Introduction
Hepatocellular carcinoma (HCC) is one of the most
prevalent malignant cancers and a major cause of mortality
worldwide, most notably in East Asia (1); however, the incidence rate of HCC is
rising in Western countries (2).
Despite the development of novel therapies for HCC patients,
prognoses remain poor; this may be due to the molecular
abnormalities of cancer cells. Identification of the genes and
proteins which regulate liver carcinogenesis is critical for the
exploration of novel targeted therapies for HCC.
Yes-associated protein (YAP), the downstream target
molecule of the Hippo signaling pathway, is a transcription
co-activator which cooperates with the transcriptional enhancer
activator domain factor. Overexpression of YAP has been reported to
aberrantly activate the target gene connective-tissue growth factor
(3) as well as induce the
proliferation of cancer cells (4).
As an oncogene (5), YAP protein
levels were shown to be elevated and localized in the nuclei of HCC
tissues (4). In addition, YAP
messenger (m)RNA levels were reported to be significantly elevated
in the majority of HCC tissues compared with those in adjacent
non-tumor tissues (6,7).
Tumor cell motility is a key determinant of HCC
progression and proliferation. YAP has been shown to have a
significant role in HCC cell motility via its role in the Hippo
signaling pathway. In addition, large tumor suppressor (LATS)1, an
inhibitor of YAP activity (8), was
found to decrease the motility of human hepatocellular carcinoma
HepaRG cells in vitro (9).
Furthermore, it has been reported that mice deficient in LATS1
developed soft-tissue sarcomas as well as ovarian stromal cell
tumors and were highly sensitive to carcinogens (10). These studies therefore suggested
that YAP acted as an oncogene, while LATS1 acted as a tumor
suppressor gene. It has been hypothesized that mutations associated
with LATS1 may occur in numerous HCC cells; therefore, YAP and
LATS1 may be promising therapeutic targets for the treatment of
HCC.
Two human HCC cell clones with high and low
metastatic potential, MHCC-97H (97H) and MHCC-97L (97L), derived
from the parental cell line MHCC97, have previously been
established (11,12). However, the YAP expression levels
and proliferation rates of these two clones of the same genetic
background have not been examined.
A previous study based on a Chinese cohort in Hong
Kong revealed that YAP was an independent prognostic marker for
overall and disease-free survival times in HCC patients (6). Another previous study showed that YAP
mRNA and protein levels were significantly higher in HCC tissues
compared with those in para-cancerous tissue (PCT) (7). Numerous HCC patients were more
amenable to surgery; however, these patients still had a poor
prognosis due to early recurrence following partial hepatectomy.
YAP levels in the resected HCC tissue may therefore provide a
valuable indicator for effective follow-up management. Further
studies on regulation of the Hippo pathway may enhance
understanding of hepatocarcinogenesis; in addition, the development
and use of a targeted therapy against the YAP gene may enable
long-term survival for patients with HCC.
Among the above-mentioned issues, it is important to
confirm whether knockdown of YAP using RNAi significantly inhibited
liver cancer cell growth; therefore, as YAP was found to be
associated with LATS1 in the Hippo pathway, the aim of the present
study was to measure the expression of LATS1 in cancer cells in
which YAP was downregulated.
Materials and methods
Patients and specimens of HCC
A total of 40 cases of HCC, and 10 cases of PCT were
used in the present study. All tissues were obtained from the
patients which received curative hepatectomy surgery at the Air
Force General Hospital (Beijing, China) between January 2010 and
June 2013. All patients were diagnosed with pathological primary
HCC and none of the patients had received any radiotherapy or
chemotherapy prior to surgery. The PCT was dissected at a margin
>1 cm from the tumor edge. Normal liver samples were taken from
benign lesions.
All clinical procedures in the present study were
approved by an institutional review board of the Air Force General
Hospital (Beijing, China) prior to patient enrollment. Written
informed consent was obtained from each patient prior to the
collection of these tissues.
Generation of HCC tissue array
A tissue microarray (TMA) was constructed from
formalin-fixed (10% paraformic aldehyde; Sigma-Aldrich, St. Louis,
MO, USA), paraffin- embedded (Weiqiboxing Co., Wuhan, China) tissue
blocks. Two core needle samples, 1.2 mm from each specimen, were
obtained from morphologically representative tumor areas of each
donor tissue paraffin block. Xylene (Beijing Chemwork, Beijing,
China) was used for dewaxing, graded ethanol (100% 10 sec and 95%
10 sec; Hongziweida Co., Beijing, China) was used for rehydration
of the samples and neutral balsam (Sinopharm Chemical Reagent Co.,
Ltd, Shanghai, China). Hematoxylin and eosin staining (Berlin
Biological, Beijing, China) of the TMAs was performed in order to
verify the histopathological findings.
Cell lines
HCC cell lines 97H (higher metastatic potential) and
97L (lower metastatic potential) were purchased from the Cell Bank
of Shanghai Institute (Shanghai, China; these cell lines were
established in nude mice). All cells were cultured in Dulbecco’s
modified Eagle medium with 10% fetal bovine serum supplemented with
100 U/ml penicillin and 100 μg/ml streptomycin.
Immunohistochemistry (IHC) and
immunocytochemistry (ICC)
Expression levels of YAP and LATS1 were measured
using IHC in a retrospective cohort of liver cancer and matched
adjacent non-tumor tissue samples from HCC patients following liver
resection. The expression of YAP was also measured using ICC in
MHCC97H cells.
Paraffin-embedded tissue blocks were cut into
4-μm sections and each was baked at 60°C for 2 h prior to
deparaffinization. Antigen retrieval was achieved through microwave
exposure (Milk 700 W; Galanz, Guangdong, China) at 100°C in citrate
buffer (0.01 M; Bioss, Inc., Wuhan, China) for 15 min. Following
immersion in 3% hydrogen peroxide (Lircon, Sandong, China) for 20
min, the sections and cells were incubated with goat serum (Yuanye
Co., Shanghai, China) for 1 h and then incubated with YAP (H-125)
rabbit polyclonal immunoglobulin (Ig)G (1:80; sc-15407; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), LATS1 rabbit polyclonal
antibodies (1:80; 17049-1-AP; Proteintech Group, Chicago, IL, USA),
anti-phospho-LATS1 (Ser909) rabbit polyclonal antibodies (1:80;
bs-3246R; Bioss, Inc., Wuhan, China) and GAPDH antibody (1:3,000;
GenScript, Piscataway, NJ, USA) at 4°C overnight. Following four
washes in phosphate-buffered saline with Tween 20 (Sigma-Aldrich),
sections were incubated with peroxidase-conjugated goat
anti-rabbit/mouse IgG secondary antibody (K5007 Bottle A, Dako
REAL™ EnVision™; Dako, Glostrup, Denmark) for 1 h at 37°C.
Visualization was performed using a diaminobenzidine chromogen kit
(K5007 Bottles B and C, Dako REAL™ Substrate Buffer and Dako REAL™
DAB+ Chromogen; Dako). Sections were then counterstained with
hematoxylin, dehydrated and mounted.
Experiments were repeated twice and the percentage
of tumor cells or hepatocytes with obvious staining in the
cytoplasm or nucleus were determined in all optical fields of the
slices (BX51TR; Olympus Corp., Tokyo, Japan). A percentage >10%
was considered to indicate positive immunoreactivity. Two
independent, blinded investigators examined all tumor slides at
random.
Lentivirus production and
transduction
The sequence for targeting the YAP gene was selected
using the short hairpin RNA (shRNA) Clone Library [http://cgap.nci.nih.gov/RNAi/RNAi2]. The
effective target point sequence of YAP gene YAP1 NM_001130145
Human, 5′-CATTGCTGCTGTTAATGTA-3′, was selected. A lentivirus
(pMagic 4.1) construct was provided by Shanghai Sunbio Medical
Biotechnology Co. Ltd (SB1262-C; Shanghai, China).
97H cells were transduced with lentiviral vectors
and control lentivirus in complete medium containing polybrene (8
mg/ml; Sigma-Aldrich). At 72 h following the first transfection,
total RNA and protein were extracted from cells with
radioimmunoprecipitation lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) with a protease inhibitor (Roche,
Basel, Switzerland) and reverse transcription quantitative
polymerase chain reactions (RT-qPCR) as well as western blot
analysis were performed in order to evaluate the inhibitory effects
of YAP.
Flow cytometric analysis of the cell
cycle
Cells were harvested during the exponential growth
phase and single-cell suspensions containing 1×106 cells
were fixed with 75% ethanol (Hongziweida Co.) for 48 h at 4°C. The
cell cycle was monitored using propidium iodide (PI; 50
μg/ml; Sigma-Aldrich, St. Louis, MO, USA) staining. The
fluorescence of DNA-bound PI in cells was measured using a flow
cytometer (FACS101; BD Biosciences; Franklin Lakes, NJ, USA).
In vitro cell growth assay
Cell Titer 96 AQueous One Solution Cell
Proliferation Assay (3582; Promega Corp., Madison, WI, USA)
containing 3-(4,5-dimethylthiazol-2-yl)-
5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS)
was used to assess cell proliferation, as previously described
(13).
A cell suspension was prepared at a concentration of
1×104/ml. Aliquots (100 μl) were dispensed into
96-well microtiter plates and incubated for five days. MTS assays
were performed by adding 20-μl MTS reagent into each well
and incubating the plate at 37°C for 3 h in a humidified, 5%
CO2 atmosphere. The optical density (OD) of each well
was measured daily using a microplate reader (550; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) at 490 nm; all OD values
were calculated relative to OD at day 1.
RT-qPCR
Total RNA was extracted using TRIzol (1382739;
Invitrogen Life Technologies, Carlsbad, CA, USA). Total RNA
concentrations were measured using spectrophotometry (DU-7; Beckman
Coulter, Inc., Fullerton, CA, USA) and then 300 ng total RNA from
each sample was reverse-transcribed to complementary (c)DNA using
the PrimeScript RT reagent kit (DRR037A; TaKaRa Bio, Inc., Shiga,
Japan). cDNA was amplified by qPCR using SYBR Green (DRR041A;
TaKaRa Bio, Inc.). The amplification process was applied in a
LightCycler 2.0 system with LightCycler software version 3.5
(Roche). Primer details are listed in Table I. Amplification conditions were as
follows: 95°C for 30 sec, then 40 cycles at 95°C for 5 sec and 64°C
for 30 sec. A dissociation procedure was performed to generate a
melting curve for confirmation of amplification specificity. The
analysis of PCR products by 1% agarose gel electrophoresis
(Aladdin, Shanghai, China) confirmed amplification specificity.
Primers were designed by the Sangon Biotech Co. (Shanghai, China).
Primer specificity was confirmed using Primer BLAST software
(National Center for Biotechnology Information, Bethesda, MD, USA).
Relative quantification was accomplished using a double standard
curve method or the 2−ΔΔCt method, as previously
described (14). Ct values of the
samples were normalized to the appropriate endogenous housekeeping
gene GAPDH. Each measurement was repeated in triplicate.
 | Table IPrimers used for reverse transcription
quantitative polymerase chain reaction. |
Table I
Primers used for reverse transcription
quantitative polymerase chain reaction.
| Gene | Primer | Length (bp) |
|---|
| Yes-associated
protein (Human) | Forward,
5′-ACCCACAGCTCAGCATCTTCG-3′
Reverse, 5′-TGGCTTGTTCCCATCCATCAG-3′ | 257 |
| GAPDH (Human) | Forward,
5′-AGAAGGCTGGGGCTCATTTG-3′
Reverse, 5′-AGGGGCCATCCACAGTCTTC-3′ | 258 |
Western blot analysis
Cells were lysed using radioimmunoprecipitation
buffer (P0013B; Beyotime Institute of Biotechnology) with a
protease inhibitor (11873580001; Roche). A bicinchoninic acid assay
(P0010; Beyotime Institute of Biotechnology) was used to determine
protein concentrations, as previously described (15). A total of 40 μg protein was
boiled for 10 min prior to loading and then separated using 10%
SDS-PAGE. Proteins were then transferred onto nitrocellulose
membranes (0.45 μm; Whatman Ltd, Clifton, NJ, USA) using a
semi-dry transfer system (Bio-Rad Laboratories, Inc.). Following
blocking with 5% bovine serum albumin (HyClone, GE Healthcare Life
Sciences, Logan, UT, USA), the membrane was probed with YAP (H-125)
rabbit polyclonal immunoglobulin (Ig) G (1:300), LATS1 rabbit
polyclonal antibody (1:200), β-actin antibody (1:500, Immunocreate,
Hoover, AL, USA) and GAPDH antibody (1:3,000; GenScript).
Horseradish peroxidase (HRP)-conjugated goat anti-rabbit polyclonal
secondary antibody (1:40,000; 31460, Thermo Fisher Scientific,
Waltham, MA, USA) and HRP-conjugated goat anti-mouse polyclonal IgG
(1:40,000; 115-035-003, Jackson Immunoresearch Laboratories Inc.,
West Grove, PA, USA) were then used. Protein band signals were
amplified using enhanced chemiluminescence detection reagents
(34079; Thermo Fisher Scientific). Protein levels were determined
semi-quantitatively using Quantity One 4.4.0 imaging software
(Bio-Rad Laboratories, Inc.).
Statistical analysis
The Pearson Chi-Square test and the independent
sample t-test were used to analyze differences between
values. All statistical analyses were performed using the SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference between
values.
Results
Compared with adjacent tissues, HCC
tissues express higher levels of YAP and lower levels of LATS1
YAP and LATS1 protein expression levels were
investigated in 40 HCC tissue samples and 10 PCT tissue samples
using IHC. As shown in Fig. 1A,
the positivity rate of YAP protein in HCC tissue was significantly
higher compared with that of PCT (72.5 and 20.0%, respectively;
P=0.002). In addition, compared with adjacent tissues, HCC tissues
expressed higher levels of YAP protein; furthermore, YAP was found
to be concentrated in the nuclei of HCC cells. The positivity rate
of LATS1 protein in HCC tissue was significantly lower compared
with that of PCT (17.5 and 60.0%, respectively; P=0.019). Compared
with the adjacent tissues, HCC tissues had lower levels of LATS1
protein, which was predominantly located in the cytoplasm of PCT
cells (Fig. 1B).
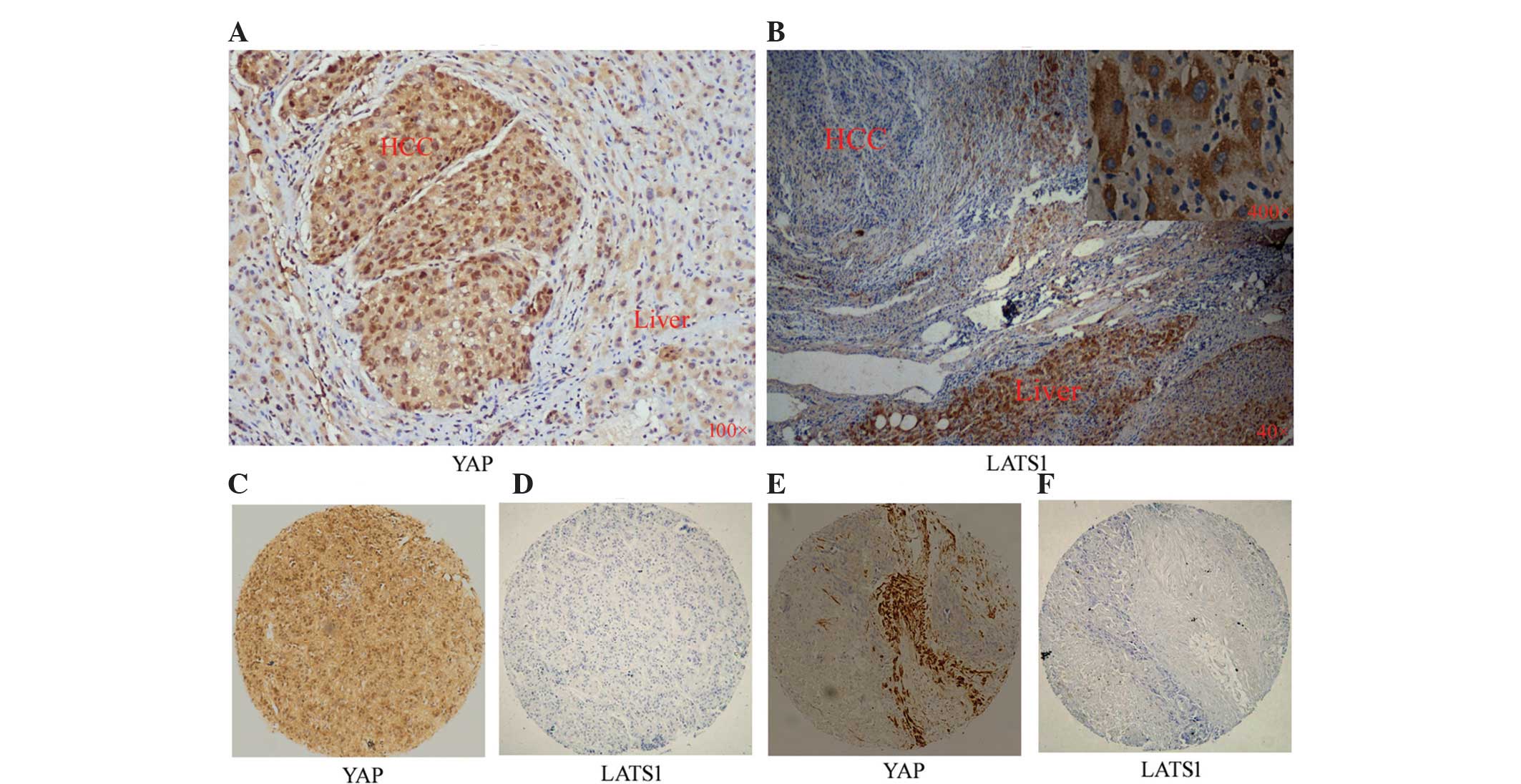 | Figure 1Expression of YAP and LATS1 in HCC and
PCT tissues. Representative images of (A) YAP staining
(magnification, ×100) or (B) LATS1 staining in HCC (magnification,
×40) and PCT (magnification, ×40 and ×400) as detected by
immunohistochemistry. (C and D) Identical HCC tissue slice stained
for YAP and LATSI, respectively. (E and F) Another identical HCC
tissue slice stained for YAP and LATSI, respectively
(magnification, ×40). YAP, yes-associated protein; LATS, large
tumor suppressor 1; HCC, hepatocellular carcinoma; PCT,
para-cancerous tissue. |
In HCC tissues, high levels of YAP are
usually accompanied by low levels of LATS1
The results of the present study also showed that
overexpression of YAP occurred primarily in HCC tissues in which
LATS1 expression was low (Fig.
1C–F). The positive rate of YAP expression demonstrated a
negative correlation with that of LATS1 expression in HCC (72.5 and
17.5%, respectively; P=0.016) (Table
II).
| Table IIAssociation between YAP and LATS1 in
hepatocellular carcinoma. |
Table II
Association between YAP and LATS1 in
hepatocellular carcinoma.
| YAP
| χ2 | P-value |
|---|
| P | N |
|---|
| LATS1 | P | 2 | 5 | 5.759 | 0.016 |
| N | 27 | 6 | | |
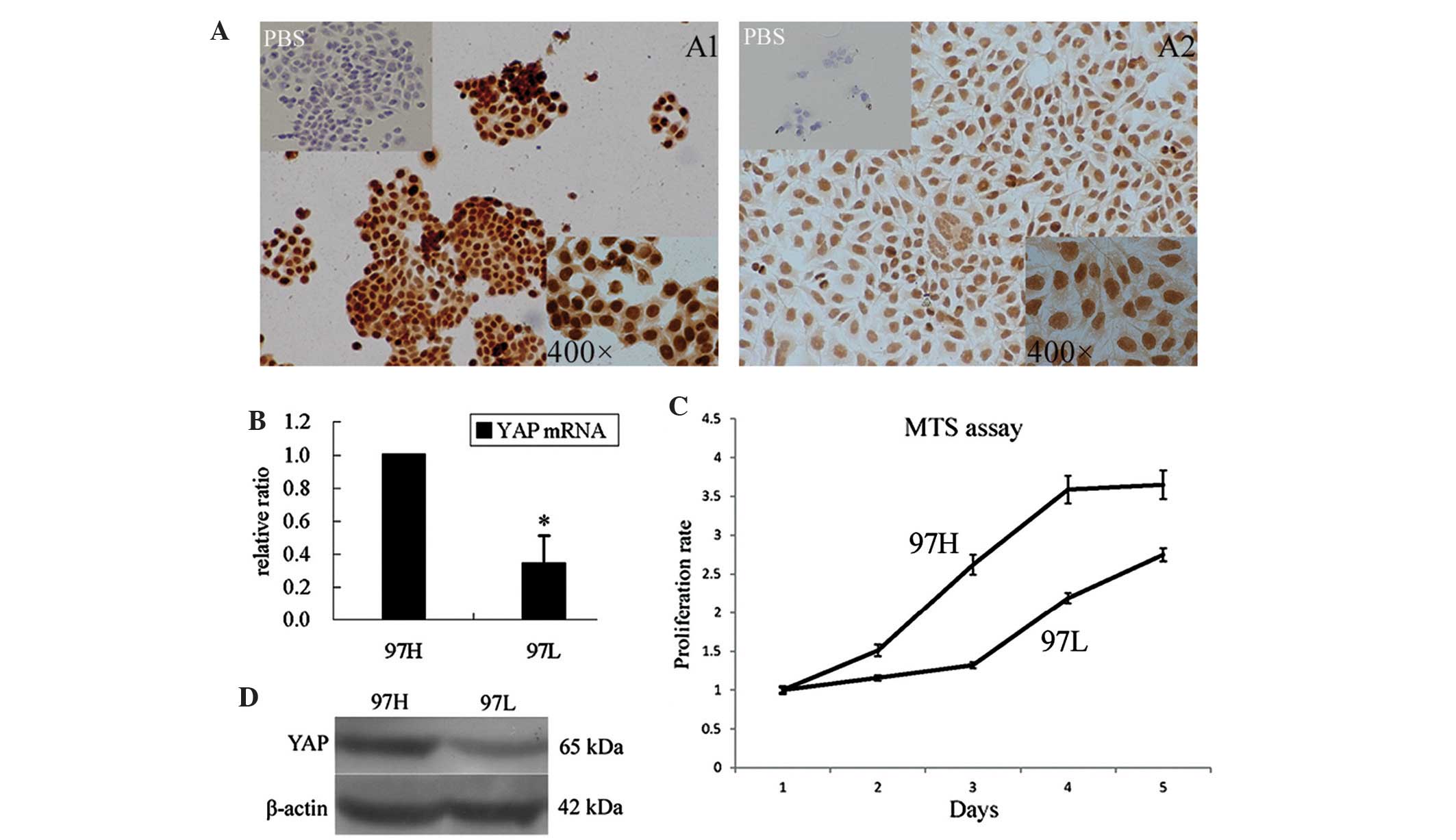
MHCC97 cells with high YAP expression
have an increased proliferative capacity
YAP expression in two liver cancer cell lines with
different metastatic properties were examined using ICC and western
blot analysis (Fig. 2A and D,
respectively). The results showed that 97H cells aggregated during
growth, whereas 97L cells spread out during growth. 97H cells,
which are highly metastatic, expressed higher protein levels of YAP
compared with those of 97L cells. As shown in Fig. 2B, 97H cells expressed higher levels
of YAP mRNA compared with those in 97L cells (P=0.021). Of note,
the results of the MTS assay showed that the proliferation of 97H
cells was significantly higher compared with that of 97L cells
(Fig. 2C); however, 97 L cells
also grew rapidly. The proliferation multiples of 97H and 97L on
day five were 3.65±0.06 and 2.75±0.03, respectively (P<0.05).
Therefore, the proliferation activity of these liver cancer cells
paralleled the expression levels of YAP.
RNAi-mediated YAP gene silencing inhibits
97H cell proliferation and cell cycle
Lentivirus-mediated RNAi was used to efficiently
downregulate YAP expression in 97H cells as shown in the
fluorescence photomicrograph images (Fig. 3A), as well as by the significantly
decreased mRNA (Fig. 3B) and
protein (Fig. 4A) levels of YAP
following shRNA YAP (shYAP) transfection compared with that of the
shRNA control-transfected (shControl) cells. The results showed
that cell proliferation was significantly suppressed and cell cycle
progression was altered in shYAP-transfected 97H cells (Fig. 3C and D). The proliferation
multiples of shYAP-transfected cells and shControl cells on day
four following transfection were 2.42±0.15 and 3.45±0.11,
respectively. Cell cycle analysis of 97H cells using flow cytometry
revealed that the proportion of shYAP-transfected cells in the G1,
G2 and S phases were 50.33, 13.94 and 35.73%, respectively, whereas
the proportion in each cell cycle phase in shControl cells were
33.67, 0.94 and 65.39%, respectively; the differences between the
two groups were significant for all phases (P<0.001). The
proportion of cells in the G1 and G2 phase was significantly
increased in shYAP-transfected cells, whereas the proportion cells
in S phase was decreased as compared with populations of shControl
cells. This therefore indicated that YAP silencing induced G1 and
G2 phase arrest as well as a decrease of cells in S phase.
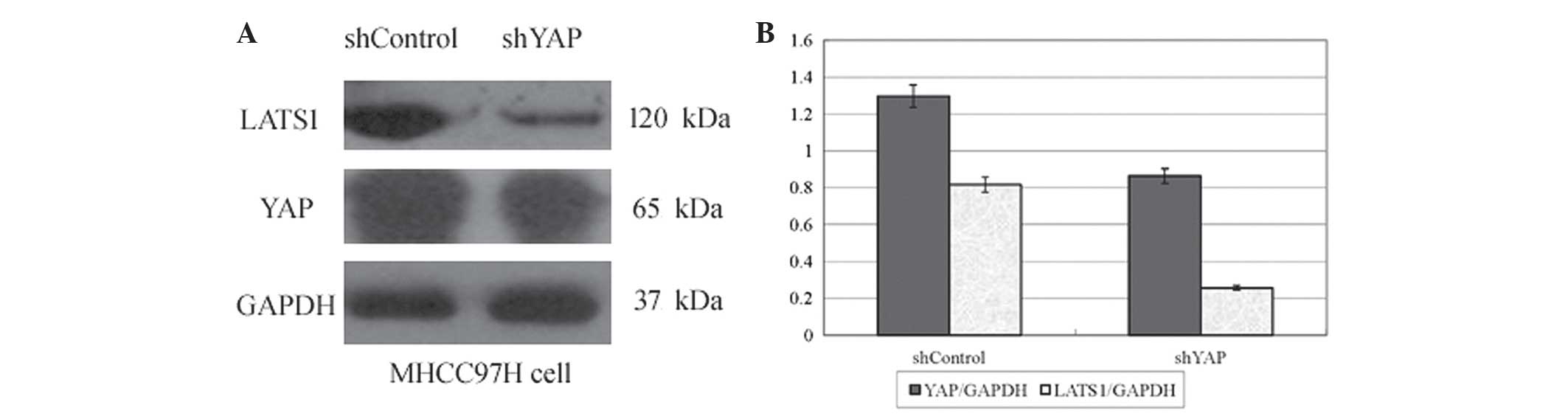
YAP and LATS1 are downregulated in
shYAP-transfected 97H cells
As shown in Fig. 4,
western blot analysis revealed that protein levels of YAP and
LATS1, the main upstream regulator of YAP, were significantly
decreased in shYAP-transfected cells compared with those in the
shControl group. Downregulation of YAP protein levels was
accompanied by downregulation of LATS1 protein levels (33.4 and
68.5%, respectively; P<0.05). These results indicated that
changes in YAP gene expression affected the expression of LATS1
through a positive regulatory association between YAP and LATS1.
Following YAP mRNA downregulation, YAP protein levels decreased and
in turn, the proliferation of cancer cells decreased; therefore, by
reducing the inhibition of YAP, cancer cells decreased the
expression of LATS1, demonstrating a novel negative feedback loop
in the shYAP cells.
Discussion
YAP is a transcriptional co-activator which has been
shown to be a regulator of cell growth in the Hippo signaling
pathway (1). It has been
previously confirmed that YAP was phosphorylated by LATS1, the
upstream factor in the Hippo pathway (8,16),
which was reported to be able to bind to and phosphorylate
transcription regulator YAP in cell culture and in vivo
(8). Phosphorylation of YAP was
found to be associated with its export from the nucleus to the
cytoplasm, where it downregulates growth-associated genes (4,8).
YAP mRNA and protein levels were previously reported
to be higher in major HCC than in para-cancerous tissue (7). One study showed that following YAP1
transformation, a non-tumorigenic hepatocyte cell line formed
visible tumors in a nude mouse model; however, this cell line was
unable to form subcutaneous tumors prior to transformation
(17). These results indicated
that YAP may be involved in the pathogenesis of HCC. In addition,
numerous studies have provided evidence linking YAP gene expression
to tumorigenicity in several solid cancers (4,18,19).
Nuclear overexpression of YAP was reported to contribute to the
growth of pulmonary adenocarcinoma (20) and the widespread upregulation of
YAP in a variety of tumor types further suggested that the YAP gene
may represent a gene which allows cancer cells to evade the effects
of growth inhibition. Furthermore, excessive proliferation may
result in the evasion of intrinsic size-control mechanisms, which
may lead to gradual increases in tumor volume. These findings
indicated that YAP may contribute to multiple aspects of tumor
progression and neoplasia; therefore, YAP may be a potential
therapeutic target molecule for HCC intervention. By contrast,
LATS1 was found to negatively regulate the transcriptional and
transformational functions of YAP through inhibiting its nuclear
translocation (8); therefore,
LATS1 may also be a potential therapeutic target molecule for the
treatment of HCC.
In order to study the association between YAP and
LATS1 in mammals, the present study aimed to investigate YAP and
LATS1 protein levels in 40 cases of HCC tissue, and 10 cases of PCT
using IHC. The results of the present study were consistent with
those of previous studies (6,9),
which showed that YAP protein was overexpressed in HCC samples
compared that of PCT; in addition, the present study revealed that
the majority of normal liver tissues exhibited extensive activation
of LATS1 protein, which was not observed in most HCCs. Exactly the
opposite was found for the expression of YAP. Another previous
study showed that western blot analysis was able to detect LATS1 in
all normal liver tissues, whereas it was only detected in three out
of seven HCC samples (9), which
indicated that LATS1 loss may be involved in the development of
liver cancer; therefore, a YAP-targeted therapy for liver cancer
must focus on LATS1.
In the present study, YAP-positive HCC cells were
found to have low LATS1 expression. It has already been confirmed
that YAP was phosphorylated by LATS1, the upstream factor in the
Hippo signaling pathway (8,16).
This therefore indicated that LATS1 inactivation decreased YAP
phosphorylation, leading to the overexpression of YAP in HCCs.
A previous study using MTT assays showed that
following YAP1 transformation, non-tumorigenic hepatocyte cell
clones demonstrated faster growth rates compared with those of
cells transfected with empty vector controls (17). This therefore indicated that YAP
may be a determinant of liver cancer cell proliferation. In the
present study, cell growth curves revealed that 97H cells with
higher levels of YAP gene expression exerted a significantly
increased proliferative activity compared with that of 97L cells
with lower levels of YAP, indicating that high expression of YAP in
liver cancer cells may induce a high proliferative activity.
Therefore, in the present study, 97H cells were selected for RNA
interference experiments.
The results of the present study demonstrated that
shRNA-mediated downregulation of YAP expression resulted in a
decrease in the number of cells in S-phase as well as significant
cell growth inhibition in vitro. This further indicated that
HCC cell growth was associated with YAP expression levels. Further
analysis revealed that downregulation of YAP in 97H cells was
accompanied by a decrease in the expression of LATS1. This
therefore led to the hypothesis that a negative feedback control
mechanism existed in the Hippo pathway of 97H cells, which may
compensate for RNA interference against YAP in 97H hepatocellular
carcinoma cells.
LATS1 has a critical role in the Hippo pathway;
however, the mechanisms by which LATS1 is regulated at the protein
level remains to be fully elucidated (21). Certain positive regulators of
LATS1, including macrophage-stimulating protein 1/2 (8), hMOB1 (22) and kidney and brain expressed
protein (23) have been identified
in previous studies, as well as negative regulators of LATS1,
including E3 ubiquitin ligase Itch (24) and WW domain-containing E3 ubiquitin
protein ligase 1 (25). These
proteins have been suggested to be involved in the negative
feedback control mechanism between YAP and LATS1; however, further
studies are required in order to clarify this hypothesis.
In the present study, a gradient of YAP expression
in PCT and HCC suggested that the occurrence, development and
progression of HCC was associated with increased YAP expression. A
previous study demonstrated that overexpression of YAP was
accompanied by high levels of YAP mRNA in HCC tissues (7). If the high levels of YAP were due to
overexpression of the YAP gene, and not LATS1 inactivation, the
normal function of LATS1 may be to feedback and suppress the
production of YAP protein. However, in the present study, LATS1
protein levels were not found to be increased; by contrast, LATS1
expression was significantly decreased in HCC cells and
YAP-positive HCCs expressed low levels of LATS1.
In conclusion, the results of the present study
demonstrated that there was a negative correlation between YAP and
LAST1 expression in HCC tissues. In addition, RNAi-mediated YAP
gene silencing inhibited 97H cell proliferation and cell cycle
progression. Furthermore, LATS1 protein levels were markedly
decreased in 97H cells in which YAP was downregulated. These
results provided additional evidence that LATS1 levels may
compensate for the effects of the RNA interference against YAP in
MHCC97H cells. Therefore, these findings may form the basis for YAP
inhibition and LATS1 stimulation as targeted therapies for the
future treatment of HCC.
Acknowledgments
The authors would like to thank Medjaden Bioscience
Limited (Hong Kong, China) for assisting in the preparation of this
manuscript.
References
|
1
|
Simonetti RG, Camma C, Fiorello F, Politi
F, D’Amico G and Pagliaro L: Hepatocellular carcinoma. a worldwide
problem and the major risk factors. Dig Dis Sci. 36:962–972. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carr BI: Hepatocellular carcinoma: current
management and future trends. Gastroenterology. 127(5 Suppl 1):
S218–S224. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao B, Ye X, Yu J, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao B, Wei X, Li W, et al: Inactivation
of YAP oncoprotein by the hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zender L, Spector MS, Xue W, et al:
Identification and validation of oncogenes in liver cancer using an
integrative oncogenomic approach. Cell. 125:1253–1267. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu MZ, Yao TJ, Lee NP, et al:
Yes-associated protein is an independent prognostic marker in
hepatocellular carcinoma. Cancer. 115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang C, Zhang L, He Q, et al: Differences
in yes-associated protein and mrna levels in regenerating liver and
hepatocellular carcinoma. Mol Med Rep. 5:410–414. 2012.
|
|
8
|
Hao Y, Chun A, Cheung K, Rashidi B and
Yang X: Tumor suppressor LATS1 is a negative regulator of oncogene
yap. J Biol Chem. 283:5496–5509. 2008. View Article : Google Scholar
|
|
9
|
Li H, Wolfe A, Septer S, et al:
Deregulation of hippo kinase signalling in human hepatic
malignancies. Liver Int. 32:38–47. 2012. View Article : Google Scholar
|
|
10
|
St John MA, Tao W, Fei X, et al: Mice
deficient of lats1 develop soft-tissue sarcomas, ovarian tumours
and pituitary dysfunction. Nat Genet. 21:182–186. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Y, Tang ZY, Ye SL, et al: Establishment
of cell clones with different metastatic potential from the
metastatic hepatocellular carcinoma cell line MHCC97. World J
Gastroenterol. 7:630–636. 2001.
|
|
12
|
Tian J, Tang ZY, Ye SL, et al: New human
hepatocellular carcinoma (HCC) cell line with highly metastatic
potential (MHCC97) and its expressions of the factors associated
with metastasis. Br J Cancer. 81:814–821. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ramachandran V, Arumugam T, Wang H and
Logsdon CD: Anterior gradient 2 is expressed and secreted during
the development of pancreatic cancer and promotes cancer cell
survival. Cancer Res. 68:7811–7818. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
15
|
Liu XS, Jiang J, Jiao XY, Wu YE, Lin JH
and Cai YM: Lycorine induces apoptosis and down-regulation of Mcl-1
in human leukemia cells. Cancer Lett. 274:16–24. 2009. View Article : Google Scholar
|
|
16
|
Zhang J, Smolen GA and Haber DA: Negative
regulation of YAP by LATS1 underscores evolutionary conservation of
the drosophila hippo pathway. Cancer Res. 68:2789–2794. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu MZ, Chan SW, Liu AM, et al: AXL
receptor kinase is a mediator of YAP-dependent oncogenic functions
in hepatocellular carcinoma. Oncogene. 30:1229–1240. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Camargo FD, Gokhale S, Johnnidis JB, et
al: YAP1 increases organ size and expands undifferentiated
progenitor cells. Curr Biol. 17:2054–2060. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang X, George J, Deb S, et al: The Hippo
pathway transcriptional co-activator, YAP, is an ovarian cancer
oncogene. Oncogene. 30:2810–2822. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim JM, Kang DW, Long LZ, et al:
Differential expression of yes-associated protein is correlated
with expression of cell cycle markers and pathologic TNM staging in
non-small-cell lung carcinoma. Hum Pathol. 42:315–323. 2011.
View Article : Google Scholar
|
|
21
|
Visser S and Yang X: LATS tumor
suppressor: a new governor of cellular homeostasis. Cell cycle.
9:3892–3903. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chow A, Hao Y and Yang X: Molecular
characterization of human homologs of yeast MOB1. Int J Cancer.
126:2079–2089. 2010.
|
|
23
|
Xiao L, Chen Y, Ji M and Dong J: KIBRA
regulates hippo signaling activity via interactions with large
tumor suppressor kinases. J Biol Chem. 286:7788–7796. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ho KC, Zhou Z, She YM, Chun A, Cyr TD and
Yang X: Itch E3 ubiquitin ligase regulates large tumor suppressor 1
stability [corrected]. Proc Natl Acad Sci USA. 108:4870–4875. 2011.
View Article : Google Scholar
|
|
25
|
Yeung B, Ho KC and Yang X: WWP1 E3 ligase
targets LATS1 for ubiquitin-mediated degradation in breast cancer
cells. PLoS One. 8:e610272013. View Article : Google Scholar : PubMed/NCBI
|