Introduction
Oculocutaneous albinism (OCA) is a group of
inherited disorders characterized by defective melanin
biosynthesis, which results in congenital hypopigmentation of the
hair, skin and eyes. At least 16 genes have been found to be
associated with various forms of albinism. Mutations in these
genes, which regulate the processes of melanin synthesis and
distribution, mcause various types of OCA (1), which are classified into 2 groups:
non-syndromic and syndromic. Non-syndromic OCA is subdivided into 4
forms based on clinical and genetic findings as follows: OCA1,
OCA2, OCA3 and OCA4. OCA1, the most severe, is caused by mutations
in the tyrosinase (TYR) gene and is further divided into two
types; OCA1A and OCA1B. OCA1A is characterized by a life-long
absence of melanin pigmentation, whereas patients with OCA1B may
develop certain cutaneous and ocular pigmentation with age. The
other forms are milder clinical phenotypes revealing some pigment
accumulation over time, and include OCA2, OCA3 and OCA4, caused by
mutations in the OCA2, TRYP1 and SLC45A2
genes, respectively (1).
The prevalence of the various forms of OCA varies
widely in different populations. OCA1 is the most common, with a
prevalence of 1 per 40,000 individuals in most populations
(2). Although OCA2 and OCA3 are
common in African OCA patients, these forms are uncommon in
Caucasian and Asian populations (3,4).
OCA4 has been reported to be the second most common form in Far
East Asian populations, with a prevalence of 24–27% in Japanese
patients (5,6) and 12.6% in Chinese patients (7). In addition, it is well known that a
certain mutation has been commonly identified in a specific ethnic
group due to a founder effect in OCA. Although different forms of
OCA are caused by mutations in a variety of genes, clinical
phenotypes are not always distinguishable; therefore, molecular
diagnosis has become a useful and essential tool for genetic
counseling.
Only 5 studies of mutational screenings involving 13
Korean OCA patients exist (8,9). Of
these studies, causative mutations in TYR and SLC45A2
were found in 6 (46.2%) patients and 1 (7.7%) patient,
respectively. The present study included 11 Korean OCA patients and
their parents, who were referred to the Genetics Clinic at Ajou
University Hospital. We analyzed the mutations found in TYR
and SLC45A2 to determine their mutation spectrums. The
clinical features of the mutations were compared and analyzed to
determine genotype-phenotype correlations.
Materials and methods
Study patients and clinical
evaluations
Korean OCA patients and their parents were recruited
for the study at the Genetics Clinic of Ajou University Hospital
between December 2004 and May 2011. The study protocol was reviewed
and approved by the Institutional Review Board of the Ajou
University Hospital, and written informed consent was obtained from
all subjects or from their parents. The diagnostic inclusion
criteria were based on the presence of the following clinical
characteristics: varying degrees of hypopigmentation of the skin
and body hair, and abnormal opthalmological findings, including
photophobia, nystagmus, reduced visual acuity, strabismus, iris
translucency, fundus hypopigmentation or foveal hypoplasia.
Patients with OCA forms, including Hermansky-Pudlak,
Chediak-Higashi and Griscelli syndromes were excluded based on
clinical features. The typical clinical features in the OCA
patients were analyzed to predict genotype-phenotype correlations
in the cases with detected mutations.
If OCA1 was determined by TYR gene analysis,
OCA1 patients were further classified into 2 types: OCA1A and
OCA1B. OCA1A was defined as the presence of white hair and skin
throughout life, and OCA1B was defined as the presence of white
hair at birth, particularly eyelash hair, that subsequently
developed pigmentation in the first decade of life.
Mutation identification
Genomic DNA was isolated from the peripheral blood
leukocytes of the study subjects using a DNA isolation kit (Qiagen
GmbH, Hilden, Germany). The DNA samples were first screened for
TYR mutations. All 5 coding exons and the intronic flanking
regions of the TYR gene were amplified using a polymerase
chain reaction (PCR) using 6 specific primer pairs (Table I). PCR was performed in a reaction
volume of 25 μl containing 100 ng of genomic DNA template, 200 nM
of each primer, 200 nM of each dNTP, 1X PCR buffer and 2.5 units of
LA Taq DNA polymerase (Takara Bio Inc., Shiga, Japan).
Amplifications were conducted over 35 cycles; each cycle consisted
of denaturation at 95°C for 30 sec, annealing at 51°C for 1 min and
extension at 72°C for 1 min, with a final extension at 72°C for 10
min. If no mutation was identified using TYR analysis, the
sample was analyzed for SLC45A2 mutations. All 7 coding
exons and the intronic flanking regions were PCR-amplified using 10
specific primer pairs (Table I).
Amplifications were conducted over 35 cycles; each cycle consisted
of denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec
and extension at 72°C for 40 sec, with a final extension at 68°C
for 10 min.
 | Table IPCR primer pairs used for the
amplification of TYR and SLC45A2 coding
sequences. |
Table I
PCR primer pairs used for the
amplification of TYR and SLC45A2 coding
sequences.
| TYR |
|---|
|
|---|
| Exon | Forward | Reverse |
|---|
| 1–1 | CCC ACT GGT GGG ATA
CGA | GGT CCC CAA AAG CCA
AAC |
| 1–2 | CCC TAG AGC CTG TGT
CTC C | CCC TGC CTG AAG AAG
TGA T |
| 2 | CCT CAG GAG AAG TCT
AAC AAC | ACA ACA CAT ATT CTT
GGT C |
| 3 | TGG GTA TCC AGA ATG
TAA A | TTT AAA TCC AAT GAG
CAC G |
| 4 | TTT TAA TAT ATG CCT
TAT TTT AC | GGT AAC ACT AGA TTC
AGC AA |
| 5 | CTC CAA AGG ACT GTG
AAA GGA | GGT CTT TAC AGA AAA
ATA C |
| SLC45A2 |
|---|
|
|---|
| Exon | Forward | Reverse |
|---|
| 1-1 | AGG CTC CAC GTC AAA
TCC AG | GGT CAC ATA CGC TGC
CTC CA |
| 1-2 | CAG ACT CAT CAT GCA
CAG CA | ATG CCC ACG AGC ATC
ATG AC |
| 1-3 | CAG CAT TGT GTG GTT
CCT CA | GGT CAA ACA CAT GAA
CAT CCT C |
| 2 | AAC GTG GAT GAT TCT
AAA ACA GGA | CTC ATT GTC TGG GGA
GCT GA |
| 3-1 | GGG AGT GTC TAT GCA
TGA GG | GAT AGA ACC ATA CTC
GTA CAT TCC |
| 3-2 | GCC CCA CTT ACA GAG
GTT GC | CAA CAA AGA GCA AGA
ATA TTT TCC CTT G |
| 4 | AGC TGG CTG AGT TTC
TGC AG | CCT CAA CAG GTG TTA
ATG GAG G |
| 5 | AGA GGT GGA GAA GCA
GAG TG | GAA GAC ATC CTT AGG
AGA GAG |
| 6 | ATG AGG CAC TGC CAG
CTG TA | CCC AAG GCA GAG GTT
CAA TG |
| 7 | GCC CTA AAT GAC AGT
TCC TTG | TGT GCT TCA CTG TCT
CTG AG |
PCR products were separated on 1.2% agarose gels and
bands were visualized with ethidium bromide. Subsequently, DNA
sequencing reactions were conducted using the same primer pairs and
a BigDye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems,
Foster City, CA, USA), according to the manufacturer’s
instructions. The sequencing reaction mixtures were electrophoresed
and analyzed using an ABI3130xl Genetic Analyzer (Applied
Biosystems) and Sequencing Analysis v.5.2 software.
To predict the functional impact of novel amino acid
changes, we performed additional molecular analyses on DNA from the
patients’ parents and 50 healthy controls. Additionally, novel
sequence alterations were assessed using in silico
prediction algorithms, Polymorphism Phenotyping (PolyPhen) and
Sorting Intolerant from Tolerant (SIFT), and the In the PolyPhen
program. A position-specific independent count (PSIC) score of
>2.0 for an amino acid variant indicates ‘probably damaging to
protein function’; a score of 1.5–2.0, ‘possibly damaging’ and a
score of <1.5, indicates ‘benign or unknown’ (10,11).
Results
Study population and clinical findings in
Korean OCA patients
A total of 12 patients (M:F=5:7) and their parents
were enrolled. The ages at enrollment ranged from 0.8 to 29.0 years
(median, 3.4 years). OCA subtypes were previously unknown. There
were 5 patients with a complete absence of pigmentation who had
been clinically diagnosed with OCA1A, and the remaining 7 patients
had minimal to moderate pigmentation that had gradually accumulated
with age. Patient clinical characteristics are shown in Table II. Ultimately, 24 alleles from 12
OCA patients from nonconsanguineous families were examined.
| Table IIClinical parameters and mutations in
12 Korean OCA families. |
Table II
Clinical parameters and mutations in
12 Korean OCA families.
| Patient | Age (years) | Gender | Diagnosis | Causing gene | Mutation | Clinical
phenotype |
|---|
| | | | |
|
|
|---|
| | | | | Paternal | Maternal | Hair color | Skin color | Nystagmus |
|---|
| 1 | 2.2 | M | OCA1A | TYR | c.929insC | c.929insC | White | White | + |
| 2 | 0.9 | F | OCA1A | TYR | p.R299H | c.929insC | White | White | + |
| 3 | 2.1 | M | OCA1A | TYR | c.929insC | p.D383N | White | White | + |
| 4 | 14.2 | M | OCA1A | TYR | p.D383N | p.R77Q | White | White | + |
| 5 | 27.0 | M | OCA1A | TYR | p.R77Q | c.1037-7T>A;
c.1037-10delTT | White | White | + |
| 6 | 1.1 | F | OCA1B | TYR | c.929insC | c.1037-7T>A;
c.1037-10delTT | Blond | White | + |
| 7 | 0.8 | F | OCA1B | TYR | c.1037-7T>A;
c.1037-10delTT | Not identified | Blond | White | + |
| 8 | 29.0 | F | OCA1B | TYR | p.R52I | c.1037-7T>A;
c.1037-10delTT | Blond | White | + |
| 9 | 4.5 | M | OCA4 | SLC45A2 | p.D93N | p.D157N | Blond | White | + |
| 10 | 5.9 | F | OCA | - | Not identified | Not identified | Blond | White | + |
| 11 | 1.0 | F | OCA | - | Not identified | Not identified | Blond | White | - |
| 12 | 29.0 | F | OCA | - | Not identified | Not identified | Blond | White | + |
Spectrum of TYR and SLC45A2
mutations
Nine of 12 (75.0%) patients were identified using
molecular analyses of TYR and SLC45A2. The remaining
3 (25.0%) patients were unable to be identified from the mutational
screening for these 2 genes. There were 2 mutational alleles
identified from 8 patients and 1 mutational allele from 1
patient.
OCA1 confirmed by mutations in the TYR gene
was observed in 8 patients (66.7%) (Table II). OCA4 was diagnosed in 1
patient (8.3%) by SLC45A2 analysis. This patient had demonstrated
improvement in pigment phenotype with age, and his hair color had
changed to light yellow.
TYR analysis identified a total of 6 distinct
mutations, consisting of 4 missense, 1 frameshift and 1 splice
site, in 15 alleles of 8 patients (Table III). The mutations were located
in exons 1, 2, and 3 and in intron 2. The 6 mutations had already
been reported as pathological mutations. The most frequently
detected mutation was c.929insC, which was found in 5 (31.3%)
alleles of 4 patients. c.1037-7T>A/c.1037-10delTT, p.D383N,
p.R77Q, p.R52I and p.R299H mutations were identified in 4 (25.0%)
alleles of 4 patients, 2 (12.5%) alleles of 2 patients, 2 (12.5%)
alleles of 2 patients, 1 (6.3%) allele of 1 patient, and 1 (6.3%)
allele of 1 patient, respectively.
| Table IIIAllelic frequencies of the TYR
gene in 8 Korean OCA1 patients. |
Table III
Allelic frequencies of the TYR
gene in 8 Korean OCA1 patients.
| Location | Nucleotide
change | Amino acid
change | No. (%) of alleles
identified |
|---|
|
|---|
| Korean patients
(n=16) | Japanese patients
(n=60) | Chinese patients
(n=178) |
|---|
| Exon 1 | c.155G>T | p.R52I | 1 (6.3) | 0 (0) | 0 (0) |
| c.230G>A | p.R77Q | 2 (12.5) | 9 (15.0) | 7 (3.9) |
| Exon 2 | c.896G>A | p.R299H | 1 (6.3) | 0 (0) | 26 (14.6) |
| c.929insC | p.R311LfsX7 | 5 (31.3) | 30 (50.0) | 25 (14.0) |
| Intron 2 | c.1037-7T>A | Mis-splicing | 4 (25.0) | 5 (8.3) | 5 (2.8) |
| c.1037-10delTT | | | | |
| Exon 3 | c.1147G>A | p.D383N | 2 (12.5) | 2 (3.3) | 1 (0.6) |
| Total | | | 15 (93.8) | 46 (76.7) | 64 (35.9) |
There were 4 patients in whom mutations could not be
identified in the coding sequences or exon/intron boundaries of
TYR. These patients were screened for SLC45A2
mutations. A known disease-causing mutation, c.469G>A (p.D157N),
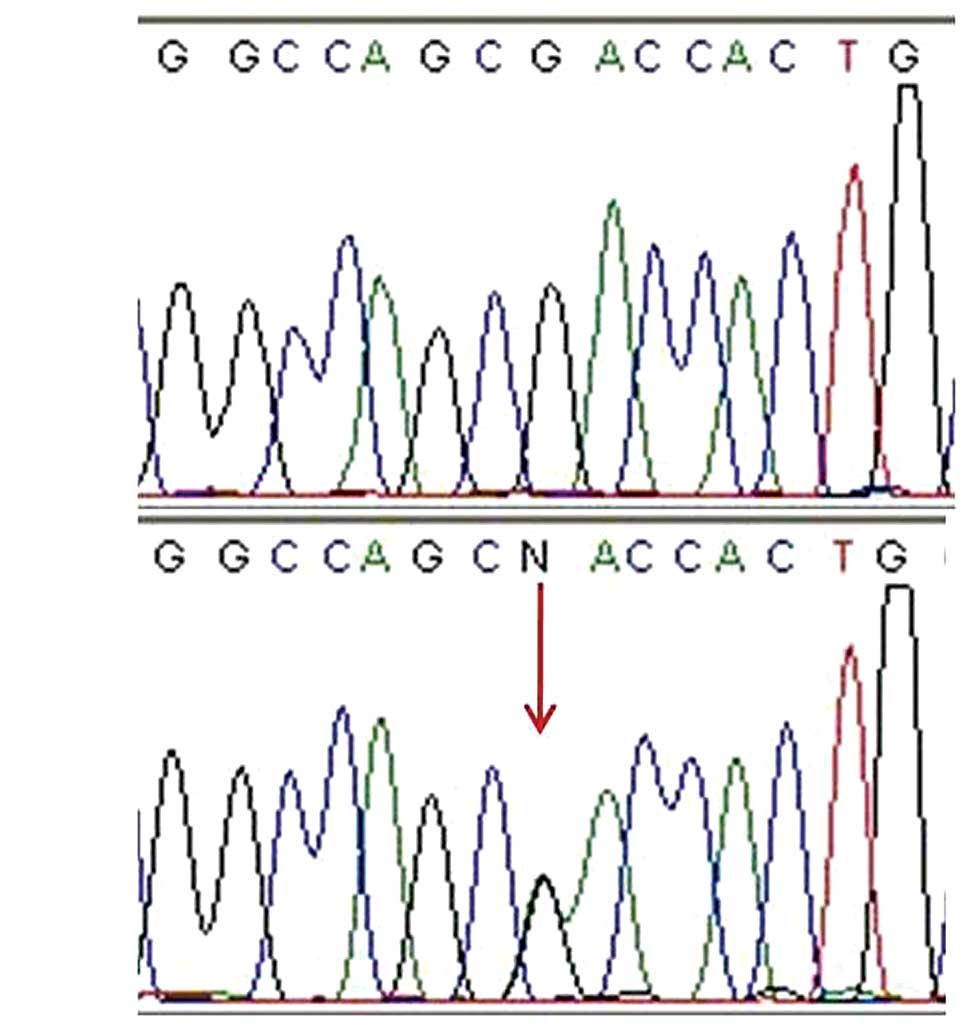
and a novel sequence alteration, c.277G>A (p.D93N), was also
detected (Fig. 1). p.D93N is
considered to be a pathological mutation, since it was not detected
in 50 unaffected controls and it is located in amino acid residues
that are conserved in zebrafish, mice, rhesus monkeys and humans.
The SIFT and PISC scores were 0.00 and 1.851, respectively,
indicating that p.D93N may affect protein structure and possibly be
a damaging mutation.
Discussion
OCA1 is caused by mutations in the TYR gene
on chromosome 11q14.3. The gene consists of 5 exons spanning
approximately 65 kb of genomic DNA, and encodes a protein of 529
amino acids (12). Tyrosinase is a
glycoprotein and a copper-containing oxidase that is involved in
the formation of pigments, including melanins and other
polyphenolic compounds. It catalyzes the rate-limiting conversions
of tyrosine to dopa, dopa to dopaquinone, and possibly
5,6-dihydroxyindole to indole-5,6 quinone (13). During melanogenesis, tyrosinase may
be released from the endoplasmic reticulum in the presence of a
protonophore or proton pump inhibitors that increase the pH of
intracellular organelles. Tyrosinase is then normally transported
to the Golgi, and then to the melanosomes via the endosomal sorting
system. For melanin synthesis, tyrosinase is sorted to the
melanosomes via a dileucine-based signal (14). Mutations in the TYR gene
appear to cause tyrosinase to be retained in the endoplasmic
reticulum, with subsequent early degradation and destruction of
tyrosinase activity (15).
At present, only 6 pathological TYR mutations
have been detected in Korean patients (8,16),
whereas more than 200 different mutations in TYR have been
reported worldwide (17,18). We examined 24 alleles from the
entire TYR gene and identified 6 different mutations in 15
(62.5%) alleles. These mutations had all been previously reported
in Korean patients (Table III).
According to a previous study, c.929insC was the most common of the
6 known Korean mutations, with an allele frequency of 0.5 (8). In this study, c.929insC was also the
most common mutation, with an allele frequency of 0.31. This
mutation is also known to be the most common TYR mutation in
Japanese patients, with an allele frequency of 0.50 (19,20),
and the second most common mutation in Chinese patients, with an
allele frequency of 0.14 (7).
c.929insC has only been identified in Far East Asian populations
thus far, including Korean, Japanese and Chinese. This mutation is
an insertion mutation, which causes a frameshift and premature
termination of tyrosinase synthesis (p.R311LfsX7). It is regarded
to be more commonly associated with OCA1A phenotypes than with
OCA1B (19), and in this study 3
(1 homozygote and 2 compound heterozygotes) of 4 patients with this
mutation also had OCA1A phenotypes. Another mutation,
c.1037-7T>A/c.1037-10delTT, was identified in 4 alleles.
Although c.1037-7T>A has been reported worldwide (21), c.1037-7T>A/c.1037-10delTT has
only been identified in Korean, Japanese and Chinese patients, with
allele frequencies of 0.25, 0.08 and 0.03, respectively. This
mutation is located in intron 2 and causes mis-splicing of exon 3.
In previous studies, it was found in patients with OCA1B phenotypes
(8,19), and in this study 3 (compound
heterozygotes) of 4 patients with this mutation had OCA1B
phenotypes. p.R77Q and p.D383N mutations were found to be compound
heterozygotes in 2 patients. These missense mutations have been
reported in various ethnic groups, including Caucasian and Far East
Asian populations, and both have been frequently found in patients
with OCA1A phenotypes (19,21).
All 3 patients (compound heterozygotes) harboring these mutations
in our study were also classified with OCA1A. The other mutations,
p.R52I and p.R299H, were each identified in 1 patient. p.R52I is an
infrequent mutation and has been reported only in Caucasian
patients with OCA1A phenotypes (17), even though two other mutations on
the same amino acid location (p.R52K and p.R52T) were found in the
Indian population (22). p.R299H
has been identified worldwide (7,8,21)
and is also the most frequent mutation in Chinese OCA1 patients,
with an allele frequency of 0.15 (7). However, its allele frequency does not
appear to be high in our Korean OCA1 patients and, there is
currently no other study of an OCA1 Korean patient with this
mutation.
Notably, the mutation p.R278X is the third most
common in Japanese and Chinese patients, accounting for 11.6 and
11.8% of all reported TYR mutations, respectively. However,
this mutant allele has yet to be identified in Korean OCA1
patients.
A comparison of the results of previous studies on
Japanese and Chinese OCA1 patients and our results reveals that the
identified mutations largely overlapped among these Far East Asian
populations. The 6 mutations found in 93.8% of alleles in the
Korean patients have been detected in 76.7% of Japanese patients
and 35.9% of Chinese patients. The index of coincidence of this
comparison suggests that the Korean population is genetically
closer to the Japanese population than the Chinese one.
Mutations in the SLC45A2 gene (also known as
MATP) on chromosome 5p13.3 cause OCA4. The SLC45A2
gene consists of 7 exons spanning approximately 40 kb of genomic
DNA. The SLC45A2 protein of 530 amino acids, which contains 12
putative transmembrane domains, reveals sequence and structural
similarity to the plant sucrose transporter, and is only expressed
in melanocytes (23).
SLC45A2 encodes a melanocyte differentiation antigen that is
expressed in a high percentage of melanoma cell lines (24). Studies on the Japanese medaka fish
reveal that this protein is significant in pigmentation and may
function as a membrane transporter in melanosomes (23). Ultrastructural analysis has
demonstrated that the vesicles secreted from OCA4 melanocytes are
mostly early-stage melanosomes. In OCA4 melanocytes, tyrosinase
processing and intracellular trafficking to the melanosome is
disrupted, and the enzyme is abnormally secreted from the cells in
immature melanosomes, which disrupts the normal maturation process
of those organelles (25).
SLC45A2 mutations were first found in a Turkish OCA patient
in 2001 (26), since then more
than 30 mutations have been reported (5,7,9,27,28).
OCA4 appears to be a rare form of OCA in Caucasians,
and mutations in SLC45A2 have been reported as the cause of
disease in 5–8% of German OCA patients (27). However, OCA4 is the second most
common form in Japanese and Chinese populations, accounting for
24–27% and 13% of all OCA, respectively (5–7).
Clinical phenotypes of OCA4 vary from complete depigmentation to
partial pigmentation; and similarly in OCA1B certain patients
improve during the first decade of life (5). Based on our results which reveal that
the mutation spectrum of Korean OCA1 patients considerably overlaps
with those of Japanese and Chinese patients, we examined
SLC45A2 in 4 TYR mutation-negative patients. One
known mutation (p.D157N) and 1 novel sequence alteration (p.D93N)
were identified. Over 20 missense mutations have been reported in
SLC45A2, and the majority of them are located within or in
close proximity to the transmembrane domains, in areas that appear
to be essential for the function of the SLC45A2 protein
(23,26). The mutation p.D157N is known to be
the most common mutation in Japanese OCA4 patients, with an allele
frequency of 0.39 (6), and 1
Korean OCA4 patient was reported as a homozygote of this allele in
a previous study (9). Therefore,
this indicates a founder effect for p.D157N in Japanese and Korean
populations (29). Of note, in
Chinese OCA4 patients, the p.D160H allele accounted for
approximately half of the mutational SLC45A2 alleles, and
p.D157N was detected in just 1 allele with an allele frequency of
0.03 (7). This finding also
suggests a founder mutation in the Chinese Han population,
revealing a genetic disparity between the Japanese and Chinese
populations. A novel sequence alteration, p.D93N, is considered to
be a pathological mutation since this change is located close to
the second transmembrane domain composed of amino acids 70 to 92,
and it was not detected in 50 unaffected controls. It is also in an
amino acid residue that is conserved among various species. We also
predicted the functional impact of p.D93N using 2 in silico
prediction algorithms, and this novel change was predicted to be a
deleterious mutation.
There are certain limitations to the present study.
Firstly, this study was performed at one institution, which may
have caused selection bias. Secondly, the wide age distribution
among the patients may have affected the phenotypes. Lastly, we did
not perform any functional studies of the one putative novel
mutation. Additional studies of Korean OCA patients, with molecular
analyses of other OCA-associated genes, including OCA2 and
TYRP, may aid in predicting more accurate genotype-phenotype
correlations and provide information for genetic counseling.
Additionally, anthropological studies are required to confirm
ethnic differences in the mutational spectrum of OCA.
In conclusion, great progress in the detection and
analysis of OCA-causative genes has been achieved in recent years;
however, OCA remains a challenging disorder. Our study provides
certain information concerning the genetic basis of OCA in Korean
patients, and allows us to estimate the relative frequencies of
OCA1 and OCA4 in Korea.
References
|
1
|
Y TomitaT SuzukiGenetics of pigmentary
disordersAm J Med Genet C Semin Med
Genet1317581200410.1002/ajmg.c.30036
|
|
2
|
K GrønskovJ EkK
Brondum-NielsenOculocutaneous albinismOrphanet J Rare
Dis243200719006216
|
|
3
|
WS OettingRA KingMolecular basis of
albinism: mutations and polymorphisms of pigmentation genes
associated with albinismHum
Mutat1399115199910.1002/(SICI)1098-1004(1999)13:2%3C99::AID-HUMU2%3E3.0.CO;2-C10094567
|
|
4
|
C RooryckC RoudautE RobineJ MusebeckB
ArveilerOculocutaneous albinism with TYRP1 gene mutations in a
Caucasian patientPigment Cell
Res19239242200610.1111/j.1600-0749.2006.00298.x16704458
|
|
5
|
K InagakiT SuzukiH ShimizuOculocutaneous
albinism type 4 is one of the most common types of albinism in
JapanAm J Hum Genet74466471200410.1086/38219514961451
|
|
6
|
T SuzukiY TomitaRecent advances in genetic
analyses of oculocutaneous albinism types 2 and 4J Dermatol
Sci5119200810.1016/j.jdermsci.2007.12.00818407468
|
|
7
|
A WeiY WangY LongA comprehensive analysis
reveals mutational spectra and common alleles in Chinese patients
with oculocutaneous albinismJ Invest
Dermatol130716724201010.1038/jid.2009.33919865097
|
|
8
|
SK ParkKH LeeKC ParkJS LeeRA SpritzST
LeePrevalent and novel mutations of the tyrosinase gene in Korean
patients with tyrosinase-deficient oculocutaneous albinismMol
Cells718719119979163730
|
|
9
|
T SuzukiK InagakiK FukaiA ObanaST LeeY
TomitaA Korean case of oculocutaneous albinism type IV caused by a
D157N mutation in the MATP geneBr J
Dermatol152174175200510.1111/j.1365-2133.2005.06403.x
|
|
10
|
PC NgS HenikoffSIFT: predicting amino acid
changes that affect protein functionNucleic Acids
Res3138123814200310.1093/nar/gkg50912824425
|
|
11
|
V RamenskyP BorkS SunyaevHuman
non-synonymous SNPs: server and surveyNucleic Acids
Res3038943900200210.1093/nar/gkf49312202775
|
|
12
|
BS KwonAK HaqSH PomerantzR
HalabanIsolation and sequence of a cDNA clone for human tyrosinase
that maps at the mouse c-albino locusProc Natl Acad Sci
USA8474737477198710.1073/pnas.84.21.74732823263
|
|
13
|
CJ CookseyPJ GarrattEJ LandEvidence of the
indirect formation of the catecholic intermediate substrate
responsible for the autoactivation kinetics of tyrosinaseJ Biol
Chem2722622626235199710.1074/jbc.272.42.262269334191
|
|
14
|
H WatabeJC ValenciaK YasumotoRegulation of
tyrosinase processing and trafficking by organellar pH and by
proteasome activityJ Biol
Chem27979717981200410.1074/jbc.M30971420014634018
|
|
15
|
K ToyofukuI WadaRA SpritzVJ HearingThe
molecular basis of oculocutaneous albinism type 1 (OCA1): sorting
failure and degradation of mutant tyrosinases results in a lack of
pigmentationBiochem
J355259269200110.1042/0264-6021:355025911284711
|
|
16
|
KC ParkSK ParkYS LeeMutations of the
tyrosinase gene in three Korean patients with type I oculocutaneous
albinismJpn J Hum Genet41299305199610.1007/BF019131728996965
|
|
17
|
The Human Gene Mutation Databasehttp://www.hgmd.cf.ac.uk/ac/all.phpAccessed September
1, 2011
|
|
18
|
Albinism database: Mutations of the
tyrosinase gene associated with OCA1http://albinismdb.med.umn.edu/oca1mut.htmlAccessed
September 1, 2011
|
|
19
|
M GotoKC Sato-MatsumuraD SawamuraK YokotaH
NakamuraH ShimizuTyrosinase gene analysis in Japanese patients with
oculocutaneous albinismJ Dermatol
Sci35215220200410.1016/j.jdermsci.2004.06.00715381243
|
|
20
|
Y TomitaY MiyamuraM KonoR NakamuraJ
MatsunagaMolecular bases of congenital hypopigmentary disorders in
humans and oculocutaneous albinism 1 in JapanPigment Cell
Res13Suppl 8130134200010.1034/j.1600-0749.13.s8.23.x11041370
|
|
21
|
RA KingJ PietschJP FryerTyrosinase gene
mutations in oculocutaneous albinism 1 (OCA1): definition of the
phenotypeHum
Genet113502513200310.1007/s00439-003-0998-113680365
|
|
22
|
K RayM ChakiM SenguptaNovel human
pathological mutations. Gene symbol: TYR Disease: tyrosinase
deficiencyHum Genet1225562007
|
|
23
|
S FukamachiA ShimadaA ShimaMutations in
the gene encoding B, a novel transporter protein, reduce melanin
content in medakaNat Genet28381385200110.1038/ng58411479596
|
|
24
|
ME RayG WistowYA SuPS MeltzerJM TrentAIM1,
a novel non-lens member of the betagamma-crystallin super-family,
is associated with the control of tumorigenicity in human malignant
melanomaProc Natl Acad Sci
USA9432293234199710.1073/pnas.94.7.32299096375
|
|
25
|
GE CostinJC ValenciaWD VieiraML LamoreuxVJ
HearingTyrosinase processing and intracellular trafficking is
disrupted in mouse primary melanocytes carrying the underwhite (uw)
mutation. A model for oculocutaneous albinism (OCA) type 4J Cell
Sci11632033212200310.1242/jcs.00598
|
|
26
|
JM NewtonO Cohen-BarakN HagiwaraMutations
in the human orthologue of the mouse underwhite gene (uw) underlie
a new form of oculocutaneous albinism, OCA4Am J Hum
Genet69981988200110.1086/32434011574907
|
|
27
|
U RundshagenC ZuhlkeS OpitzE SchwingerB
Kasmann-KellnerMutations in the MATP gene in five German patients
affected by oculocutaneous albinism type 4Hum
Mutat23106110200410.1002/humu.1031114722913
|
|
28
|
A GargiuloF TestaS RossiMolecular and
clinical characterization of albinism in a large cohort of Italian
patientsInvest Ophthalmol Vis
Sci1412811289201110.1167/iovs.10-609120861488
|
|
29
|
K InagakiT SuzukiS ItoOCA4: evidence for a
founder effect for the p.D157N mutation of the MATP gene in
Japanese and KoreanPigment Cell
Res18385388200510.1111/j.1600-0749.2005.00261.x16162179
|