Introduction
Organ transplantation is considered the best option
for treating patients who suffer from organ failure or dysfunction.
However, the long-term allograft survival rate remains
unsatisfactory due to alloimmune rejection reactions. Thus,
treating acute rejection is the only option to improve the
long-term allograft survival rate. Acute allograft rejection is
thought to be predominantly affected by T-cell mediated
processes.
Interleukin-2 (IL-2) is considered to be the unique
growth factor for T cells; however, rejection episodes are not
always mediated by IL-2. Allograft rejection still occurs despite
IL-2/IL-2R blockade (1), knockout
of the IL-2 gene, and IL-2/IL-4 double
knockout (2,3). This suggests that other cytokines may
be involved in acute IL-2-negative anti-donor responses, such as
IL-4, IL-7, IL-9 and IL-15, which can bind to the γ-chain of the
IL-2 receptor (IL-2R) complex (4,5).
IL-15 and IL-2 have similar structures. They share
several biological activities, including stimulation of the
proliferation and differentiation of T cells, and promotion of T
cell chemotaxis (6–8). However, unlike IL-2, IL-15 is derived
from a wide range of cell types, including activated macrophages,
activated vascular endothelial cells, fibroblasts, muscle cells,
and epithelial cells (9).
Moreover, although IL-15 and IL-2 share the same IL-2Rβ and γ-chain
receptor subunits, IL-15 has a unique α-chain, IL-15Rα (6–8). The
role of IL-15 in allograft rejection is not clearly defined,
although important roles for this cytokine have been suggested in
other immunopathologies, such as infectious diseases (10), atherosclerosis (11), human immunodeficiency virus
infection (12), and cancer
(13–15). A number of studies have shown an
increase in the expression of intragraft IL-15 mRNA in both
IL-2-dependent and -independent allograft rejection cases, and
suggested that increased IL-15 expression correlates with acute
rejection (2,3,16,17),
particularly in the case of IL-2-independent rejection. These
findings support the hypothesis that IL-15 may substitute IL-2 in
the mechanism underlying IL-2-independent rejection.
Cyclosporine A (CsA) and tacrolimus (FK506), are two
conventional immunosuppressant drugs widely used in the clinical
setting, which are effective in not only preventing acute
rejection, but also prolonging allograft survival time by
inhibiting IL-2 production. However, the effect of CsA and FK506 on
IL-15 production remains unclear. Although some in vitro
studies have been performed, the results were inconsistent and
contradictory (18–20). To date, no study exists that has
examined the effects of CsA and FK506 on IL-15 expression in
vivo. This study evaluated the effects of treatment with CsA
and FK506 on the alloimmune responses following heart
transplantation in mice, and specifically showed that IL-15
expression is inhibited by both drugs.
Materials and methods
Drugs
Stock solutions of CsA and FK506 were prepared as
follows: CsA was purchased from Novartis Pharmaceuticals (Basel,
Switzerland), and was dissolved in physiological saline to obtain a
working concentration of 0.4 mg/ml. The FK506 stock (Fujisawa
Pharmaceutical, Osaka Japan) was suspended in phosphate-buffered
saline (PBS) to obtain a working concentration of 0.08 mg/ml.
Animals and groups
C57BL/6 (H-2b) and Balb/c (H-2d) mice, 6–8 weeks
old, weighing 18–20 g, were purchased from the Shanghai Laboratory
Animal Center of the Chinese Academy of Sciences, and were housed
in cages inside a room with a light/dark cycle. Heterotopic
intra-abdominal cardiac transplantation was performed with the
method reported by (21), with
some modifications. Briefly, the hearts of the C57 mice were
transplanted into the abdominal cavities of the Balb/c mice, the
aortic ascent artery of the C57 mice was connected to the abdomnal
aortic artery of the Balb/c mice, and the main pulmonary artery of
the C57 mice was connected to the inferior cavae vein of the Balb/c
mice. Balb/c (H-2d) mice that received the transplant were randomly
divided in four groups: syngeneic control group, where donors were
Balb/c mice; allogeneic acute rejection group, where donors were
C57BL/6 mice; allogeneic CsA treatment group, where donors were
C57BL/6 mice and treatment with CsA was performed by
intraperitoneal injection of 10 mg/kg/day CsA from the day of
operation [post-operative day (POD) 0] to POD 13; and allogeneic
FK506 treatment group, where donors were C57BL/6 mice and
recipients were treated with FK506 1.0 mg/kg/day by intraperitoneal
injection from POD 0 to POD 13. Each of these four groups was
subdivided into four groups (n=5) corresponding to PODs 1, 3, 5 and
7 for sample harvesting, and additional subgroups (n=6) for general
observations and measurement of graft survival time. The graft
survival time was assessed daily by palpation of the heart graft.
All experiments were approved by the Animal Welfare Committee and
were performed according to the Laboratory Animal Management
Guidelines of the Zhejiang University.
RNA extraction and reverse transcription
(RT)
For RT-polymerase chain reaction (PCR) analysis,
specimens were snap frozen in liquid nitrogen and stored at −80°C.
Total RNA was extracted with the Gibco® TRIzol reagent
according to the manufacturer’s instructions (Thermo Fisher
Scientific, Waltham, MA, USA). First strand cDNA synthesis was
performed as follows: 4 μg isolated RNA and 3 μl random primers
(Fermentas, Thermo Fisher Scientific, Waltham, MA, USA) were mixed
with double distilled (dd) H2O, in order to obtain a
final volume of 11 μl. The reaction mixture was incubated at 70°C
for 5 min, and at 0°C for 5 min. A total of 5 μl 5× MMLV-RT
reaction buffer, 2 μl 10 mM dNTP, 1 μl ddH20 and 1 μl
MMLV Reverse Transcriptase (Fermentas) was subsequently added and
the samples were incubated at 42°C for 60 min, and at 70°C for 10
min to terminate the reaction. The synthesized cDNA was stored at
−20°C.
PCR analysis
PCR analysis was conducted as follows: synthesized
cDNA (2 μl) was amplified in a 25 μl reaction volume containing
sense and anti-sense primers of each cytokine (Table 1). The reagents were used first to
rule out failure of the reverse transcriptase reaction and PCR
amplification, and second to detect gross variation in cDNA
quantity among the samples. The samples were amplified in a PTC-200
Peltier Thermal cycler (MJ Research, Inc., Watertown, MA, USA). The
conditions were optimized for each primer pair to avoid the
amplification of non-specific products (Table I).
 | Table IOligonucleotide sequences for the
amplification of mouse cytokine genes. |
Table I
Oligonucleotide sequences for the
amplification of mouse cytokine genes.
| Cytokine | Product size
(bp) | Primer sequences | PCR cycling
conditions |
|---|
| IL-2 | 408 | Sense: 5′-AGC TCC ACT
TCA AGC TCT AC-3′
Antisense: 5′-GAC AGA AGG CTA TCC ATC TC-3′ | 94°C, 4 min→94°C, 30
sec, 64°C, 30 sec, 72°C, 30 sec (33 cycles)→72°C, 10 min |
| IFN-γ | 300 | Sense: 5′-TGG GGA CTG
AAG TCC TAG AAG-3′
Antisense: 5′-TTA CCC AGT CAG GGT TAC TGC TGC TGT G-3′ | 94°C, 4 min→94°C, 60
sec, 57°C, 60 sec, 72°C, 60 sec (26 cycles)→72°C, 10 min |
| IL-15 | 345 | Sense: 5′-TCC ATC TCG
TGC TAC TTG TG-3′
Antisense: 5′-CAT TCC TTG CAG CCA GAT TC-3′ | 94°C, 4 min→94°C, 30
sec, 66°C, 30 sec, 72°C, 30 sec (29 cycles)→72°C, 10 min |
| TNF-α | 446 | Sense: 5′-AGC CCA CGT
AGC AAA CCA CCA A-3′
Antisense: 5′-ACA CCC ATT CCC TTC ACA GAG CAA T-3′ | 94°C, 4 min→94°C, 30
sec, 64°C, 30 sec, 72°C, 30 sec (33 cycles)→72°C, 10 min |
| β-actin | 539 | Sense: 5′-GTG GGC CGC
CCT AGG CAC CAA-3′
Antisense: 5′-CTC TTT GAT GTC ACG CAC GAT TTC-3′ | 94°C, 4 min→94°C, 30
sec, 62°C, 30 sec, 72°C, 30 sec (28 cycles)→72°C, 10 min |
Semi-quantitative mRNA analysis
Amplified products and the Fermentas®
pUC19 DNA/MspI (HpAII) marker 23 (Thermo Fisher
Scientific) were analyzed by electrophoresis on 1.5% agarose gels
containing ethidium bromide. Images of the gels were acquired and
the band intensity was analyzed using the Kodak analysis of the gel
image software (Life Technologies, Grand Island, NY, USA). To
correct for variations in the mRNA concentration in each sample,
the densitometry value for each cytokine was divided by the
corresponding value of β-actin.
Protein extraction, titration and
storage
The heart grafts were lysed in ice-cold tissue lysis
buffer, containing 0.25% NP-40, 125 mM KCl, 10 mM MgCl2,
60 mM HEPES (pH 7.9), 0.5 mM DTT, 0.5mM phenylmethylsulfonyl
fluoride, 10 μg/l aprotinin and 10 μg/l leupeptin (all Sangon
Biotech Co., Ltd., Shanghai, China). The cell debris was removed by
centrifugation and the protein-containing supernatant was removed
and titrated according to the specifications of the DC™ Protein
Assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA), and
stored at −80°C.
Western blotting
The proteins were subjected to 15% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, and transferred to
nitrocellulose membranes (EMD Millipore, Billerica, MA, USA). The
membranes were blocked with 5% nonfat dry milk in PBS and incubated
with biotinylated goat anti-mouse IL-15 antibody (1:1,000; R&D
Systems, Minneapolis, MN, USA) or mouse anti-mouse tumor necrosis
factor-α (TNF-α) antibody (1:1,000; Perbio Science AB, Helsingborg,
Sweden) at 4°C overnight. After washing with PBS, the membranes
were incubated with horseradish peroxidase (HRP)-conjugated
anti-goat or anti-mouse immunoglobulin (Ig)G (both at 1:1,500;
Dako, Glostrup, Denmark). The bands were visualized using the
Enhanced Chemiluminescence Blotting system (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The blots were developed on
X-ray film (Eastman Kodak, Rochester, NY, USA).
Histologic evaluation
Heart grafts were excised, covered with formalin,
embedded in paraffin wax, sectioned and stained with hematoxylin
and eosin. They were then observed under a microscope (Leica
DM3000; Leica Microsystems, Wetzlar, Germany).
Statistical analysis
Results were expressed as the mean ± standard
deviation (SD). Statistical analysis was performed with the SPSS
software (SPSS Inc., Chicago, IL, USA). Independent-sample t-tests
and one-way analysis of variance (ANOVA) were used for comparisons
between groups of parametric data. A p-value (P) of <0.05 was
considered to indicate a statistically significant difference.
Results
Heart graft survival time
The heart graft survival times were all longer than
100 days in the control group. In the allogeneic acute rejection
group, the heart grafts stopped working by POD 7–9 (8±0.9 days).
The mean heart graft survival times of the CsA and FK506 treatment
groups were 20.2±4.4 and 17.3±2.1 days, respectively (Table II), which are significantly higher
compared to the allogeneic acute rejection group (P<0.001).
However, the heart graft survival time did not differ significantly
between the CSA and FK506 treatment groups (P=0.1).
| Table IIThe survival time of heart grafts in
the different groups. |
Table II
The survival time of heart grafts in
the different groups.
| Group | n | Survival timea (nb) |
|---|
| A | 8 | 100 (8) |
| B | 7 | 7 (2), 8 (3), 9
(2) |
| C | 6 | 15, 16, 18, 23, 24,
25 |
| D | 6 | 15, 17, 18, 19, 20,
15 |
Histology
No signs of rejection were detected at any
time-point in the control group. Myocardial lymphocyte infiltration
was observed in the heart grafts from PODs 3 to 7 in the allogeneic
acute rejection group. Rejection was markedly suppressed in the
transplanted hearts of the CsA and FK506 treatment groups, with
marked mononuclear infiltration observed (Fig. 1).
 | Figure 1Histological examination of the murine
heart grafts, hematoxylin and eosin staining (original
magnification, ×200). A, syngeneic control group; B, allogeneic
acute rejection group; C, allogeneic cyclosporine A (CsA) treatment
group; D, allogeneic tacrolimus (FK506) treatment group. The
numbers 3 and 5 in each group refer to the post-operative day (POD)
in which the samples were harvested. On POD 3, the heart grafts
have a normal histological appearance and show no sign of rejection
in the syngeneic control, CsA treatment, and FK506 treatment
groups. However, some lymphocyte infiltration and rejection occurs
in the heart grafts of the allogeneic acute rejection group. On POD
5, the heart grafts have a normal histological appearance in the
syngeneic control group, and rejection occurs in the other groups.
Compared to the allogeneic acute rejection group, the degree of
lymphocyte infiltration is markedly reduced in the CsA and FK506
treatment groups. |
Cytokine mRNA expression
The IL-15 mRNA was detected at low levels in
the transplanted hearts of the control group at all postoperative
time-points. Compared to the control group, the IL-15
expression was significantly increased in the allogeneic acute
rejection group on PODs 3, 5 and 7 (P=0.02, P<0.001 and P=0.03,
respectively). The expression of IL-15 mRNA peaked on POD 5
in the allogeneic acute rejection group. In the CsA and FK506
treatment groups, the IL-15 mRNA was detected at low levels
on PODs 1 and 3, but its level increased from PODs 5 to 7. Compared
to the allogeneic acute rejection group, the expression of
IL-15 was significantly inhibited on PODs 3 and 5 in the CsA
and FK506 treatment groups (Fig.
2). The expression pattern of the TNF-α mRNA was similar
to that of IL-15 in all groups (Fig. 3). The IL-2 mRNA level was
undetectable in the control group and significantly increased in
the allogeneic acute rejection group on PODs 3, 5 and 7 (all,
P<0.001), with a peak on POD 5. In the CsA and FK506 treatment
groups, the IL-2 mRNA could not be detected on PODs 1 and 3,
but ts level increased from PODs 5 to 7. Compared to the allogeneic
acute rejection group, the IL-2 expression was significantly
inhibited on PODs 3 and 5 in the CsA and FK506 treatment groups
(Fig. 4).
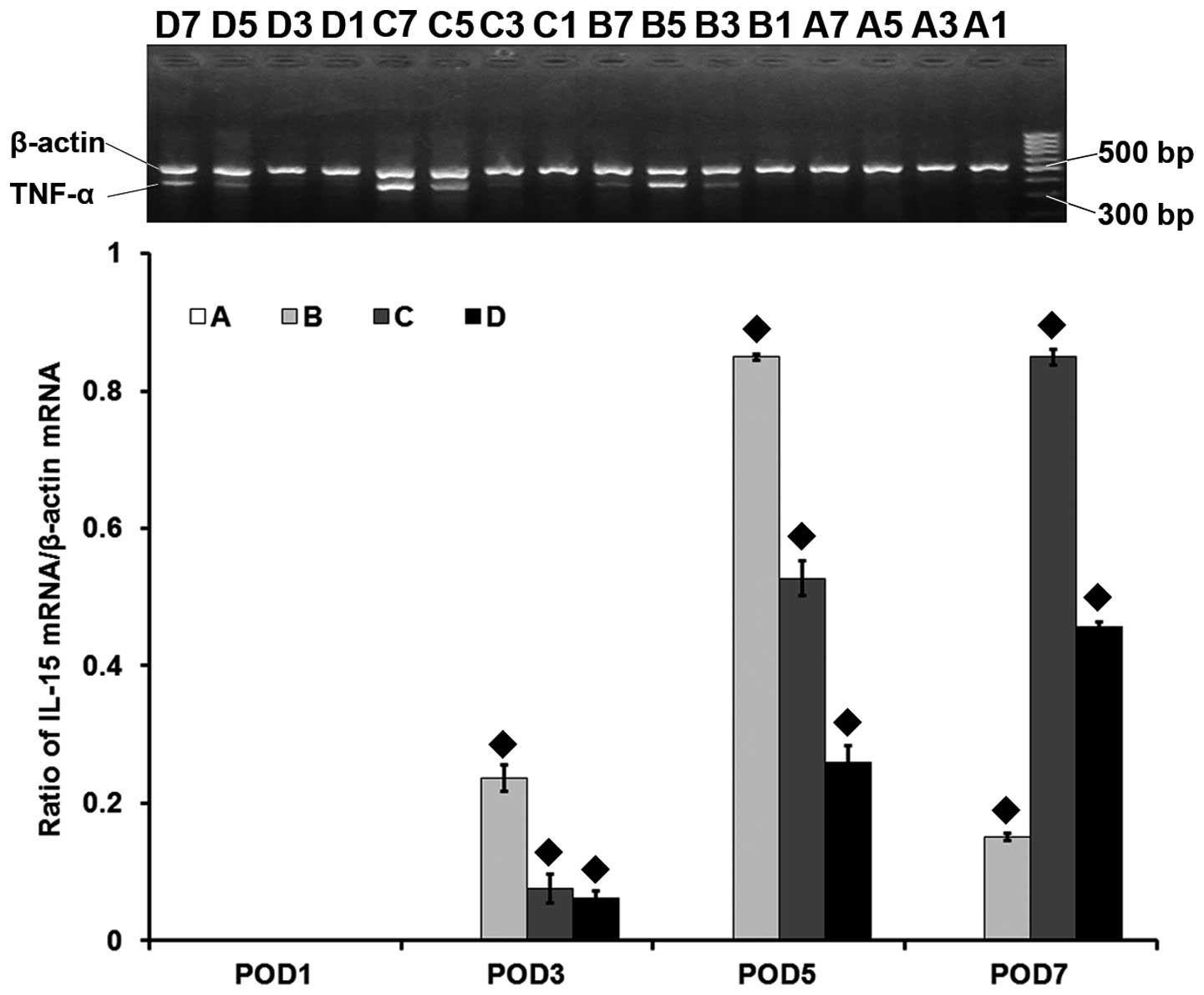
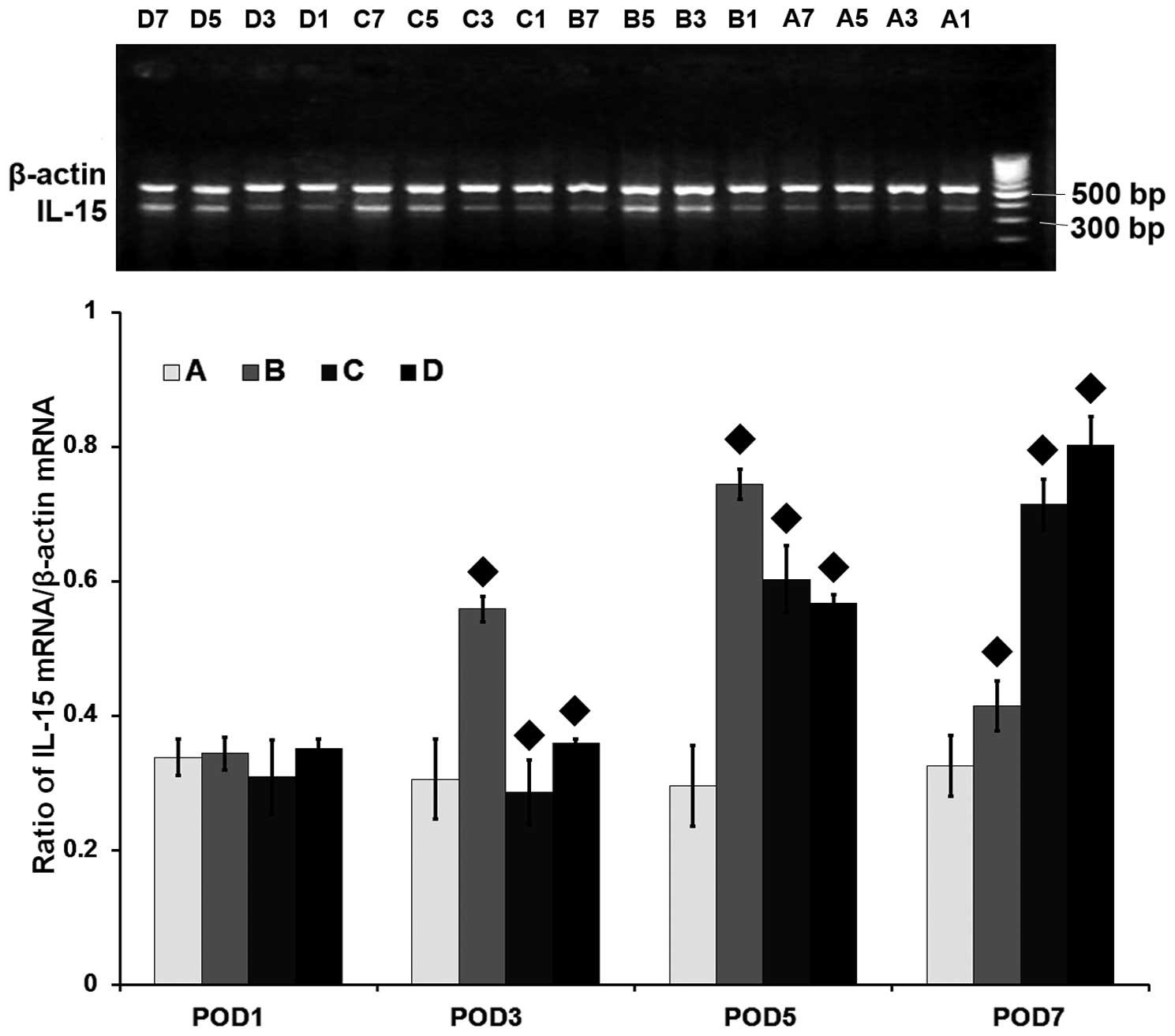 | Figure 2mRNA level of the interleukin-15 gene
(IL-15) assessed by reverse transcription-polymerase chain
reaction in the heart grafts. A, syngeneic control group; B,
allogeneic acute rejection group; C, allogeneic cyclosporine A
(CsA) treatment group; D, allogeneic tacrolimus (FK506) treatment
group. The numbers 1, 3, 5 and 7 in each group refer to the
post-operative day (POD) in which the samples were harvested. ♦
indicates statistical significance (P<0.001) on pairwise
comparison assessed by a t-test. The IL-15 gene is expressed
at low levels in the syngeneic control group at all time-points or
on POD 1 in the other groups. The IL-15 mRNA level is
significantly increased in the allogeneic acute rejection group on
PODs 3, 5 and 7 (all, P<0.05), with its highest level reached on
POD 5. Compared to the allogeneic acute rejection group, the
IL-15 expression in the CsA and FK506 treatment groups is
significantly decreased on PODs 3 and 5 (all, P<0.001), and is
again increased on POD 7 (both, P<0.001). |
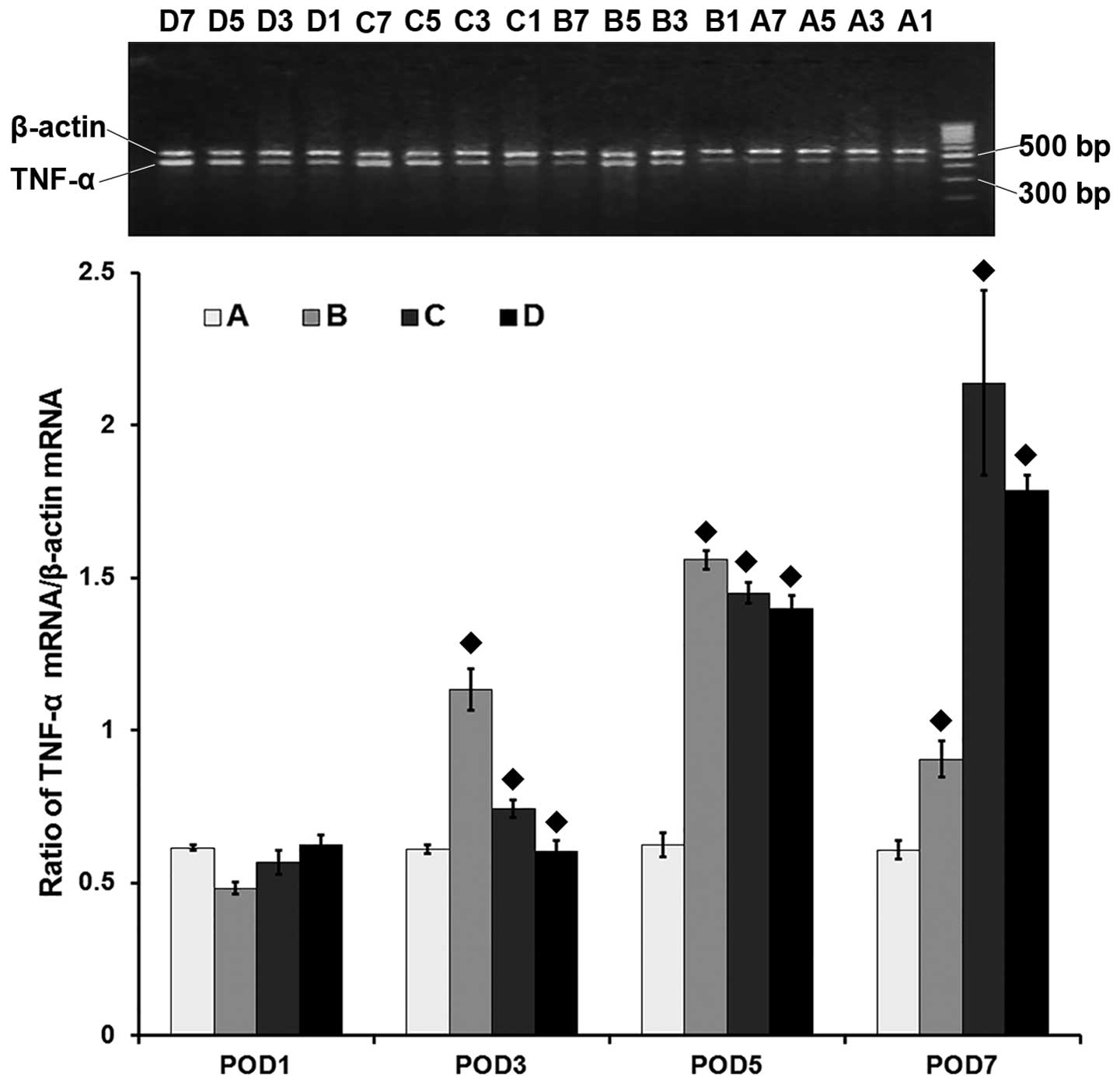 | Figure 3mRNA level of the tumor necrosis
factor-α gene (TNF-α) assessed by reverse
transcription-polymerase chain reaction in the heart grafts. A,
syngeneic control group; B, allogeneic acute rejection group; C,
allogeneic cyclosporine A (CsA) treatment group; D, allogeneic
tacrolimus (FK506) treatment group. The numbers 1, 3, 5 and 7 in
each group refer to the post-operative day (POD) in which the
samples were harvested. ♦ indicates statistical significance
(P<0.001) on pairwise comparison assessed by a t-test. The
TNF-α gene is expressed at low levels in the syngeneic
control group at all time-points or at POD 1 in the other groups.
The TNF-α mRNA level is significantly increased in the
allogeneic acute rejection group on PODs 3, 5 and 7 (all,
P<0.001), with its highest level reached on POD 5. Compared to
the allogeneic acute rejection group, TNF-α expression in
the CsA and FK506 treatment groups is significantly decreased on
PODs 3 and 5 (all, P<0.001), and is again increased on POD 7
(both, P<0.001). |
 | Figure 4mRNA level of the interleukin-2 gene
(IL-2) assessed by reverse transcription-polymerase chain
reaction in the heart grafts. A, syngeneic control group; B,
allogeneic acute rejection group; C, allogeneic cyclosporine A
(CsA) treatment group; D, allogeneic tacrolimus (FK506) treatment
group. The numbers 1, 3, 5 and 7 in each group refer to the
post-operative day (POD) in which the samples were harvested. ♦
indicates statistical significance (P<0.001) on pairwise
comparison assesssed by a t-test. The IL-2 mRNA is not
detected in the syngeneic control group at any time point or on POD
1 in the other groups. Its level is significantly increased in the
allogeneic acute rejection group on PODs 3, 5 and 7 (all,
P<0.001,) reaching its highest level on POD 5. Compared to the
allogeneic acute rejection group, IL-2 expression in the CsA
and FK506 treatment groups is significantly decreased on PODs 3 and
5 (all, P<0.001), and is again increased on POD 7 (both,
P<0.001). |
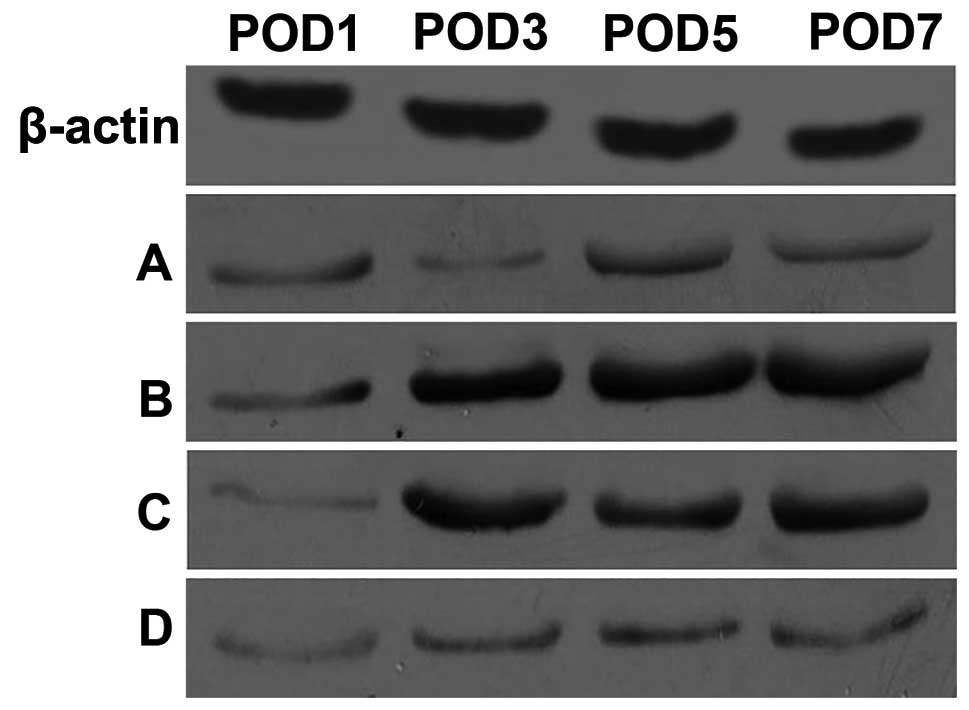
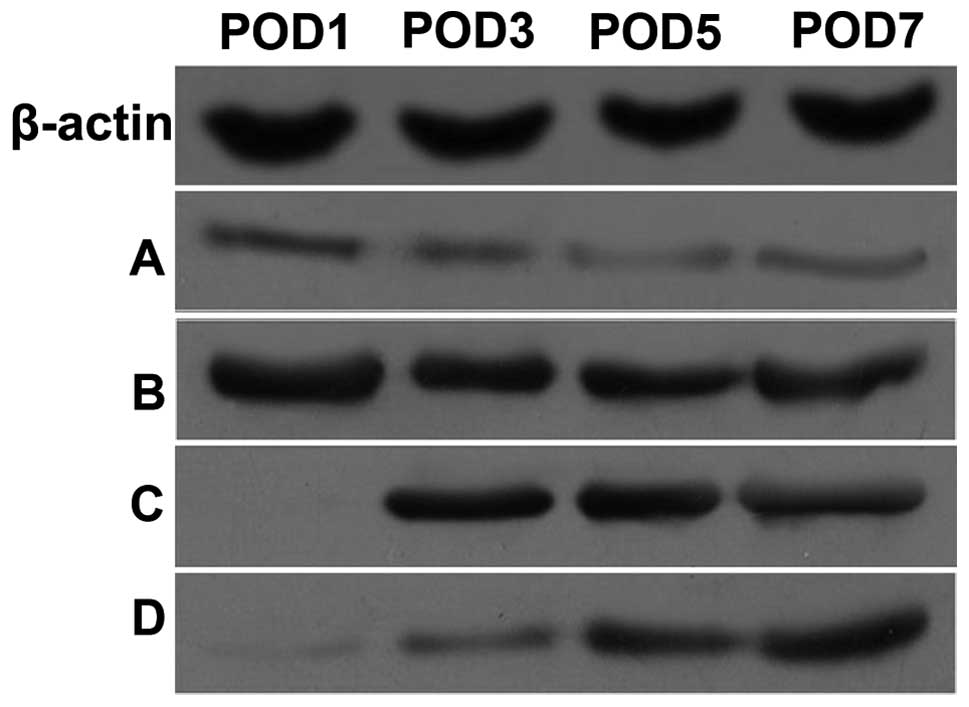
Cytokine protein expression
Low levels of the IL-15 and TNF-α proteins were
detected in the heart grafts of the control group at all
time-points. Compared to the control group, IL-15 protein
expression was significantly increased from PODs 3 to 7, and TNF-α
protein expression was increased from PODs 1 to 7 in the heart
grafts of the allogeneic acute rejection group. In the CsA and
FK506 treatment groups, despite the gradual and slight increase
from PODs 3 to 7, the expression of IL-15 (Fig. 5) and TNF-α (Fig. 6) was reduced at all time-points
relative to the allogeneic acute rejection group.
Discussion
IL-15, a T cell growth factor, is derived from
non-lymphocytes, such as activated macrophages, activated vascular
endothelial cells, fibroblasts, and muscle cells. It shares a
number of common biological activities with IL-2, including
stimulation of T cell, B cell, and natural killer cell
proliferation, as well as a T cell chemoattractant activity
(6–8). Recent studies showed that IL-15
expression is increased in certain types of immune-mediated tissue
injury, such as allograft rejection (4,5,10–12,15).
There is considerable interest in the potential role of this
cytokine in the pathogenesis of tissue destruction, particularly in
the context of alloimmune reactions.
In the present study, the level of the IL-15
mRNA was low but still detectable in non-rejected heart grafts in
the syngeneic control group, whereas it was significantly increased
in rejected heart grafts in the acute rejection group. The low
level of IL-15 mRNA expressed may be attributed to cardiac
muscle cells, which produce low levels of IL-15 (22). The increased level of IL-15
mRNA is likely primarily derived from the infiltrating activated
monocytes/macrophages in the rejected heart graft, since these
cells produce high levels of IL-15 (22). Numerous monocyte/macrophage
infiltrates are observed in acutely rejected allografts (23,24).
Unlike IL-15, the expression of IL-2 was not detected
in non-rejected heart grafts and was significantly increased in
rejected heart grafts. The difference between the expression
patterns of IL-15 and IL-2 may be due to the origin
of these two cytokines.
Increased IL-15 expression appeared
associated with the presence of acute rejection in the present
study. The strongest expression of the IL-15 gene was
observed on POD 5, the same day at which lymphocyte infiltration
peaked in the rejected heart grafts according to the pathological
examination.
It is well known that both CsA and FK506 can inhibit
IL-2 expression and prolong allograft survival time by blocking
calcineurin (25,26). However, the relationship between
IL-15 expression and the administration of CsA or FK506 is rarely
reported, and only three in vitro studies have been
performed (18–20); two of these (19,20)
reported that the production of IL-15 is unaffected by CsA, but one
(18) reported that CsA
administration decreases the level of IL-15 in a dose-dependent
manner. The contradictory results between these studies may be due
to the different cell lines used. In the present study, we
demonstrated that IL-15 expression, at both the mRNA and the
protein level, is reduced by CsA or FK506 treatment (Figs. 2 and 5). However, we observed a discordance
between IL-15 mRNA and protein expression. In the allogeneic acute
rejection group, the IL-15 mRNA level was increased on POD
3, peaked on POD 5, and decreased on POD 7, although the expression
of IL-15 was not completely blocked. Notably, on POD 7, the
IL-15 expression in the heart grafts treated with CsA and
FK506 increased again. This finding may be related to the dose of
the immunosuppressants used in this study. It is likely that CsA or
FK506 induce a decrease in the level of IL-15 in a
dose-dependent manner in vivo, and increased doses of CsA or
FK506 may result in complete inhibition of IL-15. However,
the mechanism by which CsA or FK506 inhibit the expression of
IL-15 remains unclear. In addition, the present study showed
that the TNF-α mRNA derived from activated
monocytes/macrophages is significantly reduced upon administration
of CsA and FK506. This finding indicates that the activity of
macrophages may be suppressed in these conditions. We conclude that
CsA and FK506 may affect the production of IL-15 via interactions
with the macrophages.
In conclusion, IL-15, a non-T cell-derived cytokine,
is involved in acute rejection following heart transplantation and
is partially downregulated in vivo in mice by CsA and FK506
treatment. Whether increased doses of CsA or FK506 may result in
the complete inhibition of IL-15 production requires further study,
while it is also necessary to clarify the relationship between CsA
or FK506 treatment and IL-15 expression in clinical studies.
Acknowledgements
This study was supported by grants from the Team
Program of Science and the Health Bureau of the Zhejiang Province
Foundation (B1652).
References
|
1
|
van Gelder T, Baan CC, Balk AH, et al:
Blockade of the interleukin (IL)-2/IL-2 receptor pathway with a
monoclonal anti-IL-2 receptor antibody (BT563) does not prevent the
development of acute heart allograft rejection in humans.
Transplantation. 65:405–410. 1998.PubMed/NCBI
|
|
2
|
Li XC, Roy-Chaudhury P, Hancock WW, et al:
IL-2 and IL-4 double knockout mice reject islet allografts: a role
for novel T cell growth factors in allograft rejection. J Immunol.
161:890–896. 1998.PubMed/NCBI
|
|
3
|
Steiger J, Nickerson PW, Steurer W,
Moscovitch-Lopatin M and Strom TB: IL-2 knockout recipient mice
reject islet cell allografts. J Immunol. 155:489–498.
1995.PubMed/NCBI
|
|
4
|
Lewis EC, Weiler M, Tejman-Yarden N, et
al: Involvement of graft-derived interleukin-15 in islet allograft
rejection in mice. Cytokine. 34:106–113. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zheng XX, Gao W, Donskoy E, et al: An
antagonist mutant IL-15/Fc promotes transplant tolerance.
Transplantation. 81:109–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ikemizu S, Chirifu M and Davis SJ: IL-2
and IL-15 signaling complexes: different but the same. Nat Immunol.
13:1141–1142. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ring AM, Lin JX, Feng D, et al:
Mechanistic and structural insight into the functional dichotomy
between IL-2 and IL-15. Nat Immunol. 13:1187–1195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Waldmann TA: The IL-2/IL-15 receptor
systems: targets for immunotherapy. J Clin Immunol. 22:51–56. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grabstein KH, Eisenman J, Shanebeck K, et
al: Cloning of a T cell growth factor that interacts with the beta
chain of the interleukin-2 receptor. Science. 264:965–968. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Di Sabatino A, Calarota SA, Vidali F,
MacDonald TT and Corazza GR: Role of IL-15 in immune-mediated and
infectious diseases. Cytokine Growth Factor Rev. 22:19–33.
2011.PubMed/NCBI
|
|
11
|
van Es T, van Puijvelde GH, Michon IN, et
al: IL-15 aggravates atherosclerotic lesion development in LDL
receptor deficient mice. Vaccine. 29:976–983. 2011.PubMed/NCBI
|
|
12
|
d‘Ettorre G, Andreotti M, Ceccarelli G, et
al: The role of IL-15 in challenging Acquired Immunodeficiency
Syndrome. Cytokine. 57:54–60. 2012.PubMed/NCBI
|
|
13
|
Croce M, Orengo AM, Azzarone B and Ferrini
S: Immunotherapeutic applications of IL-15. Immunotherapy.
4:957–969. 2012. View Article : Google Scholar
|
|
14
|
Roberti MP, Rocca YS, Amat M, et al: IL-2-
or IL-15-activated NK cells enhance Cetuximab-mediated activity
against triple-negative breast cancer in xenografts and in breast
cancer patients. Breast Cancer Res Treat. 136:659–671. 2012.
View Article : Google Scholar
|
|
15
|
Steel JC, Waldmann TA and Morris JC:
Interleukin-15 biology and its therapeutic implications in cancer.
Trends Pharmacol Sci. 33:35–41. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baan CC, van Riemsdijk-Overbeeke IC,
Boelaars-van Haperen MJ, Ijzermans JM and Weimar W: Inhibition of
the IL-15 pathway in anti-CD25 mAb treated renal allograft
recipients. Transpl Immunol. 10:81–87. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Smith XG, Bolton EM and Bradley JA:
Targeting IL-15 as a therapeutic strategy in organ transplant
rejection. Curr Opin Investig Drugs. 3:406–410. 2002.PubMed/NCBI
|
|
18
|
Cho ML, Kim WU, Min SY, et al:
Cyclosporine differentially regulates interleukin-10,
interleukin-15, and tumor necrosis factor α production by
rheumatoid synoviocytes. Arthritis Rheum. 46:42–51. 2002.PubMed/NCBI
|
|
19
|
Lewis E, Weiler M, Chaimovitz C and
Douvdevani A: Interleukin-15 is the main mediator of lymphocyte
proliferation in cultures mixed with human kidney tubular
epithelial cells. Transplantation. 72:886–890. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stoeck M, Schäfer M, Hofmann HP and
Gekeler V: Dexamethasone and cyclosporin A do not inhibit
interleukin-15 expression in the human lung carcinoma cell line
A549. Eur Cytokine Netw. 11:414–419. 2000.PubMed/NCBI
|
|
21
|
Ono K and Lindsey ES: Improved technique
of heart transplantation in rats. J Thorac Cardiovasc Surg.
57:225–229. 1969.PubMed/NCBI
|
|
22
|
Giri JG, Ahdieh M, Eisenman J, et al:
Utilization of the beta and gamma chains of the IL-2 receptor by
the novel cytokine IL-15. EMBO J. 13:2822–2830. 1994.PubMed/NCBI
|
|
23
|
Hubscher SG: Histological findings in
liver allograft rejection - new insights into the pathogenesis of
hepatocellular damage in liver allografts. Histopathology.
18:377–383. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
McCaughan GW, Davies JS, Waugh JA, et al:
A quantitative analysis of T lymphocyte populations in human liver
allografts undergoing rejection: the use of monoclonal antibodies
and double immunolabeling. Hepatology. 12:1305–1313. 1990.
View Article : Google Scholar
|
|
25
|
Wiederrecht G, Lam E, Hung S, Martin M and
Sigal N: The mechanism of action of FK-506 and cyclosporin A. Ann
NY Acad Sci. 696:9–19. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamamoto S and Kato R: Hair
growth-stimulating effects of cyclosporin A and FK506, potent
immunosuppressants. J Dermatol Sci. (Suppl): S47–S54. 1994.
View Article : Google Scholar : PubMed/NCBI
|