Introduction to tumor related potassium
channels
Ion channels are pore-forming transmembrane
proteins, which regulate the life activities of organisms by
controlling the ion transport, not only participating in the
remodeling of cytoskeleton and the interaction between cells, but
also regulating the processes of cell migration and invasive
growth. Moreover, it has been shown that abnormal expression or
activity change of ion channels can regulate the proliferation and
apoptosis of tumor cells (1).
Voltage-gated potassium channels (Kv) are a type of
protein channels formed on the cell membrane, which can be turned
on or off according to the voltage change of the cell membrane.
They can regulate the transmembrane flow of potassium ions, thus
maintaining the normal physiological functions of cells, such as
nerve conduction, muscle contraction and cell excitability
(2). The human genome contains 40
voltage-gated potassium channels, which are involved in a number of
physiological processes, including the repolarization of neuron or
cardiac action potential, the regulation of calcium signal and cell
volume and the promotion of cell proliferation and migration. The
Kv channel has shown promising research potential in cancer,
autoimmune diseases, metabolic, neurological and cardiovascular
diseases. It has been found that several voltage-gated potassium
channels are closely associated with tumors: KCa3.1 (KCNN4)
(3), Kv10.1 (EAG1, KCNH1)
(4), Kv1.3 (KCNA3) (5) and HERG (Kv11.1, KCNH2) (6).
Among the potassium ion channels, Kv10.1 has become
a focus of current anti-cancer research due to its high expression
characteristics in tumors and close association with tumor
occurrence and development.
Kv10.1 potassium channel: An overview
Kv10.1 (potassium voltage-gated channel subfamily H
member 1/KCNH1, known as Ether-à-go-go-1 or EAG1) is a
voltage-gated potassium channel (4). Kv10.1 has been found to be expressed
in a number of different cancer cell lines, including gastric
cancer (7), liver cancer
(8) and lung cancer (9), but its expression in normal tissues
is limited (10,11). It has been proved that it is
highly expressed in a number of tumors and plays an important role,
which makes it a potential marker and target for tumor diagnosis
and treatment (7-9,12-16).
The origin of Kv10.1
The discovery of Kv10.1 channel can be traced back
to the study of voltage-gated potassium ion channels. Kv10.1 was
first discovered by Kaplan et al (2) in 1969 in the mutant X chromosome of
Drosophila. Its name 'ether-a-go-go' comes from the
irregular leg tremor phenotype exhibited by Drosophila
mutants under ether anesthesia, which is caused by a functional
defect in voltage-gated potassium channels due to gene locus
mutations (17). Studies have
shown that molecular cloning technology confirms that the protein
encoded by the Eag gene belongs to the Kv superfamily and
reveals its direct association with the regulation of neural
excitability in Drosophila (17). Kv10.1 is located in chromosome 1,
band q32.1-32.3 and comprises 457,343 bases and 12 exons (18).
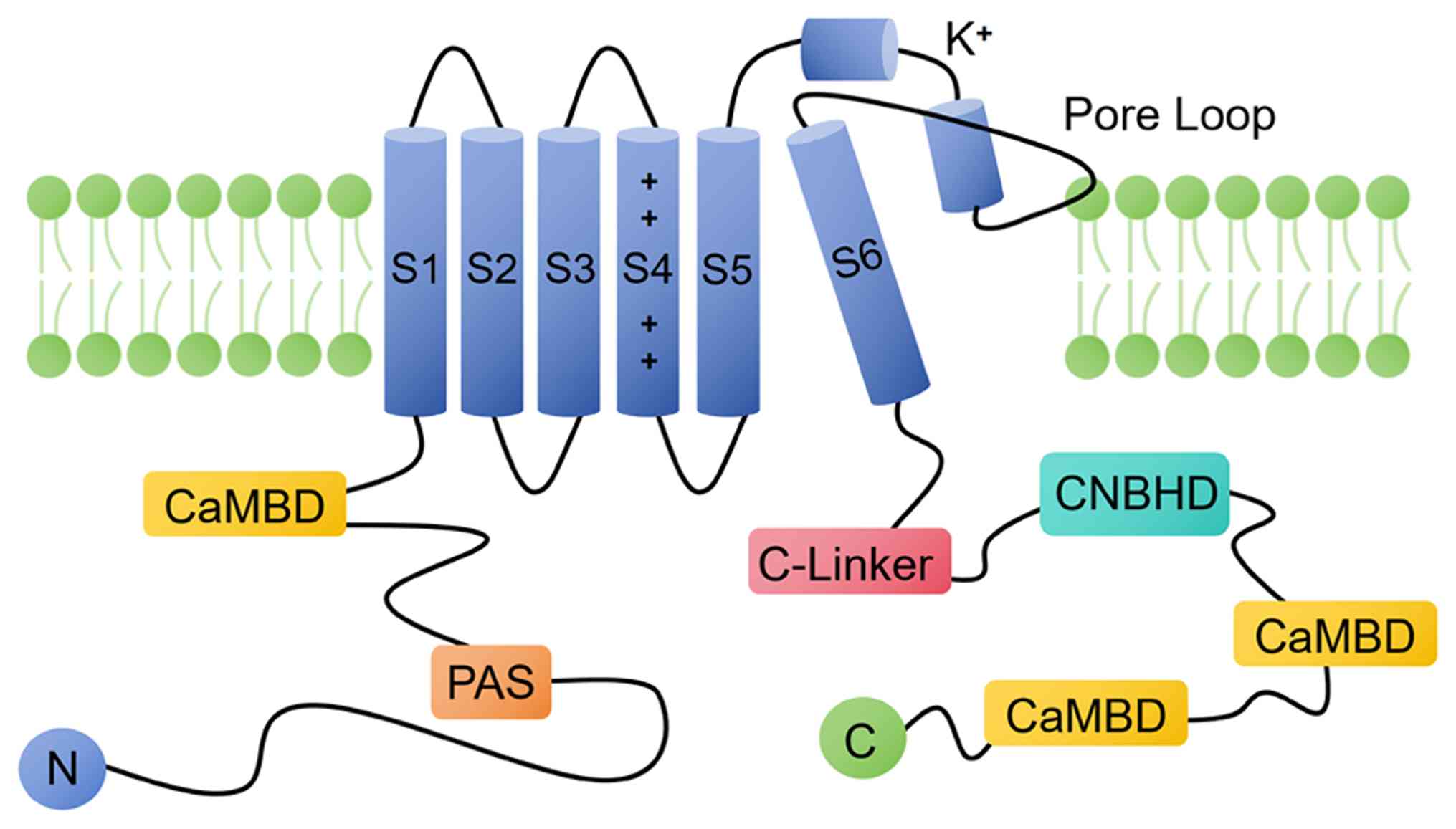
The molecular structure of Kv10.1
Kv10.1 is encoded by KCNH1 gene, which
encodes a 989 amino acid protein with an estimated molecular weight
of 111,423 Daltons. The overall structure of Kv10.1 channel is
similar to these of the other Kvs. The core region contains six
transmembrane domains (transmembrane helices S1-S6), including
voltage sensor domains (transmembrane helices S1-S4) and potassium
ion selective permeability channels (transmembrane helices S5, pore
helices S6) and the N-terminal and C-terminal of this channel
protein (Fig. 1). The single-hole
channels of these six transmembrane regions (S1-S6) are activated
by depolarization (19-21). The N-terminal contains
Per-Arnt-Sim (PAS) domain. C-terminal contains cyclic nucleotide
binding homology domain (CNBHD), C-linker, ciliary localization
signal and tetrameric coiled helix domain (19,22).
Although the structure of Kv10.1 has a number of
similarities with other Kvs, based on the three-dimensional
structure of the rat Kv10.1 obtained by single-particle
cryoelectron microscopy, it was found that Kv10.1 has a number of
unique structural features compared with other Kv channels. The
S2-S3 connector of Kv10.1 extending into cytoplasm is different
from those of other voltage-gated potassium channels. This
connector is a conservative feature of Kv family and the only other
Kv subfamily member containing the similar connectors is Kv7
(19,23). In Kv10.1, S4 region is the main
positive charge concentration region, usually with 5-7 positive
charges distributed. As a voltage receptor, this region can respond
to the electric field force when the membrane potential changes and
drive the conformational change of the channel. In addition, other
transmembrane regions (such as S1, S2 and S3) may also contain a
small amount of positively charged amino acid residues, but the
positive charge density is relatively low and the sensitivity to
voltage change is not as significant as that of S4 region. S1-S4
constitute positively charged amino acids in the field of voltage
sensors, which move membrane potential with the depolarization of
cerebral cortex (24-26). The S4-S5 linker in Kv10.1 is a
short loop composed of five amino acid residues, which contrasts
with the helical structure formed by 15 residues parallel to the
membrane in domain exchange potassium channels (such as Kv1.2)
(19,27). The segments S5-S6 form the pore
domain of the channel, while the ring P (between S5-S6) is
responsible for the K+ selectivity. These local
structural features determine that Kv10.1 has a different voltage
gating process compared with other types of Kv channels. S4 in
Kv10.1 directly interacts with the carbon terminal C-linker to
close the channel, that is, the channel can realize
voltage-dependent activation gating without covalent connection
between S4 and S5 (27,28).
Kv10.1 also has unique N-terminal and C-terminal
domains, which are necessary for subfamily-specific channel
assembly (17,29,30). Kv10.1 has a large N-terminal and a
C-terminal in cells, accounting for ~70% of its molecular weight.
In addition, there are several regulatory domains at the ends of
cells. N-terminal contains a PAS domain, which is usually involved
in the detection of redox reaction. It is considered as an oxygen
sensor, which can induce the production of hypoxia inducible factor
hypoxia-inducible factor 1 (HIF-1) in hypoxic environment, thus
causing the increase of glycolysis rate and angiogenesis, which is
beneficial to the growth of tumor cells in hypoxic environment. The
C-terminal contains a CNBHD, which is a calmodulin binding site
(31). The fragment between CNBHD
and S6 is called C-linker. S6 spirally extends to the intracellular
region and is connected to C-linker, forming an intracellular ring
above CNBHD. The C-linker couples the movements of S6 and CNBHD and
CNBHD is connected to the pore region through C-linker (32). As with other ion channels, Kv10.1
undergoes nitrogen chain glycosylation at N388 and N406 sites,
which is very important for the correct transport of the channel to
the membrane. Three-dimensional structural analysis by
cryo-electron microscopy showed that the PAS domain of Kv10.1
channel in rats was located in the intracellular region and mainly
interacted with CNBHD, a neighboring subunit. In addition, Kv10.1
channel contain amino acid residues carrying glycosylation regions
of polymers, among which glycosylation of N406 is a necessary
condition to maintain the normal structure and function of Kv10.1
and deglycosylation will lead to weakening of channel current and
slow activation (19,33).
The distribution of Kv10.1
The distribution of Kv10.1 in normal
tissues
Kv10.1 channel is a protein embedded in plasma
membrane, but it can also be found in subcellular structural
membrane including the inner nuclear membrane, intracellular
vesicles and close to the primary cilia. Under normal physiological
conditions, Kv10.1 is almost undetected in peripheral tissues,
except for a few peripheral tissues, such as pre fusion myoblasts
(34) and Kv10.1 shows abundant
expression in specific brain regions. Previous studies have shown
that Kv10.1 mainly exists in the brain, locates at the presynaptic
terminal and regulates the release of neurotransmitters, which can
be used as a regulator of local action potential (35).
The distribution of Kv10.1 in
malignant tumors and precancerous lesions
Kv10.1 potassium channel is the first
voltage-dependent potassium channel proved to be closely associated
with tumor growth (36-39). In recent years, a large number of
studies have shown that Kv10.1 potassium channels are abnormally
expressed in malignant tumors and precancerous lesions (7,40-55). Kv10.1 was highly expressed in the
primary tumor of head and neck squamous cell carcinoma and was
detected in 10 of 12 cell lines derived from head and neck squamous
cell carcinoma (40,41). Kv10.1 was positively expressed in
breast cancer cell lines, such as MCF-7 (12), T-47D (12) and MDA-MB-231 (42). Kv10.1 is highly expressed in lung
cancer. A549 and NCI-H1975 are common lung cancer cell lines
(9). In liver cancer, Kv10.1
inhibitor can inhibit the proliferation and migration of liver
cancer cells HuH-7 cells and HepG2 cells (13). By immunohistochemical methods,
>70% colon cancer tissues showed positive expression of Kv10.1,
which was markedly higher than that of normal tissues adjacent to
cancer. Kv10.1 was also highly expressed in colon cancer cell line
SW480 (14). The primary tissue
samples of gastric cancer were detected by immunohistochemistry and
it was found that 70.5% of tumor tissues showed positive expression
of Kv10.1 protein, which was markedly higher than that of normal
tissues adjacent to cancer. Compared with normal gastric mucosal
cells, the expression of Kv10.1 in gastric cancer cell lines
(SGC-7901, BGC-823) is also higher (7,43).
The expression of Kv10.1 in cervical cancer tissue was markedly
higher than that in normal cervical tissue and the expression
content of Kv10.1 in cervical cancer cells HeLa, Siha and Casci was
higher than that in normal cervical cells (15,44). Studies have shown that Kv10.1 can
be expressed in cervical intraepithelial neoplasia and the
expression level is associated with the level of cervical
intraepithelial neoplasia (16,45,46). Kv10.1 is highly expressed in
ovarian cancer and atypical adenoma hyperplasia of precancerous
lesions (16). The high
expression of Kv10.1 may be a potential prognostic marker of
prostate cancer, which is associated with the poor prognosis of
patients. Kv10.1 is highly expressed in prostate cancer tissues and
tumor cell lines, but almost not in normal prostate tissues
(47-49). The abnormally high expression of
Kv10.1 in neuroblastoma is not only driven by the transcription
level, but also depends on the fine regulation of
post-translational modifications (such as ubiquitination and
phosphorylation) (50). Kv10.1 is
also expressed in melanoma, which also affects the proliferation of
melanoma cells (51-53). Kv10.1 is highly expressed in bone
marrow primordial cells of acute myeloid leukemia patients, but
hardly expressed in normal hematopoietic stem cells and benign
blood diseases (54). The
expression level of Kv10.1 in patients with soft tissue sarcoma has
been detected by immunohistochemistry. Kv10.1 was expressed in 71%
tumors and the frequency ranged from 56% in liposarcoma to 82% in
rhabdomyosarcoma (55). Kv10.1 is
also expressed in osteosarcoma, which also affects the
proliferation of osteosarcoma cells (56). The aforementioned results are
listed in Table I. The abnormal
expression of Kv10.1 in a number of malignant tumors and
precancerous lesions will provide a certain molecular basis for the
diagnosis and treatment of corresponding diseases. Different
experimental techniques are used to analyze its mechanism of action
from multiple levels in studying the expression and function of
Kv10.1. Fresh patient samples can be quantified for protein and
mRNA expression levels through western blotting and quantitative
(q) PCR. In cell lines in good condition, channel current
characteristics can be directly measured through
electrophysiological recording, while wax blocks can be used for
immunohistochemistry and in situ localization of protein
expression. Nevertheless, each method has its limitations. Western
blotting involves semi-quantitative detection following protein
separation by electrophoresis, allowing precise measurement of
protein molecular weight and relative expression level. However, it
cannot provide protein localization information and requires
tedious experimental steps, with strict control of parameters such
as sample loading volume and antibody concentration to avoid
errors. qPCR reflects gene expression levels by detecting mRNA
levels, which is simple to operate and has high throughput.
However, it cannot directly reflect protein function and results
may be affected by factors such as RNA quality and primer
specificity. Inappropriate primer design may lead to PCR reactions
with low specificity or efficiency. Immunohistochemistry localizes
target proteins in tissue sections through antigen-antibody
reactions, visually displaying their distribution in the cell
membrane, cytoplasm, or nucleus. However, staining intensity is
easily affected by factors such as antibody concentration and
incubation time. Antibodies from different manufacturers have
varying binding abilities to target proteins, leading to
differences in results. Electrophysiological experiments directly
measure the current characteristics of ion channels to reflect
their functional activity. However, they require extremely high
cell states and have long experimental cycles, making them
difficult to apply on a large scale. In practical applications,
selection should be based on research objectives and combined with
the biological characteristics of the samples.
 | Table ICell lines with high expression of
Kv10.1. |
Table I
Cell lines with high expression of
Kv10.1.
| Cancer type | Cell line | (Refs.) |
|---|
| Head and neck
cancer | SCC29, SCC40,
SCC42B, SCC040, SCC041, SCC078, SCC094, SCC096A, SCC120,
SCC147 | (33,36) |
| Breast cancer | MCF-7, T-47D | (12) |
| MDA-MB-231 | (42) |
| Lung cancer | A549,
NCI-H1975 | (9) |
| Liver cancer | HuH-7, HepG2 | (13) |
| Colon cancer | SW480 | (14) |
| Gastric cancer | SGC-7901,
BGC-823 | (7,43) |
| Cervical
cancer | HeLa, SiHa,
CaSki | (15,46) |
| Ovarian cancer | SKOV3, OVCAR3 | (16) |
| Prostatic
cancer | DU-145, PC-3 | (47-49) |
| Neuroblastoma | SHSY-5Y | (50) |
| Melanoma | IGR1, IPC298,
IGR39, A375 | (51-53) |
| Acute Myeloid
Leukemia | HL-60, K562,
PLB-985, HEL, CMK, KASUMI, UT-7 | (54) |
|
Rhabdomyosarcoma | TE-671, A-204 | (55) |
| Fibrosarcoma | HT-1080,
Hs633t | (55) |
| Osteosarcoma | SaOS-2, MG-63 | (56) |
In addition, the ectopic expression of Kv10.1 is
associated with the phenotype of chemotherapy-resistant cells.
Therefore, the inhibition of Kv10.1 channel can improve the
response of cells to commonly used therapeutic drugs in
chemotherapy. In vitro experiments on ovarian cancer cells
showed that the expression of Kv10.1 was associated with the
chemotherapy resistance to cisplatin. Compared with cells treated
with cisplatin alone, the combination of down-regulation of Kv10.1
and cisplatin increased the apoptosis of ovarian cancer cells
(44).
Physiological and pathological functions of
Kv10.1
Physiological characteristics of
Kv10.1
Kv10.1 is a voltage-gated potassium channel, which
is highly expressed in nervous system and some cancer cells. It has
unique electrophysiological characteristics and plays an important
role in cell excitability regulation, tumor occurrence and
development.
Kv10.1 channel is voltage-dependent and its
activation degree changes with the membrane potential. This
characteristic makes Kv10.1 channel play an important role in
regulating cell excitability. In addition, the gating
characteristics of Kv10.1 channel are also different from other
potassium channels, with a slow activation and deactivation
process. The activation characteristics of Kv10.1 can be associated
with Cole-Moore shift observed in squid axons published by Cole KS
and Moore JW in 1960 (57). This
effect was originally used to describe the potassium current in
squid axons, which is characterized by a significant delay in
current activation when hyperpolarized resting membrane potential
channels are activated (58,59). The Cole-Moore shift of Kv10.1 is
~10s of milliseconds, while that of other members of the
voltage-gated potassium channel family is close to 1 msec and the
Cole-Moore shift of Kv10.1 is steeper than that of Kv10.2 (60). This electrophysiological feature
can be used as a marker to identify the expression of Kv10.1 in
cells and ectopic expression. The activation of Kv10.1 depends on
cell membrane resting potential and extracellular Mg2+
concentration and low extracellular pH can slow its activation
(61,62). Cole-Moore shift can be enhanced by
increasing extracellular Mg2+ concentration in physiological range
(63). The Cole-Moore shift of
Kv10.1 can be inhibited by using amiodarone and mibefradil
(64,65).
Kv10.1 is mainly expressed in the nervous system and
its electrophysiological characteristics are closely associated
with the function of the nervous system. In adult rats, the mRNA of
Kv10.1 can be mainly detected in olfactory bulb, cerebral cortex,
hippocampus and cerebellum and it is consistent with its protein
expression level (66). Further
research found that Kv10.1 was mostly expressed on dopaminergic
cells in physiological state, which may be associated with its
electrophysiological function (67). In the nervous system, Kv10.1 can
be used as a regulator of local action potential, especially when
other potassium channels suffer from high-frequency bursts of
cumulative inactivation. Part of the function adjustment of Kv10.1
channel involves its detergent resistant membrane part (also called
fat raft) (68,69).
The pathological function of Kv10.1 in
cancer
Kv10.1 channel is highly expressed in 70% of tumors
and its expression level is closely associated with the malignant
degree, metastatic ability and clinical prognosis of tumors. Kv10.1
is highly expressed in head and neck cancer, breast cancer and
acute myeloid cells (54,70-72) and inhibiting or knocking out
Kv10.1 can reduce tumor growth (73-76). Therefore, Kv10.1 is considered as
a potential target for cancer treatment.
Kv10.1 channel affects the production of resting
potential and action potential by regulating the potassium ion
permeability of cell membrane. This electrophysiological change may
further affect the intracellular signal transduction pathway, thus
regulating the cell proliferation process. In tumor cells, the high
expression of Kv10.1 channel may lead to uncontrolled cell
proliferation and promote tumor growth. In addition to regulating
cell proliferation, Kv10.1 channel may also participate in tumor
metastasis by regulating cell migration and invasion.
Kv10.1 is very important for the proliferation of
tumor cells. In 1997, Brüggemann et al first discovered the
function of Kv10.1 in cell proliferation in Xenopus oocytes
(77). Kv10.1 leads to abnormal
proliferation of cells, including enhanced metabolic activity and
reduced dependence of cells transfected with Kv10.1 on growth
factors in the medium. Cells transfected with Kv10.1 can continue
to grow in the medium with low concentration of serum, even without
matrix indicating the loss of contact inhibition characteristics
(17). In vitro,
inhibiting the expression of Kv10.1 can reduce the cell
proliferation, migration and invasion of cancer cells. In
vitro experiments, injecting exogenous CHO cells expressing
Kv10.1 into immunosuppressed mice induced the occurrence of
invasive tumors (78), while
using specific monoclonal antibodies to inhibit Kv10.1 can inhibit
the growth of tumors in vitro (79) and similar effects were also
observed in animal experiments (80-85). In recent years, the closed state
wild-type channel model of cryoelectron microscope structure based
on HERG and Kv10.1 channels has been constructed by using Rosetta
software and cryoelectron microscope structure. These models were
then used in molecular docking studies to explore the mechanism of
drug channel interaction (86-89).
Generally, Kv10.1 channel plays an important role in
the occurrence and development of cancer and its high expression is
closely associated with the clinical prognosis of a number of
tumors. In-depth study on the pathological function and regulation
mechanism of Kv10.1 channel is expected to provide new theoretical
basis and therapeutic strategies for cancer prevention and
treatment.
Tumor regulation mechanisms of Kv10.1
In the process of carcinogenesis, Kv10.1 may play a
role through various mechanisms. First, as a potassium ion channel,
Kv10.1 can regulate the potassium ion permeability of cell
membrane, thus affecting the generation of cell resting potential
and action potential. This electrophysiological change may further
affect the signal transduction pathway in cells and then regulate
the gene expression associated with carcinogenesis. Second, Kv10.1
may also interact with other signal pathways to jointly promote the
occurrence and development of tumors. Kv10.1 is highly expressed in
a number of tumor cells and tissues, but its specific mechanism of
action is not very clear. At present, there are only some
explorations about Kv10.1 in cell cycle regulation, cell
hyperpolarization, signaling pathway, epigenetic regulation
mechanism.
Cell cycle regulation
In 2006, it was found that the expression activity
of Kv10.1 was associated with the cell cycle (90). Cell cycle regulates the progress
of mitosis. During mitosis, if progesterone or mitotic promoter
exists, the activity of Kv10.1 channel will be inhibited, which
also proves its cell cycle sensitivity (91).
The research after 2016 gave more specific results.
That is, Kv10.1 is located in centrosome and primary cilia
(92) and promotes the
decomposition of primary cilia in G2/M phase, which is
beneficial to the progress of cell cycle (93). Following Kv10.1 knock-out, ciliary
decomposition is inhibited and proliferation is delayed. Therefore,
the regulation of Kv10.1 on ciliary development can explain the
influence of Kv10.1 expression on normal cell proliferation and may
be a main mechanism of its tumorigenesis (94). Kv10.1 affects Ca2+
entry through hyperpolarization and Ca2+ inhibits EAG1
function through calmodulin (3).
Ca2+ is associated with the cell cycle and cell
replication. Calcium influx is considered necessary for the
transition from G1 phase to S phase in the cell cycle.
This hyperpolarization also facilitates the entry of a large amount
of nutrients into the cell, thereby promoting cell proliferation
(10,95).
Similar research also support the aforementioned
view. Urrego et al (96)
reported that Kv10.1 could be detected only in G2/M
phase. As the E2F transcription factor 1 (E2F1) coordinates cell
division and induces the expression of Kv10.1 in G2
phase, Kv10.1 can only be detected in G2/M phase and
downregulation of Kv10.1 will prolong the duration of
G2/M phase. However, Kv10.1 will be diluted in
G0/G1 phase, so it cannot be detected in
other stages of cell cycle (97).
Some studies also hypothesize that the strong influence of Kv10.1
channel in cell cycle progress is also associated with ciliary
disintegration, which usually provides the signal transduction
needed to start proliferation (96,97). Cyclic expression of Kv10.1 has
been proved to exist at the base of primary cilia of hTERRPE-1
cells, which may lead to significant disposability of microtubules
in the subsequent process (10).
Regarding microtubule activity, the upregulation of
Kv10.1 is closely associated with a significant increase in
microtubule dynamics, characterized by assembly and disassembly
rates. Therefore, there is an important association between Kv10.1
overexpression and certain processes including cellular variability
(96,97). It has been proved that the
mobility of MDA-MB-231 cells decreased markedly when Kv10.1 was
exposed to a specific blocking agent such as astemizole. Some
hypotheses suggest that cell migration caused by Kv10.1 channel is
mainly due to the change of microtubule dynamics caused by
hyperpolarization of action potential, which has been proved to
induce calcium to enter through Ca2+ release and
activates ORAI calcium release-activated calcium modulator
1(ORAI1). ORAI1 association with Ca2+ ATPase seems to be
associated with the proliferation and survival of cancer cells
(98).
Hyperpolarization mechanism
The outflow of K+ through Kv10.1 channel
makes cells hyperpolarized, thus promoting the influx of
Ca2+, leading to the proliferation of non-invasive cells
(10,99,100). However, the relationship between
Kv10.1 and intracellular Ca2+ is complicated. Although
Kv10.1 is beneficial to Ca2+ influx under
hyperpolarization, high intracellular Ca2+ reversibly
inhibits the overexpression of Kv10.1 in a number of tumors
(101,102). Overexpression of Kv10.1 can lead
to membrane potential hyperpolarization and affect cell cycle
progression. Potential changes in the cell cycle are much slower,
smoother and less pronounced than rapid action potentials. Due to
membrane hyperpolarization, the driving force for calcium influx is
enhanced. Ca2+ is a major second messenger involved in
cell proliferation, migration, survival and apoptosis (103). Depletion of extracellular
calcium ions can arrest cells in the early G1 and
G1/S phase transition. Normal cell proliferation relies
on intracellular and extracellular calcium ions, especially during
the G1 phase. However, cancer cells can bypass this
requirement and continue to proliferate in calcium-deficient media
(104,105). This mechanism exhibits high
similarity across various types of cancer. Calcineurin is one of
the main regulators of intracellular Ca2+ signaling.
Calcineurin inhibits the degradation of cyclin D1 through
dephosphorylation of T286 residue, promoting cell cycle progression
(106). Ca2+ can also
form a dynamic interaction network with the Hippo pathway, jointly
regulating cell proliferation, differentiation and tissue
homeostasis (107). Although
there are currently no specific studies directly elucidating the
role of Kv10.1 in the interaction with Ca2+ regulation
of cyclin D1 and the Hippo pathway, based on the existing
understanding of the complex regulatory relationship between
Ca2+ and cyclin D1 and the Hippo pathway, it is
reasonable to speculate that Kv10.1 may play a role in this
regulatory network. Cell hyperpolarization is very sensitive to the
activation of Kv10.1, which can regulate cell hyperpolarization
through the interaction between N-terminal of PAS domain and S4-S5
connector (28).
When the cell membrane is hyperpolarized, the Kv10.1
channel can sense this potential change and open, allowing
potassium ions to flow from the inside of the cell to the outside
(17). Therefore, drug
development for Kv10.1 channel has become a potential cancer
treatment strategy, aiming at inhibiting the growth and spread of
tumor by blocking its hyperpolarization mechanism.
Epigenetic regulation mechanism
Epigenetic regulation mechanism is a mechanism that
changes biological phenotype without involving DNA sequence
changes, but including DNA methylation, histone modification and
non-coding RNA-mediated regulation. These modifications can affect
the transcription activity of genes, thus regulating gene
expression (108). For example,
the DNA methylation status in the promoter region of Kv10.1 gene
may affect its transcription activity, thus regulating the
expression level of Kv10.1. In addition, histone modification may
also regulate the transcription process of Kv10.1 gene by affecting
the position and arrangement of nucleosomes. Studies have shown
that several abnormally methylated genes, including Kv10.1, have
been found in gastric cancer tissue and the methylation level of
Kv10.1 in the samples is more than three times higher, suggesting
that the hypermethylation of Kv10.1 may play a role in the
occurrence and development of gastric cancer (17,35). Some have suggested that epigenetic
changes in early life may alter lung cell function and lead to
asthma risk (109). In mouse
models, early exposure to house dust mite allergens alters DNA
methylation and the expression of different genes detected, up to
three consecutive generations and is associated with airway
hyperresponsiveness and inflammation (109). In mice exposed to allergens, the
Kcnh1 gene was hydroxymethylated and upregulated, indicating a
susceptibility to asthma (109).
In head and neck squamous cell carcinoma (HNSCC), histone
acetylation, rather than DNA methylation, is hypothesized to be
involved in the regulation of Kv10.1 (40).
Reversal of drug resistance
Simultaneous drug resistance to multiple drugs with
different chemical structures and targets is the main obstacle to
effective cancer treatment (110-114). Multidrug resistance (MDR) is an
acquired resistance of microorganisms and tumor cells to
chemotherapeutic drugs, which is characterized by different
chemical structures and mechanisms of action. MDR is the result of
overexpression of a number of protein and these protein drugs
squeeze chemotherapy drugs out of cells to make their concentration
lower than the effective concentration (115). MDR in cancer treatment causes
tens of thousands of deaths every year and it can be endowed by a
number of transporters that pump drugs out of cells. They can
transport various substrates, including amino acids, peptides,
ions, sugars, toxins, lipids and drugs and are associated with
several serious human diseases (116). First-line therapy is usually
followed by the proliferation of a small number of surviving cancer
cells, which leads to the development of secondary tumors, which
are insensitive to initial drugs. This may lead to tumor
progression after stabilization or significant regression, because
successful chemotherapy in the first stage becomes ineffective
(117,118).
Platinum drugs (such as cisplatin and carboplatin)
are one of the commonly used chemotherapy drugs in clinic and are
widely used to treat various tumors. However, the resistance of
tumor cells to platinum drugs limits their clinical application. As
aforementioned, the ectopic expression of Kv10.1 is not only
associated with the proliferation of tumor cells, but also
associated with the phenotype of chemotherapy-resistant cells.
In vitro experiments in ovarian cancer cells showed that the
expression of Kv10.1 was associated with the chemotherapy
resistance of cisplatin. Compared with the cells treated with
cisplatin alone, the downregulation of Kv10.1 combined with
cisplatin increased the apoptosis of ovarian cancer cells (16). Similar results were also observed
in the chemotherapy-resistant glioblastoma cell line U251AR, which
showed a high level of Kv10.1 (mRNA and protein). Notably, when the
expression of Kv10.1 is downregulated, U251AR cells are more
sensitive to chemotherapy drugs (119). Therefore, the inhibition of
Kv10.1 channel can improve the response of cells to commonly used
therapeutic drugs in chemotherapy.
Other mechanisms
The tumorigenic mechanism of Kv10.1 is associated
with the fact that calcium-dependent calmodulin kinase-Ⅱ is always
activated, which can cause imbalance of cell proliferation and
apoptosis even at low calcium ion concentration (120). Kv10.1 also promotes tumor
progression by increasing the activity of hypoxia sensor
hypoxia-inducible factor-1 alpha (HIF-1α), stimulating the
secretion of vascular endothelial growth factor (VEGF) and
increasing angiogenesis under hypoxia (78). The tumor microenvironment usually
shows changes in extracellular matrix (ECM). ECM products, such as
type 1 collagen and fibronectin, promote tumor progression by
increasing cell movement (121).
Ion channels are highly fine-tuned protein necessary
for cell physiology. Therefore, the synthesis and degradation of
ion channels should be a quality control process, which is
necessary for correct cell function. However, the knowledge about
the synthesis and degradation of Kv10.1 is limited. A study has
shown that protein degradation of Kv10.1 depends on the protein
ligase cullin 7 (Cul 7) (122).
Cul7 degrades Kv10.1 protein on plasma membrane through proteasome
and lysosomal pathways respectively (122).
Signaling pathway
In the field of tumor biology, abnormal regulation
of cell signaling pathway is recognized as the core mechanism
driving tumor occurrence, progress and metastasis. In recent years,
because of its unique molecular structure, specific expression in
tumor tissues and its regulation on cell proliferation, migration
and invasion, Kv10.1 has become a typical example in the study of
connecting ion channels with tumor signal pathways (Fig. 2).
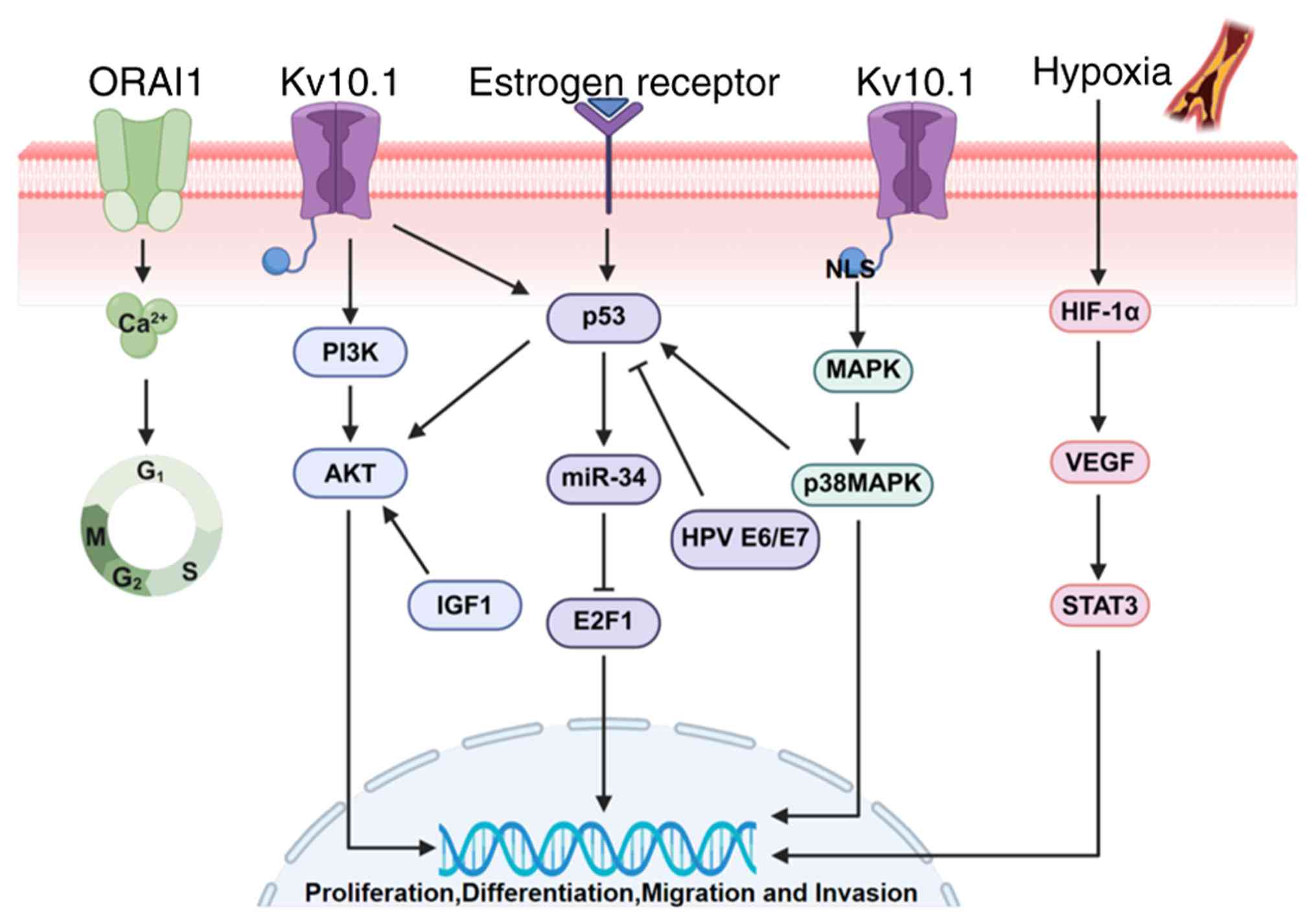 | Figure 2Kv10.1 activates multiple signaling
pathways. Kv10.1 has become a typical example of the connection
between ion channels and tumor signaling pathways due to its unique
molecular structure, specific expression in tumor tissues and
regulation of cell proliferation, migration and invasion. Kv10.1 is
negatively regulated by the negative feedback mechanism of
p53-miR-34-E2F1 pathway. P53 is regulated by a number of factors,
such as estrogen receptor, HPV and E6/E7. Kv10.1 can upregulate the
expression of HIF-1α by interacting with the intracellular hypoxia
homeostasis system and promote the transcription of VEGF under
hypoxia. In osteosarcoma cells, Kv10.1 regulates the expression of
STAT3-VEGF by affecting the activation of stat3-vegf downstream.
Kv10.1 channel and PI3K/AKT signaling pathway form cancer promoting
signal axis through functional correlation and regulate cancer cell
proliferation, invasion and metastasis, angiogenesis and glucose
metabolism. The NLS in the C-terminal domain of Kv10.1 activates
the MAPK signaling pathway. For example, the high expression of
Kv10.1 in osteosarcoma cells is regulated by p38MAPK/p53 pathway.
The increase of ORAI1 in plasma membrane can regulate intracellular
calcium ion and mediate cell cycle. miR, microRNA; E2F1, E2F
transcription factor 1; HPV, human papillomavirus; HIF-1α,
hypoxia-inducible factor-1 alpha; VEGF. vascular endothelial growth
factor; STAT3, signal transducer and activator of transcription 3;
NLS, nuclear localization signal; MAPK, mitogen-activated protein
kinase; ORAI1, ORAI calcium release-activated calcium modulator
1. |
p53-miR34-E2F1-hEAG1 signaling
pathway
Studies have shown that the tumor suppressor gene
regulatory network includes the p53-miR-34 pathway and E2F1
transcription factor, which can regulate the expression of Kv10.1
(96,123). The expression of Kv10.1 may be
regulated by the negative feedback of this signal pathway (124,125). This negative regulation may be
achieved through two mechanisms: direct inhibition of Kv10.1 at the
post-transcriptional level and negative regulation of Kv10.1
through negative feedback mechanism through the p53-miR-34-E2F1
pathway. Studies have also shown that estrogen and human
papillomavirus (HPV) regulate the level of Kv10.1 protein (126). As estrogen receptor interacts
with p53 (125), HPV inhibits
p53 and activates E2F1 (127),
suggesting that EAG1 may affect downstream gene transcription by
regulating E2F1. In addition, insulin-like growth factor 1 can also
increase the expression of EAG1 by activating kinase AKT1 (128). Studies have shown that AKT1 can
overcome the apoptosis-inducing effect of p53 (129), suggesting that p53 and Kv10.1
can act in opposite directions along the axis of apoptosis and
proliferation (130).
HIF-1α/signal transducer and activator of
transcription 3 (STAT3)/VEGF signaling pathway
The influence mechanism of silent Kv10.1 on
HIF-1α/STAT3/VEGF pathway is a multi-level and multi-step process.
As a potassium channel, the abnormal expression of Kv10.1 may
regulate the activity of intracellular signal transduction pathway
by affecting the balance of potassium concentration inside and
outside the cell. Kv10.1 can upregulate the expression of HIF-1α by
interacting with intracellular hypoxia homeostasis system. HIF-1α
is a key transcription factor to regulate the expression of VEGF,
which promotes the transcription of VEGF under hypoxia, thus
inducing angiogenesis (131). In
osteosarcoma cells, the phosphorylation level of STAT3 decreased
markedly after Kv10.1 silencing, which indicated that Kv10.1 might
regulate the expression of STAT3-VEGF downstream by influencing its
activation state (84,132).
PI3K/AKT signaling pathway
The PI3K/AKT signaling pathway is an important
signal transduction pathway with multiple biological functions
mediated by enzyme-linked receptors in mammals. The PI3K/AKT
pathway plays an important role in the occurrence and development
of tumors. When stimulated by upstream signals, PI3K activates AKT,
which further activates downstream signal molecules and regulates
cancer cell proliferation, invasion and metastasis, angiogenesis
and carbohydrate metabolism. In osteosarcoma, tumor growth and
angiogenesis in osteosarcoma can be inhibited by downregulating
VEGF/PI3K/AKT signaling pathway to silence Kv10.1 (133). The Kv10.1 channel and PI3K/AKT
signaling pathway form a cancer-promoting signal axis through
functional correlation and Kv10.1 is the key target of Nutlin-3
regulating PI3K/AKT through p53 (81). Therefore, it is of great
significance to study PI3K/AKT signaling pathway and develop
targeted drugs for tumor treatment.
The mitogen-activated protein kinase
(MAPK) signaling pathway
The nuclear localization signal at the C-terminal
of Kv10.1 potassium channel can activate the MAPK pathway to cause
changes in cell morphology (134). The Kv10.1 channel plays the role
of oncogene, such as promoting the proliferation of human
osteosarcoma cells. The high expression of Kv10.1 in osteosarcoma
cells is regulated by p38MAPK/p53 pathway (135). Inhibition of p38MAPK activation
by p38 MAPK inhibitor SB203580 or short interfering RNA also
decreased the level of Kv10.1 protein, but increased the level of
p53 protein. In addition, the activation of p53 will lead to the
growth stagnation of osteosarcoma cells and decrease the level of
Kv10.1 protein, while the inactivation of p53 will promote the cell
growth and increase the expression of Kv10.1 protein. This suggests
that Kv10.1 may affect the proliferation of osteosarcoma cells by
regulating p53 (136).
Cancer type specificities in the
anti-tumor mechanisms of Kv10.1
Although Kv10.1 exhibits mechanistic similarities
across various malignancies, namely, influencing tumor
proliferation through hyperpolarization, exerting effects via the
PI3K/AKT signaling pathway and inhibiting apoptosis (81), its downstream signaling network
still demonstrates significant cancer type specificity. For
instance, the expression of Kv10.1 in endometrial cancer tissues is
markedly higher than that in normal endometrial tissues. Kv10.1 can
be regulated by cyclin D1, p53 and other factors, thereby affecting
the proliferation, migration and invasion of tumor cells (137). Kv10.1 may form different
complexes or interaction networks with other molecules in various
cancers, thereby influencing its function. For example, in
osteosarcoma and hepatocellular carcinoma, Kv10.1 enhances tumor
angiogenesis by upregulating the expression of hypoxia-inducible
factor HIF-1α and promoting the secretion of VEGF. This effect is
not dependent on its ion transport function but is achieved through
interaction with the cellular hypoxia homeostasis system (133,138). Different mechanisms predominate
in various cancers; for instance, in breast cancer, the PI3K/AKT
pathway is central, driving cell proliferation and metabolic
reprogramming; in hepatocellular carcinoma, the HIF-1α/VEGF pathway
is predominant, promoting angiogenesis and tumor progression; in
endometrial cancer and head and neck cancer, it may affect tumor
behavior through cell cycle regulation and cytoskeleton dynamics
(137-139). Drug targets are critical in drug
discovery and therapy, as their identification is essential for
achieving precision treatment (140). Therefore, Kv10.1 is considered a
potential therapeutic target in various cancers.
Regulators and inhibitors
According to the activation mechanism of Kv10.1,
its regulators are mainly divided into two categories: activators
and inhibitors. Validation of Kv10.1 as a therapeutic target
requires specific modulators and can be used as a basic tool for
understanding channel pharmacology. Therefore, it is necessary to
find more selective and effective modulators, especially
inhibitors. According to the structure of Kv10.1, this section
introduces its regulators and inhibitors and their applications in
cancer (Table II).
| Table IIKv10.1 inhibitors. |
Table II
Kv10.1 inhibitors.
| Compound name | Binding domain | Cell line |
IC50 | (Refs.) |
|---|
| Chlorpromazine | PAS | Xenopus
oocytes | 3.7±0.7
µM | (143) |
| Mibefradil | VSD | 293 | Kd 1.3 µM,
nH0.8 | (65,145) |
| Purpurealidin
analog 5 | VSD | Xenopus
oocytes | 7.7±1.0
µM | (50) |
| 20(S)-ginsenoside
Rg3 | VSD | Xenopus
oocytes | 1.18 µM | (1547,148) |
| APETx4 | VSD | Xenopus
oocytes | 1.1µM | (149) |
| Corydaline | VSD | HepG2 | 11.3±0.6
µM | (150) |
| Tetrandrine | Selectivity
filter | CHO | 70±5.2
µM | (155) |
| ProcyanidinB1 | Selectivity
filter | 293 | 10.38±0.87
µM | (13) |
|
Tetraethylammonium | Pore domain | Xenopus
oocytes | 1.2±0.1
µM | (156) |
| Amiodarone | Pore domain | 293 | Kd 203 nM,
nH0.9 | (157) |
| Astemizole | Pore domain | 293
Xenopus oocytes | 196 nM
2.8±0.1 µM | (157) |
| Imipramine | Pore domain | Xenopus
oocytes | 40.2±0.3
µM | (157) |
| Dronedarone | Pore domain | 293 | Kd 9 µM,
nH0.9 | (76) |
| Quinidine | Pore domain | CHO | 1.4±0.1
µM | (60) |
| Haloperidol | Pore domain | CHO | 590±121 nM | (67) |
| κ-Hefutoxin 1 | Pore domain | - | 26±2 µM | (158) |
| Heme | C-linker | Xenopus
oocytes | 4nM | (161) |
PAS domain
The Kv10.1 channel is formed by assembling tetramer
subunits and each subunit contains N-terminal PAS domain and
C-terminal cyclic nucleotide binding homology domain. Small
molecular ligands can inhibit Kv10.1 channels by binding to their
PAS domains. Deleting PAS domain can cancel the inhibitory effect
of chlorpromazine (141). At low
voltage, chlorpromazine can inhibit mouse Eag1 by binding with PAS
domain, but when the channel is open at high voltage, it will be
inhibited by blocking the channel (142). Chlorpromazine, a small molecule
binding agent of PAS domain, changes the interaction between PAS
and CNBH domain and reduces the coupling between tetramer ring and
channel hole in cells, while the coupling between PAS and voltage
sensor domain (VSD) domain has little effect. In addition, the
combination of chlorpromazine and PAS domain does not change the
Cole-Moore shift characteristics of Kv10.1 channel, which further
indicates that chlorpromazine has no effect on the movement of VSD
from deep closed state to open state (143). Chlorpromazine inhibits the
proliferation of oral cancer cells by regulating the PI3K/AKT/mTOR
signaling pathway and effectively suppresses tumor growth in
zebrafish and mouse models. Given that chlorpromazine can act on
Kv10.1, it is hypothesized that it regulates tumor proliferation by
modulating the PI3K/AKT signaling pathway through Kv10.1. Based on
the mechanism of action of chlorpromazine on oral cancer cells and
its potential association with Kv10.1, it is reasonable to
hypothesize that chlorpromazine may regulate the PI3K/AKT signaling
pathway through Kv10.1, thereby affecting tumor proliferation
(144).
VSD
VSD has a small molecule binding site as a gating
regulator and extracellular VSD allows molecules to regulate
channels without transmembrane, as with toxins. Most of the known
small molecules that regulate Kv10.1 activity first cross the cell
membrane and bind to the inner side of the channel. Most of these
compounds are physically closed pore blocker ion permeation
pathways, such as mibefradil (65,145), Purpurealidin analog 5 (146), 20 (s)-Ginsenoside Rg3 (147,148), APETx4 (149) and Corydaline (150).
Mibefradil has been reported to alter the gating of
Kv10.1 by binding to the VSD (65,145). When the hyperpolarizing
potential acts outside the cell, mibefradil induces obvious open
inactivation, but not from inside the cell. In addition, mibefradil
also inhibited the Cole-Moore shift. It is also not used as a pore
size blocker because it does not compete with the known pore size
blocker, quinidine. Mibefradil 1 also inhibits other potassium
channels and L-type and T-type calcium channels (65).
Purpurealidin is considered to be a gating
modifier, which binds to the mibefradil binding site near the
voltage sensor Kv10.1. Mibefradil is a Ca2+ channel
antagonist and a Kv10.1 door control regulator. As with
purpurealin, mibefradil moves the activation curve to the left.
Studies have shown that the binding site of purpurealidin on Kv10.1
overlaps with that of mibefradil (65,145,146).
Ginsenoside is a steroidal glycoside that inhibits
the KCNH family at sub µM concentrations. Its function is
not limited to Kv channels. Studies have shown that ginsenosides
can markedly inhibit the voltage dependent activation curves of
Kv10.1 and HERG channels and increase their opening probability at
more negative potentials (147,148). When combined with cisplatin, it
can enhance chemotherapy sensitivity and reverse drug resistance by
blocking the PI3K/AKT pathway (151).
Apetx4 is a new toxin isolated from sea anemones,
which can inhibit the Kv10.1 channel. APETx4 inhibits its current
by binding to Kv10.1 channel and the inhibitory effect is
concentration dependent. APETx4 is a gating modifier that
presumably binds to the S3b-E2-S4 region (voltage sensor paddle) of
Kv10.1 (149). Corydaline is a
novel natural product that selectively inhibits Kv10.1 channels,
while being insensitive to other KCNH channels. Corydaline can
inhibit the proliferation and migration of liver cancer cells by
targeting Kv10.1. This previously unidentified new site can
specifically bind to the medicinal pocket of Corydaline, providing
possibilities for drug screening against diseases associated with
abnormal Kv10.1 channels (150).
Selectivity filter
The selectivity filter of Kv10.1 consists of a
highly conserved amino acid sequence, forming a structure similar
to the 'potassium channel signature sequence' (152). Through the spatial arrangement
of carbonyl oxygen atoms and threonine hydroxyl oxygen atoms, this
region mimics the hydration layer mechanism of water molecules
surrounding K+, forming a series of K+
binding sites (153,154).
Tetrandrine, a natural compound used in traditional
Chinese medicine, can inhibit Kv10.1 in a concentration dependent
manner with an IC50 of 69.97±5.2 µM (155). Tetrandrine specifically
inhibited Kv10.1 channel, blocked potassium outflow, induced cell
cycle arrest in G1/S phase and enhanced cisplatin
sensitivity by downregulating PI3K/AKT signaling pathway (44). Tetrandrine may also have the same
binding site as procyanidin B1, a natural compound present in grape
seeds, which inhibits Kv10.1 in a concentration dependent manner
with an IC50 value of 10.4±0.9 µM (13). Procyanidin B1 directly inhibits
channel activity and blocks tumor cell proliferation by hydrogen
bonding to i550, t552 and q557 amino acids in the filter. Similar
to tetrandrine, 100 µM procyanidin B1 did not markedly
inhibit Kv7.1, Kir2.1, or HERG (13).
Pore domain
The pore region of Kv10.1 is composed of S5 and S6
transmembrane helices and intermediate pore rings, which is the
core domain of ion selective permeation. In recent years, in the
development of targeted drugs for the Kv10.1 channel, the pore
domain has become the core target in the design of small molecular
inhibitors because of its unique structural characteristics and
functional importance. Tetraethylammonium is a commonly used
potassium channel blocker, which can physically block ion passage
(156). Tetraethylammonium can
combine with aromatic amino acid residues on the inner wall of the
channel, which hinders the selective filtration function of
potassium ions, thus inhibiting the channel current. The mechanism
of quinidine and tetraethylammonium is similar and does not depend
on the voltage gating state of the channel, so it belongs to
non-competitive inhibition (60).
The blocking effect of quinidine on Kv10.1 channel is
concentration-dependent and the inhibitory effect is more obvious
at high concentration. Astemizole competitively binds to Kv10.1
channel with tricyclic antidepressants imipramine and
tetraethylammonium (157). The
blocking effect of imipramine is voltage-dependent and can be
antagonized by intracellular tetraethylammonium. Amiodarone and
dronedarone both act through the pore region, but their blocking
effects on Kv10.1 are different and dronedarone cannot inhibit the
Cole-Molar displacement characteristics of Kv10.1 channel (76). Haloperidol also inhibited the
Kv10.1 channel through the pore region and the inhibition was
associated with the drug concentration and membrane potential
(67). κ-Hefutuxin 1, as the
first peptide inhibitor of Kv10.1 channel, inhibits its activity by
binding to Kv10.1 channel (158). As the first peptide inhibitor of
Kv10.1 channel, κ-hefutuxin 1 inhibits its activity by binding to
Kv10.1 channel and may act on or near the pore area of Kv10.1
channel. Channel residues met397 and asp398 may be the anchors to
stabilize its binding (158).
Studies have investigated the effects of
Haloperidol on the PI3K/AKT signaling pathway in PC12 cells (a
neuroblastoma cell line) (159).
The research found that Haloperidol can induce the nuclear
translocation of PI3K, generating
phosphatidylinositol-3,4,5-trisphosphate (PIP3) in the nucleus,
effectively inhibiting the phosphorylation of AKT, leading to a
decrease in AKT activity. Furthermore, Haloperidol can effectively
suppress the expression of Kv10.1 (159,160). Although no studies have yet
explored the direct relationship between these factors, it is
possible that Haloperidol exerts its anticancer effects by
indirectly influencing the PI3K/AKT signaling pathway through the
inhibition of Kv10.1 expression (159).
Other binding sites
At present, the mechanism of action of Kv10.1
inhibitors is complex and diverse and the binding sites of some
inhibitors are not yet clear. Heme is an endogenous regulator of
the Kv10.1 channel, which inhibits its activity by binding to the
CxHxH motif in the C-Linker region (161). The aforementioned results are
listed in Table II.
Inhibitors with clear binding sites are beneficial
for the development of new drugs for treating diseases, while
inhibitors with unknown binding sites reveal the complexity and
diversity of channel regulation. Future research should further
strengthen the exploration of the mechanism of action of inhibitors
with unknown binding sites and reveal the molecular mysteries of
their interaction with Kv10.1 channels. Meanwhile, based on the
mechanism of action of inhibitors, more efficient and specific
Kv10.1 inhibitors have been developed, providing new strategies and
means for the treatment of related diseases.
Clinical status
From a clinical perspective, Kv10.1 is highly
expressed in various malignant tumor tissues, markedly higher than
in normal tissues, thus possessing the potential to become a
diagnostic marker for tumors. Currently, little is known about the
mechanism of Kv10.1 in tumorigenesis and most data on Kv10.1
inhibitors are derived from in vitro cell lines or simple
animal models. This lack of accurate targeting in drug development
makes it difficult to design highly effective and specific targeted
drugs. The metabolic process of drugs in the body, potential toxic
effects and how to effectively deliver drugs to tumor tissues have
become major obstacles in the clinical translation of Kv10.1
targeted drugs.
The clinical value of Kv10.1 as a
diagnostic marker
Kv10.1 is expressed in a variety of malignant
tumors, including clear cell renal cell carcinoma, breast cancer,
cervical cancer, gastric cancer, colon cancer and cervical
intraepithelial neoplasia (14,42,43,162-165). According to clinical data
analysis reported in relevant literature, a total of 68 clinical
samples were tested in the study of esophageal squamous cell
carcinoma. The results showed that 51 cases showed positive
expression, with a positive rate of 75%. Further analysis showed
that the positive expression was closely associated with the depth
of tumor infiltration and the survival time of positive patients
was generally shorter (166). In
clinical data of ovarian cancer, there were 336 samples, of which
249 showed positive results, with a positivity rate of 65%. The
expression of Kv10.1 is associated with tumor size, differentiation
degree, staging and metastasis (167). In the clinical data of head and
neck squamous cell carcinoma, there were a total of 54 clinical
samples, of which 45 samples showed positive results, with a
positive rate as high as 83.00%. It is worth noting that Kv10.1
plays a key role in this head and neck squamous cell carcinoma and
has been proven to be a tumor marker and potential therapeutic
target. The expression of Kv10.1 is associated with important
clinical pathological features such as tumor size, differentiation
degree, staging and metastasis (40).
A research group has conducted in-depth predictive
analysis of the clinical correlation between cancer-related genes
and renal cell carcinoma cell lines using the Cancer Genome Atlas
database. The research results indicate that Kv10.1 is a key
biomarker and potential therapeutic target in clear cell renal cell
carcinoma (ccRCC) (162). The
widespread abnormal expression of Kv10.1 in numerous tumors
suggests that it may be intrinsically associated with the
occurrence and development of tumors and has potential value as a
tumor marker for tumor screening and diagnosis. However, most of
the data on Kv10.1 inhibitors to date have been derived from in
vitro cell lines or simple animal models and there is no
systematic patient-derived xenografts (PDX) model data to clearly
demonstrate the quantitative indicators of tumor inhibition rate
and survival extension effect of representative inhibitors. Due to
the lack of such data, it is difficult to directly support the
conclusion that Kv10.1 inhibitors have significant clinical
translational value in PDX models. Kv10.1 is highly expressed in
brain tissue and involved in regulating neuronal excitability,
neurotransmitter release and synaptic plasticity. When treating
brain tumors, Kv10.1 inhibitors need to penetrate the blood-brain
barrier (BBB) to target tumor cells. At this time, increased
permeability of the BBB (such as through hyperosmotic solutions or
nanocarrier technology) may facilitate drug entry into the brain,
but it also increases the risk of exposure to normal brain tissue.
When treating non-brain tumors, the BBB can prevent Kv10.1
inhibitors from entering the brain tissue, thereby avoiding
neurotoxicity. Studies have shown that knocking out Kv10.1 did not
result in significant abnormalities in multiple behavioral tests in
mice and its function may have a redundant compensation mechanism
with other channels. Therefore, Kv10.1 inhibitors have less
interference with normal tissue, especially brain tissue function
(67). Therefore, at present, it
can only be concluded that Kv10.1 is a potential therapeutic
target.
Bottleneck of clinical transformation of
targeted drugs
The BBB is a selective barrier between the central
nervous system and the circulatory system. Its unique structural
characteristics, such as tight junction, stability of cytoskeleton
and active efflux of transmembrane proteins, together constitute
the physical and chemical barrier for drugs to enter the brain
tissue. Kv10.1 is highly expressed in brain tumors (such as
glioma), but the existence of the blood-brain barrier makes it
difficult for most drugs to penetrate effectively, resulting in low
treatment efficiency of brain tumors. Even if drugs can break
through the blood-brain barrier for a short time, its action time
is often not enough to play a lasting effect (168,169).
The molecular weight of peptide inhibitors is
relatively small and the structure is relatively simple. It is
generally considered to have low immunogenicity potential and it is
easy to be degraded and excreted in the body, reducing the contact
time with the immune system, thus reducing the immunogenicity. In
addition, in the production process of peptide inhibitors, some
impurities may be introduced, such as pollutants, solvents,
enzymes, impurities formed by amino acid side chain modification,
truncation, duplication, oxidation, insertion or deletion in the
active pharmaceutical ingredient sequence. These impurities may
have potential immunogenicity and can trigger the immune response
of the body, thus affecting the safety and effectiveness of
drugs.
Kv10.1 combination therapy reverses drug
resistance
Kv10.1 inhibitors combined with chemotherapeutic
drugs can act on tumor cells simultaneously through different
mechanisms to produce synergistic killing effect. This combined
strategy is expected to improve the therapeutic effect and prolong
the survival time of patients. For example, Kv10.1 can participate
in metabolic adaptation to cancer cells by regulating mitochondrial
dynamics and inhibit Kv10.1 expression or function, resulting in
mitochondrial fragmentation, increased reactive oxygen species and
increased autophagy. Kv10.1 endogenous overexpression cells were
also more sensitive to mitochondrial metabolic inhibitors than low
expression cells, indicating that they were more dependent on
mitochondrial function. Therefore, the combined treatment of Kv10.1
functional monoclonal antibody and mitochondrial metabolism
inhibitor leads to the enhanced efficacy of the inhibitor (31). Kv10.1 specific scFv was fused with
soluble TNF-related apoptosis inducing ligand. The combination of
ligands and different chemotherapeutic drugs can overcome drug
resistance and selectively induce apoptosis (170).
Summary
The Kv10.1 channel is overexpressed in 70% of tumor
cells and has carcinogenic characteristics, regulates cell
proliferation, survival, angiogenesis, migration and invasion and
is associated with the formation and progress of invasive tumors.
However, although Kv10.1, as a potential target for cancer
treatment, has been confirmed in vivo and in vitro
experiments and some carcinogenesis mechanisms have been proved,
the specific mechanism of Kv10.1 participating in the occurrence
and development of tumors is not completely clear.
However, several specific Kv10.1 inhibitors have
been used in animal models and achieved good results, which can
inhibit the proliferation of tumor cells, induce tumor cell
apoptosis, preserve normal cells and make cancer cells more
sensitive to chemotherapy drugs. If these specific Kv10.1
inhibitors are combined with the current routine clinical
treatment, it will be a potential therapeutic strategy to make them
really useful tumor targets.
Availability of data and materials
Not applicable.
Authors' contributions
Investigation was by YX, SD, BW, JS, LD, LL, XW and
YC. Writing the original draft was by YX and WH. Data curation was
by YX, SD, BW, JS, LD, XW and YC. Conceptualization was by YX.
Writing, reviewing and editing was by WH, LL, HA, XW and YC.
Methodology was by SD. Resources were provided by BW, HA, XW and
YC. Supervision was by HA, XW and YC. Data authentication is not
applicable. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Key Research and
Development Plan of China (grant no. 2023YFF1205500), Central
Government Guides Local Funds for Science and Technology
Development for Hebei Province (grant nos. 246Z2701G and
254Z2702G), Science Research Project of Hebei Education Department
(grant no. QN2025020) and Shijiazhuang Science and Technology
Cooperation Special Project (grant no. SJZZXB24007).
References
|
1
|
Pardo LA and Stühmer W: Eag1: An emerging
oncological target. Cancer Res. 68:1611–1613. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kaplan WD and Trout WE III: The behavior
of four neurological mutants of Drosophila. Genetics. 61:399–409.
1969. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thi Hong Van N and Hyun Nam J:
Intermediate conductance calcium-activated potassium channel
(KCa3.1) in cancer: Emerging roles and therapeutic potentials.
Biochem Pharmacol. 230:1165732024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Luis E, Anaya-Hernández A, León-Sánchez P
and Durán-Pastén ML: The Kv10.1 Channel: A promising target in
cancer. Int J Mol Sci. 23:84582022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cheng S, Jiang D, Lan X, Liu K and Fan C:
Voltage-gated potassium channel 1.3: A promising molecular target
in multiple disease therapy. Biomed Pharmacother. 175:1166512024.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
He S, Moutaoufik MT, Islam S, Persad A, Wu
A, Aly KA, Fonge H, Babu M and Cayabyab FS: HERG channel and
cancer: A mechanistic review of carcinogenic processes and
therapeutic potential. Biochim Biophys Acta Rev Cancer.
1873:1883552020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gao S, Wang W, Ye W and Wang K: The
mechanism study of Eag1 potassium channel in gastric cancer. Transl
Cancer Res. 11:3827–3840. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen J, Xuan Z, Song W, Han W, Chen H, Du
Y, Xie H, Zhao Y, Zheng S and Song P: EAG1 enhances hepatocellular
carcinoma proliferation by modulating SKP2 and metastasis through
pseudopod formation. Oncogene. 40:163–176. 2021. View Article : Google Scholar
|
|
9
|
Chávez-López MG, Zúñiga-García V,
Hernández-Gallegos E, Vera E, Chasiquiza-Anchatuña CA, Viteri-Yánez
M, Sanchez-Ramos J, Garrido E and Camacho J: The combination
astemizole-gefitinib as a potential therapy for human lung cancer.
Onco Targets Ther. 10:5795–5803. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pardo LA, Contreras-Jurado C, Zientkowska
M, Alves F and Stühmer W: Role of voltage-gated potassium channels
in cancer. J Membr Biol. 205:115–124. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cázares-Ordoñez V and Pardo LA: Kv10.1
potassium channel: From the brain to the tumors. Biochem Cell Biol.
95:531–536. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Badaoui M, Mimsy-Julienne C, Saby C, Van
Gulick L, Peretti M, Jeannesson P, Morjani H and Ouadid-Ahidouch H:
Collagen type 1 promotes survival of human breast cancer cells by
overexpressing Kv10.1 potassium and Orai1 calcium channels through
DDR1-dependent pathway. Oncotarget. 9:24653–24671. 2017. View Article : Google Scholar
|
|
13
|
Na W, Ma B, Shi S, Chen Y, Zhang H, Zhan Y
and An H: Procyanidin B1, a novel and specific inhibitor of Kv10.1
channel, suppresses the evolution of hepatoma. Biochem Pharmacol.
178:1140892020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding XW, Yan JJ, An P, Lü P and Luo HS:
Aberrant expression of ether à go-go potassium channel in
colorectal cancer patients and cell lines. World J Gastroenterol.
13:1257–1261. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
de Guadalupe Chávez-López M,
Hernández-Gallegos E, Vázquez-Sánchez AY, Gariglio P and Camacho J:
Antiproliferative and proapoptotic effects of astemizole on
cervical cancer cells. Int J Gynecol Cancer. 24:824–828. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hui C, Lan Z, Yue-Li L and Li-Lin H and
Li-Lin H: Knockdown of Eag1 expression by RNA interference
increases chemosensitivity to cisplatin in ovarian cancer cells.
Reprod Sci. 22:1618–1626. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guy HR, Durell SR, Warmke J, Drysdale R
and Ganetzky B: Similarities in amino acid sequences of Drosophila
eag and cyclic nucleotide-gated channels. Science. 254:7301991.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Occhiodoro T, Bernheim L, Liu JH, Bijlenga
P, Sinnreich M, Bader CR and Fischer-Lougheed J: Cloning of a human
Ether-a-Go-Go potassium channel expressed in myoblasts at the onset
of fusion. FEBS Lett. 434:177–182. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Whicher JR and MacKinnon R: Structure of
the Voltage-Gated K+ Channel Eag1 reveals an alternative voltage
sensing mechanism. Science. 353:664–669. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barros F, de la Peña P, Domínguez P,
Sierra LM and Pardo LA: The EAG Voltage-dependent K+ channel
subfamily: Similarities and differences in structural organization
and gating. Front Pharmacol. 11:4112020. View Article : Google Scholar
|
|
21
|
Yellen G: The voltage-gated potassium
channels and their relatives. Nature. 419:35–42. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Morais Cabral JH, Lee A, Cohen SL, Chait
BT, Li M and Mackinnon R: Crystal structure and functional analysis
of the HERG potassium channel N terminus: A eukaryotic PAS domain.
Cell. 95:649–655. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li S, Yang F, Sun D, Zhang Y, Zhang M, Liu
S, Zhou P, Shi C, Zhang L and Tian C: Cryo-EM structure of the
hyperpolarization-activated inwardly rectifying potassium channel
KAT1 from Arabidopsis. Cell Res. 30:1049–1052. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ju M and Wray D: Molecular regions
responsible for differences in activation between heag channels.
Biochem Biophys Res Commun. 342:1088–1097. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Terlau H, Heinemann SH, Stuhmer W, Pongs O
and Ludwig J: Amino terminal-dependent gating of the potassium
channel rat eag is compensated by a mutation in the S4 segment. J
Physiol. 502:537–543. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ludwig J, Terlau H, Wunder F, Brüggemann
A, Pardo LA, Marquardt A, Stühmer W and Pongs O: Functional
expression of a rat homologue of the voltage gated either á go-go
potassium channel reveals differences in selectivity and activation
kinetics between the Drosophila channel and its mammalian
counterpart. EMBO J. 13:4451–4458. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lorinczi E, Gómez-Posada JC, de la Peña P,
Tomczak AP, Fernández-Trillo J, Leipscher U, Stühmer W, Barros F
and Pardo LA: Voltage-dependent gating of KCNH potassium channels
lacking a covalent link between voltage-sensing and pore domains.
Nat Commun. 6:66722015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tomczak AP, Fernández-Trillo J, Bharill S,
Papp F, Panyi G, Stühmer W, Isacoff EY and Pardo LA: A new
mechanism of voltage-dependent gating exposed by KV10.1 channels
interrupted between voltage sensor and pore. J Gen Physiol.
149:577–593. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fukai R, Saitsu H, Tsurusaki Y, Sakai Y,
Haginoya K, Takahashi K, Hubshman MW, Okamoto N, Nakashima M,
Tanaka F, et al: De novo KCNH1 mutations in four patients with
syndromic developmental delay, hypotonia and seizures. J Hum Genet.
61:381–387. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin TF, Lin IW, Chen SC, Wu HH, Yang CS,
Fang HY, Chiu MM and Jeng CJ: The Subfamily-specific Assembly of
Eag and ErgK+ Channels Is Determined by both the amino and the
carboxyl recognition domains. J Biol Chem. 289:22815–22834. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schӧnherr R, Lober K and Heinemann SH:
Inhibition of human ether à go-go potassium channels by
Ca(2+)/calmodulin. Embo J. 19:3263–3271. 2000. View Article : Google Scholar
|
|
32
|
Warmke J, Drysdale R and Ganetzky B: A
distinct potassium channel polypeptide encoded by the Drosophila
eag locus. Science. 252:1560–1562. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Napp J, Monje F, Stühmer W and Pardo LA:
Glycosylation of Eag1 (Kv10.1) potassium channels: Intracellular
trafficking and functional consequences. J Biol Chem.
280:29506–29512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pardo LA, Camino D, Sánchez A, Alves F,
Brüggemann A, Beckh S and Stühmer W: Oncogenic potential of EAG
K(+) channels. EMBO J. 18:5540–5547. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mortensen LS, Schmidt H, Farsi Z,
Barrantes-Freer A, Rubio ME, Ufartes R, Eilers J, Sakaba T, Stühmer
W and Pardo LA: KV 10.1 opposes activity-dependent increase in
Ca2+ influx into the presynaptic terminal of the
parallel fibre-Purkinje cell synapse. J Physiol. 593:181–196. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hernández-Reséndiz I, Pacheu-Grau D,
Sánchez A and Pardo LA: Inhibition of Kv10.1 channels sensitizes
mitochondria of cancer cells to antimetabolic agents. Cancers
(Basel). 12:9202020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gubič Š, Toplak Ž, Shi X, Dernovšek J,
Hendrickx LA, Pinheiro-Junior EL, Peigneur S, Tytgat J, Pardo LA,
Peterlin Mašič L and Tomašič T: New diarylamine KV10.1 inhibitors
and their anticancer potential. Pharmaceutics. 14:19632022.
View Article : Google Scholar
|
|
38
|
Girault A, Peretti M, Badaoui M, Hémon A,
Morjani H and Ouadid-Ahidouch H: The N and C-termini of SPCA2
regulate differently Kv10.1 function: Role in the collagen
1-induced breast cancer cell survival. Am J Cancer Res. 11:251–263.
2021.PubMed/NCBI
|
|
39
|
Chávez-López MG, Zúñiga-García V,
Pérez-Carreón JI, Avalos-Fuentes A, Escobar Y and Camacho J: Eag1
channels as potential early-stage biomarkers of hepatocellular
carcinoma. Biologics. 10:139–148. 2016.PubMed/NCBI
|
|
40
|
Menéndez ST, Villaronga MA, Rodrigo JP,
Alvarez-Teijeiro S, García-Carracedo D, Urdinguio RG, Fraga MF,
Pardo LA, Viloria CG, Suárez C and García-Pedrero JM: Frequent
aberrant expression of the human ether à go-go (hEAG1) potassium
channel in head and neck cancer: Pathobiological mechanisms and
clinical implications. J Mol Med (Berl). 90:1173–1184. 2012.
View Article : Google Scholar
|
|
41
|
Kang C, Li X, Yang X, Cheng X, Zhang D and
Wei X: Voltage-gated potassium channels associated with head and
neck cancer. Biochim Biophys Acta Rev Cancer. 1880:1893402025.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Canella R, Brugnoli F, Gallo M, Keillor
JW, Terrazzan A, Ferrari E, Grassilli S, Gates EWJ, Volinia S,
Bertagnolo V, et al: A multidisciplinary approach establishes a
link between transglutaminase 2 and the Kv10.1 Voltage-dependent K+
channel in breast cancer. Cancers (Basel). 15:1782022. View Article : Google Scholar
|
|
43
|
Ding XW, Luo HS, Jin X, Yan JJ and Ai YW:
Aberrant expression of Eag1 potassium channels in gastric cancer
patients and cell lines. Med Oncol. 24:345–350. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang BC and Liu WX: Study on the effect
and mechanism of tetrandrine combined with cisplatin in inhibiting
the growth of cervical cancer cells. Modern Oncol. 32:2364–2370.
2024.
|
|
45
|
Qiu S, Wang Q, Jiang H and Feng L:
Immunohistochemistry staining of Eag1 and p16/Ki-67 can help
improve the management of patients with cervical intraepithelial
Neoplasia after cold knife conversion. Diagn Pathol. 19:972024.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ortiz CS, Montante-Montes D, Saqui-Salces
M, Hinojosa LM, Gamboa-Dominguez A, Hernández-Gallegos E,
Martínez-Benítez B, Del Rosario Solís-Pancoatl M, Garcia-Villa E,
Ramírez A, et al: Eag1 potassium channels as markers of cervical
dysplasia. Oncol Rep. 26:1377–1383. 2011.PubMed/NCBI
|
|
47
|
Hartung F, Krüwel T, Shi X, Pfizenmaier K,
Kontermann R, Chames P, Alves F and Pardo LA: A Novel Anti-Kv10.1
nanobody fused to Single-Chain TRAIL enhances apoptosis induction
in cancer cells. Front Pharmacol. 11:6862020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bernal-Ramos G, Hernández-Gallegos E, Vera
E, Chávez-López MG, Zúñiga-García V, Sánchez-Pérez Y, Garrido E and
Camacho J: Astemizole inhibits cell proliferation in human prostate
tumorigenic cells expressing ether à-go-go-1 potassium channels.
Cell Mol Biol (Noisy-le-grand). 63:11–13. 2017.
|
|
49
|
Söğüt F, Çömelekoğlu Ü, Dervişoğlu H,
Eroğlu P, Yalin S and Yilmaz NŞ: Effect of imipramine on ether
à-go-go potassium channel (Kv1.10) expression in DU145 prostate
cancer cells. Andrologia. 54:e142912022. View Article : Google Scholar
|
|
50
|
Guasti L, Crociani O, Redaelli E, Pillozzi
S, Polvani S, Masselli M, Mello T, Galli A, Amedei A, Wymore RS, et
al: Identification of a posttranslational mechanism for the
regulation of hERG1 K+ channel expression and hERG1 current density
in tumor cells. Mol Cell Biol. 28:5043–5060. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gavrilova-Ruch O, Schönherr K, Gessner G,
Schönherr R, Klapperstück T, Wohlrab W and Heinemann SH: Effects of
imipramine on ion channels and proliferation of IGR1 melanoma
cells. J Membr Biol. 188:137–149. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Meyer R, Schönherr R, Gavrilova-Ruch O,
Wohlrab W and Heinemann SH: Identification of ether à go-go and
calcium-activated potassium channels in human melanoma cells. J
Membr Biol. 171:107–115. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ma B, Shi S, Guo W, Zhang H, Zhao Z and An
H: Liensinine, a novel and Food-derived compound, exerts potent
antihepatoma efficacy via inhibiting the Kv10.1 channel. J Agric
Food Chem. 72:4689–4702. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Agarwal JR, Griesinger F, Stühmer W and
Pardo LA: The potassium channel Ether à go-go is a novel prognostic
factor with functional relevance in acute myeloid leukemia. Mol
Cancer. 9:182010. View Article : Google Scholar
|
|
55
|
Mello de Queiroz F, Suarez-Kurtz G,
Stühmer W and Pardo LA: Ether à go-go potassium channel expression
in soft tissue sarcoma patients. Mol Cancer. 5:422006. View Article : Google Scholar
|
|
56
|
Mészáros B, Csoti A, Szanto TG, Telek A,
Kovács K, Toth A, Volkó J and Panyi G: The hEag1 K+ channel
inhibitor astemizole stimulates Ca2+ Deposition in SaOS-2 and MG-63
osteosarcoma cultures. Int J Mol Sci. 23:105332022. View Article : Google Scholar
|
|
57
|
Cole KS and Moore JW: Liquid junction and
membrane potentials of the squid giant axon. J Gen Physiol.
43:971–980. 1960. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cole KS and Moore JW: Ionic current
measurements in the squid giant axon membrane. J Gen Physiol.
44:123–167. 1960. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Cole KS and Moore JW: Potassium ion
current in the squid giant axon: Dynamic characteristic. Biophys J.
1:1–14. 1960. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Schönherr R, Gessner G, Löber K and
Heinemann SH: Functional distinction of human EAG1 and EAG2
potassium channels. FEBS Lett. 514:204–208. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Terlau H, Ludwig J, Steffan R, Pongs O,
Stühmer W and Heinemann SH: Extracellular Mg2+ regulates activation
of rat eag potassium channel. Pflugers Arch. 432:301–312. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kazmierczak M, Zhang X, Chen B, Mulkey DK,
Shi Y, Wagner PG, Pivaroff-Ward K, Sassic JK, Bayliss DA and Jegla
T: External pH modulates EAG superfamily K+ channels through
EAG-specific acidic residues in the voltage sensor. J Gen Physiol.
141:721–735. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Brelidze TI, Carlson AE and Zagotta WN:
Absence of direct cyclic nucleotide modulation of mEAG1 and hERG1
channels revealed with fluorescence and electrophysiological
methods. J Biol Chem. 284:27989–27997. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Barriga-Montoya C, Huanosta-Gutiérrez A,
Reyes-Vaca A, Hernández-Cruz A, Picones A and Gómez-Lagunas F:
Inhibition of the K+ conductance and Cole-Moore shift of the
oncogenic Kv10.1 channel by amiodarone. Pflugers Arch. 470:491–503.
2018. View Article : Google Scholar
|
|
65
|
Gómez-Lagunas F and Barriga-Montoya C:
Mibefradil inhibition of the Cole-Moore shift and K+-conductance of
the tumor-related Kv10.1 channel. Channels (Austin). 11:373–376.
2017. View Article : Google Scholar
|
|
66
|
Martin S, Lino de Oliveira C, Mello de
Queiroz F, Pardo LA, Stühmer W and Del Bel E: Eag1 potassium
channel immunohistochemistry in the CNS of adult rat and selected
regions of human brain. Neuroscience. 155:833–844. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ufartes R, Schneider T, Mortensen LS, de
Juan Romero C, Hentrich K, Knoetgen H, Beilinson V, Moebius W,
Tarabykin V, Alves F, et al: Behavioural and functional
characterization of Kv10.1 (Eag1) knockout mice. Hum Mol Genet.
22:2247–2262. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kohl T, Lörinczi E, Pardo LA and Stühmer
W: Rapid internalization of the oncogenic K+ channel K(V)10.1. PLoS
One. 6:e263292011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Pardo LA and Stühmer W: The roles of K(+)
channels in cancer. Nat Rev Cancer. 14:39–48. 2014. View Article : Google Scholar
|
|
70
|
Del-Río-Ibisate N, Granda-Díaz R, Rodrigo
JP, Menéndez ST and García-Pedrero JM: Ion Channel dysregulation in
head and neck cancers: Perspectives for clinical application. Rev
Physiol Biochem Pharmacol. 181:375–427. 2021. View Article : Google Scholar
|
|
71
|
Li Z, Zhu K, Gong X, Vasilescu S, Sun Y,
Hong K, Li H, Li L and Shan Y: Inducing polyclonal Eag1-Specific
antibodies by vaccination with a linear epitope immunogen and its
relation to breast tumorigenesis. Pathol Oncol Res. 23:761–767.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
García-Quiroz J, González-González ME,
Díaz L, Ordaz-Rosado D, Segovia-Mendoza M, Prado-García H, Larrea F
and García-Becerra R: Astemizole, an Inhibitor of Ether-à-go-go-1
potassium channel, increases the activity of the tyrosine kinase
inhibitor gefitinib in breast cancer cells. Rev Invest Clin.
71:186–194. 2019.
|
|
73
|
Haas B, Roth I, Säcker L, Wos-Maganga M,
Beltzig L and Kaina B: Apoptotic and senolytic effects of hERG/Eag1
channel blockers in combination with temozolomide in human
glioblastoma cells. Naunyn Schmiedebergs Arch Pharmacol.
398:12267–12278. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Rashno Z, Rismani E, Ghasemi JB, Mansouri
M, Shabani M, Afgar A, Dabiri S, Rezaei Makhouri F, Hatami A and
Harandi MF: Design of ion channel blocking, toxin-like Kunitz
inhibitor peptides from the tapeworm, Echinococcus granulosus, with
potential anti-cancer activity. Sci Rep. 13:114652023. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Toplak Ž, Hendrickx LA, Gubič Š, Možina Š,
Žegura B, Štern A, Novak M, Shi X, Peigneur S, Tytgat J, et al: 3D
Pharmacophore-based discovery of novel KV10.1 inhibitors with
antiproliferative activity. Cancers (Basel). 13:12442021.
View Article : Google Scholar
|
|
76
|
Meléndez TA, Huanosta-Gutiérrez A,
Barriga-Montoya C, González-Andrade M and Gómez-Lagunas F:
Dronedarone blockage of the tumor-related Kv10.1 channel: A
comparison with amiodarone. Pflugers Arch. 472:75–87. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Brüggemann A, Stühmer W and Pardo LA:
Mitosis-promoting factor-mediated suppression of a cloned delayed
rectifier potassium channel expressed in Xenopus oocytes. Proc Natl
Acad Sci USA. 94:537–542. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Downie BR, Sánchez A, Knötgen H,
Contreras-Jurado C, Gymnopoulos M, Weber C, Stühmer W and Pardo LA:
Eag1 expression interferes with hypoxia homeostasis and induces
angiogenesis in tumors. J Biol Chem. 283:36234–36240. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Gómez-Varela D, Zwick-Wallasch E, Knötgen
H, Sánchez A, Hettmann T, Ossipov D, Weseloh R, Contreras-Jurado C,
Rothe M, Stühmer W and Pardo LA: Monoclonal antibody blockade of
the human Eag1 potassium channel function exerts antitumor
activity. Cancer Res. 67:7343–7349. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Sales TT, Resende FF, Chaves NL,
Titze-De-Almeida SS, Báo SN, Brettas ML and Titze-De-Almeida R:
Suppression of the Eag1 potassium channel sensitizes glioblastoma
cells to injury caused by temozolomide. Oncol Lett. 12:2581–2589.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang X, Chen Y, Liu H, Guo S, Hu Y, Zhan Y
and An H: A novel anti-cancer mechanism of Nutlin-3 through
downregulation of Eag1 channel and PI3K/AKT pathway. Biochem
Biophys Res Commun. 517:445–451. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
García-Quiroz J, García-Becerra R,
Santos-Martínez N, Barrera D, Ordaz-Rosado D, Avila E, Halhali A,
Villanueva O, Ibarra-Sánchez MJ, Esparza-López J, et al: In vivo
dual targeting of the oncogenic Ether-à-go-go-1 potassium channel
by calcitriol and astemizole results in enhanced antineoplastic
effects in breast tumors. BMC Cancer. 14:7452014. View Article : Google Scholar
|
|
83
|
Shen C, Kuang Y, Xu S, Li R, Wang J, Zou
Y, Wang C, Xu S, Liang L, Lin C, et al: Nitidine chloride inhibits
fibroblast like synoviocytes-mediated rheumatoid synovial
inflammation and joint destruction by targeting KCNH1. Int
Immunopharmacol. 101:1082732021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Chen ZD, Liu QJ, Zeng WR, Wu XY, Lin B and
Wu J: Expression of Ether à go-go 1 and its molecular mechanism of
regulating the malignant phenotype of osteosarcoma. Zhonghua Zhong
Liu Za Zhi. 38:818–825. 2016.In Chinese. PubMed/NCBI
|
|
85
|
Valdés-Abadía B, Morán-Zendejas R,
Rangel-Flores JM and Rodríguez-Menchaca AA: Chloroquine inhibits
tumor-related Kv10.1 channel and decreases migration of MDA-MB-231
breast cancer cells in vitro. Eur J Pharmacol. 855:262–266. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Emigh Cortez AM, DeMarco KR, Furutani K,
Bekker S, Sack JT, Wulff H, Clancy CE, Vorobyov I and Yarov-Yarovoy
V: Structural modeling of hERG channel-drug interactions using
Rosetta. Front Pharmacol. 14:12441662023. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Abdelaziz R, Tomczak AP, Neef A and Pardo
LA: Revealing a hidden conducting state by manipulating the
intracellular domains in KV10.1 exposes the coupling between two
gating mechanisms. Elife. 12:RP914202024. View Article : Google Scholar
|
|
88
|
Wei AD, Burgraff NJ, Oliveira LM, Moreira
TS and Ramirez JM: Fentanyl blockade of K+ channels contributes to
wooden chest syndrome. J Physiol. 604:582–604. 2026. View Article : Google Scholar :
|
|
89
|
Wang ZJ, Ghorbani M, Chen X, Tiwari PB,
Klauda JB and Brelidze TI: Molecular mechanism of EAG1 channel
inhibition by imipramine binding to the PAS domain. J Biol Chem.
299:1053912023. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Camacho J: Ether à go-go potassium
channels and cancer. Cancer Lett. 233:1–9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Herrmann S, Ninkovic M, Kohl T, Lörinczi É
and Pardo LA: Cortactin controls surface expression of the
voltage-gated potassium channel K(V)10.1. J Biol Chem.
287:44151–44163. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Sánchez A, Urrego D and Pardo LA: Cyclic
expression of the voltage-gated potassium channel KV10.1 promotes
disassembly of the primary cilium. EMBO Rep. 17:708–723. 2016.
View Article : Google Scholar
|
|
93
|
Urrego D, Sánchez A, Tomczak AP and Pardo
LA: The electric fence to cell-cycle progression: Do local changes
in membrane potential facilitate disassembly of the primary
cilium?: Timely and localized expression of a potassium channel may
set the conditions that allow retraction of the primary cilium.
Bioessays. 39:16001902017. View Article : Google Scholar
|
|
94
|
Martinez N, Boire A and Deangelis LM:
Molecular interactions in the development of brain metastases. Int
J Mol Sci. 14:17157–17167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Rosendo-Pineda MJ, Moreno CM and Vaca L:
Role of ion channels during cell division. Cell Calcium.
91:1022582020. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Urrego D, Movsisyan N, Ufartes R and Pardo
LA: Periodic expression of Kv10.1 driven by pRb/E2F1 contributes to
G2/M progression of cancer and non-transformed cells. Cell Cycle.
15:799–811. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Movsisyan N and Pardo LA: Kv10.1 regulates
microtubule dynamics during mitosis. Cancers (Basel). 12:24092020.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Peretti M, Badaoui M, Girault A, Van
Gulick L, Mabille MP, Tebbakha R, Sevestre H, Morjani H and
Ouadid-Ahidouch H: Original association of ion transporters
mediates the ECM-induced breast cancer cell survival:
Kv10.1-Orai1-SPCA2 partnership. Sci Rep. 9:11752019. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ouadid-Ahidouch H, Le Bourhis X,
Roudbaraki M, Toillon RA, Delcourt P and Prevarskaya N: Changes in
the K+ current-density of MCF-7 cells during progression through
the cell cycle: Possible involvement of a h-ether.a-gogo K+
channel. Recept Channels. 7:345–356. 2001.PubMed/NCBI
|
|
100
|
Ouadid-Ahidouch H and Ahidouch A: K+
channel expression in human breast cancer cells: Involvement in
cell cycle regulation and carcinogenesis. J Membr Biol. 221:1–6.
2008. View Article : Google Scholar
|
|
101
|
Stansfeld CE, Röper J, Ludwig J, Weseloh
RM, Marsh SJ, Brown DA and Pongs O: Elevation of intracellular
calcium by muscarinic receptor activation induces a block of
voltage-activated rat ether-à-go-go channels in a stably
transfected cell line. Proc Natl Acad Sci USA. 93:9910–9914. 1996.
View Article : Google Scholar
|
|
102
|
Bronk P, Kuklin EA, Gorur-Shandilya S, Liu
C, Wiggin TD, Reed ML, Marder E and Griffith LC: Regulation of Eag
by Ca2+/calmodulin controls presynaptic excitability in Drosophila.
J Neurophysiol. 119:1665–1680. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Huang X and Jan LY: Targeting potassium
channels in cancer. J Cell Biol. 206:151–162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cook SJ and Lockyer PJ: Recent advances in
Ca(2+)-dependent Ras regulation and cell proliferation. Cell
Calcium. 39:101–112. 2006. View Article : Google Scholar
|
|
105
|
Monteith GR, McAndrew D, Faddy HM and
Roberts-Thomson SJ: Calcium and cancer: Targeting Ca2+ transport.
Nat Rev Cancer. 7:519–530. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Goshima T, Habara M, Maeda K, Hanaki S,
Kato Y and Shimada M: Calcineurin regulates cyclin D1 stability
through dephosphorylation at T286. Sci Rep. 9:127792019. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Sayedyahossein S, Thines L and Sacks DB:
Ca2+ signaling and the Hippo pathway: Intersections in cellular
regulation. Cell Signal. 110:1108462023. View Article : Google Scholar
|
|
108
|
Cavalli G and Heard E: Advances in
epigenetics link genetics to the environment and disease. Nature.
571:489–499. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Pulczinski JC, Shang Y, Dao T,
Limjunyawong N, Sun Q, Mitzner W, Cheng RY and Tang WY:
Multigenerational epigenetic regulation of allergic diseases:
Utilizing an experimental dust Mite-induced asthma model. Front
Genet. 12:6245612021. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Wang J, Seebacher N, Shi H, Kan Q and Duan
Z: Novel strategies to prevent the development of multidrug
resistance (MDR) in cancer. Oncotarget. 8:84559–84571. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Iacopetta D, Rosano C, Sirignano M,
Mariconda A, Ceramella J, Ponassi M, Saturnino C, Sinicropi MS and
Longo P: Is the way to fight cancer paved with gold? Metal-based
carbene complexes with multiple and fascinating biological
features. Pharmaceuticals (Basel). 13:912020. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Kulkarni P, Ramisetty S, Bruno D, Tan T,
Merla A and Salgia R: Phenotypic plasticity, Non-genetic
mechanisms, and immune drug resistance in cancer. Cancer Treat Res.
129:309–324. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wei JR, Lu MY, Wei TH, Fleishman JS, Yu H,
Chen XL, Kong XT, Sun SL, Li NG, Yang Y and Ni HW: Overcoming
cancer therapy resistance: From drug innovation to therapeutics.
Drug Resist Updat. 81:1012292025. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Liu S, Jiang A, Tang F, Duan M and Li B:
Drug-induced tolerant persisters in tumor: Mechanism, vulnerability
and perspective implication for clinical treatment. Mol Cancer.
24:1502025. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Madrid MF, Mendoza EN, Padilla AL,
Choquenaira-Quispe C, de Jesus Guimarães C, de Melo Pereira JV,
Barros-Nepomuceno FWA, Lopes Dos Santos I, Pessoa C, de Moraes
Filho Mo, et al: In vitro models to evaluate multidrug resistance
in cancer cells: Biochemical and morphological techniques and
pharmacological strategies. J Toxicol Environ Health B Crit Rev.
28:1–27. 2025. View Article : Google Scholar
|
|
116
|
Thomas C and Tampe R: Structural and
mechanistic principles of ABC transporters. Annu Rev Biochem.
89:605–636. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Mansoori B, Mohammadi A, Davudian S,
Shirjang S and Baradaran B: The different mechanisms of cancer drug
resistance: A brief review. Adv Pharm Bull. 7:339–348. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zheng HC: The molecular mechanisms of
chemoresistance in cancers. Oncotarget. 8:59950–59964. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Bai Y, Liao H, Liu T, Zeng X, Xiao F, Luo
L, Guo H and Guo L: MiR-296-3p regulates cell growth and multi-drug
resistance of human glioblastoma by targeting ether-à-go-go (EAG1).
Eur J Cancer. 49:710–724. 2013. View Article : Google Scholar
|
|
120
|
Sun XX, Hodge JJ, Zhou Y, Nguyen M and
Griffith LC: The eag potassium channel binds and locally activates
calcium/calmodulin-dependent protein kinase II. J Biol Chem.
279:10206–10214. 2004. View Article : Google Scholar
|
|
121
|
Huttenlocher A and Horwitz AR: Integrins
in cell migration. Cold Spring Harb Perspect Biol. 3:a0050742011.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Hsu PH, Ma YT, Fang YC, Huang JJ, Gan YL,
Chang PT, Jow GM, Tang CY and Jeng CJ: Cullin 7 mediates
proteasomal and lysosomal degradations of rat Eag1 potassium
channels. Sci Rep. 7:408252017. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Horst CH, Titze-de-Almeida R and
Titze-de-Almeida SS: The involvement of Eag1 potassium channels and
miR-34a in rotenone-induced death of dopaminergic SH-SY5Y cells.
Mol Med Rep. 15:1479–1488. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Lin H, Li Z, Chen C, Luo X, Xiao J, Dong
D, Lu Y, Yang B and Wang Z: Transcriptional and
post-transcriptional mechanisms for oncogenic overexpression of
ether à go-go K+ channel. PLoS One. 6:e203622011. View Article : Google Scholar
|
|
125
|
Wu X, Zhong D, Gao Q, Zhai W, Ding Z and
Wu J: MicroRNA-34a inhibits human osteosarcoma proliferation by
downregulating ether à go-go 1 expression. Int J Med Sci.
10:676–682. 2013. View Article : Google Scholar
|
|
126
|
Diaz L, Ceja-Ochoa L, Restrepo-Angulo I,
Larrea F, Avila-Chávez E, García-Becerra R, Borja-Cacho E, Barrera
D, Ahumada E, Gariglio P, et al: Estrogens and human papilloma
virus oncogenes regulate human ether-à-go-go-1 potassium channel
expression. Cancer Res. 69:3300–3307. 2009. View Article : Google Scholar
|
|
127
|
Scheffner M, Werness BA, Huibregtse JM,
Levine AJ and Howley PM: The E6 oncoprotein encoded by human
papillomavirus types 16 and 18 promotes the degradation of p53.
Cell. 63:1129–1136. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Borowiec AS, Hague F, Harir N, Guénin S,
Guerineau F, Gouilleux F, Roudbaraki M, Lassoued K and
Ouadid-Ahidouch H: IGF-1 activates hEAG K(+) channels through an
Akt-dependent signaling pathway in breast cancer cells: Role in
cell proliferation. J Cell Phvsiol. 212:690–701. 2007. View Article : Google Scholar
|
|
129
|
Sabbatini P and McCormick F:
Phosphoinositide 3-OH kinase (PI3K) and PKB/Akt delay the onset of
p53-mediated, transcriptionally dependent apoptosis. J Biol Chem.
274:24263–24269. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Yoon AR, Gao R, Kaul Z, Choi IK, Ryu J,
Noble JR, Kato Y, Saito S, Hirano T, Ishii T, et al: MicroRNA-296
is enriched in cancer cells and downregulates p21WAF1 mRNA
expression via interaction with its 3′ untranslated region. Nucleic
Acids Res. 39:8078–8091. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Song S, Zhang G, Chen X, Zheng J, Liu X,
Wang Y, Chen Z, Wang Y, Song Y and Zhou Q: HIF-1α increases the
osteogenic capacity of ADSCs by coupling angiogenesis and
osteogenesis via the HIF-1α/VEGF/AKT/mTOR signaling pathway. J
Nanobiotechnology. 21:2572023. View Article : Google Scholar
|
|
132
|
Wu X, Chen Z, Zeng W, Zhong Y, Liu Q and
Wu J: Silencing of Eag1 gene inhibits osteosarcoma proliferation
and migration by targeting STAT3-VEGF pathway. Biomed Res Int.
2015:6173162015. View Article : Google Scholar
|
|
133
|
Wu J, Wu X, Zhong D, Zhai W, Ding Z and
Zhou Y: Short Hairpin RNA (shRNA) Ether à go-go 1 (Eag1) inhibition
of human osteosarcoma angiogenesis via VEGF/PI3K/AKT signaling. Int
J Mol Sci. 13:12573–12583. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Sun XX, Bostrom SL and Grifith LC:
Alternative splicing of the eag potassium channel gene in
Drosophila generates a novel signal transduction scaffolding
protein. Mol Cel1 Neurosci. 40:338–343. 2009. View Article : Google Scholar
|
|
135
|
Wu J, Zhong D, Wei Y, Wu X, Kang L and
Ding Z: Potassium channel ether à go-go1 is aberrantly expressed in
human liposarcoma and promotes tumorigenesis. Biomed Res Int.
2014:3456782014. View Article : Google Scholar
|
|
136
|
Wu X, Zhong D, Lin B, Zhai W, Ding Z and
Wu J: p38 MAPK regulates the expression of ether à go-go potassium
channel in human osteosarcoma cells. Radiol Oncol. 47:42–49. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Hemmerlein B, Weseloh RM, Mello de Queiroz
F, Knötgen H, Sánchez A, Rubio ME, Martin S, Schliephacke T, Jenke
M, Heinz-Joachim-Radzun, Stühmer W and Pardo LA: Overexpression of
Eag1 potassium channels in clinical tumours. Mol Cancer. 5:412006.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Lei Z, Luo Y, Lu J, Fu Q, Wang C, Chen Q,
Zhang Z and Zhang L: FBXO22 promotes HCC angiogenesis and
metastasis via RPS5/AKT/HIF-1α/VEGF-A signaling axis. Cancer Gene
Ther. 32:198–213. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Zhang HP, Jiang RY, Zhu JY, Sun KN, Huang
Y, Zhou HH, Zheng YB and Wang XJ: PI3K/AKT/mTOR signaling pathway:
An important driver and therapeutic target in triple-negative
breast cancer. Breast Cancer. 31:539–551. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Liao Y, Wei Z, Xu H, Zhang Z and Zhu F:
Unlocking precision medicine: Innovative strategies for druggable
target identification and therapeutic enhancement. Precision
Medication. 1:1000022024. View Article : Google Scholar
|
|
141
|
Wang ZJ, Soohoo SM, Tiwari PB, Piszczek G
and Brelidze TL: Chlorpromazine binding to the PAS domains uncovers
the effect of ligand modulation on EAG channel activity. J Biol
Chem. 295:4114–4123. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Toplak Ž, Hendrickx LA, Abdelaziz R, Shi
X, Peigneur S, Tomašič T, Tytgat J, Peterlin-Mašič L and Pardo LA:
Overcoming challenges of HERG potassium channel liability through
rational design: Eag1 inhibitors for cancer treatment. Med Res Rev.
42:183–226. 2022. View Article : Google Scholar
|
|
143
|
Ghorbani M, Wang ZJ, Chen X, Tiwari PB,
Klauda JB and Brelidze TI: Chlorpromazine inhibits EAG1 channels by
altering the coupling between the PAS, CNBH and pore domains.
bioRxiv. Feb 26–2024. View Article : Google Scholar
|
|
144
|
Jhou AJ, Chang HC, Hung CC, Lin HC, Lee
YC, Liu WT, Han KF, Lai YW, Lin MY and Lee CH: Chlorpromazine, an
antipsychotic agent, induces G2/M phase arrest and apoptosis via
regulation of the PI3K/AKT/mTOR-mediated autophagy pathways in
human oral cancer. Biochem Pharmacol. 184:1144032021. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Gómez-Lagunas F, Carrillo E, Pardo LA and
Stühmer W: Gating modulation of the Tumor-related Kv10.1 Channel by
Mibefradil. J Cell Physiol. 232:2019–2032. 2017. View Article : Google Scholar
|
|
146
|
Moreels L, Bhat C, Voráčová M, Peigneur S,
Goovaerts H, Mäki-Lohiluoma E, Zahed F, Pardo LA, Yli-Kauhaluoma J,
Kiuru P and Tytgat J: Synthesis of novel purpurealidin analogs and
evaluation of their effect on the cancer-relevant potassium channel
KV10.1. PLoS One. 12:e01888112017. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Gardner A, Wu W, Thomson S, Zangerl-Plessl
EM, Stary-Weinzinger A and Sanguinetti MC: Molecular basis of
altered hERG1 channel gating induced by ginsenoside Rg3. Mol
Pharmacol. 92:437–450. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Wu W, Gardner A, Sachse FB and Sanguinetti
MC: Ginsenoside Rg3, a gating modifier of EAG Family K+ channels.
Mol Pharmacol. 90:469–482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Moreels L, Peigneur S, Galan DT, De Pauw
E, Béress L, Waelkens E, Pardo LA, Quinton L and Tytgat J: APETx4,
a novel sea anemone toxin and a modulator of the cancer-relevant
potassium channel KV10.1. Mar Drugs. 15:2872017. View Article : Google Scholar
|
|
150
|
Ma B, Shi S, Ren S, Qu C, Zhao Z and An H:
Corydaline binds to a druggable pocket of hEAG1 channel and
inhibits hepatic carcinoma cell viability. Eur J Pharmacol.
962:1762402024. View Article : Google Scholar
|
|
151
|
Wang JH, Nao JF, Zhang M and He P:
20(s)-ginsenoside Rg3 promotes apoptosis in human ovarian cancer
HO-8910 cells through PI3K/Akt and XIAP pathways. Tumour Biol Dec.
35:11985–11994. 2014. View Article : Google Scholar
|
|
152
|
Doyle DA, Morais Cabral J, Pfuetzner RA,
Kuo A, Gulbis JM, Cohen SL, Chait BT and MacKinnon R: The structure
of the potassium channel: Molecular basis of K+ conduction and
selectivity. Science. 280:69–77. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Bezanilla F and Armstrong CM: Negative
conductance caused by entry of sodium and cesium ions into the
potassium channels of squid axons. J Gen Physiol. 60:588–608. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Kim DM and Nimigean CM: Voltage-gated
potassium channels: A structural examination of selectivity and
gating. Cold Spring Harb Perspect Biol. 8:a0292312016. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Wang X, Chen Y, Li J, Guo S, Lin X, Zhang
H, Zhan Y and An H: Tetrandrine, a novel inhibitor of
ether-à-go-go-1 (Eag1), targeted to cervical cancer development. J
Cell Physiol. 234:7161–7173. 2019. View Article : Google Scholar
|
|
156
|
Garg V, Sachse FB and Sanguinetti MC:
Tuning of EAG K(+) channel inactivation: Molecular determinants of
amplification by mutations and a small molecule. J Gen Physiol.
140:307–324. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
García-Ferreiro RE, Kerschensteiner D,
Major F, Monje F, Stühmer W and Pardo LA: Mechanism of block of
hEag1 K+ channels by imipramine and astemizole. J Gen Physiol.
124:301–317. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Hernández-Meza JM, Mares-Sámano S and
Garduño-Juárez R: Insights into the molecular inhibition of the
oncogenic channel KV10.1 by globular toxins. J Chem Inf Model.
61:2328–2340. 2021. View Article : Google Scholar
|
|
159
|
Dai Y, Wei Z, Sephton CF, Zhang D,
Anderson DH and Mousseau DD: Haloperidol induces the nuclear
translocation of phosphatidylinositol 3′-kinase to disrupt Akt
phosphorylation in PC12 cells. J Psychiatry Neurosci. 32:323–330.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Bai YH, Yang X and Zeng ZW: Protective
effect of haloperidol on serum-deprived PC12 cells and its
underlying mechanism. J Psychiatry. 34:119–123. 2021.
|
|
161
|
Sahoo N, Yang K, Coburger I, Bernert A,
Swain SM, Gessner G, Kappl R, Kühl T, Imhof D, Hoshi T, et al:
Intracellular hemin is a potent inhibitor of the voltage-gated
potassium channel Kv10.1. Sci Rep. 12:146452022. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Ke Z, Tang Z, Shen D, Liu Y, Shu Y, Mu X,
Li Z, Xiang P, Zhong B, Hu X, et al: Co-Highly expressed SLC17A9
and KCNH1 as potential prognostic biomarkers and therapeutic
targets in clear cell renal cell carcinoma. Front Biosci (Landmark
Ed). 30:380612025. View Article : Google Scholar
|
|
163
|
Cortes-Hernández U, Lizardi-Aguilera TM,
Noriega-Mejía BJ, González-Macías J, García-Quiroz J, Díaz L,
Larrea F and Avila E: Prostaglandin E2 suppresses KCNH1 gene
expression and inhibits the proliferation of CaSki cervical cells
through its four prostanoid PTGER subtypes. Gene. 933:1489972025.
View Article : Google Scholar
|
|
164
|
Vargas-Castro R, García-Becerra R, Díaz L,
Avila E, Ordaz-Rosado D, Bernadez-Vallejo SV, Cano-Colín S, Camacho
J, Larrea F and García-Quiroz J: Enhancing tamoxifen therapy with
α-Mangostin: Synergistic antiproliferative effects on breast cancer
cells and potential reduced endometrial impact. Pharmaceuticals
(Basel). 16:15762023. View Article : Google Scholar
|
|
165
|
Fan Z, Chen Y, Yan D and Li Q: Effects of
differentially methylated CpG sites in enhancer and promoter
regions on the chromatin structures of target LncRNAs in breast
cancer. Int J Mol Sci. 25:110482024. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Ding XW, Wang XG, Luo HS, Tan SY, Gao S,
Luo B and Jiang H: Expression and prognostic roles of Eag1 in
resected esophageal squamous cell carcinomas. Dig Dis Sci.
53:2039–2044. 2008. View Article : Google Scholar
|
|
167
|
Asher V, Khan R, Warren A, Shaw R,
Schalkwyk GV, Bali A and Sowter HM: The Eag potassium channel as a
new prognostic marker in ovarian cancer. Diagn Pathol. 5:782010.
View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Martínez R, Stühmer W, Martin S, Schell J,
Reichmann A, Rohde V and Pardo L: Analysis of the expression of
Kv10.1 potassium channel in patients with brain metastases and
glioblastoma multiforme: Impact on survival. BMC Cancer.
15:8392015. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Pardridge WM: The blood-brain barrier:
Bottleneck in brain drug development. NeuroRx. 2:3–14. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Hartung F and Pardo LA: Guiding TRAIL to
cancer cells through Kv10.1 potassium channel overcomes resistance
to doxorubicin. Eur Biophys J. 45:709–719. 2016. View Article : Google Scholar : PubMed/NCBI
|