Introduction
Streptococcus suis is an important emerging
human threat that can cause severe systemic infection (1–3).
Since the first reported cases of human infection by S. suis
in 1968, the number of cases has significantly increased in the
past few years, reaching ~1,000 cases by now, with the majority
occurring in Southeast Asia. S. suis infections are the
third most common cause of community-acquired bacterial meningitis
in Hong Kong, and the leading cause of adult meningitis in Vietnam
(4–5).
The S. suis serotype 2 (SS2) is the most
common cause of the disease in humans, although serotypes 1, 4, 14
and 16 have also been reported to cause severe disease in a limited
number of individuals. Most previous studies have concerned
sporadic cases of S. suis infection, but two Chinese
outbreaks (in 1998 and 2005) that involved >200 cases and 50
deaths, emphasized the importance of S. suis as an emerging
zoonotic pathogen (6,7). The most important feature of these
outbreaks was the high incidence of systemic disease, the
proportionally few cases of meningitis, and the high mortality.
Epidemiological surveys have indicated that all patients had a
history of close contact with diseased pigs and pork-derived
products, and that the emergence of the highly virulent SS2 strains
played a key role in the severe outbreaks (8,9).
The current understanding of the pathogenesis of
S. suis remains limited. Whole-genome sequencing and
comparative genomic analysis (10)
revealed that a DNA fragment of ~89 kb, designated as 89K, is
present in the two Chinese strains (98HAH12 and 05ZYH33), and
absent in the P1/7, which is a widely epidemic virulent
Streptococcus suis serotype 2 strain. The 89K fragment was
proposed to be a pathogenicity island, and genetic studies
indicated that disruption of the Salk/SalR two-component system
(TCS) inside the 89K considerably attenuates the virulence of the
pathogen, whereas functional complementation restores virulence in
infection experiments of piglets (11). A recent study (12) showed that 89K can spontaneously
excise to form an extrachromosomal circular product, and the 89K
excision intermediate can act as a substrate for lateral transfer
to non-89K SS2 recipients via a genomic island type IV secretion
system (T4SS) encoded in 89K. The authors proposed that these
genetic events are important for the emergence, pathogenesis and
persistence of epidemic SS2 strains. Based on sequence typing (ST),
it was also suggested that ST7 strains are prevalent in China and
possess a stronger capacity to stimulate T cells, naive T cells and
peripheral blood mononuclear cell proliferation compared to ST1
strains (13–15). The authors of these studies
proposed a two-stage hypothesis to explain streptococcal toxic
shock syndrome in the Chinese outbreaks. However, both hypotheses
need to be further investigated in the future.
Pore-forming toxins (PFTs) are the most prevalent
virulence factors produced by disease-causing bacteria, and are
required for the virulence of numerous important human pathogens,
including Staphylococcus aureus, S. pyogenes,
Clostridium perfringens and Aeromonas hydrophilia
(16). A PFT produced by S.
suis known as suilysin (sly) has been recognized as a virulence
factor owing to its toxicity to the host epithelial cells,
endothelial cells and macrophages (17–19).
The sly protein belongs to a family of thiol-activated toxins and
has a molecular weight of 54 kDa. It is a cholesterol-dependent
cytolysin related to the streptolysin O of S. pyogenes and
to pneumolysin of S. pneumoniae. Immunization to sly
provides protection against lethal challenge with a serotype 2
strain in both mice and pig modelss (20,21).
However, the expression profile of sly in certain S. suis
populations and its role in causing invasive infections have not
been characterized. In this study, we found that increased
production of sly in the S. suis strain 05ZYH33 is
associated with enhanced severity of S. suis infections and
thus, may contribute to the translocation of the pathogen across
the epithelial barrier. These results indicate that enhanced sly
production may play an important role during S. suis
invasive infections.
Materials and methods
Ethics
CD1 mice were obtained from the Experimental Animal
Centre of the Academy of Military Medical Sciences (Beijing,
China). Animal welfare and experimental procedures were approved by
the Academy of Military Medical Sciences Animal Care and Use
Committee, and were conducted in strict accordance with the Guide
for the Care and Use of Laboratory Animals (National Research
Council of USA, 1996). Efforts were made to minimize animal
suffering and to reduce the number of animals used. For the
survival experiments, the decisions were made following the
relevant guidelines of the Organisation for Economic Co-operation
and Development [OECD Environmental Health and Safety Publications,
Series on Testing and Assessment no. 19, ENV/JM/MONO(2000)7].
Bacterial strains and growth
conditions
The bacterial strains used in this study are listed
in Table I. S. suis was
grown in Difco™ Todd-Hewitt broth (THB; Becton, Dickinson and
Company, Franklin Lakes, NJ, USA) at 37°C. For antibiotic
selection, 5 μg/ml of chloramphenicol (Cm) or 100 μg/ml
spectinomycin (Spc) were used.
 | Table IBacterial strains and plasmids used
in this study. |
Table I
Bacterial strains and plasmids used
in this study.
| Material | Description | Area
(year)-Origin | Status | Source/Ref. |
|---|
| Streptococcus
suis strains |
| Δsly | In frame deletion
of sly in 05ZYH33 | Beijing, China
(2011) - Our lab | Mutant | This study |
| 1330 |
Sly−89K− | Canada-HPL | Avirulent | (25) |
| 05ZYH33 |
Sly+89K+ | Sichuan, China
(2005)-HP | Epidemic | This study |
| 98001 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 99001 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 98003 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 98005 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 98012 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 98015 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 98242 |
Sly+89K+ | Jiangsu, China
(1998)-HP | Epidemic | This study |
| 4 |
Sly+89K+ | Sichuan, China
(2005)-DP | Epidemic | This study |
| 5 |
Sly+89K+ | Sichuan, China
(2005)-DP | Epidemic | This study |
| SUN |
Sly+89K+ | Jiangsu, China
(2006)-HP | Epidemic | This study |
| 606 |
Sly+89K− | China
(1980)-DP | Non-epidemic | This study |
| 607 |
Sly+89K− | Japan-DP | Non-epidemic | This study |
| 1940 |
Sly+89K− | China
(1980)-DP | Non-epidemic | This study |
| 1941 |
Sly+89K− | China
(1980)-DP | Non-epidemic | This study |
| NJ |
Sly+89K− | Jiangsu,
China-DP | Non-epidemic | This study |
| 4005 |
Sly+89K− | The
Netherlands-DP | Non-epidemic | (26) |
| s735 |
Sly+89K− | The
Netherlands-DP | Non-epidemic | (27) |
| T15 |
Sly+89K− | Europe-HPL | Non-epidemic | (28) |
| Plasmids |
| pCR2.1 | na | na | na | Invitrogen™ |
| pSET4s | na | na | na | (29) |
Cell culture and lactate dehydrogenase
(LDH) release assay
Human umbilical vein endothelial cells (HUVECs;
Institute of Biochemistry and Cell Biology, Shanghai, China) were
cultured in RPMI-1640 medium containing 10% fetal bovine serum
(FBS). Cells (200 μl) were added at a density of 5×104
cells/ml into 96-well culture plates. S. suis strains
05ZYH33 or 1940 at the mid-log phase of growth were resuspended in
cell culture medium and were added to the cell culture wells at
multiplicity of infection (MOI) of 100. The amount of released
lactate dehydrogenase (LDH) from HUVECs was calculated with an LDH
release assay kit (Promega, Madison, WI, USA) after 4 h of
incubation.
Targeted mutagenesis of the sly gene
Polymerase chain reaction (PCR) was used to generate
an in-frame substitution of the sly gene with the Cm
gene using a previously described method (22). Briefly, 644 and 590 bp immediately
upstream and downstream of the sly coding sequence
(SSU05_1403) were amplified with the primer pairs
sly_upF/_upR and sly_downF/_downR, respectively. The
Cm gene was amplified using the primer pair Cm_F/_R
(Table II). The PCR products were
cloned into the Invitrogen™ pCR2.1 plasmid (Thermo Fisher
Scientific, Waltham, MA, USA), digested using restriction enzymes
and subcloned into the plamsid pSET4s one by one. Allelic exchange
mutagenesis in S. suis 05ZYH33 was performed as previously
described (22) to generate the
mutant Δsly. Allelic replacement of sly with
Cm in the S. suis chromosome was confirmed by PCR and
sequence analysis using the Phusion High-Fidelity DNA Polymerase
reagents (New England Biolabs, Ipswich, MA, USA) on a ABI 3730XL
machine (Applied Biosystems, Carlsbad, CA, USA).
| Table IIOligonucleotide primer sequences used
in this study. |
Table II
Oligonucleotide primer sequences used
in this study.
| Primer | Sequence
(5′-3′) | Restriction
enzyme |
|---|
| Sly_upF | GGGGGAAGCTTCTAGTCGGGGGAGTTTTTGTG | HindIII |
| Sly_upR | GGGGGGGTCGACTTAATATCTTGTTTGGAATCTG | SalI |
| Cm_F | GCGGTCGACTAATTCGATGGGTTCCGAGG | SalI |
| Cm_R | CGCGGATCCCACCGAACTAGAGCTTGATG | BamHI |
| Sly_downF | GGGGGGATCCCATGGGAGTGGTGGAGAACAGT | BamHI |
| Sly_downR | GGGGGGGGAATTCTTGGCCCGAATACCGACAG | EcoRI |
Enzyme-linked immunosorbent assay (ELISA)
assay
The sly level in the supernatant of different S.
suis strain cultures was determined using a double antibody
sandwich ELISA assay. The antibodies used were polyclonal. Briefly,
purified rabbit anti-sly IgG (1:5,000 dilution) was used to coat
the wells of a microtiter plate. Purified rat anti-sly IgG
(1:5,000) was used as the primary antibody, peroxidase-conjugated
goat anti-rat antibody (1:5,000) was used as the secondary
antibody, and K-Blue was used as the substrate (all from Santa Cruz
Biotechnology, Santa Cruz, CA, USA). After 10 min of incubation at
room temperature, the reaction was terminated with addition of 2 N
H2SO4, and the optical density was measured
at 450 nm (OD450) using the SpectraMax®
Plus384 Absorbance Microplate reader (Molecular Devices LLC,
Sunnyvale, CA, USA). Purified sly protein (23) was used to establish the standard
curve.
Hemolytic activity assay
The haemolytic activity assay was performed using a
previously described method (24).
Briefly, the culture supernatant of S. suis strains was
diluted 2-fold in phosphate-buffered saline (PBS). An equal amount
of human red blood cells (RBCs), obtained from the Department of
Hematology (Hospital 307 of Chinese People’s Liberation Army,
Beijing China) and washed twice in PBS, was added to 0.5 ml of each
dilution (final concentration of RBCs, 1.4%). Following incubation
for 1 h at 37°C, the mixtures were sedimented by centrifugation
(1,500 × g for 10 min), and the supernatants were transferred to
microplates. Following correction with the controls, which lacked
either haemolysin or erythrocytes, the OD540 of the
supernatant of each dilution was measured on a SpectraMax Plus
Absorbance Microplate reader.
Reverse transcription-quantitative-PCR
(RT-qPCR) analysis
The RT-qPCR analysis was performed with a sly
gene-specific primer set (forward, ACTTACGAGCCACAAGAGATTC and
reverse, GCAGCCTTAGCATCAATAACAG) on cDNA from the S. suis
strains 05ZYH33 and 1940. The assays were carried out in triplicate
using reagents of the SYBR Premix Ex Taq™ mastermix (Takara Bio,
Co., Ltd., Shiga, Japan) on an Opticon 2 system (MJ Research,
Waltham, MA, USA) using RNA isolated from three independent
cultures for each strain. RNA was extracted using the TRIzol RNA
Reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) following
the manufacturer’s instructions. The PCR cycling conditions were as
follows: 95°C for 10 min followed by 40 cycles of 95°C for 15 sec
and 60°C for 1 min. The gene encoding the 16S rRNA was used as an
internal control. The Ct values were normalized to the average Ct
(30) and the mean fold-changes
for the target genes were calculated as described by Livak and
Schmittgen (31).
Polymorphonuclear neutrophil (PMN)
phagocytosis assay
The PMN phagocytosis assay was performed as
previously described (32). Human
PMNs were isolated from healthy human blood as follows: blood was
incubated for 40 min at room temperature in a ratio of 3:1 ratio of
6.0% Dextran to sediment erythrocytes. Ficoll, 70% percoll and
blood supernatant were combined (1:1:1) and centrifuged at 400 × g
for 30 min at room temperature to separate PMNs. The PMNs were
aspirated in to a new cuvette, resuspended in 1640-RPMI, and
enumerated by microscopy. Briefly, 1 ml of human PMNs
(107 cells) were mixed with an equal number of
pre-opsonized S. suis colonies and incubated for 15 min at
37°C under continuous rotation to allow phagocytosis. When needed,
different concentrations of recombinant sly protein (23) were added (50–500 ng). To quantify
the intracellular bacteria, extracellular S. suis were
killed by incubating with 100 μg/ml gentamicin and 5 g/ml
penicillin for 1 h at 4°C. Subsequently, the cells were washed
twice with PBS and lysed for 15 min at room temperature in 400 μl
of 1% saponin. The viable bacteria were counted on an inverted IX81
microscope (Olympus, Tokyo, Japan) after plating serial dilutions
of the supernatants on THB agar plates.
Mouse infections
CD1 female mice (6-week-old) were chosen to evaluate
the virulence of the S. suis strains 05ZYH33 and 1940.
Briefly, mice (n=10 per group) were intraperitoneally injected with
108 living colony forming units (CFUs) of S. suis
strains 05ZYH33 and 1940 in 0.1 ml sterile saline. The mortality of
mice infected with these bacterial strains was recorded at 4-h
intervals for 32 h. To count the bacteria present in the blood,
blood samples were collected from the tail vein and plated onto THB
agar to accurately determine the viable bacteria at 5 h
post-infection. To assess the necrosis of PMNs, we sacrificed the
mice and injected 5 ml of RPMI-1640 medium containing 10% FBS into
the abdominal cavities. Then, mice were surgically opened and 4-ml
exudates were collected with 23-G needles. The exudates were
centrifuged at 500 × g for 5 min, and cell pellets were resuspended
in 100 ml of staining buffer (PBS containing 1% goat serum). The
samples were stained either with fluorescein isothiocyanate
(FITC)-conjugated anti-mouse anti-Ly-6G (a neutrophil marker), or
with the appropriate isotype control antibody (eB149/10H5; both
from eBiosciences, San Diego, CA, USA). Propidium iodide (0.5
mg/ml; BD Biosciences, San Jose, CA, USA) was used to determine the
cell viability. Samples were analyzed on a FACSCalibur flow
cytometer and the necrotic PMNs were quantified with the CellQuest
software (both from BD Biosciences). Data were obtained from 30,000
cells for each sample.
CD1 female mice (6-week-old, ~22 g) were chosen to
evaluate the virulence of the S. suis strain 05ZYH33 and the
Δsly mutant. Mice (n=10, per group) were intraperitoneally
injected with 0.5 ml (~5×108 CFUs) of S. suis at
the late-exponential growth phase in THB medium. The mortality of
the infected mice was recorded for 50 h, and the bacteria in the
blood were counted at different time-points post-infection. To
histopathologically assess the severity of changes in the abdominal
wall tissues, mice were surgically opened 6 h after injection. The
abdominal wall tissues were fixed in 10% Formaldehyde (Sigma, St.
Louis, MO, USA), sectioned and stained with hematoxylin and eosin.
The slides were observed and photographed using a BX53 fluorescence
microscope (Olympus, Tokyo, Japan) equipped with a DP72 CCD
camera.
Statistical analysis
Statistical analysis was performed using the SPSS
13.0 version software (SPSS Inc., Chicago, IL, USA). Statistical
significance of differences was assessed with Student’s t-tests,
with a p-value <0.05 considered to indicate statistically
significant differences.
Results
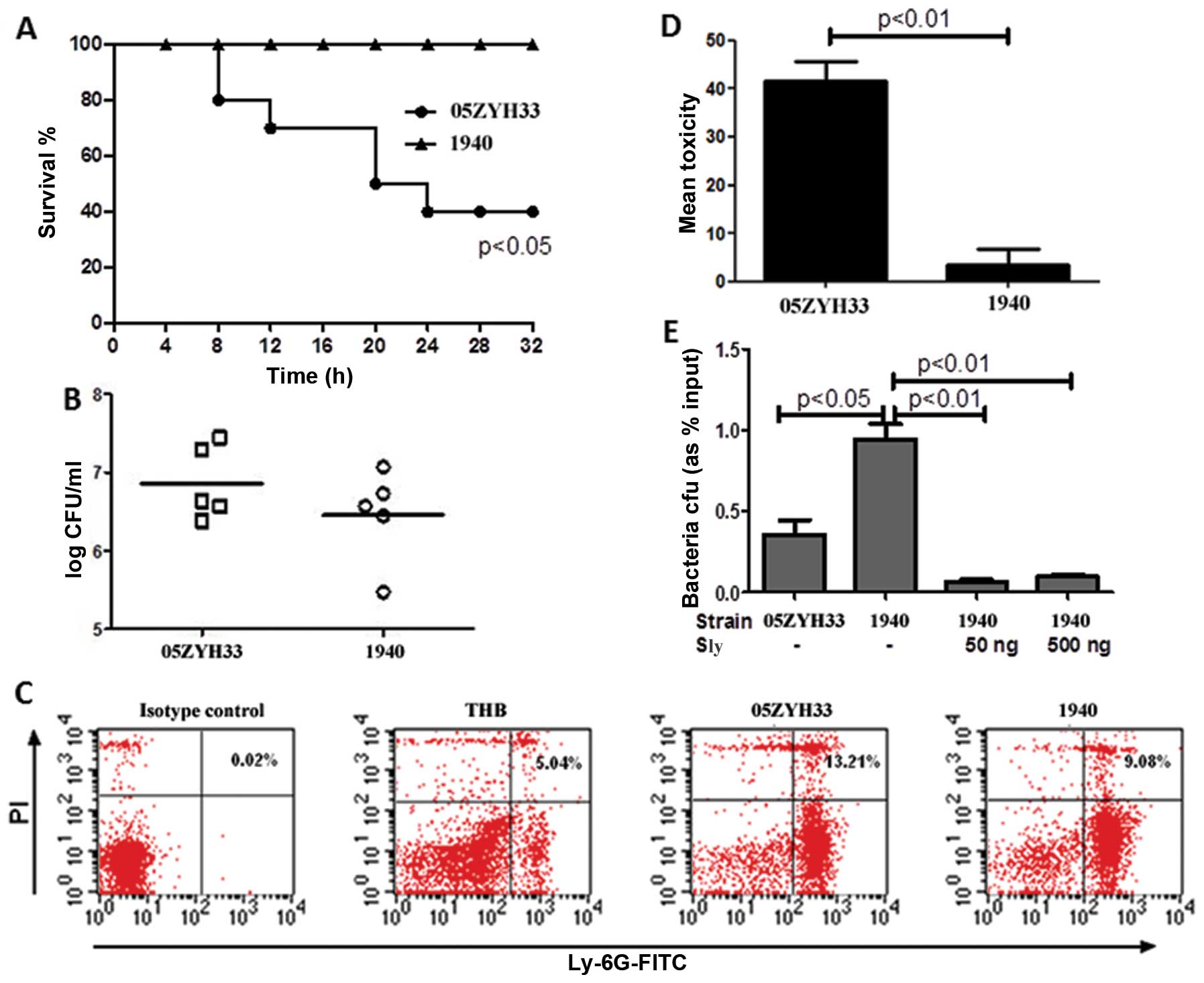
The S. suis epidemic strain is more
virulent than the non-epidemic strain
The S. suis epidemic strains are expected to
show high virulence, and need to resist host defence mechanisms and
be able to invade the epithelial or the endothelial barrier in
order to cause successful infection. To evaluate the potential
differences in virulence between S. suis epidemic and
non-epidemic strains, CD1 female mice were intraperitoneally
injected with 108 CFUs of the S. suis strains
05ZYH33 (epidemic, ST7) or 1940 (non-epidemic) in 0.1 ml sterile
saline. The strain 05ZYH33 caused significantly higher mortality
(p<0.05) than the 1940 strain (Fig.
1A). The bacterial concentration in the blood of the infected
mice also appeared different at 5 h post-infection, although the
difference was not statistically significant (Fig. 1B). In addition, S. suis
05ZYH33 caused higher necrosis of PMNs (Fig. 1C), the major cell type mediating
acute inflammatory responses to bacterial infections, compared to
S. suis 1940 (13.21±3.11 vs. 9.08±2.56%). Next, we used
HUVECs to test the ability of the two S. suis strains to
damage the cells. The results showed that 05ZYH33 has a higher
cytotoxicity than 1940, as determined by the LDH assay (Fig. 1D). Taken together, these data
demonstrate that the S. suis epidemic strain 05ZYH33 is more
virulent than the non-epidemic strain 1940.
Next, we investigated whether the sly protein may
contribute to the enhanced virulence of the S. suis epidemic
strain. Pre-opsonised S. suis 05ZYH33 or 1940 were incubated
with purified human PMNs (MOI =1) for 15 min in the presence or
absence of the sly protein, and the bacteria phagocytized by PMNs
were counted. This assay showed that PMNs phagocytized the S.
suis 1940 strain at higher rates compared to the 05ZYH33
strain, (Fig. 1E) while adding sly
protein significantly decreased the phagocytosis of S. suis
1940 by the PMNs.
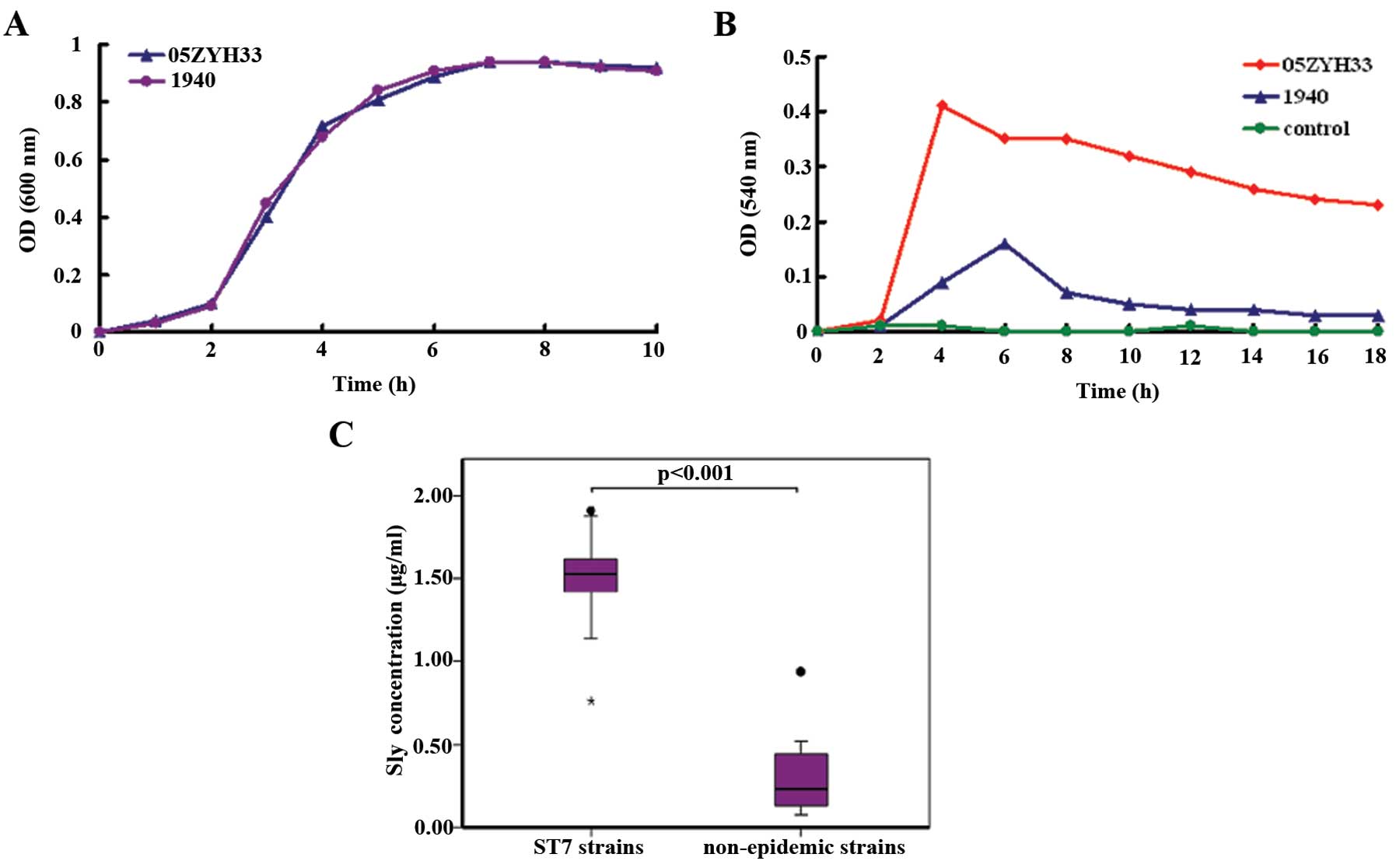
The S. suis epidemic strain 05ZYH33 shows
increased production of sly compared to the non-epidemic
strains
Since the sly protein displays hemolytic activity,
we then compared the relative amounts of sly produced by the S.
suis epidemic strain 05ZYH33 and the non-epidemic strain 1940.
Although both strains showed the same growth rate (Fig. 2A), the supernatant of S.
suis 05ZYH33 displayed higher hemolytic activity than that of
S. suis 1940 (Fig. 2B).
This phenotypic difference was further confirmed by qRT-PCR, which
indicated a difference in sly expression between the two
S. suis strains; a 6.3-fold difference at the mRNA
transcription level of sly was observed between the two
strains (data not shown).
In order to confirm the hypothesis that the S.
suis ST7 epidemic strains may have increased production of sly
compared to the non-epidemic strains, 11 S. suis ST7
epidemic strains and eight non-epidemic strains were collected
(Table I), and the amount of sly
was determined in the supernatant at 7 h of culture, using a
sandwich ELISA assay (Fig. 2C).
The epidemic ST7 strains produced more sly (mean, 1.49 g/ml; range,
0.76–1.91 g/ml) than the non-epidemic strains (mean, 0.33 g/ml;
range, 0.07–0.94 g/ml). Moreover, the difference between the two
groups was significant (p<0.001).
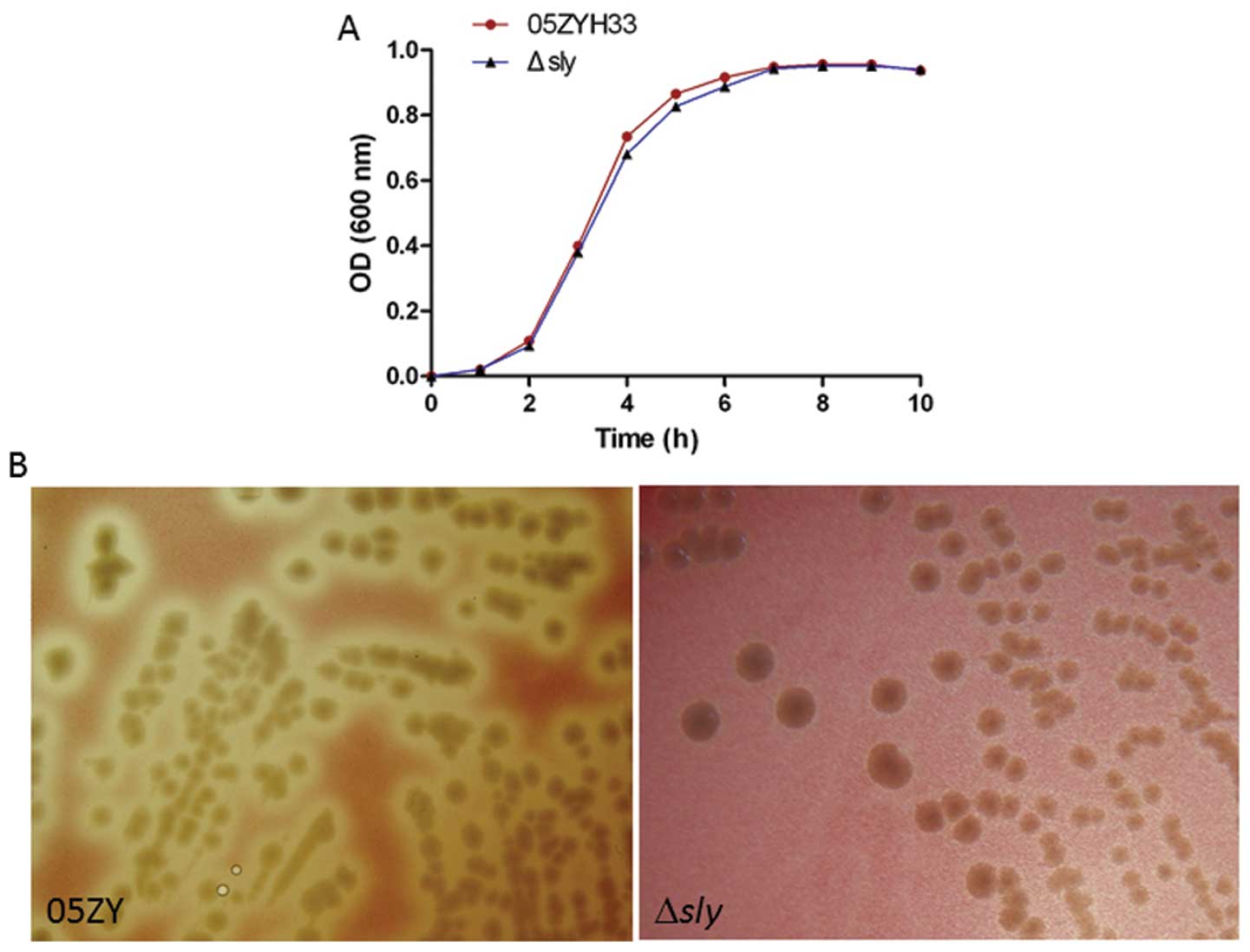
Sly is associated with enhanced severity
of S. suis infections
To further explore the roles of sly in invasive
infection of the 05ZYH33 strain, the sly gene was in-frame
substituted by the Cm gene in the S. suis 05ZYH33
strain, to generate the nonpolar mutant Δsly. The mutant
Δsly strain showed reduced hemolytic activity, but similar
growth rates compared to the wild-type strain S. suis
05ZYH33 (Fig. 3). We further
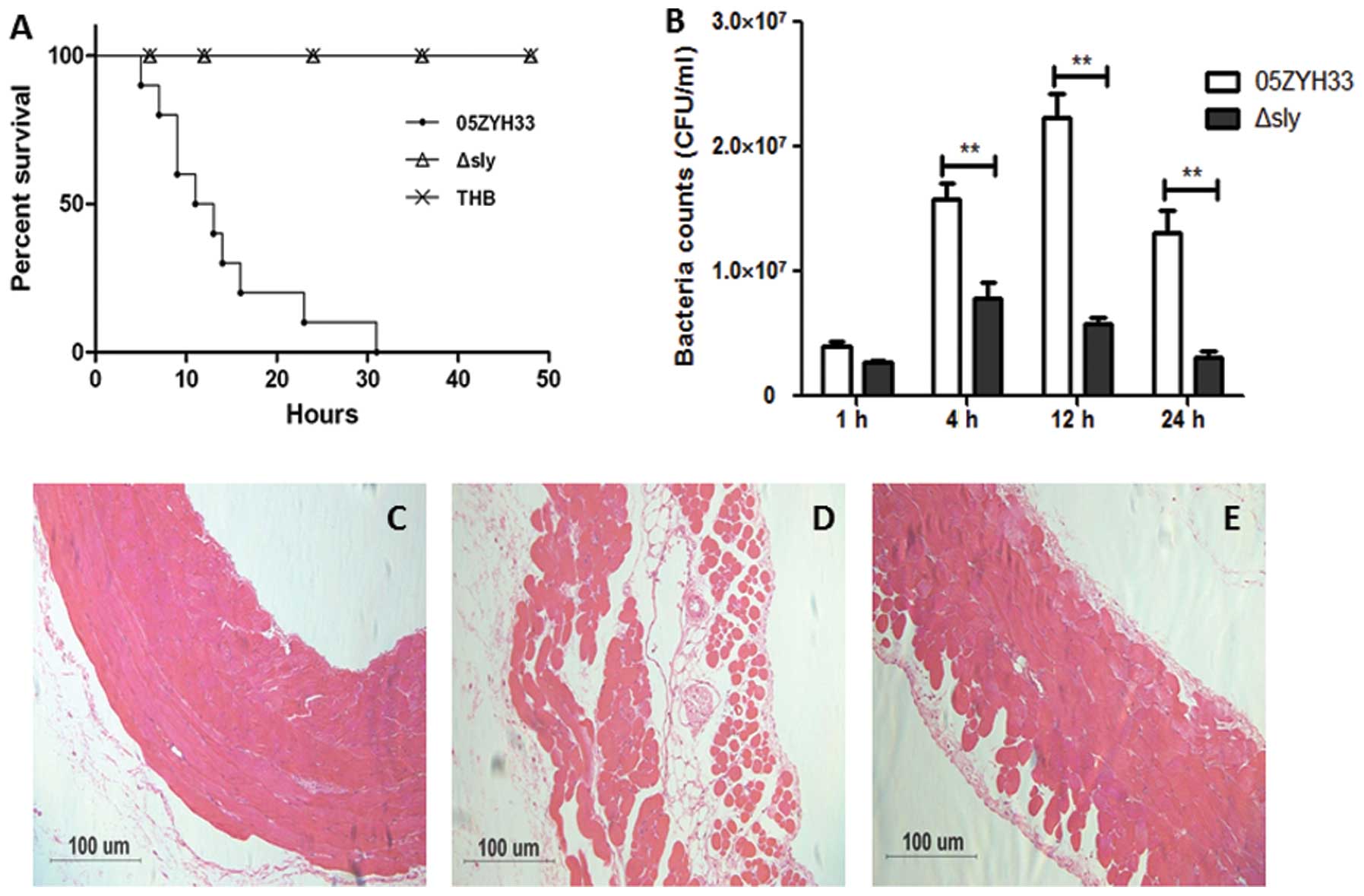
investigated the virulence of this mutant. CD1 mice (6-week-old
female, ~22 g) were infected by intraperitoneal injection with 0.5
ml (~5×108 CFUs) S. suis at the late-exponential
growth phase in THB. The mortality of the infected mice was
recorded for 50 h. No mice died in the Δsly or the THB
control group, while the mice infected with S. suis 05ZYH33
all died within 32 h post-infection. The bacterial concentration in
the blood was evaluated at different time-points post-infection
(Fig. 4A). The parental 05ZYH33
strain survived/multiplied in the blood at higher rates than
Δsly at 4, 12 and 24 h (Fig.
4B). Histopathological examination of the abdominal wall
tissues was also performed 6 h after the infection. The abdominal
epithelial cells of mice infected by S. suis 05ZYH33 lost
their normal morphological features and the integrity of the tight
junctions, while the epithelial cells infected by S. suis
Δsly showed a few changes (Fig.
4C–E). Taken together, these data demonstrate that sly is
associated with enhanced severity of S. suis 05ZYH33
infections.
Discussion
S. suis is a swine pathogen responsible for a
number of infections, including meningitis, endocarditis and
septicemiae, and is also an important zoonotic agent. The two
outbreaks that occurred in China in 1998 and 2005 affected the
world perspective regarding the threat that this pathogen
represents for humans. Subsequent investigations on S. suis
at the molecular and the genomic level confirmed that the emergence
of the highly virulent SS2 strains, or epidemic ST7 strains, played
a key role in these severe outbreaks (13). In the present study, we sought to
determine the capacity of the S. suis protein sly to enhance
the virulence of the epidemic ST7 strain 05ZYH33.
A previous study showed that S. suis ST7
strains express the proposed virulence markers muramidase-released
protein (MRP), extracellular protein factor (EF), and sly (15). However to date, there is no study
that specifically focused on how the differences in the expression
abundance of bacterial virulence factors may affect the severity of
S. suis infections. We hereby showed, using an animal model,
that the S. suis ST7 epidemic strain 05ZYH33 causes higher
mortality, higher necrosis of PMNs and significantly higher damage
to HUVECs than the S. suis non-epidemic strain 1940. In
agreement with these results, epidemic ST7 strains produced more
sly than non-epidemic strains. This significant difference in sly
secretion may be of outmost importance in S. suis 05ZYH33
infections. Phagocytosis is the first line of cellular defense
against acute infectious diseases. A clinical association with
invasive human infections implies that bacteria can survive the
innate host defense responses raised in order to clear the bacteria
from the bloodstream and the tissues. In other pathogenic
streptococcal infections, hemolysins have been shown to be involved
in bacteria-phagocyte interactions. For example, in Group B
Streptococcus (GBS) infections, Liu et al (32) elegantly demonstrated the
multi-factorial roles of the pore-forming toxin
β-hemolysin/cytolysin (βH/C) in thwarting the immune phagocytic
defenses. In the present study, we demonstrated that sly mediates
inhibition of the phagocytic uptake by human PMNs in vitro.
Moreover, we demonstrated that the S. suis strain with
enhanced sly expression causes enhanced PMN necrosis in an in
vivo mouse model. Taken together, these findings suggest that
the sly-mediated inhibition of phagocytosis and cytolysis is
directly associated with a mechanism impairing the function and
viability of PMNs, and thus contributing to the enhanced survival
of epidemic S. suis strains.
The ability of sly to enhance the severity of S.
suis infection was confirmed using a mutant of S. suis
05ZYH33 deficient in sly expression. The survival rate of
mice challenged by the Δsly strain was significantly
increased, and the bacterial count in the blood decreased compared
to the parental strain 05ZYH33. These data strongly support the
contribution of sly in enhancing the severity of S. suis ST7
strain infections, and are consistent with the previous study of
Allen et al (33).
In our intraperitoneal infection animal model, S.
suis needs to successfully cross the abdominal epithelial
barrier and survive in the bloodstream in order to cause invasive
infection. Among the potential strategies that can allow this are:
transcellular transport by passive or adhesion-induced
transcytosis, paracellular passage through opened tight junctions,
disruption of the barrier due to a direct cytotoxic effect,
leukocyte-facilitated transport by infected phagocytes (34). Histopathological examination showed
that the mice infected by S. suis 05ZYH33 lost their
abdominal epithelial cell morphological features and the integrity
of the tight junctions compared to mice infected by the mutant
Δsly. The S. pneumoniae toxin pneumolysin recognizes
Toll-like receptor (TLR)4 on dendritic cells (35), and S. pneumoniae and H.
influenzae were shown to exploit TLR-dependent downregulation
of claudins 7 and 10, tight junction key components for the
maintenance of the epithelial barrier integrity(36), to facilitate translocation across
the epithelium (37). However,
in vivo studies carried out in our laboratory indicate that
the S. suis sly protein contributes to the release of
inflammatory cytokines in a TLR4-independent manner (data not
shown). Whether activation of TLR4 by sly can facilitate the S.
suis translocation across the epithelium needs to be further
studied.
In conclusion, we have shown that differences in sly
production are linked to differences in S. suis virulence.
The pathogenic S. suis ST7 strain 05ZYH33 appeared to have
an increased ability to subvert host clearance mechanisms, which
may allow its survival and dissemination into the bloodstream. The
cytotoxic effects of sly may inhibit the uptake of microorganisms
by the host phagocytes and can directly cause epithelial cell
damage, enhancing the microbial spread into deeper tissues. Thus,
in a murine model, sly contributes to increased bacterial load and
death. Further understanding of the precise molecular mechanisms
underlying the sly-mediated pathogenicity may provide potential
therapeutic targets for a better control of S. suis
infections.
Acknowledgements
This study was supported by grants from the Natural
Sciences Foundation of China (30870091 and 81171528). We thank
Professor Henk J. Wisselink and Marcelo Gottschalk for kindly
providing S. suis strains.
References
|
1
|
Lun ZR, Wang QP, Chen XG, Li AX and Zhu
XQ: Streptococcus suis: an emerging zoonotic pathogen.
Lancet Infect Dis. 7:201–209. 2007. View Article : Google Scholar
|
|
2
|
Segura M: Streptococcus suis: an
emerging human threat. J Infect Dis. 199:4–6. 2009. View Article : Google Scholar
|
|
3
|
Wertheim HF, Nghia HD, Taylor W and
Schultsz C: Streptococcus suis: an emerging human pathogen.
Clin Infect Dis. 48:617–625. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hui AC, Ng KC, Tong PY, Mok V, Chow KM, Wu
A and Wong LK: Bacterial meningitis in Hong Kong: 10-years’
experience. Clin Neurol Neurosurg. 107:366–370. 2005.PubMed/NCBI
|
|
5
|
Mai NT, Hoa NT, Nga TV, et al:
Streptococcus suis meningitis in adults in Vietnam. Clin
Infect Dis. 46:659–667. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu H, Jing H, Chen Z, et al: Human
Streptococcus suis outbreak, Sichuan, China. Emerg Infect
Dis. 12:914–920. 2006.
|
|
7
|
Gottschalk M, Segura M and Xu J:
Streptococcus suis infections in humans: the Chinese
experience and the situation in North America. Anim Health Res Rev.
8:29–45. 2007. View Article : Google Scholar
|
|
8
|
Normile D: Infectious diseases. WHO probes
deadliness of China’s pig-borne disease. Science. 309:1308–1309.
2005.PubMed/NCBI
|
|
9
|
Ngo TH, Tran TB, Tran TT, et al:
Slaughterhouse pigs are a major reservoir of Streptococcus suis
serotype 2 capable of causing human infection in southern Vietnam.
PloS one. 6:e179432011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen C, Tang J, Dong W, et al: A glimpse
of streptococcal toxic shock syndrome from comparative genomics of
S. suis 2 Chinese isolates. PLoS One. 2:e3152007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li M, Wang C, Feng Y, Pan X, et al:
SalK/SalR, a two-component signal transduction system, is essential
for full virulence of highly invasive Streptococcus suis
serotype 2. PLoS One. 3:e20802008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li M, Shen X, Yan J, et al: GI-type
T4SS-mediated horizontal transfer of the 89K pathogenicity island
in epidemic Streptococcus suis serotype 2. Mol Microbiol.
79:1670–1683. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ye C, Zhu X, Jing H, et al:
Streptococcus suis sequence type 7 outbreak, Sichuan, China.
Emerg Infect Dis. 12:1203–1208. 2006. View Article : Google Scholar
|
|
14
|
Ye C, Bai X, Zhang J, et al: Spread of
Streptococcus suis sequence type 7, China. Emerg Infect Dis.
14:787–791. 2008.
|
|
15
|
Ye C, Zheng H, Zhang J, et al: Clinical,
experimental, and genomic differences between intermediately
pathogenic, highly pathogenic, and epidemic Streptococcus
suis. J Infect Dis. 199:97–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Alouf JE: Molecular features of the
cytolytic pore-forming bacterial protein toxins. Folia Microbiol
(Praha). 48:5–16. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Segura M and Gottschalk M:
Streptococcus suis interactions with the murine macrophage
cell line J774: adhesion and cytotoxicity. Infect Immun.
70:4312–4322. 2002. View Article : Google Scholar
|
|
18
|
Lalonde M, Segura M, Lacouture S and
Gottschalk M: Interactions between Streptococcus suis
serotype 2 and different epithelial cell lines. Microbiology.
146:1913–1921. 2000.
|
|
19
|
Charland N, Nizet V, Rubens CE, Kim KS,
Lacouture S and Gottschalk M: Streptococcus suis serotype 2
interactions with human brain microvascular endothelial cells.
Infect Immun. 68:637–643. 2000. View Article : Google Scholar
|
|
20
|
Jacobs AA, Loeffen PL, van den Berg AJ and
Storm PK: Identification, purification, and characterization of a
thiol-activated hemolysin (suilysin) of Streptococcus suis.
Infect Immun. 62:1742–1748. 1994.PubMed/NCBI
|
|
21
|
Jacobs AA, van den Berg AJ and Loeffen PL:
Protection of experimentally infected pigs by suilysin, the
thiol-activated haemolysin of Streptococcus suis. Vet Rec.
139:225–228. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pian Y, Gan S, Wang S, et al: Fhb, a novel
factor H-binding surface protein, contributes to the antiphagocytic
ability and virulence of Streptococcus suis. Infect Immun.
80:2402–2413. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lv QY, Hao HJ, Bi LL, Zheng YL, Jiang YQ
and Lv SX: Purification and biological activities analysis of
streptococcus suis Serotype 2 suilysin. Xi Bao Yu Fen Zi Mian Yi
Xue Za Zhi. 27:374–376. 2011.(In Chinese).
|
|
24
|
Gottschalk MG, Lacouture S and Dubreuil
JD: Characterization of Streptococcus suis capsular type 2
haemolysin. Microbiology. 141:189–195. 1995. View Article : Google Scholar
|
|
25
|
Berthelot-Hérault F, Gottschalk M, Morvan
H and Kobisch M: Dilemma of virulence of Streptococcus suis:
Canadian isolate 89–1591 characterized as a virulent strain using a
standardized experimental model in pigs. Can J Vet Res. 69:236–240.
2005.PubMed/NCBI
|
|
26
|
Wisselink HJ, Joosten JJ and Smith HE:
Multiplex PCR assays for simultaneous detection of six major
serotypes and two virulence-associated phenotypes of
Streptococcus suis in tonsillar specimens from pigs. J Clin
Microbiol. 40:2922–2929. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Martinez G, Gottschalk M, et al:
Identification of a surface protein of Streptococcus suis and
evaluation of its immunogenic and protective capacity in pigs.
Infect Immun. 74:305–312. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wisselink HJ, Reek FH, Vecht U,
Stockhofe-Zurwieden N, Smits MA and Smith HE: Detection of virulent
strains of Streptococcus suis type 2 and highly virulent
strains of Streptococcus suis type 1 in tonsillar specimens
of pigs by PCR. Vet Microbiol. 67:143–157. 1999.
|
|
29
|
Takamatsu D, Osaki M and Sekizaki T:
Thermosensitive suicide vectors for gene replacement in
Streptococcus suis. Plasmid. 46:140–148. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schmittgen TD and Zakrajsek BA: Effect of
experimental treatment on housekeeping gene expression: validation
by real-time, quantitative RT-PCR. J Biochem Biophys Methods.
46:69–81. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu GY, Doran KS, Lawrence T, Turkson N,
Puliti M, Tissi L and Nizet V: Sword and shield: linked group B
streptococcal beta-hemolysin/cytolysin and carotenoid pigment
function to subvert host phagocyte defense. Proc Natl Acad Sci USA.
101:14491–14496. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Allen AG, Bolitho S, Lindsay H, et al:
Generation and characterization of a defined mutant of
Streptococcus suis lacking suilysin. Infect Immun.
69:2732–2735. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Join-Lambert O, Morand PC, Carbonnelle E,
Coureuil M, Bille E, Bourdoulous S and Nassif X: Mechanisms of
meningeal invasion by a bacterial extracellular pathogen, the
example of Neisseria meningitidis. Prog Neurobiol.
91:130–139. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bernatoniene J, Zhang Q, Dogan S, Mitchell
TJ, Paton JC and Finn A: Induction of CC and CXC chemokines in
human antigen-presenting dendritic cells by the pneumococcal
proteins pneumolysin and CbpA, and the role played by toll-like
receptor 4, NF-kappaB, and mitogen-activated protein kinases. J
Infect Dis. 198:1823–1833. 2008. View
Article : Google Scholar
|
|
36
|
Dominguez-Punaro MC, Segura M, Plante MM,
Lacouture S, Rivest S and Gottschalk M: Streptococcus suis
serotype 2, an important swine and human pathogen, induces strong
systemic and cerebral inflammatory responses in a mouse model of
infection. J Immunol. 179:1842–1854. 2007. View Article : Google Scholar
|
|
37
|
Clarke TB, Francella N, Huegel A and
Weiser JN: Invasive bacterial pathogens exploit TLR-mediated
downregulation of tight junction components to facilitate
translocation across the epithelium. Cell Host Microbe. 9:404–414.
2011. View Article : Google Scholar
|