Introduction
The endocrine function of adipose tissue has been
elucidated in recent years, with the identification of
adipocytokines such as leptin, adiponectin and resistin. Adipose
tissue produces numerous cytokines and hormones, which act as
autocrine, paracrine and endocrine factors and participate in the
pathophysiological process of insulin resistance (IR) and chronic
inflammation (1,2). Retinol binding protein 4 (RBP4) is an
adipocytokine related to IR (3).
This was first shown in mice by Yang et al (3), who observed that knockout mice for
the gene encoding adipose-specific glucose transporter-4 (GLUT-4)
were insulin resistant in muscle and liver, and displayed increased
expression of the RBP4 gene. Graham et al (4) subsequently measured the serum RBP4
level, insulin resistance, and components of the metabolic syndrome
in lean and obese individuals with or without type 2 diabetes, and
found that the serum level of RBP4 correlates with insulin
resistance. Additional studies further proved that the level of
RBP4 in the blood associates with IR (5–8). IR
is followed by compensatory hyperinsulinemia. It is widely accepted
that hyperinsulinemia and insulin resistance are the main risk
factors of cardiovascular diseases (CVD), eventually leading to the
formation and development of atherosclerosis (9).
Vascular smooth muscle cells (VSMCs) have been
extensively used to study the pathological mechanisms underlying
atherosclerosis. Proliferation and migration of VSMCs is of
important value for the formation of coronary atherosclerosis and
the development of coronary heart disease (CHD). Insulin is a
highly potent cell growth factor, which can promote VSMC
proliferation and DNA synthesis, and plays an important role in the
formation of atherosclerosis (10,11).
Recently, RBP4, an adipocytokine related to IR, has been suggested
to play an important role in the occurrence and development of
atherosclerosis and CVD (12–14).
However, whether RBP4 is involved in insulin-induced proliferation
of VSMCs leading to atherosclerosis remains unclear.
Proliferation and migration of VSMCs are related to
a variety of signal transduction pathways, such as the
mitogen-activated protein kinase (MAPK) and the JAK/STAT pathway.
Insulin activates the MAPK pathway through the Grb2/SOS and RAS
proteins to promote cell growth and proliferation, and collagen
synthesis (15–17). IR is followed by compensatory
hyperinsulinemia, which promotes insulin-induced proliferation of
VSMCs via the SHC/Raf/MAPK pathway, and accelerates artery
atherosclerosis (18). JAK/STAT is
another important signal transduction pathway mediating cell
proliferation. Binding of cytokines such as interferon,
5-hydroxytryptamine, platelet-derived growth factor and others to
the specific receptor activates the protein tyrosine kinase (PTK)
Janus kinase (JAK), thereby activating signal transducer and
activator of transcription (STAT), and inducing cell proliferation.
Previous studies (19–21) have shown that the JAK/STAT
signaling pathway plays an important role in VSMC
proliferation.
Insulin promotes proliferation of VSMCs to induce
formation of atherosclerosis through the MAPK pathway. Ost et
al (22) examined the
mechanisms of action of RBP4 in primary human adipocytes.
RBP4-treated adipocytes displayed the same molecular defects in
insulin signaling, mediated by the insulin receptor substrate (IRS)
protein 1 and the MAP kinase, as adipocytes from patients with type
2 diabetes. Takebayashi et al (23) further showed that RBP4 has a robust
acute effect on the enhancement of NO production via stimulating
part of the PI3K/Akt/eNOS pathway and inhibiting insulin-induced
ET-1 secretion, probably via the MAPK pathway, eventually causing
vasodilatation. However, whether RBP4 is involved in
insulin-induced proliferation of VSMCs leading to atherosclerosis
remains unclear. In the present study, we evaluated the role of
RBP4 in this process and the underlying signaling pathways.
Materials and methods
Reagents
RBP4 protein was purchased from Sino Biological Inc.
(Beijing, China) and was dissolved in a solution comprising sterile
50 mM Tris, 10 mM CaCl2 and 150 mM NaCl at pH 7.5, at a
final concentration of 500 μg/ml. Mouse anti-extracellular
signal-regulated kinase (ERK)1/2 and -phospho-ERK1/2 (p-ERK1/2)
monoclonal antibodies, and rabbit polyclonal anti-JAK2, -p-JAK2,
-signal transducer and activator of transcription (STAT) 3 and
-p-STAT3 antibodies were purchased from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). HRP-conjugated goat anti-rabbit and
anti-rat IgGs were purchased from ZSGB-Bio (Beijing, China). The
specific inhibitors of ERK1/2 PD098059, and of JAK2 AG490, were
purchased from Sigma-Aldrich (St. Louis, MO, USA). Supersignal West
Pico Chemiluminescent substrate was purchased from Thermo
Scientific (Rockford, IL, USA).
Cell culture
The rat aortic smooth muscle cell (RASMC) line A10
was obtained from the American Type Culture Collection (Manassas,
VA, USA). The cells were cultured in Dulbecco’s modified Eagle’s
medium (DMEM) containing 10% fetal bovine serum (FBS), penicillin
(100 U/ml), streptomycin (100 μg/ml) and NaHCO3 (3.7
g/l), in a humidified atmosphere of 5% CO2 at 37°C.
Cells from passages 7 and 15 were used for the experiments.
Cell proliferation assays
Cell proliferation was analyzed by using the MTT and
a cell cycle assay. For the MTT assay, RASMCs were seeded in
96-well plates at a density of 0.6–1.0×104 cells/well in
DMEM supplemented with 10% FBS. After 24 h, the medium was replaced
with DMEM containing 1% FBS to render the cells quiescent for 24 h.
Then, cells were incubated for different time periods in DMEM
containing 1% FBS and different concentrations of insulin. DMEM
containing 1% FBS and RBP4 (1 or 4 μg/ml), PD098059
(5×10−5 M), or AG490 (5×10−5 M) was added to
the cells, followed by treatment with insulin (10−5 M).
Finally, 20 μl of 5 mg/ml MTT solution was added to each well and
incubated for 4 h. The supernatants were aspirated, and the
formazan crystals in each well were dissolved in 150 μl dimethyl
sulfoxide. Cell proliferation was assessed by measuring the
absorbance at 490 nm using a microplate reader (DTX800; Beckman
Coulter, Brea, CA, USA).
For cell cycle analysis by flow cytometry, RASMCs
were seeded in 6-well culture plates (1×105 cells/well).
After 24 h, the medium was replaced with DMEM containing 1% FBS to
render the cells quiescent for 24 h. Then, RASMCs were pre-treated
with RBP4 (1 or 4 μg/ml) for 1 h, or PD098059 (5×10−5
M)/AG490 (5×10−5 M) for 10 min, followed by treatment
with insulin (10−5 M) for 24 h. After 24 h, cells were
trypsinized and washed with phosphate-buffered saline (PBS) twice
before fixing in ice-cold 70% ethanol at 4°C overnight. After
removing ethanol by centrifugation, cells were washed with PBS and
treated, for 20 min in the dark, with 50 μg/ml propidium iodide
solution (Sigma-Aldrich) combined with 100 μg/ml DNase-free RNase,
0.1% Triton X-100 and 0.1 mM EDTA in PBS. After washing with PBS,
propidium iodide-stained cells were subjected to cell cycle
analysis using a FACScan flow cytometer (Becton Dickinson, Mountain
View, CA, USA) and the FlowJo 7.1.0 software (Tree Star Inc.,
Ashland, OR, USA). At least 10,000 cells were counted for each
sample. Data are presented as the percentage of cells in a given
subpopulation.
Western blotting
RASMCs were lysed in RIPA lysis buffer containing
0.1 M phenylmethylsulfonyl fluoride. After centrifugation at 12,000
rpm for 20 min, the protein concentrations of the supernatants were
determined with the bicinchoninic acid assay (Sigma-Aldrich). Equal
amounts of protein were mixed with sodium dodecyl sulfate (SDS)
buffer and incubated at 100°C for 5 min before loading. Then, equal
amounts of total protein were loaded and separated on a 15% SDS
polyacrylamide gel by electrophoresis at a constant 90-V voltage
for 2.5 h. Proteins were transferred under a standard module onto a
polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA,
USA) for 45 min. The membrane was blocked with a 5% dry
milk/Tris-buffered saline-Tween-20 (TBST) solution for 1 h at room
temperature, and then probed with primary antibodies (anti-ERK1/2,
-p-ERK1/2, -JAK2, -p-JAK2, -STAT3, -p-STAT3, dilution 1:1,000;
-β-actin, 1:3,000) in a 5% dry milk/TBST solution overnight at 4°C.
The membrane was rinsed several times with TBST buffer and then
incubated with HRP-conjugated secondary antibodies (1:5,000) for 1
h at room temperature. Excess secondary antibody was removed by 3–4
washes in TBST buffer and the targeted protein bands were detected
using an enhanced chemiluminescence (ECL) detection kit, followed
by exposure of the membrane to an X-ray film. Protein bands were
scanned and quantified using Image J software version 1.40
(National Institutes of Health, Bethesda, MD, USA). The quantified
data for each protein were normalized to those for β-actin.
Statistical analysis
Data are expressed as means ± SD and were
statistically analyzed by Student’s paired t-test for pairwise
comparisons or ANOVA followed by Newman-Student-Keuls test for
comparisons among multiple groups. P<0.05 was considered to
indicate a statistically significant difference. The SPSS 13.0
software was used for the analyses (IBM, Armonk, NY, USA).
Results
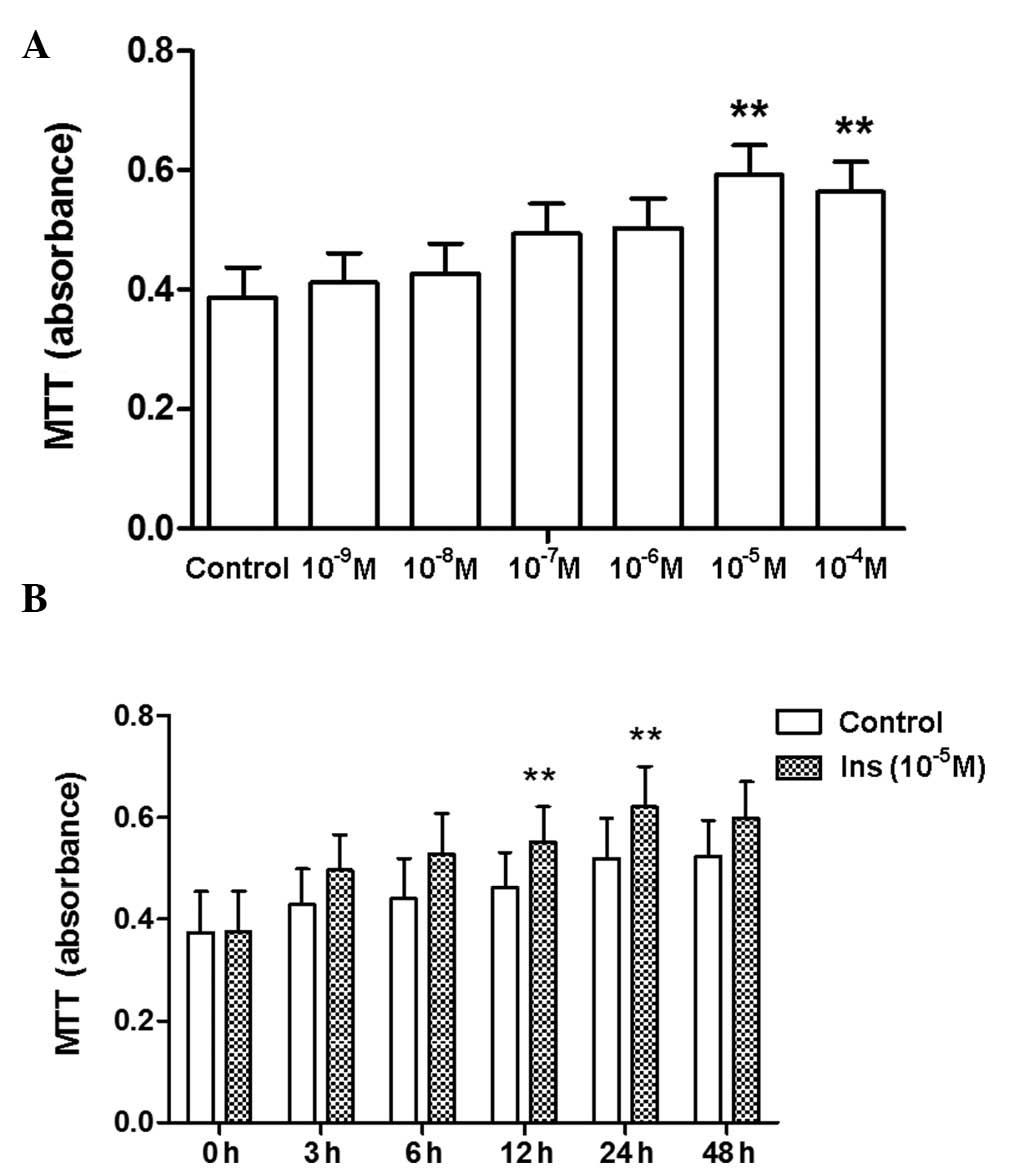
Proliferation of RAVSMCs induced by
insulin
As previously mentioned, hyperinsulinemia is
considered an independent risk factor for atherosclerosis and CVD.
Insulin can promote VSMC proliferation and DNA synthesis, playing
an important role in the formation of atherosclerosis. We therefore
used insulin to stimulate proliferation in vitro. In line
with results from a previous study (24), insulin induced proliferation of
RASMCs in a concentration- and time-dependent manner, as shown by
the increase in formazan absorbance (Fig. 1) and in the proportion of cells
detected at the S and G2 phases (data not shown).
Expression of ERK1/2, p-ERK1/2, JAK2,
p-JAK2, STAT3, p-STAT3 in RASMCs is induced by insulin
Since insulin induced proliferation of RASMCs in a
concentration- and time-dependent manner, we used the most
effective concentration (10−5 M) of insulin to further
study the activation of signal transduction pathways related to
cell proliferation. Insulin treatment enhanced the protein levels
of ERK1/2, p-ERK1/2, JAK2, p-JAK2, STAT3, and p-STAT3 in a
time-dependent manner, as shown by western blotting analysis
(Fig. 2).
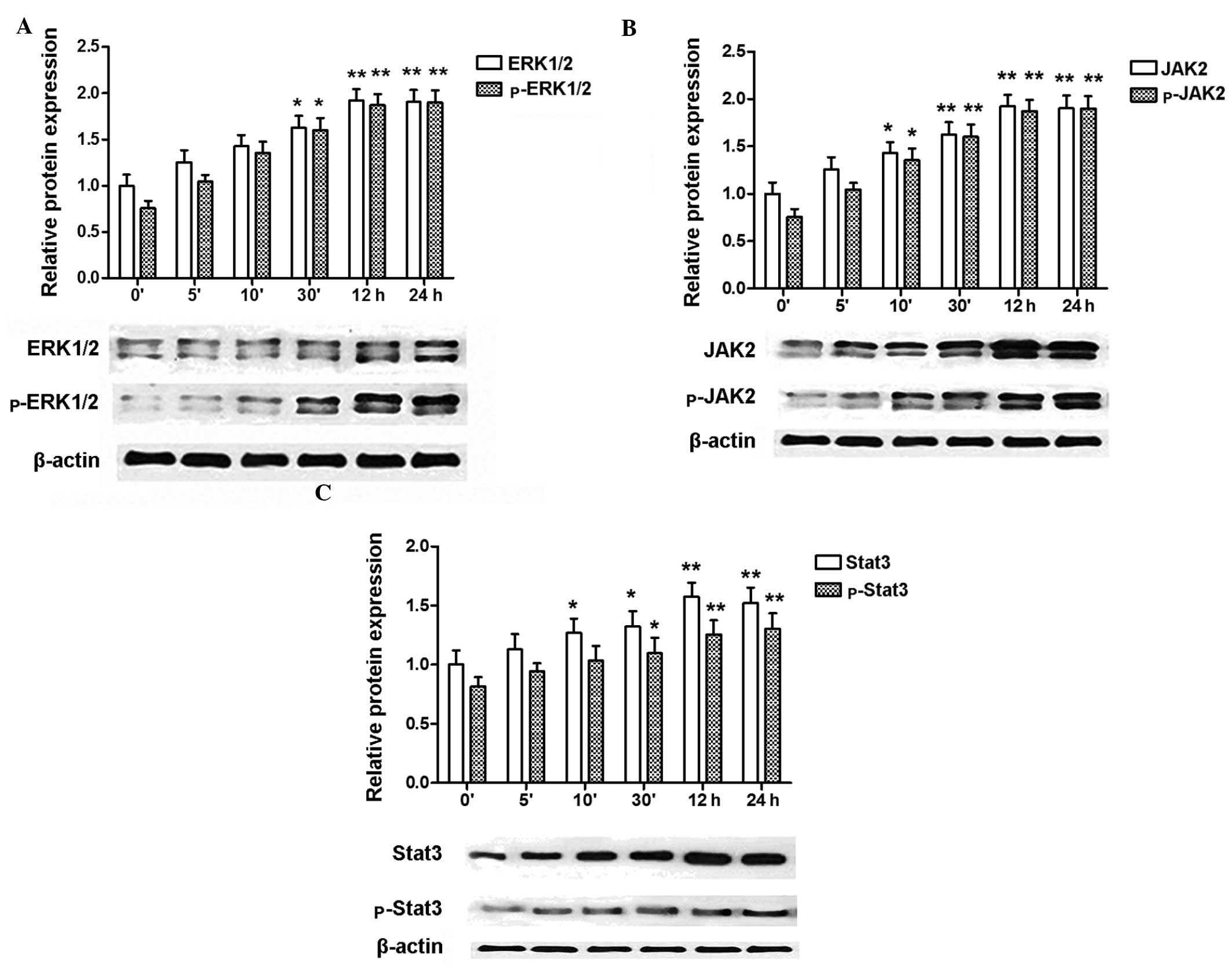 | Figure 2Expression of ERK2, p-ERK2, JAK2,
p-JAK2, STAT3, p-STAT3 in rat aortic smooth muscle cells (RASMCs)
induced by insulin. (A) Insulin induces the expression of ERK1/2,
p-ERK1/2 in a time-dependent manner, with the maximum level of both
proteins detected at 24 h. (B) Insulin induces the expression of
JAK2, p-JAK2 in a time-dependent manner, with the maximum level of
JAK2 detected at 12 h, and of p-JAK2 at 24 h. (C) Insulin induces
the expression of STAT3, p-STAT3 in a time-dependent manner, with
the maximum level of STAT3 detected at 12 h, and of p-STAT3 at 24
h. Blots are representative of three independent experiments.
*P<0.05, **P<0.01 vs. control (0′
without insulin). |
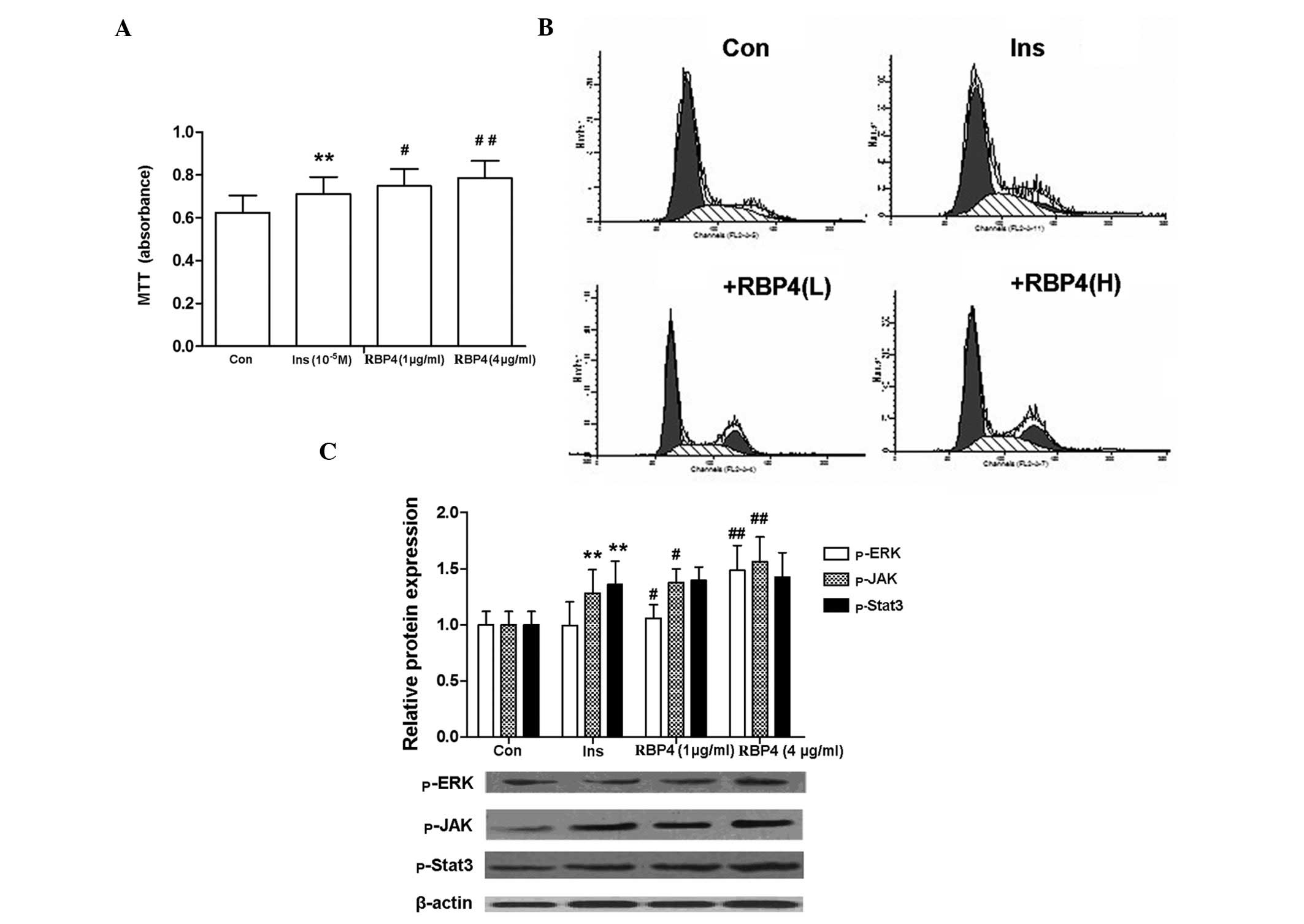
RBP4 enhances insulin-induced
proliferation of RASMCs
RBP4 enhanced the insulin-induced proliferation of
RASMCs, as shown by the increase in formazan absorbance and in the
proportion of cells at the S + G2 phases (Fig. 3A and B). The phosphorylation of
ERK1/2, JAK2 and STAT3 is an indicator of the activation of the
MAPK and the JAK/STAT signal transduction pathways. Notably, the
expression of p-ERK and p-JAK2 that was induced by insulin was
further enhanced by RBP4 treatment, while that of p-STAT3 remained
unchanged (Fig. 3C).
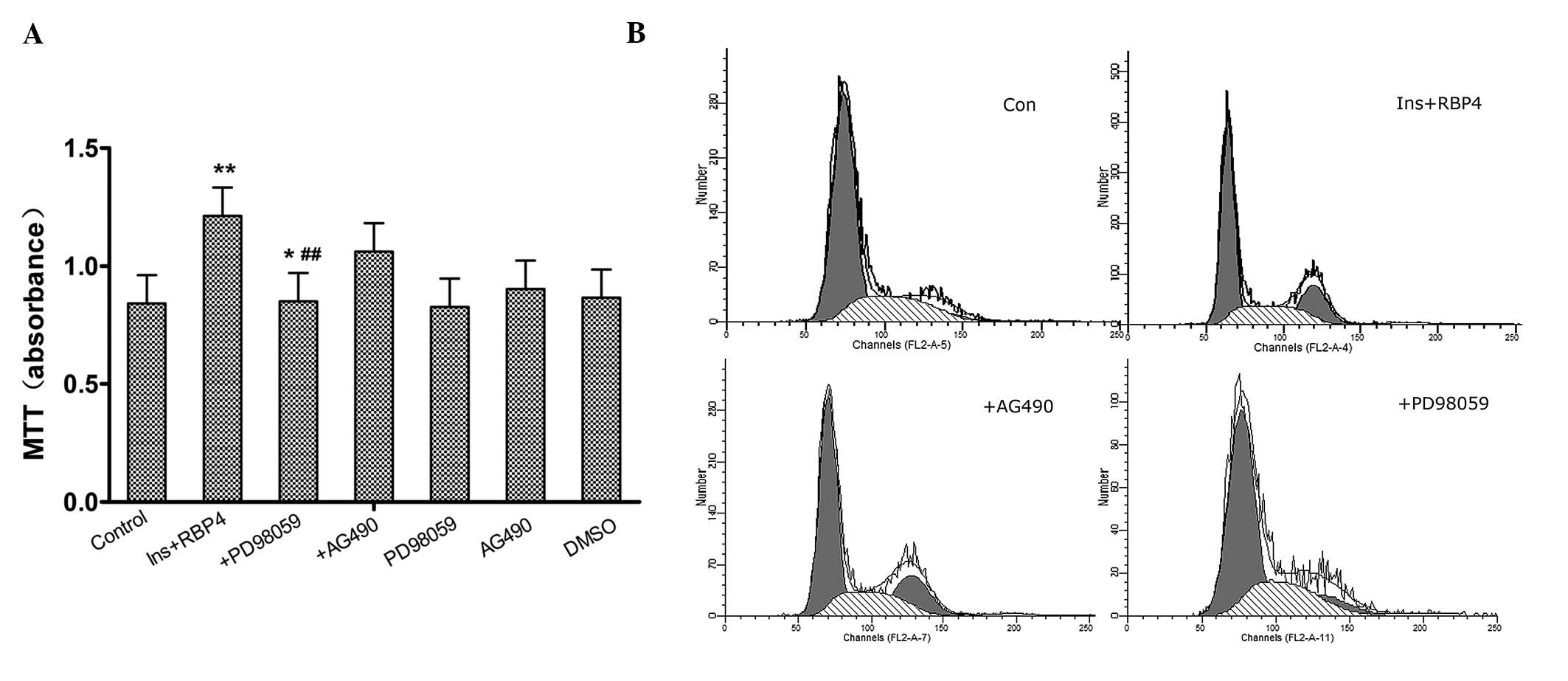
Involvement of ERK1/2 and JAK2/STAT3
pathways in the enhancement of RASMC proliferation by RBP4
To determine whether the ERK1/2 and/or the
JAK2/STAT3 pathway are involved in the enhancement of
insulin-induced cell proliferation caused by RBP4, PD98059 (the
specific ERK1/2 inhibitor) and AG490 (the specific JAK2 inhibitor)
were used. As shown by the MTT assay and flow cytometry,
pre-treatment of cells with PD98059 (5×10−5 M)
significantly (P<0.01) inhibited proliferation of RASMCs, while
pre-treatment with AG490 (5×10−5 M) had no effect on
proliferation (Fig. 4).
Discussion
The present study examined the effect of RBP4 on
insulin-induced proliferation of RASMCs. The main findings are the
following: i) insulin induced proliferation of RASMCs via the MAPK
and JAK2/STAT3 pathways; ii) RBP4 enhanced insulin-induced
proliferation of RASMCs and iii) RBP4 enhanced insulin-induced
proliferation of RASMCs via the MAPK, but not the JAK2/STAT3
pathway. Collectively, these findings indicate that RBP4 may play
an important role in the mediation of signals related to
proliferation of VSMCs induced by insulin, which suggests that RBP4
may contribute to vascular remodeling in hyperinsulinemia.
Since the first study of Yang et al (3) on RBP4 effects in mice, related
studies on RBP4 and insulin resistance, obesity, type 2 diabetes,
and CHD have provided contradictory results. Mahmoudi et al
(11) did not find a significant
difference in the RBP4 level between a non-diabetic population with
CHD and healthy controls. Mallat et al (12) attempted to prove that RBP4 can be
used as a predictor of CHD. The authors studied 1,036 patients with
CHD in a period of six years, and found that the risk of CHD
increased in cases of increased RBP4 level. However, following
adjustment of the data for other CHD risk factors, the positive
correlation between the RBP4 level and the risk of heart disease
proved not to be statistically significant, thereby suggesting that
RBP4 may not be a robust predictor of CHD. However, there are
studies that showed that RBP4 positively and significantly
correlates to carotid intima-media thickness, which is an
atherosclerosis indicator, and negatively correlates to
flow-mediated dilatation (25,26).
Independent correlations between RBP4 and the low-density
lipoprotein or the oxidized low-density lipoprotein have also been
reported (27,28). In addition, Cabré et al
(29) showed that the RBP4 level
is significantly higher in atherosclerotic patients with primary
type 2 diabetes mellitus than in those without atherosclerosis.
Overall, a potential correlation between RBP4 and atherosclerosis
and CHD can not be unequivocally claimed. Atherosclerosis is the
basic pathogenetic cause of CHD, while abnormal proliferation and
migration of VSMCs is one of the most common causes of
atherosclerosis, vascular restenosis and other CVDs.
Binding of insulin to insulin receptors (InsR) in
the target organs and tissues activates the MAPK pathway through
the Grb2/SOS and RAS proteins, thereby regulating gene
transcription to promote cell growth and proliferation. In IR, even
when pancreatic β cells show a good function, compensatory
hyperinsulinemia occurs and promotes insulin-induced proliferation
of VSMCs via the SHC/Raf/MAPK pathway; in these conditions,
hyperinsulinemia accelerates arterial atherosclerosis.
In agreement with a previous study (24), we showed that insulin induces
proliferation of RASMCs in a concentration-dependent manner, with a
10−5 M concentration exerting the strongest effect. We
further showed that insulin induces expression of ERK1/2 and
p-ERK1/2 in a time-dependent manner, and that this effect is the
strongest at 24 h of treatment. The present and previous studies
overall confirm that treatment with high concentrations of insulin
stimulates RASMC proliferation through activation of MAPKs.
Notably, we also observed a time-dependent stimulation of the
expression of JAK, p-JAK, STAT3 and p-STAT3 by high concentrations
of insulin in RASMCs. This result strongly suggests that the
JAK/STAT cell proliferation signal transduction pathway is also
regulated by high concentrations of insulin. Therefore, JAK/STAT
pathways not only mediate proliferation of VSMCs induced by IL-6
(19), platelet-derived growth
factor BB (30) and IL-18
(31), but may also have a role in
hyperinsulinism-induced atherosclerosis.
In the present study, RASMCs were pre-incubated with
RBP4 (1 or 4 μg/ml) for 1 h prior to treatment with 10−5
M insulin, and these experiments demonstrated that RBP4
significantly increases the ability of insulin to induce smooth
muscle cell proliferation. The percentage of RBP4-treated cells
that were detected at the S and G2 phases was considerably higher
than that of the control cells. This study also found that RBP4
significantly enhances the insulin-induced level of p-ERK and p-JAK
proteins, from which we can infer that the activation
(phosphorylation) of ERK and JAK is significantly enhanced. No
significant effect on STAT3 phosphorylation was observed under
these conditions, thus we conclude that RBP4 enhances
insulin-induced smooth muscle cell proliferation via the MAPK
pathway. Pre-treatment of RASMCs with the ERK1/2 inhibitor PD98059
effectively inhibited the observed effects of RBP4, including cell
cycle changes. However, treatment with the JAK inhibitor AG490 had
no significant effect on RBP4-induced cell proliferation, and cell
cycle distribution did not change. These findings argue for the
involvement of the MAPK pathway in the promotion of the
insulin-induced proliferation of RASMCs by RBP4.
Binding of insulin to the insulin receptor (InsR)
mediates, via a number of signal transduction pathways, a variety
of physiological effects in the target organs and tissues. There
are at least two insulin-related signal transduction pathways: one
is the phosphatidylinositide 3 kinase (PI-3K) pathway, which is
activated by IRS; the second is the MAPK pathway, which is
activated via the Grb2/SOS and RAS proteins. A high level of plasma
RBP4 can enhance insulin resistance through inhibition of IRS-1 and
activation of phosphatidylinositol 3-kinase (PI3-K) in skeletal
muscles (3). In agreement with a
study (22) reporting that RBP4
reduces insulin-induced phosphorylation of IRS-1 and ERK1/2 in
human adipose tissue, Takebayashi et al (23) found that RBP4 increases the
production of NO by stimulating part of the PI3K/Akt/eNOS pathway
in vascular endothelial cells, and leads to vasodilation through
inhibition of phosphorylation of ERK1/2 and insulin-induced
secretion of endothelin. However, in the present study, RBP4 was
shown to enhance phosphorylation of ERK1/2 in RASMCs, thereby
promoting the proliferation of smooth muscle cells, and this effect
was attenuated by the specific inhibitor of ERK1/2 PD98059. In
target tissues and organs of individuals with obesity and type 2
diabetes, the IRS/PI-3K pathway is impaired, but the Shc/Raf/MAPK
pathway remains intact, and is even stimulated (18). This ‘selective insulin resistance’
phenomenon mitigates insulin metabolic regulation, including its
antagonistic effect in atherosclerosis. RBP4 can inhibit
phosphorylation of IRS-1 (3,22),
then promote selective insulin resistance to enhance
insulin-induced proliferation of vascular smooth muscle cells, and
synthesis of collagen and growth factors.
In conclusion, the present study showed that insulin
induces proliferation of RASMCs in a concentration- and
time-dependent manner via the MAPK and JAK2/STAT3 pathways. In
addition, we report, for the first time to the best of our
knowledge, that RBP4 enhances RASMC proliferation induced by
insulin via activation of the MAPK signaling pathway. Since this is
a novel finding, additional research is needed to confirm it and
further investigate the underlying mechanisms. The results from the
present and previous studies indicate that RBP4 has a prominent
role as a modulator of atherosclerosis in hyperinsulinemia, and may
contribute to a better understanding of the numerous unexpected
aspects of CVDs.
References
|
1
|
Al-Harithy RN and Al-Ghamdi S: Serum
resistin, adiposity and insulin resistance in Saudi women with type
2 diabetes mellitus. Ann Saudi Med. 25:283–287. 2005.PubMed/NCBI
|
|
2
|
Bauche IB, Ait El Mkadem S, Rezsohazy R,
Funahashi T, Maeda N, Miranda LM and Brichard SM: Adiponectin
downregulates its own production and the expression of its AdipoR2
receptor in transgenic mice. Biochem Biophys Res Commun.
345:1414–1424. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang Q, Graham TE, Mody N, Preitner F,
Peroni OD, Zabolotny JM, Kotani K, Quadro L and Kahn BB: Serum
retinol binding protein 4 contributes to insulin resistance in
obesity and type 2 diabetes. Nature. 436:356–362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Graham TE, Yang Q, Bluher M, Hammarstedt
A, Ciaraldi TP, Henry RR, Wason CJ, Oberbach A, Jansson P, Smith U
and Kahn BB: Retinol-binding protein 4 and insulin resistance in
lean, obese, and diabetic subjects. N Engl J Med. 354:2552–2563.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stefan N, Hennige AM, Staiger H, Machann
J, Schick F, Schleicher E, Fritsche A and Häring HU: High
circulating retinol-binding protein 4 is associated with elevated
liver fat but not with total, subcutaneous, visceral, or
intramyocellular fat in humans. Diabetes Care. 30:1173–1178. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cho YM, Youn BS, Lee H, Lee N, Min SS,
Kwak SH, Lee HK and Park KS: Plasma retinol-binding protein-4
concentrations are elevated in human subjects with impaired glucose
tolerance and type 2 diabetes. Diabetes Care. 29:2457–2461. 2006.
View Article : Google Scholar
|
|
7
|
Ribel-Madsen R, Friedrichsen M, Vaag A and
Poulsen P: Retinol-binding protein 4 in twins: regulatory
mechanisms and impact of circulating and tissue expression levels
on insulin secretion and action. Diabetes. 58:54–60. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Klöting N, Graham TE, Berndt J, Kralisch
S, Kovacs P, Wason CJ, Fasshauer M, Schön MR, Stumvoll M, Blüher M
and Kahn BB: Serum retinol-binding protein is more highly expressed
in visceral than in subcutaneous adipose tissue and is a marker of
intra-abdominal fat mass. Cell Metab. 6:79–87. 2007.PubMed/NCBI
|
|
9
|
Despres JP, Lamarche B, Mauriege P, Cantin
B, Dagenais GR, Moorjani S and Lupien PJ: Hyperinsulinemia as an
independent risk factor for ischemic heart disease. N Engl J Med.
334:952–957. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pepe MG, Ginzton NH, Lee PD, Hintz RL and
Greenberg PL: Receptor binding and mitogenic effects of insulin and
insulin-like growth factors I and II for human myeloid leukemic
cells. J Cell Physiol. 133:219–227. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Faries PL, Rohan DI, Wyers MC, Marin ML,
Hollier LH, Quist WC and LoGerfo FW: Vascular smooth muscle cells
derived from atherosclerotic human arteries exhibit greater
adhesion, migration, and proliferation than venous cells. J Surg
Res. 104:22–28. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mahmoudi MJ, Mahmoudi M, Siassi F, Hedayat
M, Pasalar P, Chamari M, Abolhassani H, Rezaei N and Saboor-Yaraghi
AA: Circulating retinol-binding protein 4 concentrations in
patients with coronary artery disease and patients with type 2
diabetes mellitus. Int J Diabetes Dev Ctries. 32:105–110. 2012.
View Article : Google Scholar
|
|
13
|
Mallat Z, Simon T, Benessiano J, Clement
K, Taleb S, Wareham NJ, Luben R, Khaw KT, Tedgui A and Boekholdt
SM: Retinol-binding protein 4 and prediction of incident coronary
events in healthy men and women. J Clin Endocrinol Metab.
94:255–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsumoto K, Miyake S, Yano M, Ueki Y,
Yamaguchi Y, Akazawa S and Tominaga Y: Insulin resistance and
arteriosclerosis obliterans in patients with NIDDM. Diabetes Care.
20:1738–1743. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Giannattasio C and Mancia G: Arterial
distensibility in humans: modulating mechanisms, alterations in
diseases and effects of treatment. J Hypertens. 20:1889–1899. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Page C and Doubell AF: Mitogen-activated
protein kinase (MAPK) in cardiac tissues. Mol Cell Biochem.
157:49–57. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pessin JE and Saltiel AR: Signaling
pathways in insulin action: molecular targets of insulin
resistance. J Clin Invest. 106:165–169. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cusi K, Maezono K, Osman A, Pendergrass M,
Patti ME, Pratipanawatr T, DeFronzo RA, Kahn CR and Mandarino LJ:
Insulin resistance differentially affects the PI3-kinase- and MAP
kinase-mediated signaling in human muscle. J Clin Invest.
105:311–320. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Horvath CM: STAT proteins and
transcriptional responses to extracellular signals. Trends Biochem
Sci. 25:496–502. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Watanabe S, Mu W, Kahn A, Jing N, Li JH,
Lan HY, Nakagawa T, Ohashi R and Johnson RJ: Role of JAK/STAT
pathway in IL-6-induced activation of vascular smooth muscle cells.
Am J Nephrol. 24:387–392. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo F, Zarella C and Wagner WD: STAT4 and
the proliferation of artery smooth muscle cells in atherosclerosis.
Exp Mol Pathol. 81:15–22. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ost A, Danielsson A, Liden M, Eriksson U,
Nystrom FH and Stralfors P: Retinol-binding protein-4 attenuates
insulin-induced phosphorylation of IRS1 and ERK1/2 in primary human
adipocytes. FASEB J. 21:3696–3704. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takebayashi K, Sohma R, Aso Y and Inukai
T: Effects of retinol binding protein-4 on vascular endothelial
cells. Biochem Biophys Res Commun. 408:58–64. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang CC, Gurevich I and Draznin B: Insulin
affects vascular smooth muscle cell phenotype and migration via
distinct signaling pathways. Diabetes. 52:2562–2569. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ingelsson E, Sundström J, Melhus H,
Michaëlsson K, Berne C, Vasan RS, Risérus U, Blomhoff R, Lind L and
Ärnlöv J: Circulating retinol-binding protein 4, cardiovascular
risk factors and prevalent cardiovascular disease in elderly.
Atherosclerosis. 206:239–244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bobbert T, Raila J, Schwarz F, Mai K,
Henze A, Pfeiffer AF, Schweigert FJ and Spranger J: Relation
between retinol, retinol-binding protein 4, transthyretin and
carotid intima media thickness. Atherosclerosis. 213:549–551. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu J, Shi YH, Niu DM, Li HQ, Zhang CN and
Wang JJ: Association among retinol-binding protein 4, small dense
LDL cholesterol and oxidized LDL levels in dyslipidemia subjects.
Clin Biochem. 45:619–622. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
von Eynatten M, Lepper PM, Liu D, Lang K,
Baumann M, Nawroth PP, Bierhaus A, Dugi KA, Heemann U, Allolio B
and Humpert PM: Retinol-binding protein 4 is associated with
components of the metabolic syndrome, but not with insulin
resistance, in men with type 2 diabetes or coronary artery disease.
Diabetologia. 50:1930–1937. 2007.
|
|
29
|
Cabré A, Lázaro I, Girona J, Manzanares J,
Marimón F, Plana N, Heras M and Masana L: Retinol-binding protein 4
as a plasma biomarker of renal dysfunction and cardiovascular
disease in type 2 diabetes. J Intern Med. 262:496–503.
2007.PubMed/NCBI
|
|
30
|
Neeli I, Liu ZM, Dronadula N, Ma ZA and
Rao GN: An essential role of the Jak-2/STAT-3/cytosolic
phospholipase A(2) axis in platelet-derived growth factor
BB-induced vascular smooth muscle cell motility. J Biol Chem.
279:46122–46128. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sahar S, Dwarakanath RS, Reddy MA, Lanting
L, Todorov I and Natarajan R: Angiotensin II enhances
interleukin-18 mediated inflammatory gene expression in vascular
smooth muscle cells: a novel cross-talk in the pathogenesis of
atherosclerosis. Circ Res. 96:1064–1071. 2005. View Article : Google Scholar : PubMed/NCBI
|