Introduction
Adhering junctions, including adherens junctions and
desmosomes, are essential for cell unity in sheets of epithelial
cells (1). Generally, adhering
junctions are comprised of a transmembrane cadherin component,
which is involved in homophilic or heterophilic interactions in a
subclass-specific manner on the extracellular side and a variety of
cytoplasmic adapter proteins that, in turn, link cytoskeletal
structures to sites of cell-cell contact (2). The issue of how adhesive dysfunction
contributes to cancer biology has become an active area of
investigation in cancer research (3). One such protein is desmocollin-2
(DSC2), a transmembrane cadherin of the desmosomal cell-cell
adhesion structure. DSC2 is involved in the interaction of plaque
proteins and intermediate filaments mediating cell-cell adhesion,
and may also contribute to epidermal cell positioning (4).
The desmosomal cadherins family members mediate a
variety of biological processes, including cell growth,
invasiveness, adhesion and apoptosis (5–8).
With respect to tumorigenesis, DSC2 has been demonstrated to be
involved in the development of several types of tumor (9–14). A
reduction in the expression of DSC2 has been reported in numerous
types of human carcinoma, including colorectal, pancreatic,
gastric, lung and urothelial cancer (9–14).
It has been suggested that a reduction in the expression of DSC2
may act as an independent prognostic biomarker for reduced survival
rate in patients (13,14). A previous study of RNA interference
(RNAi) in transformed colonic epithelial cells revealed the
promotion of tumor cell proliferation in vitro and growth
in vivo following knock down of DSC2 (7).
In human esophageal squamous cell carcinoma (ESCC),
the expression of DSC2 has only recently been described (14). Our previous study demonstrated that
the expression of DSC2 in ESCC gradually decreases between regions
exhibiting esophageal hyperplasia to regions of dysplasia and
carcinoma in situ (14).
The depletion of DSC2 is highly associated with poor tumor
differentiation, regional lymph node metastasis and a poor
prognosis. Therefore, DSC2 may act as a new molecular marker in
predicting the prognosis of patients with ESCC. In addition, our
previous study also revealed that DSC2 has a causative effect in
esophageal cellular invasion and metastasis (6). The loss of DSC2 initiates tumor cell
metastasis by activating the β-catenin pathway and eventually
inducing an epithelial-mesenchymal transition-like process
(6). However, the contribution of
DSC2 to overall cell cohesion remains to be elucidated.
To investigate the possible role of DSC2 in
cell-cell adhesion, the present in vitro study was performed
based on the RNAi strategy in two ESCC cell lines, SHEEC and
KYSE510. The results supported our previous findings and the
proposal that DSC2 may be involved in the regulation of cell
invasion by a mechanism that controls cell-cell attachment and
cytoskeleton rearrangement. Altered DSC2 protein levels and
localization may, therefore, have several unexpected effects in
ESCC.
Materials and methods
Cell culture and transfection
The human esophageal squamous carcinoma cell lines
SHEEC and KYSE510 (Chinese Academy of Sciences, Shanghai, China)
were cultured in Dulbecco’s modified Eagle’s medium (Invitrogen
Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal
calf serum (Invitrogen Life Technologies). For siRNA transfection,
~5×104 cells/well were inoculated into 6-well plates,
cultured for 24 h and then transfected with the relevant siRNA (50
nM) using a Lipofectamine 2000 transfection reagent (Invitrogen
Life Technologies). The siRNA were synthesized by Shanghai
GenePharma Co., Ltd. (Shanghai, China) and contained two siRNAs
against human DSC2 (siDSC2-1 5′-CUGGAGAUGACAAAGUGUA-3′ and siDSC2-2
5′-CUUUACAGCUGCAAAUCUA-3′). A control siRNA oligonucleotide, not
matching any known human coding cDNA, was used as a control.
RNA extraction and reverse transcription
quantitative polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent
(Invitrogen Life Technologies) according to the manufacturer’s
instructions. Reverse transcription was performed using a total
volume of 20 μl with 1 μg total RNA using a Reverse Transcription
system (Promega Corporation, Madison, WI, USA). RT-qPCR was
performed on the Rotor-Gene 6000 system (Corbett Life Science,
Sydney, Australia). SYBR® Premix Ex Taq™ (Takara Bio,
Inc., Shiga, Japan) was used according to the manufacturer’s
instructions. The DSC2 PCR primers were designed based on the human
DSC2 mRNA sequence (GenBank accession no. NM_024422). The following
sequences were used: forward 5′-CCCAAGCTTGAAAAGCCCCTTGGATGAGA-3′
and reverse 5′-CGCGGATCCCCACTGGCTTTCAGAGACTT-3′. As an internal
control, a fragment of human β-actin was amplified using the
following primers: β-actin, forward 5′-CAACTGGGACGACATGGAGAAA-3′
and reverse 5′-GATAGCAACGTACATGGCTGGG-3′. The PCR conditions were
an initial denaturation step of 10 sec at 95°C, followed by 40
cycles consisting of 5 sec at 95°C, 20 sec at 60°C and 15 sec at
72°C.
Western blot analysis
Total cell lysates were prepared in
radioimmunoprecipitation assay buffer, separated by SDS-PAGE and
transferred onto polyvinylidene difluoride membranes (Millipore,
Billerica, MA, USA). The membranes were incubated in 10 ml of
blocking buffer [Tris-buffered saline containing 0.1% (v/v)
Tween-20 and 5% (w/v) non-fat dried milk powder (Sangon Biotech,
Shanghai, China)] for 1 h at room temperature and then incubated
with the indicated antibody. Finally, immunoreactive bands were
revealed using a luminol reagent (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA). Images were captured and quantitative
analyses were performed using FluorChem™ IS-8900 (Alpha Innotech
Co., San Leandro, CA, USA). The following antibodies were used:
Mouse monoclonal anti-DSC2 (7G6; Invitrogen Life Technologies),
mouse monoclonal anti-DSG2 (10G11; Progen Biotechnik GmbH,
Heidelberg, Germany), mouse monoclonal anti-PKP2 (PP2/62, PP2/86,
PP2/150; Progen Biotechnik GmbH) and mouse monoclonal anti-β-actin
(Sigma, St. Louis, MO, USA).
Confocal laser scanning microscopy
Staining was performed, as previously described
(6). The cells were fixed in 4%
paraformaldehyde solution (Thermo Fisher Scientific Inc., Rockford,
IL, USA) for 15 min, following which they were incubated with
donkey serum blocking buffer [Tris-buffered saline containing 5%
(w/v) normal donkey serum (Jackson ImmunoResearch, Hamburg,
Germany)] for 30 min and the primary antibody for 60 min.
Subsequently, the cells were incubated with donkey anti-mouse
polyclonal immunoglobulin G (IgG) and/or donkey anti-rabbit
polyclonal IgG (DyLight 488 and 594, respectively; Jackson
ImmunoResearch) for 60 min at room temperature. The following
antibodies were used: Mouse monoclonal anti-DSC2 (7G6; Invitrogen
Life Technologies), rabbit polyclonal anti-E-cadherin (Santa Cruz
Biotechnology, Inc.), mouse monoclonal anti-γ-catenin (Santa Cruz
Biotechnology, Inc.), mouse monoclonal anti-DSG2 (10G11; Progen
Biotechnik GmbH), mouse monoclonal anti-PKP2 (PP2/62, PP2/86,
PP2/150, Progen Biotechnik GmbH), mouse monoclonal
anti-pan-cytokeratin (Santa Cruz Biotechnology, Inc.) and
Acti-stain™ 555 Fluorescent Phalloidin (Cytoskeleton Inc., Denver,
CO, USA).
Hanging drop assay
Hanging drop cultures of aggregated cells were
generated from 1×103 cells. The cells were left
overnight to enable aggregation on the underside of a culture dish,
as previously described (15).
Following this, pipetting was performed 30 times on the resulting
cell clusters through a 200 μl Gilson pipette (Gilson Scientific
Ltd., Luton, UK). The degree of dissociation was then determined by
counting the number of particles present following trituration.
Dispase-based dissociation assay
The cell cultures were seeded in triplicate into
6-well plates. Subsequently, 24 h after reaching confluency, the
cultures were washed twice in phosphate-buffered saline (PBS) and
incubated in 1 ml dispase (2.4 U/ml; Sigma) for 30 min, as
previously described (15). The
monolayers liberated during this process were carefully washed two
times in PBS and then transferred into 15 ml conical tubes.
Additional PBS was added to reach a final volume of 2 ml. The tubes
were secured to a rocker and subjected to 50 inversion cycles. The
fragments were then counted using an inverted microscope
(Olympus).
Statistical analysis
All data are expressed as the mean ± standard
deviation and were analyzed using SPSS statistical software (SPSS
13.0; SPSS, Inc., Chicago, IL, USA). Comparisons between data sets
were performed using the χ2 test and a two-tailed
independent sample t-test when appropriate. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of DSC2 in ESCC cell
lines
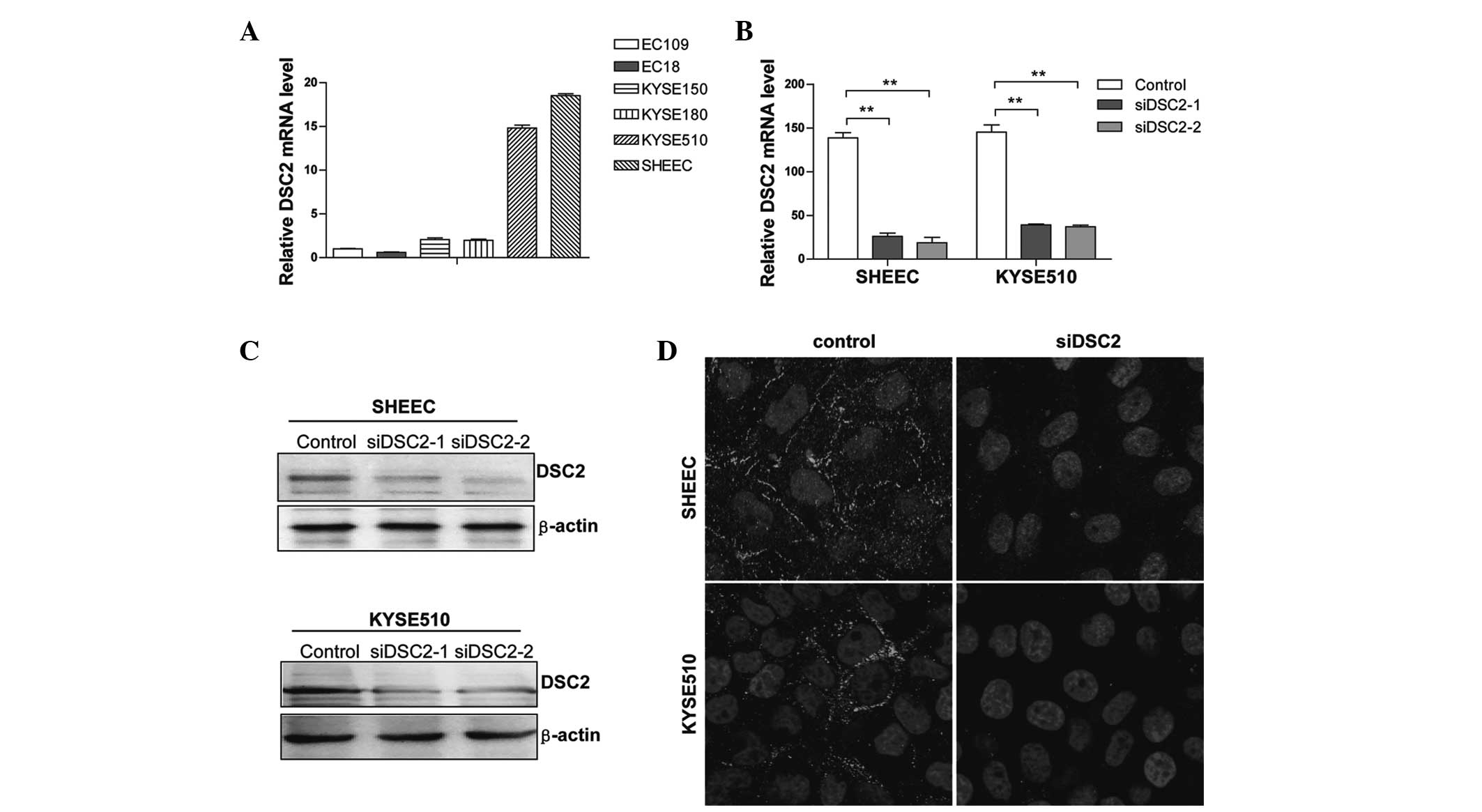
Initially, the mRNA expression levels of DSC2 were
examined in several ESCC cell lines by RT-qPCR. As shown in
Fig. 1A, DSC2 mRNA expression was
detected in all the cell lines evaluated, with SHEEC and KYSE510
cells containing the highest levels. Therefore, these cell lines
were selected for use as the model in the subsequent functional
studies.
DSC2 knockdown by transient siRNA
transfection in ESCC cells
Two double-stranded siRNAs (siRNA-1 and 2) targeting
the human DSC2 gene were synthesized. These sequences were specific
to DSC2 mRNA and matched no other genes in the NCBI nucleotide
database, in particular, they matched no other desmosomal cadherins
family members, in BLAST searching. RT-qPCR and western blot
analysis revealed that the DSC2 mRNA and protein expression
decreased markedly in the treated cells compared with the control
cells (Fig. 1B and C).
Transfection with DSC2 siRNAs resulted in a 70–80% reduction in the
expression of DSC2 in the SHEEC cells (Fig. 1C, upper panel) and a 60–70%
reduction in expression in the KYSE510 cells (Fig. 1C, lower panel), when compared with
those transfected with the scramble sequence. In addition, confocal
analysis of the cells labeled for DSC2 revealed a significant
reduction in the membrane localization of DSC2 in the cells treated
with siRNA (Fig. 1D).
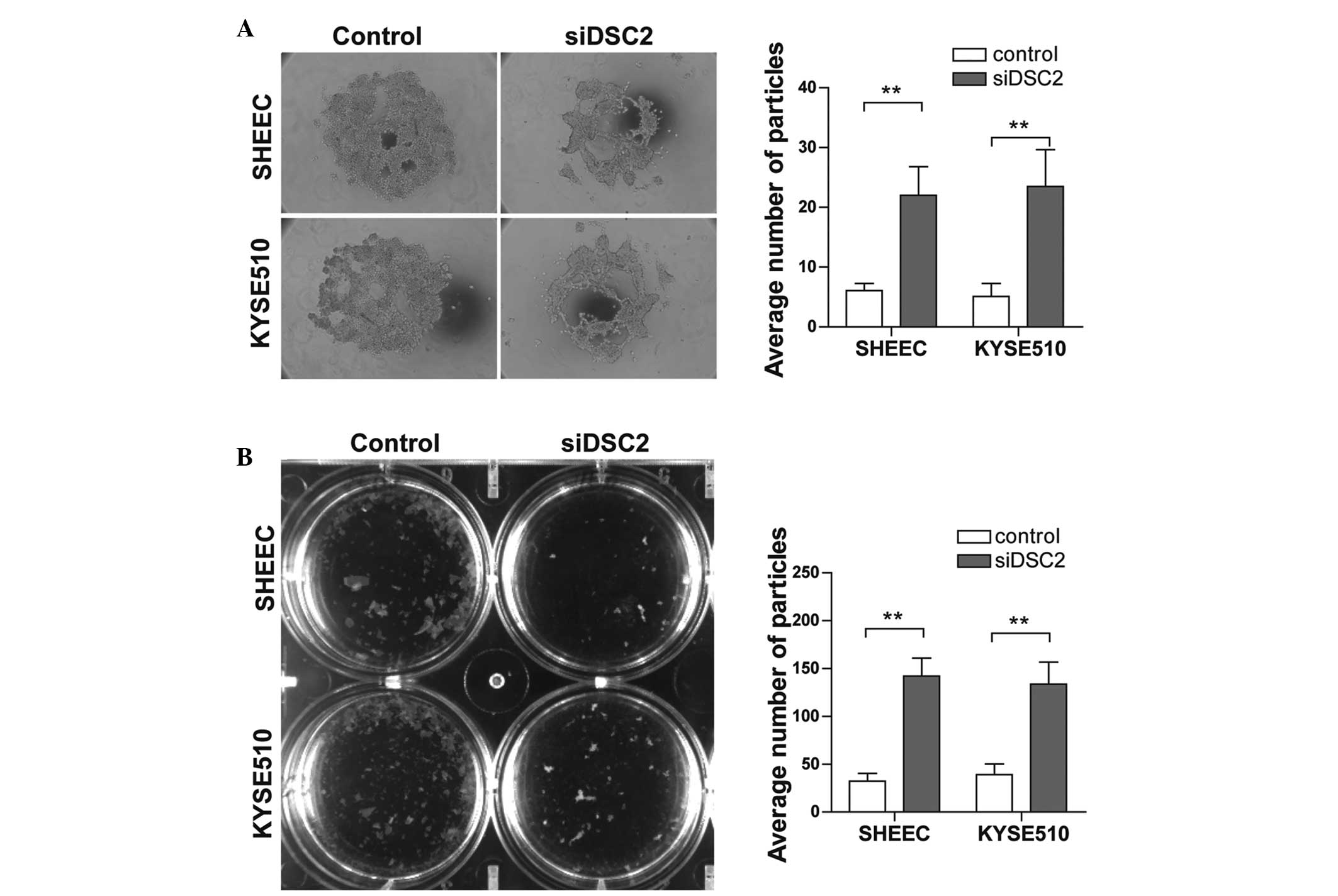
DSC2 silencing in ESCC cells compromises
cell-cell adhesion
To assess the cell-cell adhesion capacity of ESCC
cells following the suppression of DSC2, a hanging drop assay was
performed. After 24 h aggregation, in a hanging drop on the
underside of a culture dish, the cells were subjected to
trituration through a 200 μl Gilson pipette tip to disrupt
intercellular adhesion. The degree of dissociation was then
calculated by counting the particles that dissociated from the
original cluster. The results indicated a significant increase in
the number of particles and reduction in particle size in the
DSC2-depleted SHEEC and KYSE510 cells (Fig. 2A; P<0.01). To further confirm
this effect, a Dispase-based dissociation assay was also performed.
Confluent monolayers of siRNA-transfected SHEEC and KYSE510 cells
were harvested from tissue culture dishes by incubation with
dispase. The monolayers were then transferred into 15-ml conical
tubes and, following inversion of the tubes 50 times on a rocker,
the monolayer fragments were counted. As shown in Fig. 2B, following siRNA transfection for
2 days, the DSC2-knockdown cells were less cohesive and formed
smaller aggregates compared with the control cells (P<0.01).
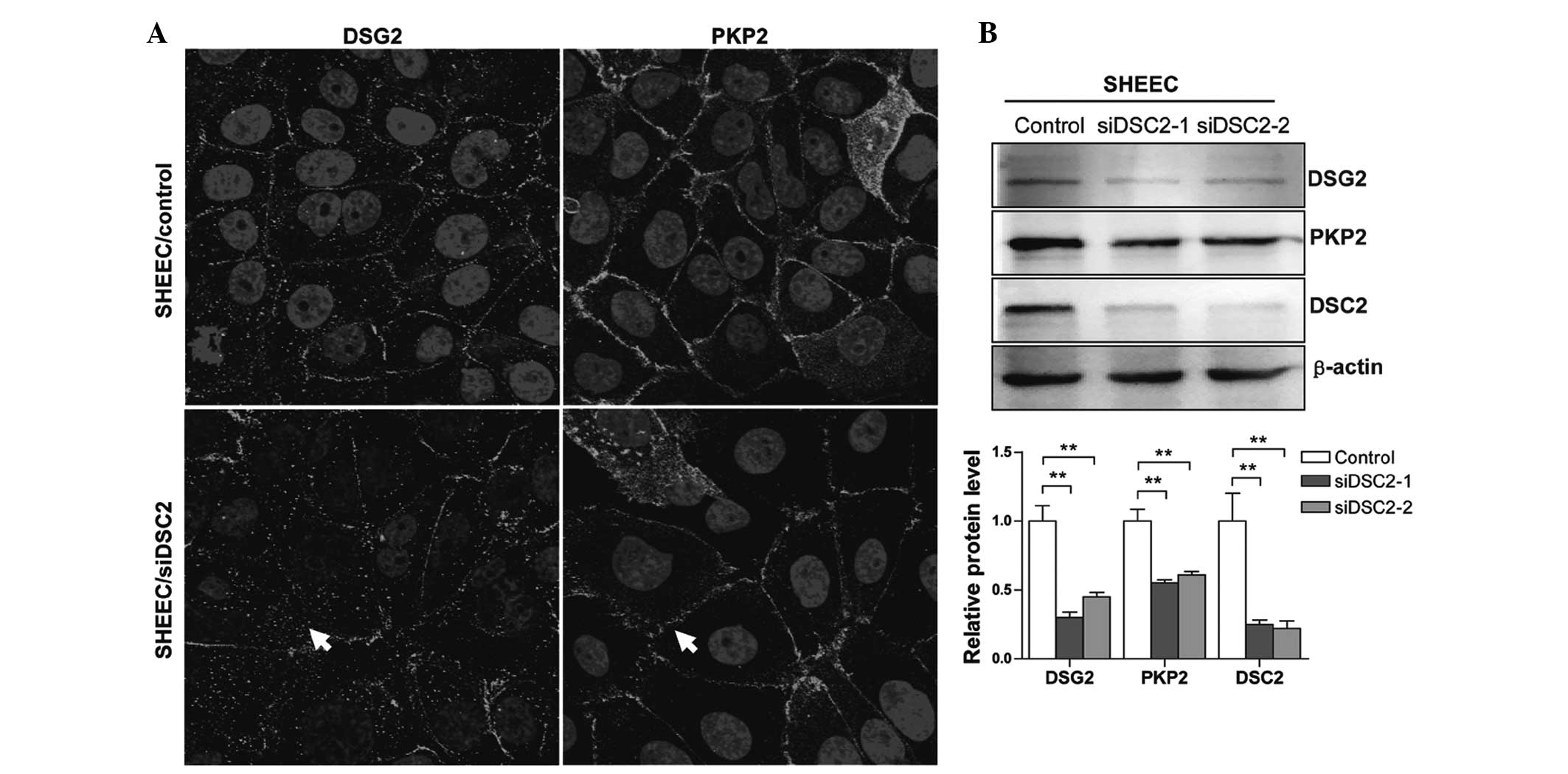
Effects of DSC2 depletion on desmosomal
proteins and adherens junction molecules
To assess the effects of RNAi-mediated DSC2
silencing on desmosome junctions in esophageal carcinoma cells, the
cells treated with either scrambled or DSC2-specific siRNA were
analyzed using fluorescence confocal microscopy to examine the
impact on desmosomal junction formation. As shown in Fig. 3A, marked staining for desmoglein-2
(DSG2) and plakophilin-2 (PKP2) was observed in the control cells
with localization at the cell membrane. Silencing DSC2 had a marked
effect on the membrane localization of these desmosomal proteins
and on desmosomal junction formation (Fig. 3A; arrows). To further confirm this
effect, the levels of desmosomal protein expression were examined
using western blot analysis. The protein expression levels of DSG2
and PKP2 were significantly reduced in the DSC2-specific
siRNA-transfected SHEEC cells (Fig.
3B). These results suggested that DSC2 silencing in ESCC cells
affects the expression and localization of desmosome proteins and
the formation of desmosomal junctions.
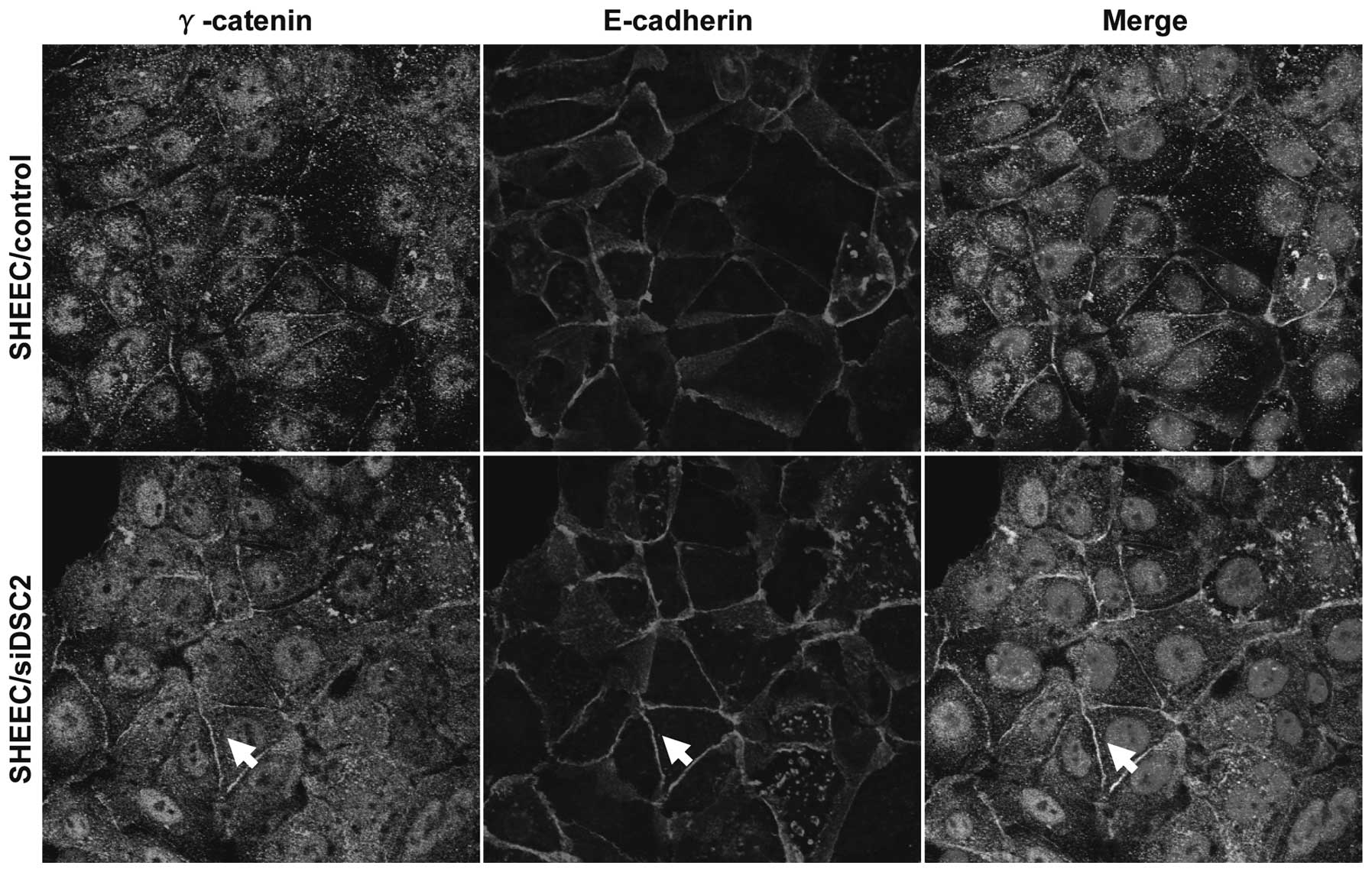
Desmosomes and adherens junctions have associated
effects in maintaining cellular adhesion and have similar
structural compositions. In addition, E-cadherin and DSC2 protein
bind directly to an armadillo family member, γ-catenin (16). Therefore, the present study
investigated the effects of DSC2 knock down on the localization of
γ-catenin and E-cadherin. As shown in Fig. 4, depletion of DSC2 resulted in
increased levels of free γ-catenin levels, promoting the
recruitment of γ-catenin to the adherens junction complex. This
contributes to our previous findings and supports the concept that
DSC2 may be involved in the regulation of cell invasive behavior by
a mechanism that controls cell-cell attachment, including adherens
junctions and desmosomes.
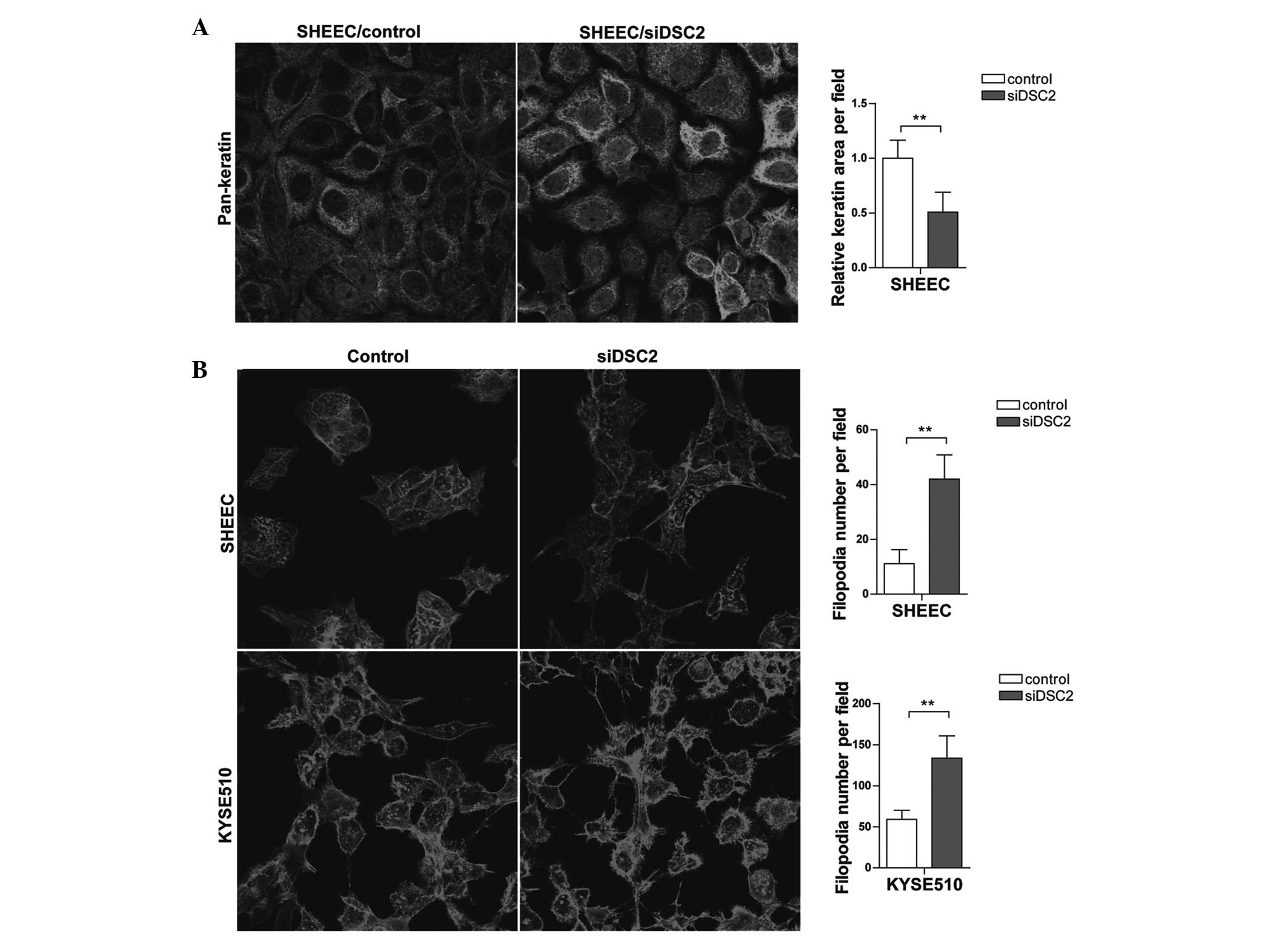
DSC2 depletion leads to keratin
intermediate filament retraction and filamentous (F)-actin
cytoskeleton rearrangement
Tumor metastasis requires the rearrangement of the
adhesive contacts of tumor cells to enable cells to migrate
relative to neighboring cells It has been suggested that, in this
process, actin filament and intermediate filament-based junctions
act synergistically to affect cell-cell adhesion (15). Therefore, the present study
investigated whether DSC2 depletion affects cytoskeleton
rearrangement. Immunofluorescence analysis revealed retracted
keratin intermediate filaments from the plasma membranes in
DSC2-specific siRNA-transfected SHEEC cells, whereas, in cells
transfected with the scramble sequence, the keratin intermediate
filaments lining the plasma membranes remained (Fig. 5A). Furthermore, phalloidin-coumarin
staining indicated that DSC2 depletion affected the F-actin
arrangement (Fig. 5B). Compared
with the control group, the cells with reduced DSC2 exhibited more
filopodia. These results indicated that DSC2 depletion resulted in
cytoskeleton rearrangement, ultimately promoting cell invasive
behavior.
Discussion
The present study is one of the first attempts, to
the best of our knowledge, to associate the expression of DSC2 with
cell-cell adhesion and cytoskeleton rearrangement in ESCC. Several
studies have found that DSC2 proteins are abnormally expressed in
various types of cancer and correlate with cell proliferation and
invasive behavior (6,7). However, fewer studies have focused on
investigating the role of DSC2 in regulating adhesive strength. In
the present study, an RNAi strategy was adopted to investigate
whether DSC2 has a role in the regulation of cell-cell adhesion in
ESCC cell lines. Using various approaches, the present study
demonstrated that knock down of DSC2 in these cell lines caused
defects in cell-cell adhesion, accompanied by reduced desmosomal
junction formation, retraction of keratin intermediate filaments
and F-actin cytoskeleton rearrangement.
Our previous studies have demonstrated that DSC2 has
a causative effect in cellular invasion and metastasis in
laboratory models and in clinical ESCC samples (6,14).
DSC2 depletion is highly associated with poor tumor
differentiation, regional lymph node metastasis and poor prognosis.
In the present study, the results revealed that DSC2 may be
involved in the regulation of cell invasive behavior by a mechanism
that controls cell-cell attachment, including adherens junctions
and desmosomes. As a junctional protein, DSC2 interacts
extracellularly with DSG2/3 to mediate cell-cell adhesion (17). It has been suggested that specific
desmosomal cadherins have a different contribution in cell-cell
adhesion (18). Consistent with
this observation, the data presented in the present study suggested
that a decreased level of DSC2 in ESCC cells leads to a decrease in
the expression of desmosomal proteins and desmosomal junction
formation, which was accompanied by a decrease in cell-cell
attachment. These results are consistent with data from Mannan
et al that DSG3 silencing in HaCaT cells causes disruption
of desmosome junctions and compromises cell-cell adhesion (19). This leads to the conclusion that
knocking down the expression of DSC2 by siRNA destabilizes other
desmosomal proteins. Although, it remains unclear whether this
represents transcriptional downregulation or increased protein
turnover, possibly complexed with DSC2, the reduction of cell
adhesion protein suggests that the inhibition of cell-cell adhesion
is partly dependent upon the interaction of DSC2 with other
desmosomal molecules.
Previous studies have revealed that adherens
junctions and desmosomes are dependant on each other for
appropriate assembly and maintenance (15,16).
In addition to their independent roles in tissue morphogenesis and
homeostasis, adherens junctions and desmosomes-based cell-cell
adhesive contacts engage in ‘cross-talk’ mechanisms, in which one
junction type affects the expression, assembly, turnover and/or
function of the other junctions or junction components. It has been
suggested that achieving maximal mechanical integrity of the
epithelial cell sheet requires proper junctional attachment to
intermediate filaments and cortical actin (15). Furthermore, it has been
demonstrated that the loss of desmosomal components leads to
defects in the maturation of adherens junctions and the associated
cortical actin cytoskeleton (20).
In comparison, the present study demonstrated that DSC2 suppresses
tumor cell-cell adhesion by opposing the localization of adherens
junction molecules and keratin intermediate filament retraction and
F-actin cytoskeleton rearrangement. DSC2 depletion from the
desmosome structure may compromise adhesive strength in several
ways. DSC2 may destabilize the desmosomal cadherin clusters formed
and affect the proper disposition of desmosomal cadherin
extracellular domains, as discussed previously. Another possibility
is that DSC2 anchors the armadillo repeat-containing proteins
γ-catenin and PKP2 that, in turn, recruit the keratin intermediate
filament cytoskeleton to sites of cell-cell contact. Uncoupling of
these connections results in increased levels of free γ-catenin and
PKP2. It has been suggested that PKP2 may functionally link the Ras
homolog (Rho)A and protein kinase C-dependent pathways to drive
actin reorganization and to regulate the assembly of desmosomes
(21,22). PKP2 may provide a structural link
between junctional components and the actin and the intermediate
filament cytoskeletons to coordinate their activities during
assembly of junctions. In addition, γ-catenin is required for
effective anchorage of intermediate filaments to desmosomes
(23). A study by Todorović et
al reported that γ-catenin regulates cell motility through
Rho-and fibronectin-dependent Src signaling (24). Therefore, in esophageal carcinoma
cells, DSC2 may affect cytoskeleton rearrangement via alternative
signaling pathways, which requires extensive investigation to
determine.
Taken together, the present study suggests that DSC2
is important in the regulation of cell-cell adhesion dependent
motility. Depletion of the transmembrane desmosomal cadherin
components is likely to occur by destabilizing other desmosomal
proteins. Uncoupling of these connections resulted in increased
levels of free desmosomal plaque proteins. This in turn affected
cytoskeleton arrangement, possibly through alternative signaling
pathways, which ultimately led to uncontrolled migration of tumor
cells lacking DSC2.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (no. 81101613), the Guangdong
Provincial Natural Science Foundation of China (no.
S2011040004363), the Specialized Research Fund for the Doctoral
Program of Higher Education of China (no. 20114402120005) and the
Natural Science Foundation of China-Guangdong Joint Fund (no.
U0932001).
References
|
1
|
Green KJ, Getsios S, Troyanovsky S and
Godsel LM: Intercellular junction assembly, dynamics, and
homeostasis. Cold Spring Harb Perspect Biol. 2:a0001252010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Angst BD, Marcozzi C and Magee AI: The
cadherin superfamily: diversity in form and function. J Cell Sci.
114:629–641. 2001.PubMed/NCBI
|
|
3
|
Jeanes A, Gottardi CJ and Yap AS:
Cadherins and cancer: how does cadherin dysfunction promote tumor
progression? Oncogene. 27:6920–6929. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nuber UA, Schäfer S, Schmidt A, Koch PJ
and Franke WW: The widespread human desmocollin Dsc2 and
tissue-specific patterns of synthesis of various desmocollin
subtypes. Eur J Cell Biol. 66:69–74. 1995.PubMed/NCBI
|
|
5
|
Chen YJ, Chang JT, Lee L, Wang HM, Liao
CT, Chiu CC, Chen PJ and Cheng AJ: DSG3 is overexpressed in head
neck cancer and is a potential molecular target for inhibition of
oncogenesis. Oncogene. 26:467–476. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fang WK, Liao LD, Li LY, Xie YM, Xu XE,
Zhao WJ, Wu JY, Zhu MX, Wu ZY, Du ZP, Wu BL, Xie D, Guo MZ, Xu LY
and Li EM: Down-regulated desmocollin-2 promotes cell
aggressiveness through redistributing adherens junctions and
activating beta-catenin signalling in oesophageal squamous cell
carcinoma. J Pathol. 231:257–270. 2013. View Article : Google Scholar
|
|
7
|
Kolegraff K, Nava P, Helms MN, Parkos CA
and Nusrat A: Loss of desmocollin-2 confers a tumorigenic phenotype
to colonic epithelial cells through activation of
Akt/{beta}-catenin signaling. Mol Biol Cell. 22:1121–1134. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dusek RL, Getsios S, Chen F, Park JK,
Amargo EV, Cryns VL and Green KJ: The differentiation-dependent
desmosomal cadherin desmoglein 1 is a novel caspase-3 target that
regulates apoptosis in keratinocytes. J Biol Chem. 281:3614–3624.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khan K, Hardy R, Haq A, Ogunbiyi O, Morton
D and Chidgey M: Desmocollin switching in colorectal cancer. Br J
Cancer. 95:1367–1370. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Anami K, Oue N, Noguchi T, Sakamoto N,
Sentani K, Hayashi T, Hinoi T, Okajima M, Graff JM and Yasui W:
Search for transmembrane protein in gastric cancer by the
Escherichia coli ampicillin secretion trap: expression of
DSC2 in gastric cancer with intestinal phenotype. J Pathol.
221:275–284. 2010.PubMed/NCBI
|
|
11
|
Cui T, Chen Y, Yang L, Mireskandari M,
Knösel T, Zhang Q, Kohler LH, Kunze A, Presselt N and Petersen I:
Diagnostic and prognostic impact of desmocollins in human lung
cancer. J Clin Pathol. 65:1100–1106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hayashi T, Sentani K, Oue N, Anami K,
Sakamoto N, Ohara S, Teishima J, Noguchi T, Nakayama H, Taniyama K,
Matsubara A and Yasui W: Desmocollin 2 is a new immunohistochemical
marker indicative of squamous differentiation in urothelial
carcinoma. Histopathology. 59:710–721. 2011. View Article : Google Scholar
|
|
13
|
Hamidov Z, Altendorf-Hofmann A, Chen Y,
Settmacher U, Petersen I and Knösel T: Reduced expression of
desmocollin 2 is an independent prognostic biomarker for shorter
patients survival in pancreatic ductal adenocarcinoma. J Clin
Pathol. 64:990–994. 2011. View Article : Google Scholar
|
|
14
|
Fang WK, Gu W, Li EM, Wu ZY, Shen ZY, Shen
JH, Wu JY, Pan F, Lv Z, Xu XE, Huang Q and Xu LY: Reduced
membranous and ectopic cytoplasmic expression of DSC2 in esophageal
squamous cell carcinoma: an independent prognostic factor. Hum
Pathol. 41:1456–1465. 2010. View Article : Google Scholar
|
|
15
|
Huen AC, Park JK, Godsel LM, Chen X,
Bannon LJ, Amargo EV, Hudson TY, Mongiu AK, Leigh IM, Kelsell DP,
Gumbiner BM and Green KJ: Intermediate filament-membrane
attachments function synergistically with actin-dependent contacts
to regulate intercellular adhesive strength. J Cell Biol.
159:1005–1017. 2002. View Article : Google Scholar
|
|
16
|
Lewis JE, Wahl JK 3rd, Sass KM, Jensen PJ,
Johnson KR and Wheelock MJ: Cross-talk between adherens junctions
and desmosomes depends on plakoglobin. J Cell Biol. 136:919–934.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garrod D and Chidgey M: Desmosome
structure, composition and function. Biochim Biophys Acta.
1778:572–587. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hartlieb E, Kempf B, Partilla M, Vigh B,
Spindler V and Waschke J: Desmoglein 2 is less important than
desmoglein 3 for keratinocyte cohesion. PLoS One. 8:e537392013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mannan T, Jing S, Foroushania SH, Fortune
F and Wan H: RNAi-mediated inhibition of the desmosomal cadherin
(desmoglein 3) impairs epithelial cell proliferation. Cell Prolif.
44:301–310. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vasioukhin V, Bowers E, Bauer C,
Degenstein L and Fuchs E: Desmoplakin is essential in epidermal
sheet formation. Nat Cell Biol. 3:1076–1085. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bass-Zubek AE, Hobbs RP, Amargo EV, Garcia
NJ, Hsieh SN, Chen X, Wahl JK 3rd, Denning MF and Green KJ:
Plakophilin 2: a critical scaffold for PKC alpha that regulates
intercellular junction assembly. J Cell Biol. 181:605–613. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Godsel LM, Dubash AD, Bass-Zubek AE,
Amargo EV, Klessner JL, Hobbs RP, Chen X and Green KJ: Plakophilin
2 couples actomyosin remodeling to desmosomal plaque assembly via
RhoA. Mol Biol Cell. 21:2844–2859. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Acehan D, Petzold C, Gumper I, Sabatini
DD, Müller EJ, Cowin P and Stokes DL: Plakoglobin is required for
effective intermediate filament anchorage to desmosomes. J Invest
Dermatol. 128:2665–2675. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Todorović V, Desai BV, Patterson MJ,
Amargo EV, Dubash AD, Yin T, Jones JC and Green KJ: Plakoglobin
regulates cell motility through Rho- and fibronectin-dependent Src
signaling. J Cell Sci. 123:3576–3586. 2010.PubMed/NCBI
|