Introduction
Type 1 diabetes mellitus (T1D) is an autoimmune
disease that occurs due to autoreactive T cell-induced damage to
pancreatic insulin-producing β cells. In addition, it was reported
that by the time the first clinical symptoms of T1D become evident,
only 15–20% of β cells remain (1,2).
Inflammatory cells in the islet milieu produce cytokines and
cytokine-induced proteins, which have a critical role in the
pathogenesis of insulitis and T1D. The onset of T1D is typically
associated with altered cytokine expression, from primarily T
helper cell (Th)2 to Th1 cytokines, which was thought to occur due
to continuous inflammation in the pancreas (3). Therefore, regulating the immune
system with regard to Th2-like immunity represents a potential
therapeutic strategy for T1D (4).
In animal models of T1D, approaches favoring Th2 activity have led
to diabetes prevention; in addition, there is evidence to suggest
that the administration of these immunoregulatory cytokines reduces
autoimmune diabetes (4–6). The Th2-like cytokine interleukin
(IL)-10 has been extensively investigated and was suggested to be a
potential candidate for effective immune diversion for diabetes
treatment (7).
IL-10 is known to be an anti-inflammatory cytokine,
which exhibits potent suppressive effects in preventing autoimmune
disease (8). IL-10 was reported to
be effective in the suppression of cytokine production in T cells
(IL-2), natural killer cells [interferon (IFN)-γ] and
monocyte/macrophages [IL-1β, IL-6, IL-8, IL-12, tumor necrosis
factor (TNF)-α and granulocyte-macrophage colony-stimulating factor
(GM-CSF)]; IL-10 was also implicated in the induction of anergy in
T cells (9,10). It was demonstrated that IL-10
overexpression prevented graft rejection in transplant organs
(7). IL-10 treatment was shown to
inhibit the onset of T1D in non-obese diabetic (NOD) mice and
prevent disease recurrence, which was dependent on the time and
mode of administration (early vs. late, systemic vs. local). Of
note, systemic delivery of the IL-10 gene and IL-10 protein or
virus-mediated IL-10 delivery was suggested to delay the onset of
diabetes symptoms as well as reduce the occurrence of diabetes in
NOD mice (11,12).
IL-10 has been reported to serve as a growth factor
for T regulatory cells (Tregs) (13,14);
it was demonstrated that IL-10 increased the
CD4+CD25+ cell population in vitro and
in vivo (15,16). In addition, a previous study
revealed that IL-10 gene transfer prevented IL-1β-induced nitric
oxide production in β cells in vitro, which were also found
to retain their insulin secretion function, even at high glucose
levels (17). Furthermore, IL-10
overexpression inhibited IL-1β-induced Fas expression and insulin
secreting cell apoptosis (18).
These findings supported the functional importance of IL-10 in
immune regulation. In addition, evidence suggested that IL-10
exhibited a partial, direct protective effect on human islets
exposed to pro-inflammatory and Th1 cytokines (19). However, to the best of our
knowledge, few direct measurements have been published regarding
the anti-apoptotic mechanism of IL-10 in vivo. Recombinant
IL-10 protein treatment requires repeated or continuous
administrations due to its short half-life. Therefore, viral
vectors are a promising type of gene transfer vehicle, as they
often mediate highly efficient gene transfer and stably express
genes.
The aim of the present study was to assess the
recombinant adenovirus-based transfer of genes encoding IL-10 to
manipulate β cell autoimmunity in NOD mice. The present study aimed
to examine if overexpression of IL-10 in NOD mice was able to
prevent the development of diabetes by downregulating experimental
autoimmune disorders.
Materials and methods
Animals
A total of 42 (n=14/group) female NOD mice, 8 weeks
of age, were purchased from the Shanghai Laboratory Animal Center,
Chinese Academy of Sciences (Shanghai, China) and housed in
specific pathogen-free conditions. The animals were housed four per
cage in a room, which was maintained under standard laboratory
conditions (12 h light/dark cycle), at a constant temperature
(25±1°C) and a humidity of 50–60%. Adenovirus treatments of NOD
mice were initiated at age 9 weeks, which was 2 weeks prior to
diabetes onset (from age 11–24 weeks). Diabetes was defined as two
consecutive blood glucose values >250 mg/dl in NOD mice. A total
of seven NOD mice were sacrificed by cervical dislocation at 12
weeks of age and blood samples (1 ml) were obtained from the
abdominal vein and stored at −80°C, until further use. The
remainder of the mice were allowed to develop diabetes and were
sacrificed 1 week post diabetes onset or at 30 weeks. All
procedures were approved by the Ethics Committee for the Use of
Experimental Animals at Guangzhou Women and Children's Medical
Center (Guangzhou, China). All efforts were made to minimize the
animals' suffering.
Preparation of the recombinant
adenovirus
An E1- and E3-deleted, first-generation recombinant
adenovirus (replication-deficient) was used as a vector in these
experiments. The recombinant Adv-IL-10 vector was generated at the
Qingdao University (Qingdao, China) by triple transfection protocol
as previously described (18).
Briefly, the IL-10 gene was cloned into the shuttle vector,
pAdTrack-CMV, and the produced plasmid was linearized with
PmeI digestion (New England Biolabs, Beverly, MA, USA). The
linearized plasmid was co-transfected into Escherichia coli
BJ5183 cells with an adenoviral backbone plasmid, pAdEasy-1. The
recombinant adenovirus encoding murine IL-10 (Adv-IL-10) was
confirmed by restriction endonuclease analysis. The linearized
recombinant plasmid was transfected into HEK293 cells (kindly
provided by Professor Bing Luo, Medical College of Qingdao
University, China) and high-titer viral stocks were prepared. The
adenovirus vector expressing enhanced green fluorescent protein was
also produced for use as the viral control. Viruses were then
purified from HEK293 cells by freeze/thaw cycles and cesium
chloride (Solarbio Science & Technology. Co., Ltd., Beijing,
China) step gradients. All the viral titers were
5.5×1010 plaque forming unit/ml. Aliquots of the
obtained recombinant adenoviruses were frozen and stored in the
ultra-low temperature freezers (Thermo Forma 900; Thermo Fisher
Scientific, Inc., MA, USA).
Treatment of NOD mice with
Adenoviruses
Female NOD mice were intraperitoneally (IP) injected
with 100 µl Adv-IL-10 or Adv-GFP (1×1010
pfuv/ml). Age-matched mice were injected with 100 µl normal
saline (Hualu Pharmaceutical Co., Ltd,. Shandong, China) as
controls. The general condition (i.e. alertness and physical
activity) and body weight of mice were observed and measured
weekly.
Incidence rate of T1D
During the experimental period, blood samples (0.5
ml) were taken weekly from the tail vein of NOD mice and glucose
levels were measured using a one-touch ultra glucometer (Johnson
& Johnson Medical Ltd, Suzhou, China). When glucose levels
exceeded 250 mg/dl on two consecutive occasions (initial
measurement and then measured again 24 h later), animals were
considered to have T1D and were sacrificed 1 week post diagnosis
for pathological examinations.
Reverse transcription polymerase chain
reaction (RT-PCR) for detecting transgene expression
Prior to sacrifice, splenocytes and liver were
isolated from all NOD mice. The spleens were harvested and pressed
through a nylon mesh to produce a single-cell suspension. The red
blood cells were treated with ACK lysis buffer (Beyotime Institute
of Biotechnology, Shanghai, China) for 5 min. The isolated
splenocytes were washed three times and resuspended in RPMI-1640
(Sigma-Aldrich, St. Louis, MO, USA), supplemented with 10% fetal
calf serum (FCS; Gibco Life Technologies, Carlsbad, CA, USA),
penicillin and streptomycin (Gibco Life Technologies) for the
subsequent experiments. Total RNA (1 µg from liver and
spleen) was extracted and purified using TRIzol® (GE
Healthcare, Shanghai, China) according to the manufacturer's
instructions. Equal amounts (1 µg) of RNA were then
reverse-transcribed into complementary DNA using a Prime Script RT
Reagent kit (Takara Bio, Inc., Dalian, China). The primers (Sangon
Biotech Co., Ltd., Shanghai, China) used for PCR were as follows:
IL-10 forward, 5′-GGCATGCTTGGCTCAGCACTG-3′ and reverse,
5′-GCCCTGCAGTCCAGTAGACG-3′ (580 bp). Cycling conditions were as
follows: Initial activation at 95°C for 5 min, followed by 40
cycles of 95°C for 30 sec, 55°C for 60 sec and 72°C for 60 sec. The
RT-PCR products were detected using 1.2% agarose gel
electrophoresis (Qiagen, Hilden, Germany). The products were
visualized by staining with ethidium bromide (Solarbio Biotech Co.,
Ltd.), observed and photographed using Bio-Rad gel imaging system
(Bio-Rad Gel Doc 2000 UV Chemi Doc; Bio-Rad Laboratories, Inc.,
Pleasanton, CA, USA).
Histological and morphological
analyses
Pancreatic tissues from Adv-IL-10- and
Adv-GFP-treated NOD mice were removed, fixed in 10% buffered
formalin (Xilong Chemical Co., Ltd., Guangzhou, China), embedded in
paraffin (Junjie Electronics Co., Ltd., Wuhan, China) and sectioned
at 5-µm thickness. Sections were stained with hematoxylin
and eosin (Beyotime Institute of Biotechnology) to assess
pancreatic islets histology and morphology in the experimental
animals. Between 5 and 10 islets of each pancreatic tissue sample
were observed and the inflammation of the islets was graded. The
grading of insulitis was scored as follows: 0, no inflammation; 1,
peri-insulitis; 2, inflammatory infiltration <50% of islet area;
and 3, inflammation >50% of islet area and islet structure
disruption (20).
Analyses of serum cytokine levels
Sera from Adv-IL-10- or Adv-GFP-treated NOD mice
were frozen at −20°C and stored until further use. Quantitative
determination of cytokine (IL-10 and IFN-γ) production was assessed
using a murine IL-10 and a murine IFN-γ ELISA commercial kits
(R&D System, Inc., Minneapolis, MN, USA). The optical density
(OD) value was determined at 450 nm using an ELISA reader (Bio-Rad
Laboratories, Inc.) at 405 nm. The amount of cytokine present was
determined from standard curves using purified recombinant
cytokines.
Analyses of cytokine-secreting
splenocytes
Splenocytes were collected and stimulated with 10
µg/ml Concanavalin A (Sigma-Aldrich, St. Louis, MO, USA).
Following incubation for 48 h, the culture supernatants were
harvested for determination of IL-10 and IFN-γ using ELISA kits
(R&D Systems, Inc.) according to the manufacturer's
instructions. All experiments were performed in triplicate for each
sample.
Flow cytometric analysis of Treg cells in
IL-10-treated NOD mice
NOD mice were randomly selected for the detection of
Tregs in the spleen. These mice were sacrificed following IL-10
treatment. Cells from the spleen were prepared and incubated with 1
µg anti-mouse monocolnal CD4, anti-mouse monoclonal CD25 and
anti-mouse monoclonal Foxp3 conjugated with fluorescein
isothiocyanate, phycoerythrin (PE) or CyChrome, respectively
(eBioscience, San Diego, CA, USA) on ice for 30 min. Cells were
then washed in phosphate-buffered saline (PBS; Solarbio Biotech
Co., Ltd.)-2% fetal calf serum (Gibco Life Technologies). Stained
cells were analyzed using a FACSCalibur flow cytometer
(Becton-Dickinson, San Jose, CA, USA).
Terminal deoxynucleotidyl transferase
deoxyuridine triphosphate (dUTP) nick-end labeling (TUNEL)
assay
TUNEL staining was performed in order to detect
apoptosis using an in situ cell death detection kit (Roche,
Indianapolis, IN, USA). Paraffin sections (4 µm) were
deparaffinized and dehydrated in serial alcohol concentrations
(100, 95, 90, 80 and 70%, diluted in double distilled water).
Protein was digested using proteinase K (Sigma-Aldrich) at 37°C for
10 min and then incubated with terminal transferase and
digoxigenin-dUTP. Endogenous peroxidase was quenched with 3%
H2O2 (Xilong Chemical Co., Ltd.) in methanol
(Xilong Chemical Co., Ltd.) for 5 min. Labeled end was detected
using streptavidin-horseradish peroxidase conjugate. Specimens were
further processed with 3,3-diaminobenzidine coloration
(Sigma-Aldrich) and slightly redyed with Mayer hematoxylin
(Sigma-Aldrich). Specimens were subsequently washed with PBS twice
for 5 min each. The sections were susbequently incubated with 50
µl TUNEL reaction mixture at 37°C for 1 h in a humidified
chamber. Following incubation, the slides were rinsed with PBS
three times for 5 min each and the apoptotic cells were visualized
using an Olympus BX40F microscope (Olympus, Melville, NY, USA).
Apoptotic cells were identified by their dark brown nuclei under a
light microscope (Olympus IX81; Olympus, Tokyo, Japan). The number
of apoptotic cells was counted in five randomly selected fields of
vision (magnification, ×400) in a blinded manner.
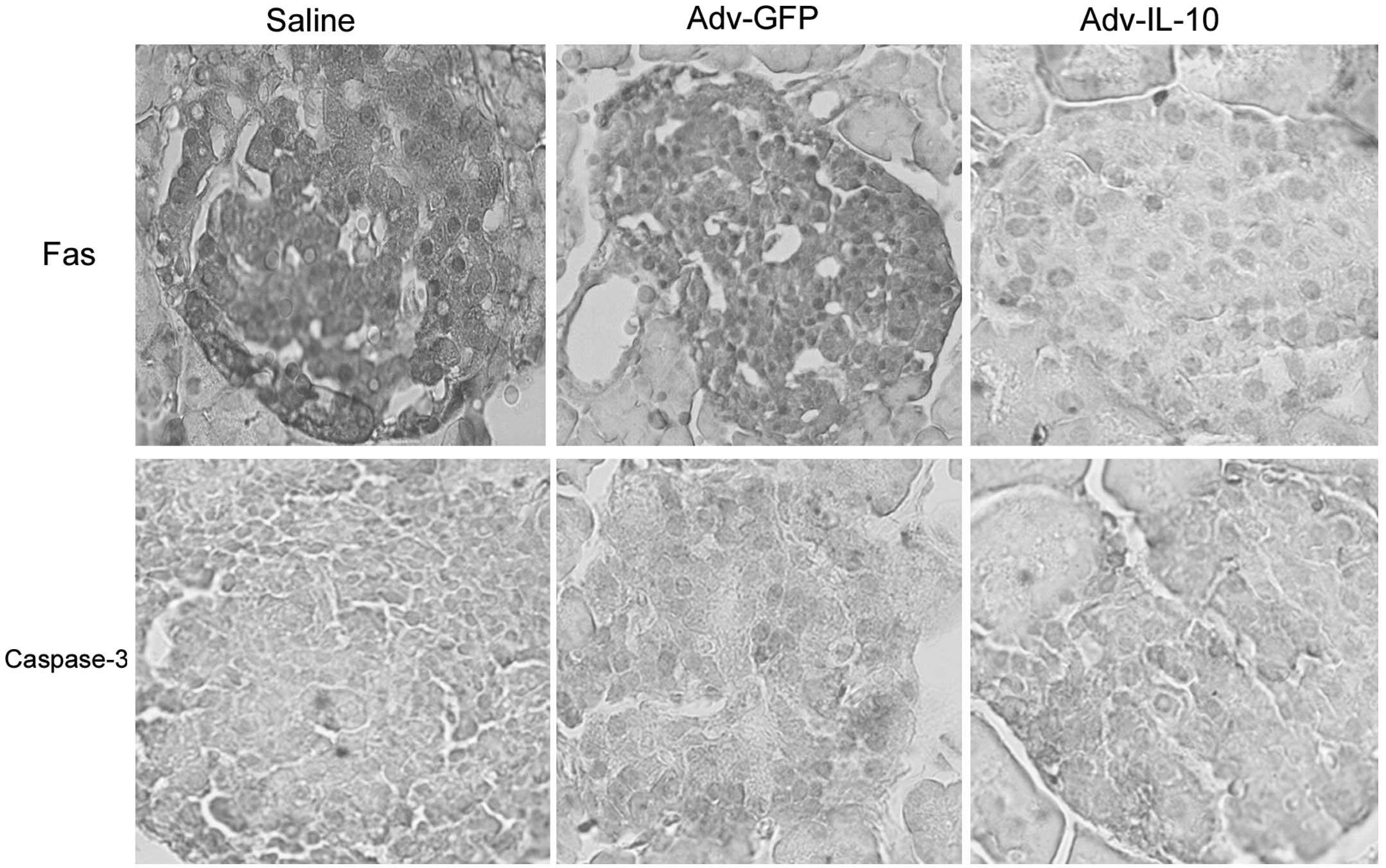
Immunohistochemical analysis
Fas staining was performed in order to determine the
presence of apoptosis pathway protein and caspase-3 staining was
used to detect early apoptosis. Briefly, the pancreata were fixed
overnight in 10% formalin and embedded in paraffin. Tissues were
then sectioned (4 µm-thick) and deparaffinized. The
endogenous peroxidase activity was quenched by 3% hydrogen
peroxidemethanol (Boster Biological Technology, Ltd., Wuhan, China)
for 10 min at room temperature. Samples were then incubated
overnight with primary antibodies against rabbit anti-caspase-3
polyclonal antibody and rabbit anti-Fas rabbit polyclonal antibody
(1:100; BD Pharmingen, San Diego, CA, USA) at 4°C. Following
washing with PBS, samples were incubated with biotinylated
secondary antibody (Boster Biological Technology, Ltd.) for 20 min
at 37°C. Slides were visualized with diaminobenzidine (Boster
Biological Technology, Ltd, Wuhan, China), counterstained with
hematoxylin, dehydrated through increasing concentrations of
alcohol, cleared in xylene and then observed with microscope
(Olympus). Images were captured using a Sony 3CCD color video
camera (Sony, Tokyo, Japan).
Statistical analysis
Values are presented as the mean ± standard error of
the mean. Parametric tests or non-parametric tests were performed
to determine statistically significant differences between groups,
as appropriate, using SPSS 17.0 software (International Business
Machines, Armonk, NY, USA). Student's t-test was used for
statistical analysis of the data. P<0.05 was considered to
indicate a statistically significant difference between values.
Results
IL-10 gene transfer protects pancreatic β
cells from autoimmune attack in T1D
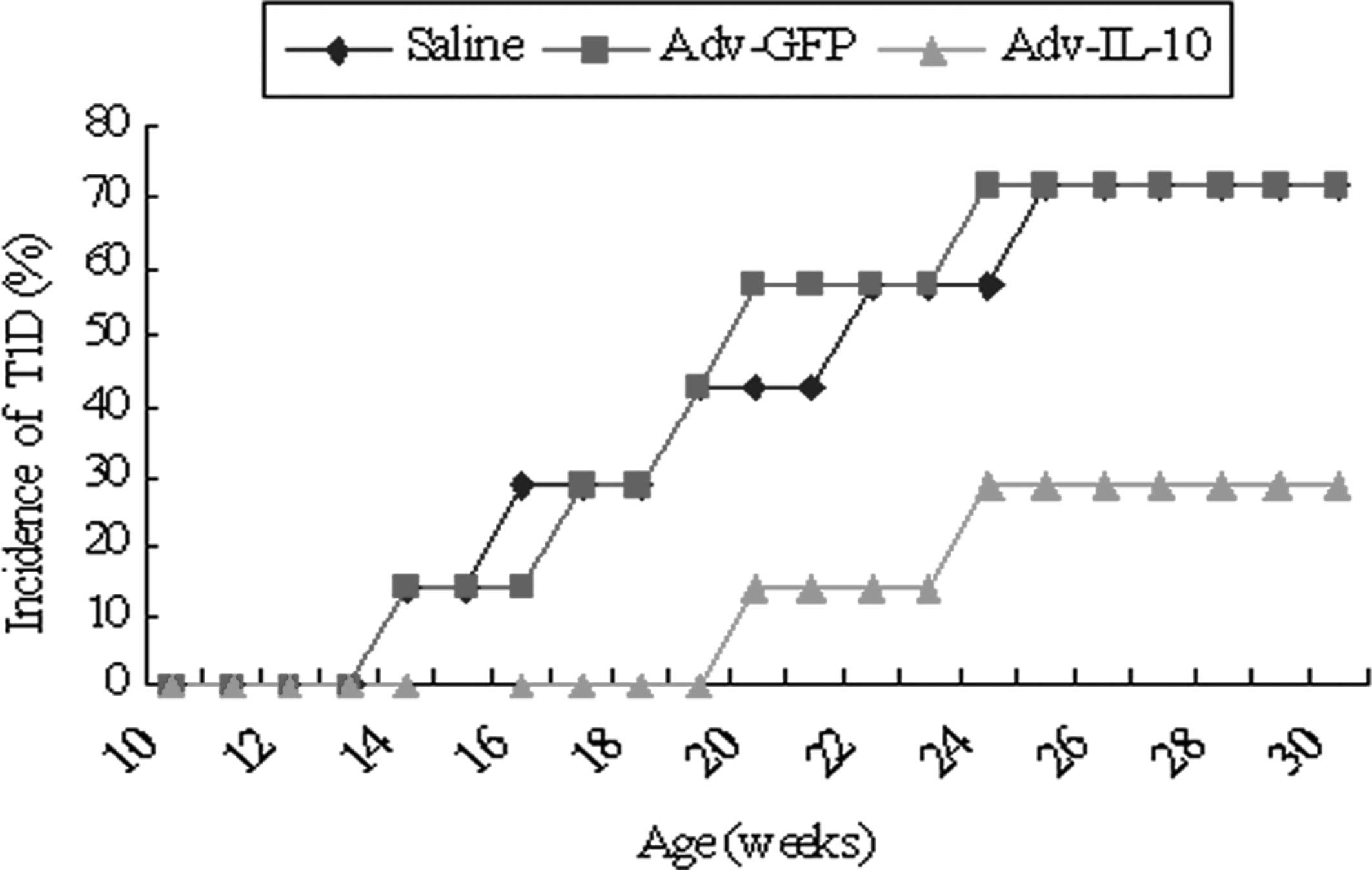
To determine whether IL-10 gene overexpression
delayed or prevented diabetes, 9-week-old NOD mice were IP injected
with Adv-IL-10. As shown in Fig.
1, the incidence of diabetes in Adv-GFP-injected NOD mice was
57.1% at 20 weeks, which increased to 71.4% by 30 weeks of age. By
contrast, only 28.6% of Adv-IL-10-injected NOD mice developed
diabetes throughout the entire experimental protocol (Fig. 1).
Gene transfer of IL-10 reduces insulitis
in prediabetic NOD mice
It was investigated whether the insulitis of NOD
mice was suppressed following IP injections of Adv-IL-10 plasmid
complexes. As shown in Fig. 2A,
severe mononuclear cell infiltration was observed in and around
islets in control mice groups. By contrast, IL-10-injected mice
displayed reduced mononuclear cell infiltration and insulitis was
only observed at the periphery of islets (peri-insulitis). Average
insulitis scores for mice receiving the IL-10 virus were decreased
compared with those of the control mice groups (Fig. 2B). The transfer of the IL-10 gene
into islets can lessen the degree of lymphocytic infiltration.
These results indicated that systemic overexpression of IL-10
protected pancreatic islets against immunological destruction.
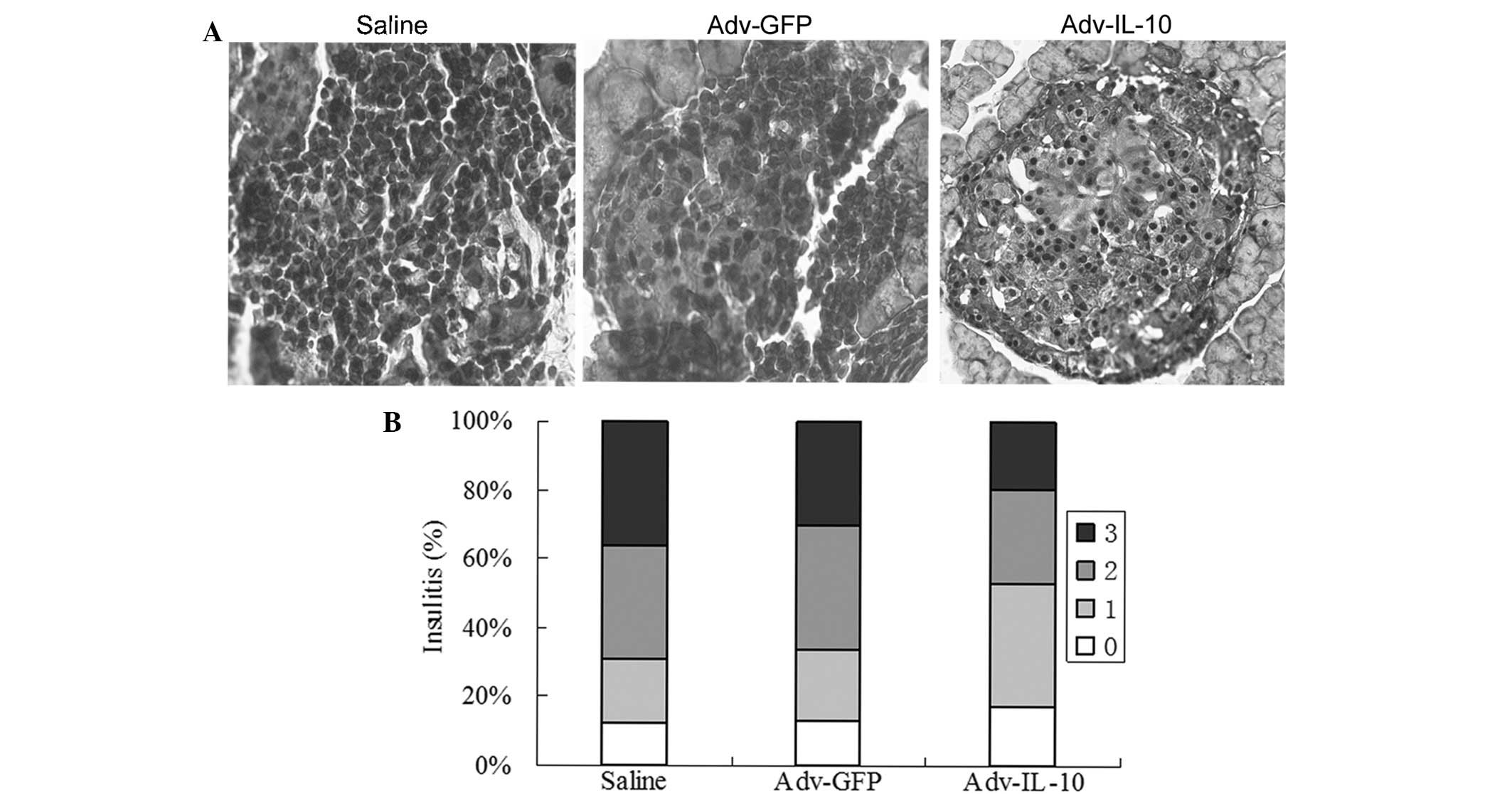 | Figure 2IL-10 gene transfer reduces
insulitis. (A) Pancreatic sections were stained with hematoxylin
and eosin, then assessed for insulitis. For each mouse, five random
sections were scored (n=7 per group; magnification, ×400). A
representative image from each group is shown. (B) Histograms
depict the percentage of normal islets (stage 0, unfilled bar),
peri-insulitis (stage 1, light gray bar), insulitis involving
<50% of the islet in cross-section (stage 2, dark gray bar) or
insulitis involving >50% of the islet (stage 3, black bar).
IL-10, interleukin-10; Adv-GFP, adenovirus carrying green
fluorescent protein; Adv-IL-10, adenovirus carrying IL-10. |
RT-PCR detection of IL-10 messenger
(m)RNA in NOD mice
To evaluate whether transgene expression of IL-10
was involved in the development of autoimmune diabetes, a RT-PCR
assay of total RNA prepared from the liver and splenocytes of NOD
mice was developed to detect transcripts of IL-10 genes. The
results showed that mRNA levels of IL-10 in the liver and
splenocytes were increased in the Adv-IL-10-treated mice compared
with those of mice in the control groups (Fig. 3). This demonstrated that Adv-IL-10
expressed IL-10 genes with high efficiency.
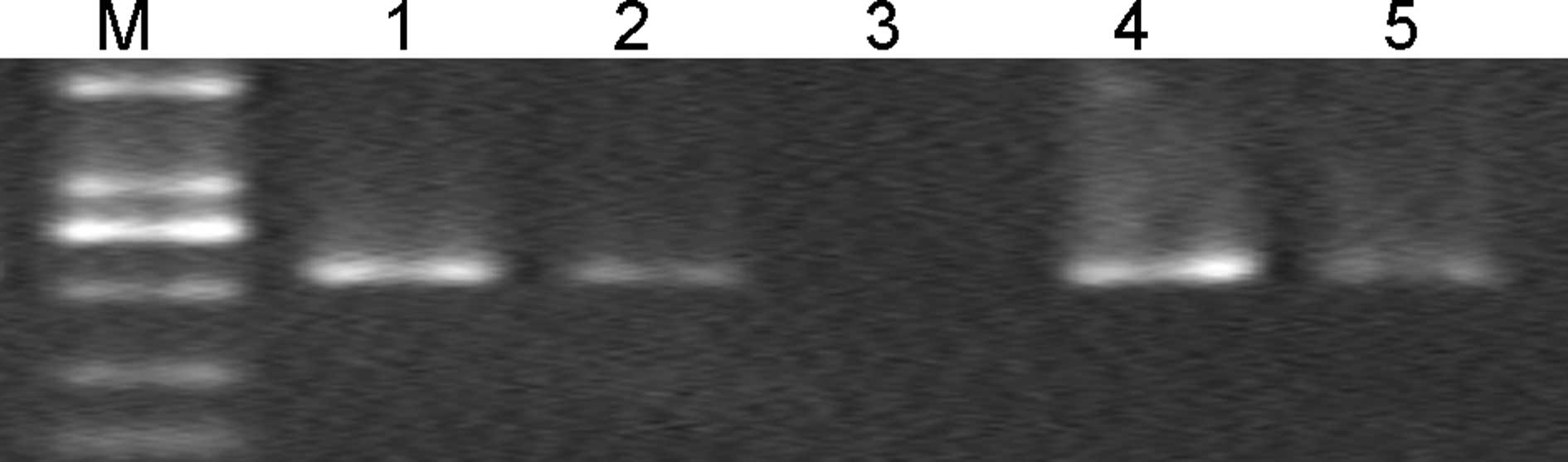 | Figure 3IL-10 messenger RNA expression in
liver cells and splenocytes of non-obese diabetic mice, as
determined by RT-PCR. Total RNA (5 µg) of liver and
splenocyte samples from non-obsese diabetic mice was used for
RT-PCR. Lanes are representative results from each group as
follows: M, DL2000 marker; 1 and 2, liver from mice injected with
Adv-IL-10 or Adv-GFP, respectively; 3, saline-injected mice; 4 and
5, isolated splenocytes from mice injected with Adv-IL-10 or
Adv-GFP, respectively. Experiments were performed in triplicate
(n=3 per group). IL-10, interleukin-10; RT-PCR, reverse
transcription polymerase chain reaction; Adv-GFP, adenovirus
carrying green fluorescent protein; Adv-IL-10, adenovirus carrying
IL-10. |
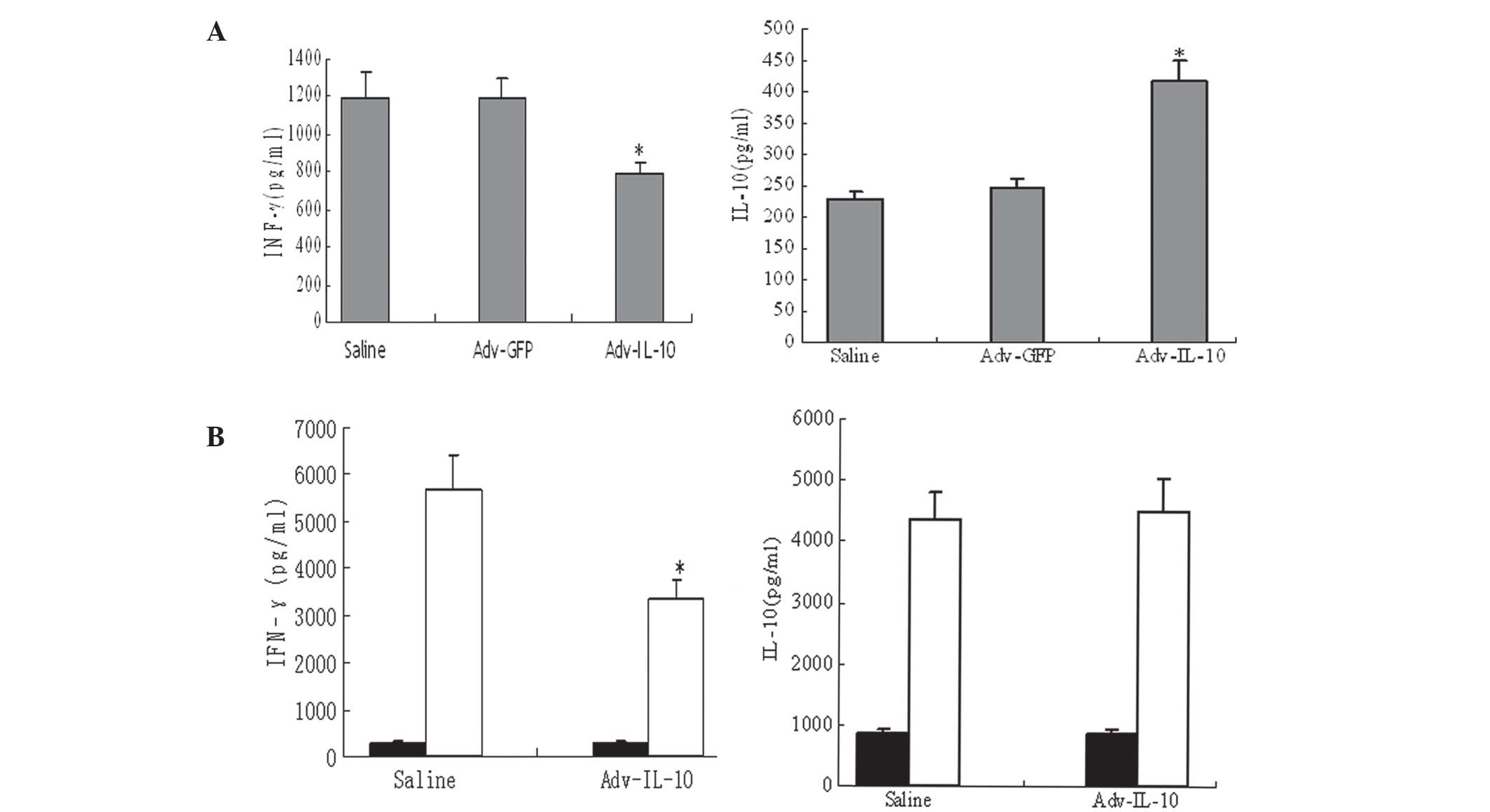
Effect of Adv-IL-10 delivered
immunomodulatory cytokines on T1D
In order to determine whether the balance of Th1/Th2
responses was affected by IL-10 expression, Th1 and Th2 cytokine
levels were determined in serum samples and in media from
splenocytes isolated from animals 3 weeks following saline or
Adv-IL-10 administration. As shown in Fig. 4A, serum levels of IFN-γ were
significantly decreased in mice injected with Adv-IL-10 compared
with the saline and Adv-GFP control groups (P<0.05); however,
serum levels of IL-10 were increased in the Adv-IL-10-treated group
(P<0.05). In addition, the production of IFN-γ by activated
splenocytes was substantially lower in Adv-IL-10-treated mice
(P<0.05); however, IL-10 levels were not significantly different
between the saline- and Adv-IL-10-treated groups (Fig. 4B).
Treg cell activity in vivo
Previous studies have highlighted the role of IL-10
in the function of Treg cells (15,16).
The present study investigated the effect of IL-10 gene therapy on
the function of Tregs. NOD mice were treated with Adv-IL-10 or
saline and flow cytometry was then used to investigate the presence
of Tregs in pancreatic islets. Flow cytometric analysis showed that
the number of CD4+CD25+Foxp3+ Treg
cells in the Adv-IL-10 group (7.03±0.30%) was significantly
increased compared with the saline (6.63±0.44%) or Adv-GFP
(6.58±0.53%) controls (P<0.01; n=7).
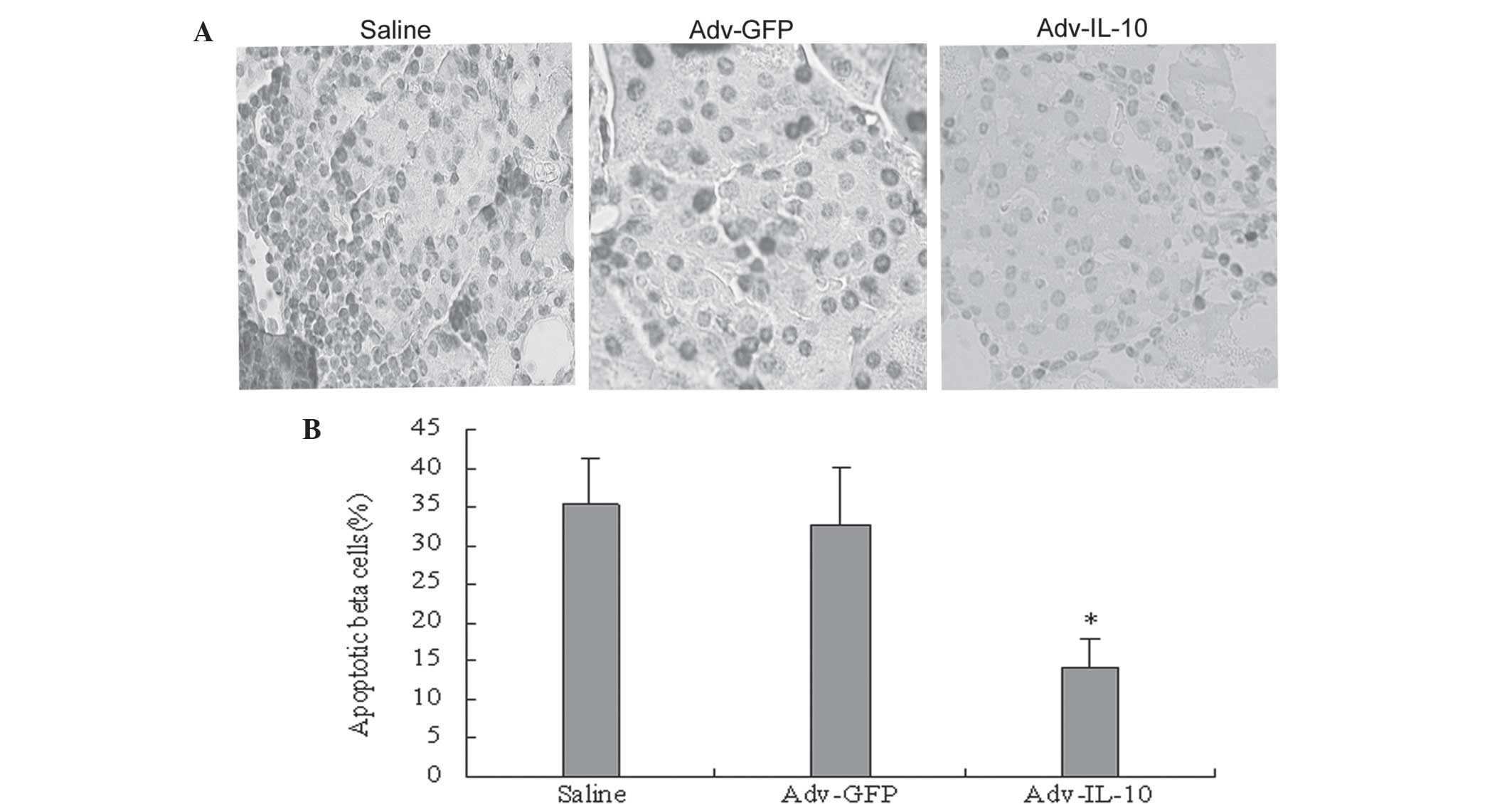
IL-10 therapies prevent β cell
apoptosis
As shown in Fig. 5,
the apoptotic rate of β cells was decreased in mice treated with
IL-10. The apoptosis rate of β cells was lower in the Adv-IL-I0
group (14.10±2.70%) compared with the Adv-GFP group (32.80±7.5%)
and the normal control group (35.40±6.10%; P<0.05) by TUNEL
assay (Fig. 5A and B). Consistent
with the results of the TUNEL staining assay, staining for
caspase-3 showed a significant reduction in caspase-3 expression in
IL-10-treated mice (28.6+4.64%; n=7) compared with the control
(48.8+10%) and Ad-GFP (42.5±7.8%) groups These data indicated that
IL-10 blocked general apoptotic pathways via inhibition of
caspase-3. Therefore, the present study examined whether the
decreased apoptosis observed in the islets of mice treated with
Adv-IL-10 were associated with the downregulation of the
anti-apoptotic protein Fas (Fig.
6). Immunohistochemical staining revealed that Fas and
caspase-3 protein expression were markedly reduced in the
pancreatic islets of mice treated with the Adv-IL-10 compared with
mice in the Adv-GFP- and saline-treated groups.
Discussion
T1D occurs due to cell-mediated
autoimmune-associated damage of pancreatic insulin-producing β
cells (21). Pro-inflammatory
cytokine-mediated pancreatic β cell disruption is a critical
pathological event in the pathogenesis of T1D (22). It was reported that Th1 cells were
involved in establishing autoimmunity in NOD mice; in addition,
endogenous Th2 cells were suggested to have a protective effect on
the development of diabetes in mice (23). IL-10 exhibited pleiotropic effects
in immunoregulation and inflammation (24); in addition, IL-10 was reported to
have various roles in mediating autoimmune responses, which were
dependent on the dosage and site of administration. For example,
systemic administration of IL-10 was found to restrict the
progression of insulitis and prevent the onset of diabetes
(25). In NOD mice, systemic
administration of recombinant adeno-associated virus (rAAV)- or
plasmid-expressed IL-10 resulted in the prevention of spontaneous
diabetes (11,26). A previous study suggested that
IFN-γ/TNF-α synergism may be the final effector molecules involved
in autoimmune diabetes in NOD mice (27). Serreze et al (23) reported that non-specific
immunostimulatory agents prevented the development of diabetes in
NOD mice, which was reported not to be dependent on the expression
of the Th1 cytokine IFN-γ, rather than a Th1 to Th2 cytokine shift
(23).
The present study aimed to examine the regulatory
mechanisms of IL-10-induced protection against diabetes in NOD mice
and it was observed that the injection of Adv-IL-10 into
prediabetic NOD mice was able to significantly attenuate insulitis
progression and the development of diabetes. RT-PCR revealed that
IL-10 was highly expressed in islets of prediabetic NOD mice. In
addition, the present study provided evidence to suggest that
systemic IL-10 gene transfer suppressed the Th1 cell response in
NOD mice islets as wells as significantly reduced the expression of
Th1-type cytokine IFN-γ in sera and in activated splenocytes. These
results suggested that inhibition of IFN-γ expression by IL-10 may,
at least in part, contribute to the anti-inflammatory properties of
IL-10 in islet inflammation.
Numerous previous studies have reported that Tregs
have a critical role in the maintenance of peripheral pancreatic
self-antigen tolerance (28–33).
The decreased ability of Tregs to suppress autoreactive T cells was
indicated to be associated with disease progression in mice and
humans (34). Injection of
polyclonal CD4+CD25+Foxp3+ Treg
cells into NOD mice was reported to prevent diabetes (35). Previous studies have indicated that
systemic IL-10 overexpression in NOD mice enhanced the
CD4+CD25+ Treg population and attenuated
diabetes development (16). In
addition, rAAV-IL-10 treatment was demonstrated to have a positive
dose-dependent effect on the number of regulatory cells in
vivo (16). The results of the
present study demonstrated that the Adv-IL-10-treated group had an
increased number of CD4+CD25+ Treg cells in
the splenocyte population compared with that of the saline- and
Adv-GFP-treated control mice. This therefore indicated that β
cell-specific IL-10 expression reduced the occurrence of
disease-inducing lymphocytes, at least in part, through maintaining
the number of Treg cells in pancreatic islets. It was therefore
hypothesized that IL-10 treatment may also be involved in
regulating the immune response, resulting in the prevention of
diabetes.
Apoptosis is the primary mechanism of β cell death
in NOD mice (36). It was
demonstrated that β cell apoptosis induced a gradual decrease in
the number of β cells in rodent models of T1D (37). Autoreactive T lymphocytes are
essential effector cells in murine autoimmune diabetes (38), which results in the induction of β
islet cells apoptosis (39,40).
Pro-inflammatory cytokines promote cell death via complex signaling
pathways (41,42), which result in the direct
activation of caspases, the primary effector molecules of
programmed cell death. The synergistic effect of IFNγ and TNFα was
reported to induce classical caspase-dependent apoptosis in murine
insulinoma and pancreatic islet cells (27,40);
however, independently these cytokines exhibited no significant
effects. The process of apoptosis involves caspase activity.
Caspase-3 activation induces apoptosis in all cells previously
examined, including β cells, and is an early mediator of Fas
signaling (42,43). Fas (CD95) is a member of the TNF
receptor superfamily. Normal human pancreatic β cells do not
constitutively express Fas; however, numerous studies on isolated
rodent and human islets have demonstrated cytokine-induced Fas
upregulation in the β cells (44,45).
IL-10 has been reported to inhibit apoptotic pathways in different
cell types (46–48), although there have been few studies
regarding the protective effect of IL-10 against pro-inflammatory
cytokine-induced β cell apoptosis. The present study demonstrated
that the apoptosis of β cells was decreased in islets of
Adv-IL-10-treated mice, which indicated that IL-10 production
protected β cells from death. This was consistent with previous
studies that reported the direct protective effect of a combination
of Th2 cytokines on human islet cells (19), as well as the anti-apoptotic effect
of IL-10 in human promyeloid cells (46), cardiomyocytes (47) and endothelial cells (48). In addition, the present study
demonstrated that IL-10 treatment protected β cells against
apoptosis through inhibition of caspase-3 activity and attenuated
cytokine-induced expression of Fas. IL-10 was previously
demonstrated to suppress Fas-induced apoptosis by reducing
caspase-3 activity via Fas-associated death domain-like
interleukin-1-converting enzyme-like inhibitory protein
upregulation and the downregulation of caspase-8 activity (49). It has been suggested that the
inhibition of pro-inflammatory cytokine-mediated caspase-3
activation by IL-10 may be critical for pancreatic β cell
protection and survival. Therefore, the results of the present
study may implicate IL-10 as a potential therapeutic agent for the
treatment of T1D.
In conclusion, the present study demonstrated that
IL-10 gene transfer protected against autoimmune diabetes in the
pancreatic β cells of NOD mice. The mechanism of which involved
enhancing the anti-apoptotic and anti-inflammatory capacity of
islets without altering the status of systemic immunity.
Furthermore, these results suggested that genetic manipulation of
IL-10 levels in islets may be a potential therapeutic strategy for
the treatment of T1D.
Acknowledgments
The present study was supported by grants from the
Natural Science Foundation of Guangdong Province (nos.
S2012040006330 and S2012040006311) and the Science and Technology
Program of Guangdong Province (no. 2012B031800016). The authors
would like to thank Professor Zhihong Chen for technical
assistance.
References
|
1
|
Bekris LM, Kavanagh TJ and Lernmark A:
Targeting type 1 diabetes before and at the clinical onset of
disease. Endocr Metab Immune Disord Drug Targets. 6:103–124. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karlsson FA, Berne C, Björk E, et al:
Beta-cell activity and destruction in type 1 diabetes. Ups J Med
Sci. 105:85–95. 2000.PubMed/NCBI
|
|
3
|
Wilson SB, Kent SC, Patton KT, et al:
Extreme Th1 bias of invariant Valpha24JalphaQ T cells in type 1
diabetes. Nature. 391:177–181. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rabinovitch A: Immunoregulatory and
cytokine imbalances in the pathogenesis of IDDM. Therapeutic
intervention by immunostimulation? Diabetes. 43:613–621. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tisch R and McDevitt H: Insulin-dependent
diabetes mellitus. Cell. 85:291–297. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Delovitch TL and Singh B: The nonobese
diabetic mouse as a model of autoimmune diabetes: immune
dysregulation gets the NOD. Immunity. 7:727–738. 1997. View Article : Google Scholar
|
|
7
|
Zhang YC, Pileggi A, Agarwal A, et al:
Adeno-associated virus-mediated IL-10 gene therapy inhibits
diabetes recurrence in syngeneic islet cell transplantation of NOD
mice. Diabetes. 52:708–716. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saraiva M and O'Garra A: The regulation of
IL-10 production by immune cells. Nat Rev Immunol. 10:170–181.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moore KW, de Waal Malefyt R, Coffman RL,
et al: Interleukin-10 and the interleukin-10 receptor. Annu Rev
Immunol. 19:683–765. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding Y, Chen D, Tarcsafalvi A, et al:
Suppressor of cytokine signaling 1 inhibits IL-10-mediated immune
responses. J Immunol. 170:1383–1391. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nitta Y, Tashiro F, Tokui M, et al:
Systemic delivery of interleukin 10 by intramuscular injection of
expression plasmid DNA prevents autoimmune diabetes in nonobese
diabetic mice. Hum Gene Ther. 9:1701–1707. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang Z, Chen M, Wu R, et al: Suppression
of autoimmune diabetes by viral IL-10 gene transfer. J Immunol.
168:6479–6485. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Groux H, O'Garra A, Bigler M, et al: A
CD4+ T-cell subset inhibits antigen-specific T-cell responses and
prevents colitis. Nature. 389:737–742. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Asseman C, Mauze S, Leach MW, Coffman RL
and Powrie F: An essential role for interleukin 10 in the function
of regulatory T cells that inhibit intestinal inflammation. J Exp
Med. 190:995–1004. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Levings MK, Sangregorio R, Galbiati F, et
al: IFN-alpha and IL-10 induce the differentiation of human type 1
T regulatory cells. J Immunol. 166:5530–5539. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goudy KS, Burkhardt BR, Wasserfall C, et
al: Systemic overexpression of IL-10 induces CD4+CD25+cell
populations in vivo and ameliorates type 1 diabetes in nonobese
diabetic mice in a dose-dependent fashion. J Immunol.
171:2270–2278. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu AJ, Chen ZH, Tian F, Yan LH and Li T:
Effects of adenovirus-mediated interleukin-10 gene transfer on
apoptosis and insulin secretion function of beta cell. Chinese
Medical Journal. 90:1711–1715. 2010.In Chinese.
|
|
18
|
Xu AJ, Zhu W, Tian F, Yan LH and Li T:
Recombinant adenoviral expression of IL-10 protects beta cell from
impairment induced by pro-inflammatory cytokine. Mol Cell Biochem.
344:163–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Marselli L, Dotta F, Piro S, et al: Th2
cytokines have a partial, direct protective effect on the function
and survival of isolated human islets exposed to combined
proinflammatory and Th1 cytokines. J Clin Endocrinol Metab.
86:4974–4978. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Casellas A, Salavert A, Agudo J, et al:
Expression of IGF-I in pancreatic islets prevents lymphocytic
infiltration and protects mice from type 1 diabetes. Diabetes.
55:3246–3255. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van Belle TL, Coppieters KT and von
Herrath MG: Type 1 diabetes: Etiology, immunology, and therapeutic
strategies. Physiol Rev. 91:79–118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thomas HE, Graham KL, Chee J, et al:
Proinflammatory cytokines contribute to development and function of
regulatory T cells in type 1 diabetes. Ann NY Acad Sci. 1283:81–86.
2013. View Article : Google Scholar
|
|
23
|
Serreze DV, Chapman HD, Post CM, et al:
Th1 to Th2 cytokine shifts in nonobese diabetic mice: sometimes an
outcome, rather than the cause, of diabetes resistance elicited by
immunostimulation. J Immunol. 166:1352–1359. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Asadullah K, Sterry W and Volk HD:
Interleukin-10 therapy – review of a new approach. Pharmacol Rev.
55:241–269. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pennline KJ, Roque-Gaffney E and Monahan
M: Recombinant human IL-10 prevents the onset of diabetes in the
nonobese diabetic mouse. Clin Immunol Immunopathol. 71:169–175.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goudy K, Song S, Wasserfall C, et al:
Adeno-associated virus vector-mediated IL-10 gene delivery prevents
type 1 diabetes in NOD mice. Proc Natl Acad Sci USA.
98:13913–13918. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Suk K, Kim S, Kim YH, et al:
IFN-gamma/TNF-alpha synergism as the final effector in autoimmune
diabetes: a key role for STAT1/IFN regulatory factor-1 pathway in
pancreatic beta cell death. J Immunol. 166:4481–4489. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tang Q, Henriksen KJ, Boden EK, et al:
Cutting edge: CD28 controls peripheral homeostasis of CD4+CD25+
regulatory T cells. J Immunol. 171:3348–3352. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen Z, Herman AE, Matos M, Mathis D and
Benoist C: Where CD4+CD25+ Treg cells impinge on autoimmune
diabetes. J Exp Med. 202:1387–1397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feuerer M, Shen Y, Littman DR, Benoist C
and Mathis D: How punctual ablation of regulatory T cells unleashes
an autoimmune lesion within the pancreatic islets. Immunity.
31:654–664. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Herman AE, Freeman GJ, Mathis D and
Benoist C: CD4+CD25+ T regulatory cells dependent on ICOS promote
regulation of effector cells in the prediabetic lesion. J Exp Med.
199:1479–1489. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gregori S, Giarratana N, Smiroldo S and
Adorini L: Dynamics of pathogenic and suppressor T cells in
autoimmune diabetes development. J Immunol. 171:4040–4047. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Salomon B, Lenschow DJ, Rhee L, et al:
B7/CD28 costimulation is essential for the homeostasis of the
CD4+CD25+ immunoregulatory T cells that control autoimmune
diabetes. Immunity. 12:431–440. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Walker LS: Natural Treg in autoimmune
diabetes: all present and correct? Expert Opin Biol Ther.
8:1691–1703. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Szanya V, Ermann J, Taylor C, Holness C
and Fathman CG: The subpopulation of CD4+CD25+ splenocytes that
delays adoptive transfer of diabetes expresses L-selectin and high
levels of CCR7. J Immunol. 169:2461–2465. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kay TW, Thomas HE, Harrison LC and Allison
J: The beta cell in autoimmune diabetes: many mechanisms and
pathways of loss. Trends Endocrinol Metab. 11:11–15. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cnop M, Welsh N, Jonas JC, et al:
Mechanisms of pancreatic beta-cell death in type 1 and type 2
diabetes: many differences, few similarities. Diabetes. 54(Suppl
2): S97–S107. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Katz JD, Benoist C and Mathis D: T helper
cell subsets in insulin-dependent diabetes. Science. 268:185–1188.
1995. View Article : Google Scholar
|
|
39
|
Kurrer MO, Pakala SV, Hanson HL and Katz
JD: Beta cell apoptosis in T cell-mediated autoimmune diabetes.
Proc Natl Acad Sci USA. 94:213–218. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim YH, Kim S, Kim KA, et al: Apoptosis of
pancreatic beta-cells detected in accelerated diabetes of NOD mice:
no role of Fas-Fas ligand interaction in autoimmune diabetes. Eur J
Immunol. 29:455–465. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Eizirik DL and Mandrup-Poulsen T: A choice
of death – the signal-transduction of immune-mediated beta-cell
apoptosis. Diabetologia. 44:2115–2133. 2001. View Article : Google Scholar
|
|
42
|
Slee EA, Adrain C and Martin SJ: Serial
killers: ordering caspase activation events in apoptosis. Cell
Death Differ. 6:1067–1074. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wolf BB, Schuler M, Echeverri F and Green
DR: Caspase-3 is the primary activator of apoptotic DNA
fragmentation via DNA fragmentation factor-45/inhibitor of
caspase-activated DNase inactivation. J Biol Chem. 274:30651–30656.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Loweth AC, Williams GT, James RF,
Scarpello JH and Morgan NG: Human islets of Langerhans express Fas
ligand and undergo apoptosis in response to interleukin-1beta and
Fas ligation. Diabetes. 47:727–732. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Riachy R, Vandewalle B, Moerman E, et al:
1, 25-Dihydroxyvitamin D3 protects human pancreatic islets against
cytokine-induced apoptosis via down-regulation of the Fas receptor.
Apoptosis. 11:151–159. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou JH, Broussard SR, Strle K, et al:
IL-10 inhibits apoptosis of promyeloid cells by activating insulin
receptor substrate-2 and phosphatidylinositol 3′-kinase. J Immunol.
167:4436–4442. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dhingra S, Sharma AK, Arora RC, Slezak J
and Singal PK: IL-10 attenuates TNF-alpha-induced NF kappaB pathway
activation and cardiomyocyte apoptosis. Cardiovasc Res. 82:59–66.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Londoño D, Carvajal J, Strle K, Kim KS and
Cadavid D: IL-10 Prevents apoptosis of brain endothelium during
bacteremia. J Immunol. 186:7176–7186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bharhani MS, Borojevic R, Basak S, et al:
IL-10 protects mouse intestinal epithelial cells from Fas-induced
apoptosis via modulating Fas expression and altering caspase-8 and
FLIP expression. Am J Physiol Gastrointest Liver Physiol.
291:G820–G829. 2006. View Article : Google Scholar : PubMed/NCBI
|