Introduction
Adipose tissue is an active metabolic and endocrine
organ that secretes adipokines acting locally or peripherally to
influence multiple biological processes, including glucose and
fatty acid metabolism, insulin sensitivity, adipocyte
differentiation, inflammation and immunity (1). The adipose tissue that accumulates in
the body is classified into two categories: subcutaneous adipose
tissue, which is stored under the skin, and visceral adipose
tissue, which is stored in or surrounding the internal organs
(2,3). Excess visceral adipose tissue may
result in serious health problems as it surrounds the vital organs
and is metabolised by the liver into cholesterol. Accumulation of
the two types of adipose tissues may be age-related and the
accumulation of visceral adipose tissue has been demonstrated to
increase with age (4). The
visceral adipose tissue and the adipokines secreted by this tissue
appear to be key components that determine and predict the
development of numerous visceral adipose tissue-related
diseases.
Excessive accumulation of visceral adipose tissues
in humans is associated with various chronic diseases, including
type II diabetes, high blood pressure, hypertension, dyslipidemia,
various types of cancer and cardiovascular diseases (5,6).
Evidence has suggested that the excessive accumulation of adipose
tissue alters the secretion of adipokines, including leptin,
interleukin (IL)-6, resistin, CCL2, adiponectin and tumour necrosis
factor (TNF)-α, which promote proliferation, angiogenesis and
invasion in certain types of cancer (7). Changes in the adipokine levels affect
cell proliferation, apoptosis and invasive growth of cancer cells
(8). The adipokines produced by
visceral adipose tissue have been shown to be associated with the
risk and the progression of breast cancer (9,10).
Therefore, elucidating the genes that regulate visceral adipose
tissue accumulation and the adipokine network is important to
understand the progression of visceral adipose tissue-related
diseases further. This research may result in the development of
novel therapies to regulate the secretion of adipokines involved in
disease pathogenesis, and the identification of potential
biomarkers for the detection and prevention of the visceral adipose
tissue-related diseases.
A number of studies have been conducted to
understand the general mechanisms that regulate alterations in
adipose tissue mass. However, the mechanism underlying visceral
adipose tissue accumulation remains unclear. Therefore, the present
study investigated the gene expression profiles in accumulated
visceral adipose tissues and the levels of adipokines in serum
samples, using an animal model. The aim was to identify the
molecular targets that facilitate excessive visceral adipose tissue
accumulation. The focus was on peroxisome proliferator-activated
receptors (PPARs) since these genes are key mediators in
adipogenesis and regulate the adipokine network (11). Following an excessive feeding
regimen for 8 and 16 weeks, the expression levels of PPAR mRNA and
protein in visceral adipose tissue were analysed using reverse
transcription-polymerase chain reaction (RT-PCR) and Western
blotting, respectively. In addition, the levels of adipokines in
the sera of the animals were determined using ELISA.
Materials and methods
Animal feeding, serum and adipose tissue
collection
The use of animals was approved by the Research
Ethics (Animal) Committee, Universiti Sains Malaysia (Penang,
Malaysia). Male Sprague Dawley rats (weight, 80–100 g; age, 4
weeks) were obtained from the Animal House of the Universiti Sains
Malaysia (Kubang Kerian, Kelantan, Malaysia). The rats were
randomly assigned to two groups: Six control rats were fed a normal
intake of food (5 g pellet/100 g body weight per day) and six
experimental rats were fed an excessive intake of food (10 g
pellet/100 g body weight per day). The animal feed was obtained
from Gold Coin Feedmills Sdn. Bhd (Penang, Malaysia). The rats were
provided pre-weighed quantities of food each day and the quantity
remaining in the food dish was weighed every 24 h. The body weight
of each rat was measured weekly to determine growth. A total of
four groups were used, with six rats in the 8 week group (three
control rats; three experimental rats) and six in the 16 week group
(three control rats; three experimental rats). The experiment was
repeated twice. Following a feeding period of 8 weeks, the waist
circumferences of the three rats in each group were measured prior
to sacrifice via brief exposure to 10% (v/v) diethyl ether in a
chamber. The same measurement was peformed prior to sacrifice in
the three rats in each of the two 16 week feeding period groups.
Blood was also collected from the rats in each group following the
feeding periods of 8 and 16 weeks by cardiac puncture, left to clot
and centrifuged at 3,000 × g for 15 mins to obtain serum for ELISA.
For long-term storage, the serum samples were maintained at −80°C.
The adipose tissues (visceral and subcutaneous) were collected and
weighed. The visceral adipose tissues were used for total RNA
extraction or rinsed in TRI Reagent® (Molecular Research
Center, Inc., Cincinnati, OH, USA), frozen in liquid nitrogen and
stored at −80°C until use.
Total RNA extraction and cDNA
synthesis
Total RNA extraction was performed using the TRI
Reagent®, according to the manufacturer’s instructions.
For this, total RNA was extracted from the aqueous phase
(colourless top layer) of the homogenised tissue samples and the
remaining organic phase was stored at −80°C for subsequent protein
extraction. The integrity of the total RNA extracted was confirmed
using agarose gel electrophoresis, while the purity and yield of
the RNA were measured using a nanophotometer (Implen, München,
Germany). Subsequently, 1 μg total RNA was reverse-transcribed to
cDNA using the RevertAid™ First Strand cDNA Synthesis kit
(Fermentas, Waltham, MA, USA), according to the manufacturer’s
instructions. The success of cDNA synthesis was validated by
conventional PCR using rat GAPDH primers. The sequences of the
GAPDH and PPAR primers used in the present study are listed in
Table I.
 | Table IPrimers used for RT-PCR
amplification. |
Table I
Primers used for RT-PCR
amplification.
| Gene | Primer sequences | Amplicon (bp) |
|---|
| PPARα | F:
5′-TGGAGTCCACGCATGTGAAG-3′
R: 5′-TTGTCGTACGCCAGCTTTAGC-3′ | 72 |
| PPARδ | F:
5′-CATGAGTTCTTGCGCAGTATCC-3′
R: 5′-AGAGCATTGAACTTGACAGCAAAC-3′ | 83 |
| PPARγ | F:
5′-CATGACCAGGGAGTTCCTCAA-3′
R: 5′-AGCAAACTCAAACTTAGGCTCCAT-3′ | 73 |
| GAPDH | F: 5′-CAAGTTCAA
CGGCACAGTCAAG-3′
R: 5′-CTCCTGGAAGATGGTGATTGGT-3′ | 76 |
Determination of PPAR expression
levels
Following cDNA synthesis, the PPAR expression levels
were determined using 2X Power SYBR Green PCR master mix (Applied
Biosystems, Inc., Foster City, CA, USA) in a Rotor-Gene 6000 PCR
machine (Qiagen, Hilden, Germany). The final concentrations of cDNA
and each primer used were 5 ng/μl and 0.4 μM, respectively. The
GAPDH gene served as a reference gene, to compare the expression
levels amongst different samples. All primers were designed with an
annealing temperature of 55°C using the Primer Express 2.0 software
(Applied Biosystems, Inc.). All PCR reactions were performed at
94°C for 10 min followed by 40 cycles of denaturation at 94°C for
20 sec, primer annealing at 55°C for 20 sec and extension at 72°C
for 30 sec. A dissociation curve program was added following the
completion of the reaction in order to generate the derivative
dissociation curve. The derivative dissociation curve was created
to confirm the primer-dimer artifact and to ensure the RT-qPCR
specificity. The dissociation curve program was conducted at 95°C
for 90 sec and 72°C to 95°C at 1°C/5 sec (1°C increase every 5 sec
between 72 and 95°C) using the same PCR machine. Each PCR reaction
was performed as three replicates in two independent experiments.
The PPAR gene expression levels were determined using the relative
quantification approach. The 2(−ΔΔCt) method was used to
analyse relative change in gene expression level (12). The value for fold-change in gene
expression level was calculated as determined by the target gene
expression level versus the reference gene expression level.
Protein extraction
The total protein from visceral adipose tissue was
extracted from the organic phase of the samples as described above.
The protein concentration was determined using the DC Protein Assay
kit (Bio-Rad, Hercules, CA, USA)and then used for Western blot
analysis.
Western blot analysis
The protein samples (30 μg per well) were separated
on a 10% SDS-polyacrylamide gel. The samples were then
electrophoretically transferred to a nitrocellulose membrane, then
blocked using 5% skimmed milk powder for 60 min at room temperature
with constant agitation. The membrane was washed three times for 10
min each using 1X Tris-buffered saline and Tween-20, and then
incubated with 1:250 responsive primary rabbit polyclonal antibody
for PPARα (H-98: sc-9000), PPARδ (H-74: sc-7197) and PPARγ (H-100:
sc-7196) at 4°C, overnight. All antibodies were obtained from Santa
Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Subsequent to
washing, the membrane was incubated at room temperature for 1 h
with 1:5,000 horseradish peroxidase-conjugated rat anti-rabbit
antibody (Santa Cruz Biotechnology, Inc.). Subsequent to another
washing step, the blot was incubated with the SuperSignal West Pico
Chemiluminescent Substrate (Pierce Biotechnology, Inc., Rockford,
IL, USA). The blot was then developed on film in a dark room.
Anti-rabbit GAPDH antibody (Santa Cruz Biotechnology, Inc.) was
used as a control, to verify equal sample loading.
Determination of adipokine expression
levels
The expression levels of the CCL2, TNF-α,
adiponectin, leptin, resistin, IL-6 adipokines and C-reactive
protein (CRP; control) in the serum samples of control and
experimental rats, were determined using commercially available
ELISA kits (Abnova, Taipei, Taiwan). In these kits, antibodies
specific for the adipokines had been pre-coated onto 96-well
microtitre plates. The serum samples were then added to the wells
and allowed to react with the bound antibody for 2.5 h at room
temperature. The unbound antibodies were washed away with a Wash
Solution, according to the manufacturer’s instructions.
Subsequently, an enzyme-linked antibody specific to the adipokine
was added to the wells. The antibody was incubated with the
targeted protein for 1 h. Following another washing step, a
substrate solution was added to the wells for colour development.
The colour developed was proportional to the quantity of adipokine
present in the samples. The colour intensity was then measured
using an ELISA reader (Tecan Group, Ltd., Männedorf, Switzerland)
at a wavelength of 450 nm and the level of each adipokine in the
serum samples was calculated. The ELISAs were performed in
triplicate and repeated in two independent experiments.
Statistical analysis
All graphs and statistical analyses were performed
using the GraphPad Prism5 software (GraphPad Software Inc., La
Jolla, CA, USA). All values are expressed as the mean ± standard
deviation. Student’s t-test was used to compare the means of two
groups and one way analysis of variance (ANOVA) was used to compare
the means between the groups. P<0.05 was considered to indicate
a statistically significant difference.
Results
Weight gain
The results demonstrated that weight gain increased
with time and that the rats in the experimental group, which
consumed more food, gained more weight compared with the rats in
the control group. The weight gain reached a plateau phase at a
feeding period of 14 weeks, whereby the weights of the control and
experimental rats were ~325 and 400 g, respectively. However,
following feeding periods of 14 weeks, no more weight gain was
observed in either the control or the experimental rats. In the
experimental rats (fed with excessive food intake), the
accumulation of adipose tissue mass was first observed at 8
weeks.
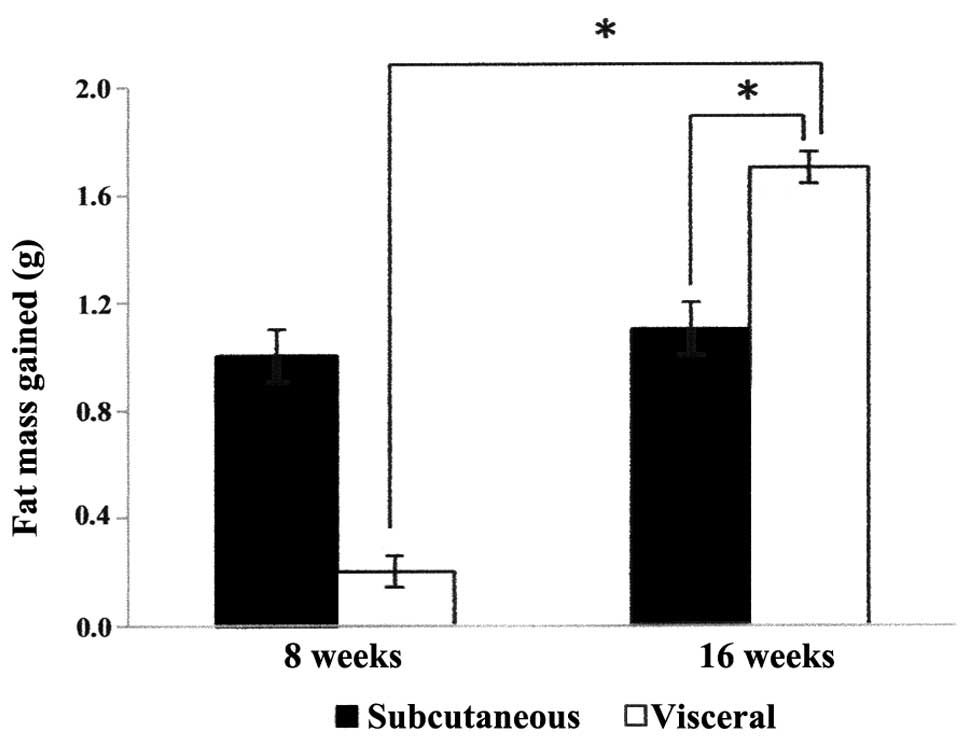
Adipose tissue mass
In the experimental rats (fed with excessive food
intake), the accumulation of adipose tissue mass was first observed
at 8 weeks. The subcutaneous adipose tissue mass collected was
higher than the visceral adipose tissue mass (Fig. 1). At 16 weeks, the quantity of
subcutaneous adipose tissue mass in the experimental rats was
unchanged as compared with the fat mass at 8 weeks. However, the
accumulation of visceral adipose tissue mass was significantly
increased (P<0.05) in the experimental rats at 16 weeks, when
compared with the fat mass at 8 weeks. The visceral adipose tissue
mass in the experimental rats was also significantly higher than
the mass of the subcutaneous adipose tissue (P<0.05) at 16
weeks.
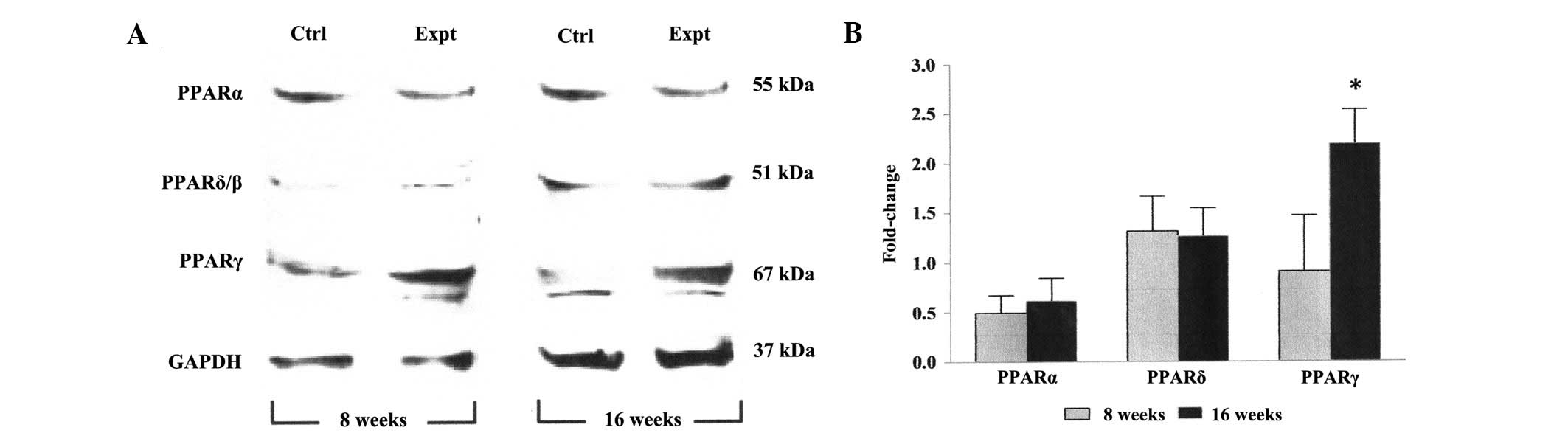
Expression levels of PPARs in visceral
adipose tissue of rats on an excessive feeding regimen
The PCR results detected no significant difference
(P>0.05) in the PPARα and PPARδ mRNA expression levels in the
visceral adipose tissue between samples obtained at 8 and 16 weeks
(Fig. 2). Conversely, at 16 weeks,
the experimental rats exhibited a significant increase (P<0.05;
~2-fold) in the mRNA expression level of PPARγ relative to the
result obtained at 8 weeks. A similar pattern in the protein
expression levels of all PPARs, as assessed by Western blotting,
was observed in the visceral adipose tissue of experimental rats at
8 and 16 weeks (Fig. 3).
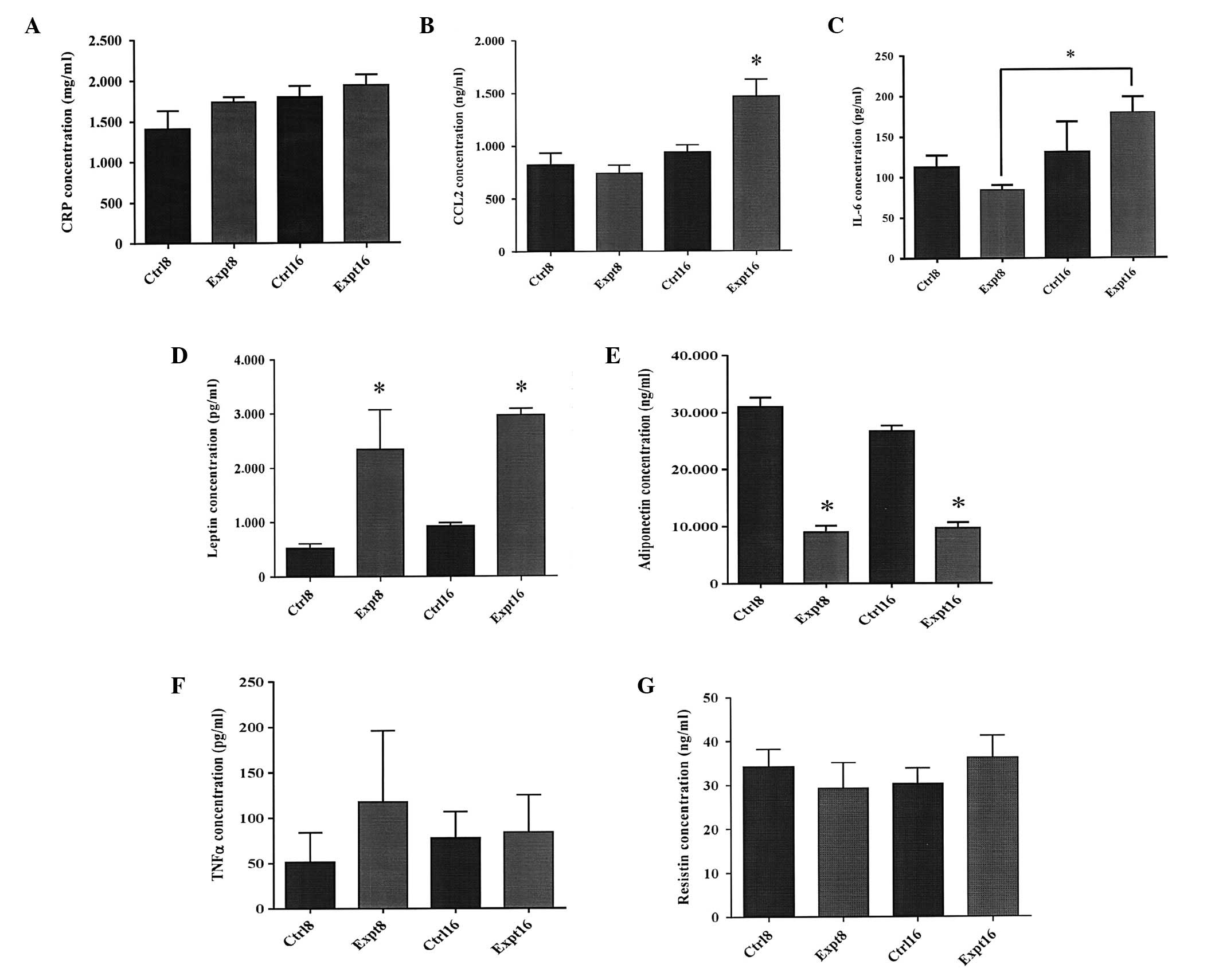
Levels of adipokines in the sera of rats
on an excessive feeding regimen
As shown in Fig. 4,
no significant differences (P>0.05) were observed in the levels
of CCL2 and IL-6 in serum samples between experimental and control
rats at 8 weeks. However, at 16 weeks, significantly higher levels
of CCL2 (P<0.05) were detected in the serum samples of the
experimental rats (1,465 ng/ml), compared with those of the control
rats (939.8 ng/ml; Fig. 4B). In
addition, at 16 weeks, the levels of IL-6 in the experimental rats
(179.1 pg/ml) were significantly higher (P<0.05), when compared
with those at 8 weeks (84.3 pg/ml; Fig. 4C). The levels of leptin at 8 weeks
were found to be significantly increased (P<0.05) in the serum
samples of experimental rats (2,341 pg/ml), compared with those of
the control rats (526.7 pg/ml). This phenomenon was also observed
at 16 weeks, with 2,971 pg/ml in experimental rats and 928.9 pg/ml
in control rats (P<0.05; Fig.
4D). Conversely, significantly reduced levels of adiponectin
(P<0.05) were observed in the serum samples of experimental rats
compared with the control rats at 8 and 16 weeks (Fig. 4E). No significant differences were
identified in the levels of the resistin and TNF-α between
experimental and control rats in all serum samples (Fig. 4F and G).
Discussion
In a previous study, female rats fed an excessive
food intake developed more mesenteric adipose tissue compared with
male rats (13). However, the
female rats were able to oxidise the tissue faster, thus male rats
were considered to exhibit a higher tendency towards
obesity-related diseases. Therefore, in the present study, only
male rats were used. An increase in the visceral adipose tissue
mass was observed in the Sprague Dawley experimental rats with
excessive food intake for 16 weeks. An increase in the expression
levels of PPARγ mRNA and protein were also detected in the visceral
adipose tissue of experimental rats, along with increased levels of
CCL2 and IL-6 in the sera. These increases were correlated with
increased accumulation of visceral adipose tissue in the rats.
In the majority of previously reported studies, a
high fat diet was used to analyse weight gain and adipose tissue
accumulation in the animals (14,15).
However, in the present study, accumulation of subcutaneous and
visceral adipose tissues was investigated via excessive food
intake. Nevertheless, similar results were obtained, in which
weight gain and accumulation of adipose tissue in the rats with
excessive food intake was observed. Weight gain and an increase in
subcutaneous adipose tissue was observed in the experimental rats
at 8 weeks. After 8 weeks of the feeding regimen, marginal weight
gain was observed in the experimental rats, despite an increase in
visceral adipose tissue. This phenomenon suggests that accumulation
of visceral adipose tissue at 16 weeks may be independent of weight
gain.
At the molecular level, the expression levels of
PPARs were analysed to identify the genes associated with visceral
adipose tissue accumulation. The PPARγ expression level exhibited a
positive correlation with the expression levels of genes involved
in the formation of visceral fat deposits following excessive food
intake for 16 weeks. However, the expression levels of PPARα and
PPARδ in the visceral adipose tissues collected from the rats of
experiment and control groups, at 8 and 16 weeks, were not
identified to be significantly different in the present study.
Thus, this finding reveals the important role of PPARγ in visceral
adipose tissue accumulation. Furthermore, the PPARγ mRNA and
protein expression levels were found to correlate with visceral
adipose tissue accumulation at the later phase (16 weeks) of the
adipose tissue development. PPARγ is predominant in the regulation
of adipocyte proliferation, lipid accumulation and specific
adipocyte biomarker expression (16). A study conducted by Duffield et
al (17) demonstrated that
early change in the PPARγ expression level in the visceral adipose
tissue of lambs results in obesity in later life. This may be due
to PPARγ activation, which enhances hyperplasia in adipocytes and
results in greater accumulation of adipose tissue in the body.
PPARγ has also been reported to activate other factors involved in
regulating the development of adipose tissue (18). When PPARγ is upregulated due to a
high-fat diet, other adipocyte differentiation-related genes, such
as aP2 and C/EBPα, are also overexpressed (18). Omentum fat accumulation also
exhibits a correlation with the expression levels of PPARγ
(19), which is consistent with
the findings of the present study. Furthermore, PPARγ-knockout mice
were unable to survive due to the inhibition of adipogenesis
(21). The PPARγ gene has also
been reported to be involved in the pathogenesis of numerous other
diseases, such as type 2 diabetes, glomerulosclerosis and several
types of cancer (21–23).
Adipocytes are known to secrete a large number of
growth hormones, growth factors, enzymes, cytokines, complement
factors and matrix proteins, which are indirectly correlated with
obesity (24). The expression of
PPARγ in adipose tissue is considered to induce the production of
various enzymes and proteins responsible for the activation of
soluble factors, including adipokines and growth hormones, which
are detectable in the serum (25).
In the present study, the results revealed that the expression
levels of PPARγ in visceral adipose tissue may be correlated with
upregulation of serum CCL2 and IL-6. Upregulation of serum leptin
and downregulation of serum adiponectin were also observed in the
experimental rats. However, the levels of these adipokines may be
associated with adipose tissue accumulation in general and not
specifically with the accumulation of visceral adipose tissue.
CCL2 is an important cytokine that has been
associated with adipose tissue-related diseases (26,27).
In the present study, the level of serum CCL2 increased in rats
with accumulated visceral adipose tissue. A similar finding was
reported in a study conducted by Chen et al (28) who revealed that CCL2/monocyte
chemoattractant proteins-1 and -3 were elevated in mice
administered a high-fat diet for 4 weeks, and this phenomenom was
ascribed to systemic inflammation. A study by Park et al
(29) demonstrated correlations
among serum levels of CRP, IL-6 and TNF-α, obesity and visceral
adiposity. Significant correlations were observed between
concentrations of serum CRP and IL-6 and visceral adiposity, while
concentrations of TNF-α exhibited a significant correlation with
weight, body-mass index and waist circumference. Notably, the
findings of the present study revealed that serum CCL2 and IL-6
levels were increased in rats with visceral adipose tissue
accumulation following excessive food intake, which is in
concordance with the results of a number of these previous studies.
However, no significance differences were identified in the levels
of TNF-α amongst all serum samples. A large number of studies have
demonstrated that CCL2 and IL-6 facilitate tumour progression in
pancreatic (30), breast (31,32)
and gastric cancer (33). Thus,
CCL2 and IL-6 are potential molecular targets that indicate the
expression level of PPARγ in visceral adipose tissue as well as the
accumulation of visceral adipose tissue in the body.
In conclusion, accumulation of visceral adipose
tissue mass was observed in rats after 16 weeks of an excessive
feeding regimen and the expression level of PPARγ may have
facilitated the process of visceral adipose tissue accumulation.
The increase of PPARγ expression in visceral adipose tissue was
correlated with increased levels of CCL2 and IL-6 in the serum. The
CCL2 and IL-6, which are easily detected in the serum, are thus
potential surrogate biomarkers for the level of PPARγ expression in
accumulated visceral adipose tissue. This may have applications for
the development of novel diagnostic and therapeutic strategies to
address visceral adipose tissue-related diseases. However, further
in-depth confirmatory studies are required in the near future.
Acknowledgements
This study was funded by a Fundamental Research
Grant Scheme Fasa 1/2008 (grant no. 203/CIPPM/6711119) from the
Ministry of Higher Education and the Postgraduate Research Grant
Scheme (grant no. 1001/CIPPM/834092) from the Universiti Sains
Malaysia. Ms. Thaneswary was supported by a Universiti Sains
Malaysia fellowship from the Institute of Postgraduate Studies. The
authors would also like to thank the staff at the Genomic
Laboratory of the Institute for Research in Molecular Medicine for
their technical support.
References
|
1
|
Balistreri CR, Caruso C and Candore G: The
role of adipose tissue and adipokines in obesity-related
inflammatory diseases. Mediators Inflamm. 2010:8020782010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mårin P, Andersson B, Ottosson M, et al:
The morphology and metabolism of intraabdominal adipose tissue in
men. Metabolism. 41:1242–1248. 1992.PubMed/NCBI
|
|
3
|
Wajchenberg BL: Subcutaneous and visceral
adipose tissue: their relation to the metabolic syndrome. Endocr
Rev. 21:697–738. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pascot A, Lemieux S, Lemieux I, et al:
Age-related increase in visceral adipose tissue and body fat and
the metabolic risk profile of premenopausal women. Diabetes Care.
22:1471–1478. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sharma AM and Staels B: Review: Peroxisome
proliferator-activated receptor gamma and adipose tissue -
understanding obesity-related changes in regulation of lipid and
glucose metabolism. J Clin Endocrinol Metab. 92:386–395. 2007.
View Article : Google Scholar
|
|
6
|
Fusco R, Galgani M, Procaccini C, et al:
Cellular and molecular crosstalk between leptin receptor and
estrogen receptor-{alpha} in breast cancer: molecular basis for a
novel therapeutic setting. Endocr Relat Cancer. 17:373–382. 2010.
View Article : Google Scholar
|
|
7
|
Hlavna M, Kohut L, Lipkova J, et al:
Relationship of resistin levels with endometrial cancer risk.
Neoplasma. 58:124–128. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Masaki T and Yoshimatsu H: Obesity,
adipokines and cancer. Transl Oncogenomics. 3:45–52. 2008.
|
|
9
|
Nyante SJ, Gammon MD, Kaufman JS, et al:
Common genetic variation in adiponectin, leptin, and leptin
receptor and association with breast cancer subtypes. Breast Cancer
Res Treat. 129:593–606. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim SH, Nagalingam A, Saxena NK, Singh SV
and Sharma D: Benzyl isothiocyanate inhibits oncogenic actions of
leptin in human breast cancer cells by suppressing activation of
signal transducer and activator of transcription 3. Carcinogenesis.
32:359–367. 2011. View Article : Google Scholar
|
|
11
|
Carter JC and Church FC: Obesity and
breast cancer: the roles of peroxisome proliferator-activated
receptor-γ and plasminogen activator inhibitor-1. PPAR Res.
2009:3453202009.
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
|
|
13
|
Priego T, Sánchez J, Picó C and Palou A:
Sex-differential expression of metabolism-related genes in response
to a high-fat diet. Obesity (Silver Spring). 16:819–826. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nomaguchi K, Tanaka M, Misawa E, et al:
Aloe vera phytosterols act as ligands for PPAR and improve the
expression levels of PPAR target genes in the livers of mice with
diet-induced obesity. Obes Res Clin Pract. 5:e169–e266. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kavanagh K, Jones KL, Sawyer J, et al:
Trans fat diet induces abdominal obesity and changes in insulin
sensitivity in monkeys. Obesity (Silver Spring). 15:1675–1684.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hamdy O, Porramatikul S and Al-Ozairi E:
Metabolic obesity: the paradox between visceral and subcutaneous
fat. Curr Diabetes Rev. 2:367–373. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Duffield JA, Vuocolo T, Tellam R, et al:
Intrauterine growth restriction and the sex specific programming of
leptin and peroxisome proliferator-activated receptor gamma
(PPARgamma) mRNA expression in visceral fat in the lamb. Pediatr
Res. 66:59–65. 2009. View Article : Google Scholar
|
|
18
|
Dewulf EM, Cani PD, Neyrinck AM, et al:
Inulin-type fructans with prebiotic properties counteract GPR43
overexpression and PPARgamma-related adipogenesis in the white
adipose tissue of high-fat diet-fed mice. J Nutr Biochem.
22:712–722. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arai K, Soga T, Ohata H, Otagiri A and
Shibasaki T: Effects of food restriction on peroxisome
proliferator-activated receptor-gamma and glucocorticoid receptor
signaling in adipose tissues of normal rats. Metabolism. 53:28–36.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barak Y and Kim S: Genetic manipulations
of PPARs: effects on obesity and metabolic disease. PPAR Res.
2007:127812007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nicholas SB, Kawano Y, Wakino S, Collins
AR and Hsueh WA: Expression and function of peroxisome
proliferator-activated receptor-gamma in mesangial cells.
Hypertension. 37:722–727. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Larsen TM, Toubro S and Astrup A:
PPARgamma agonists in the treatment of type II diabetes: is
increased fatness commensurate with long-term efficacy? Int J Obes
Relat Metab Disord. 27:147–161. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lehrke M and Lazar MA: The many faces of
PPARgamma. Cell. 123:993–999. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Frühbeck G and Salvador J: Role of
adipocytokines in metabolism and disease. Nutr Res. 24:803–826.
2004.
|
|
25
|
Tontonoz P and Spiegelman BM: Fat and
beyond: the diverse biology of PPARgamma. Annu Rev Biochem.
77:289–312. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Malavazos AE, Cereda E, Morricone L, et
al: Monocyte chemoattractant protein 1: a possible link between
visceral adipose tissue-associated inflammation and subclinical
echocardiographic abnormalities in uncomplicated obesity. Eur J
Endocrinol. 153:871–877. 2005. View Article : Google Scholar
|
|
27
|
Huber J, Kiefer FW, Zeyda M, et al: CC
chemokine and CC chemokine receptor profiles in visceral and
subcutaneous adipose tissue are altered in human obesity. J Clin
Endocrinol Metab. 93:3215–3221. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen A, Mumick S, Zhang C, et al: Diet
induction of monocyte chemoattractant protein-1 and its impact on
obesity. Obes Res. 13:1311–1320. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Park HS, Park JY and Yu R: Relationship of
obesity and visceral adiposity with serum concentrations of CRP,
TNF-alpha and IL-6. Diabetes Res Clin Pract. 69:29–35. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Monti P, Leone BE, Marchesi F, et al: The
CC chemokine MCP-1/CCL2 in pancreatic cancer progression:
regulation of expression and potential mechanisms of antimalignant
activity. Cancer Res. 63:7451–7461. 2003.PubMed/NCBI
|
|
31
|
Ueno T, Toi M, et al: Significance of
macrophage chemoattractant protein-1 in macrophage recruitment,
angiogenesis, and survival in human breast cancer. Clin Cancer Res.
6:3282–3289. 2000.PubMed/NCBI
|
|
32
|
Saji H, Koike M, Yamori T, et al:
Significant correlation of monocyte chemoattractant protein-1
expression with neovascularization and progression of breast
carcinoma. Cancer. 92:1085–1091. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ohta M, Kitadai Y, Tanaka S, et al:
Monocyte chemoattractant protein-1 expression correlates with
macrophage infiltration and tumor vascularity in human gastric
carcinomas. Int J Oncol. 22:773–778. 2003.
|