, et al: Cinnamaldehyde induces
endogenous apoptosis of the prostate cancer-associated fibroblasts
via interfering the Glutathione-associated mitochondria function.
Med Oncol. 37:912020. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Chang PM, Cheng CT, Wu RC, Chung YH,
Chiang KC, Yeh TS, Liu CY, Chen MH, Chen MH and Yeh CN:
Nab-paclitaxel is effective against intrahepatic cholangiocarcinoma
via disruption of desmoplastic stroma. Oncol Lett. 16:566–572.
2018.PubMed/NCBI
|
|
140
|
Donthireddy L, Vonteddu P, Murthy T, Kwak
T, Eraslan RN, Podojil JR, Elhofy A, Boyne MT II, Puisis JJ, Veglia
F, et al: ONP-302 Nanoparticles inhibit tumor growth by altering
tumor-associated macrophages and cancer-associated fibroblasts. J
Cancer. 13:1933–1944. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Chen B, Wang Z, Sun J, Song Q, He B, Zhang
H, Wang X, Dai W and Zhang Q: A tenascin C targeted nanoliposome
with navitoclax for specifically eradicating of cancer-associated
fibroblasts. Nanomedicine. 12:131–141. 2016. View Article : Google Scholar
|
|
142
|
Shen W, Yao PA, Li W, Gu C, Gao T, Cao Y,
Wang Z, Pei R and Xing C: Cancer-associated fibroblast-targeted
nanodrugs reshape colorectal tumor microenvironments to suppress
tumor proliferation, metastasis and improve drug penetration. J
Mater Chem B. 11:1871–1880. 2023. View Article : Google Scholar
|
|
143
|
Sousa CM, Biancur DE, Wang X, Halbrook CJ,
Sherman MH, Zhang L, Kremer D, Hwang RF, Witkiewicz AK, Ying H, et
al: Pancreatic stellate cells support tumour metabolism through
autophagic alanine secretion. Nature. 536:479–483. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Amornsupak K, Insawang T, Thuwajit P,
O-Charoenrat P, Eccles SA and Thuwajit C: Cancer-associated
fibroblasts induce high mobility group box 1 and contribute to
resistance to doxorubicin in breast cancer cells. BMC Cancer.
14:9552014. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Ren Y, Cao L, Wang L, Zheng S, Zhang Q,
Guo X, Li X, Chen M, Wu X, Furlong F, et al: Autophagic secretion
of HMGB1 from cancer-associated fibroblasts promotes metastatic
potential of non-small cell lung cancer cells via NFκB signaling.
Cell Death Dis. 12:8582021. View Article : Google Scholar
|
|
146
|
Zhao Z, Bai S, Wang R, Xiong S, Li Y, Wang
X, Chen W and Cheng B: Cancer-associated fibroblasts endow
stem-like qualities to liver cancer cells by modulating autophagy.
Cancer Manag Res. 11:5737–5744. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Martinez-Outschoorn UE, Pavlides S,
Whitaker-Menezes D, Daumer KM, Milliman JN, Chiavarina B, Migneco
G, Witkiewicz AK, Martinez-Cantarin MP, Flomenberg N, et al: Tumor
cells induce the cancer associated fibroblast phenotype via
caveolin-1 degradation: Implications for breast cancer and DCIS
therapy with autophagy inhibitors. Cell Cycle. 9:2423–2433. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Han SY, Hu MH, Qi GY, Ma CX, Wang YY, Ma
FL, Tao N and Qin ZH: Polysaccharides from polygonatum inhibit the
proliferation of prostate cancer-associated fibroblasts. Asian Pac
J Cancer Prev. 17:3829–3833. 2016.PubMed/NCBI
|
|
149
|
Ferraresi A, Phadngam S, Morani F, Galetto
A, Alabiso O, Chiorino G and Isidoro C: Resveratrol inhibits
IL-6-induced ovarian cancer cell migration through epigenetic
up-regulation of autophagy. Mol Carcinog. 56:1164–1181. 2017.
View Article : Google Scholar
|
|
150
|
Savio M, Ferraresi A, Corpina C,
Vandenberghe S, Scarlata C, Sottile V, Morini L, Garavaglia B,
Isidoro C and Stivala LA: Resveratrol and its analogue
4,4'-Dihydroxy-trans-stilbene inhibit lewis lung carcinoma growth
in vivo through apoptosis, autophagy and modulation of the tumour
microenvironment in a murine model. Biomedicines. 10:17842022.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Pereira BA, Vennin C, Papanicolaou M,
Chambers CR, Herrmann D, Morton JP, Cox TR and Timpson P: CAF
subpopulations: A new reservoir of stromal targets in pancreatic
cancer. Trends Cancer. 5:724–741. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Elyada E, Bolisetty M, Laise P, Flynn WF,
Courtois ET, Burkhart RA, Teinor JA, Belleau P, Biffi G, Lucito MS,
et al: Cross-species single-cell analysis of pancreatic ductal
adenocarcinoma reveals antigen-presenting cancer-associated
fibroblasts. Cancer Discov. 9:1102–1123. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Chen M, Cai Y, Han F, Li B, Xu Z, Cui K,
Bai W and Li F: Single-cell and bulk RNA-sequencing reveal
PRRX2-driven cancer-associated fibroblast-mediated perineural
invasion for predicting the immunotherapy outcome in colorectal
cancer. Front Cell Dev Biol. 13:16203882025. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Giorgio CD, Sette MR, Sensini B, Giannelli
E, Lachi G, Marchianò S, Paniconi F, Massa C, Urbani G, Gregorio R,
et al: LIFR antagonism reverses epithelial pro-CAF programs in
pancreatic ductal adenocarcinoma. Biochem Pharmacol.
246:1177072026. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Song J, Wei R, Liu C, Zhao Z and Liu X,
Wang Y, Liu F and Liu X: Antigen-presenting cancer associated
fibroblasts enhance antitumor immunity and predict immunotherapy
response. Nat Commun. 16:21752025. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Geng X, Chen H, Zhao L, Hu J, Yang W, Li
G, Cheng C, Zhao Z, Zhang T, Li L and Sun B: Cancer-Associated
fibroblast (CAF) heterogeneity and targeting therapy of CAFs in
pancreatic cancer. Front Cell Dev Biol. 9:6551522021. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Peng D, Fu M, Wang M, Wei Y and Wei X:
Targeting TGF-β signal transduction for fibrosis and cancer
therapy. Mol Cancer. 21:1042022. View Article : Google Scholar
|
|
158
|
Yang Y, Li X, Wang T, Guo Q, Xi T and
Zheng L: Emerging agents that target signaling pathways in cancer
stem cells. J Hematol Oncol. 13:602020. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Staff NP, Grisold A, Grisold W and
Windebank AJ: Chemotherapy-induced peripheral neuropathy: A current
review. Ann Neurol. 81:772–781. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Wang X, Zhou Y, Wang D, Wang Y, Zhou Z, Ma
X, Liu X and Dong Y: Cisplatin-induced ototoxicity: From signaling
network to therapeutic targets. Biomed Pharmacother.
157:1140452023. View Article : Google Scholar
|
|
161
|
Deac AL, Burz CC, Bocsan IC and Buzoianu
AD: Fluoropyrimidine-induced cardiotoxicity. World J Clin Oncol.
11:1008–1017. 2020. View Article : Google Scholar
|
|
162
|
Limper AH: Chemotherapy-induced lung
disease. Clin Chest Med. 25:53–64. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Zhang L, Song J, Kong L, Yuan T, Li W,
Zhang W, Hou B, Lu Y and Du G: The strategies and techniques of
drug discovery from natural products. Pharmacol Ther.
216:1076862020. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Harvey AL, Edrada-Ebel R and Quinn RJ: The
re-emergence of natural products for drug discovery in the genomics
era. Nat Rev Drug Discov. 14:111–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Huo JL, Fu WJ, Liu ZH, Lu N, Jia XQ and
Liu ZS: Research advance of natural products in tumor
immunotherapy. Front Immunol. 13:9723452022. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Pei L, Liu Y, Liu L, Gao S, Gao X, Feng Y,
Sun Z, Zhang Y and Wang C: Roles of cancer-associated fibroblasts
(CAFs) in anti-PD-1/PD-L1 immunotherapy for solid cancers. Mol
Cancer. 22:292023. View Article : Google Scholar
|
|
167
|
Yang D, Liu J, Qian H and Zhuang Q:
Cancer-associated fibroblasts: From basic science to anticancer
therapy. Exp Mol Med. 55:1322–1332. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Zhang F, Ma Y, Li D, Wei J, Chen K, Zhang
E, Liu G, Chu X, Liu X, Liu W, et al: Cancer associated fibroblasts
and metabolic reprogramming: Unraveling the intricate crosstalk in
tumor evolution. J Hematol Oncol. 17:802024. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Chen Y, McAndrews KM and Kalluri R:
Clinical and therapeutic relevance of cancer-associated
fibroblasts. Nat Rev Clin Oncol. 18:792–804. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Lavie D, Ben-Shmuel A, Erez N and
Scherz-Shouval R: Cancer-associated fibroblasts in the single-cell
era. Nat Cancer. 3:793–807. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Chen ZY, Liu HZ, Shang ZJ, Luo GF and
Zhang XZ: Nanomedicine for targeting cancer-associated fibroblasts
in cancer therapy. Theranostics. 16:1545–1576. 2026. View Article : Google Scholar :
|