Introduction
Hepatitis B virus (HBV) infection remains to be a
serious health problem worldwide (1). Carriers of HBV are at increased risk
of developing cirrhosis and hepatocellular carcinoma (HCC), with an
estimated 563,000 mortalities annually worldwide from cirrhosis and
HCC (2). Increasing evidence
suggests that host-immune responses are important in determining
the outcome of HBV infection (3).
Acutely infected individuals typically develop a strong,
multispecific cytotoxic T-lymphocyte (CTL) response and a
polyclonal T-helper (Th) cell response to the virus (4,5),
while these responses are weak or undetected in patients with
chronic HBV infection (6).
Therefore, novel immunotherapeutic approaches are required in order
to alter the T-cell response into a predominant Th1 pathway and
enhance HBV-specific CTL responses, which may facilitate the
eradication of chronic HBV infection.
HBV core antigen (HBcAg) is a peptide that induces
strong immune responses characterized by clear T-cell activity in
natural and recombinant forms (7,8).
Despite the high immunogenicity of exogenous HBcAg, HBV-specific
CTL responses induced by simple exogenous HBcAg are commonly weak,
predominantly due to restricted intracellular antigen delivery by
the lipophilic and selectively permeable biological membranes
(9). The cytoplasmic transduction
peptide (CTP) was specifically designed to ensure the efficient
delivery of CTP-fused biomolecules into the cytoplasm of cells
(10). Increasing evidence has
demonstrated that HBV-specific CTL responses are able to become
robust, polyclonal and multispecific reactions when exogenous HBcAg
is combined with CTP; such fusion leads HBcAg through cellular
membranes and into the antigen-presenting cell (APC) cytoplasm
(9,11).
The 76-residue polypeptide ubiquitin (Ub) is highly
conserved among eukaryotes (12)
and covalently binds the majority of proteins destined for
degradation by the ubiquitin-proteasome system (UPS) (13). This is critical for the generation
of the majority of peptides presented by class I major
histocompatibility complex (MHC) molecules (14,15).
The modification of antigens by ubiquitin or ubiquitin-like
proteins remodels their surface, triggering rapid degradation, thus
resulting in increased in vivo CTL responses to the
conjugated antigen (16,17). Therefore, Ub has been suggested to
be a potential therapeutic target for HBV treatment (18).
There is a possibility for exogenous HBcAg, with the
assistance of CTP, to enter the cytoplasm of APCs. When covalently
attached to exogenous Ub, HBcAg is rapidly degraded and presented
by MHC-I molecules, allowing fast and efficient action of CTL
against viral infections (19).
Consequently, a novel recombinant fusion protein, Ub-HBcAg-CTP, was
developed in the present study. Of note, Ub-HBcAg-CTP was able to
enter the cytoplasm of dendritic cells (DCs) and elicit robust
specific HBV immune responses in vitro (data not shown). The
present study aimed to assess whether the Ub-HBcAg-CTP fusion
protein were able to improve HBV-specific CTL immune responses and
anti-viral immunity in HBV transgenic mice.
Materials and methods
Fusion proteins and cell culture
The plasmid pcDNA3.1(-)-Ub-HBcAg was constructed and
maintained in the Laboratory Centre of The Sixth Hospital
affiliated to Shanghai Jiao Tong University (Shanghai, China)
(11). Briefly, the Ub-HBcAg cDNA
sequence was generated via polymerase chain reaction (PCR) to
obtain an 820 bp PCR product. The Ub-HBcAg-CTP gene and other
control genes, Ub-HBcAg and HBcAg-CTP, were amplified via PCR and
inserted into the pMAL-c2X prokaryotic expression vector
(Invitrogen Life Technologies, Carlsbad, CA, USA), respectively.
The constructed plasmids were further identified by restriction
enzyme digestion and bidirectional DNA sequencing (New England
Biolabs, Ipswich, MA, USA). Subsequently, the recombinant plasmids
were transformed into the host Escherichia coli BL21 (DE3)
cells (Tiangen, Beijing, China), which were induced to express the
recombinant fusion proteins. All fusion proteins were analyzed via
western blot analysis.
HBV transgenic mice and immunization
HBV transgenic mice (n=42; weight, 22–28g),
BALB/c-HBV1.3 (ayw subtype), which contained the 1.3-fold
over-length HBV genome, were purchased from the Key Liver Army
Laboratory (458 Hospital, Guangzhou, China). The mice were
maintained in 12 h light/dark cycles at a temperature of 2–25°C,
with free access to food and water. The detailed characterization
of these mice has been described previously (20). Groups of mice in all experiments
were matched for age (6–8 weeks; female) and all animals were
housed in the experimental animal centre of the Sixth Hospital
affiliated to Shanghai Jiao Tong University (Shanghai, China) under
specific pathogen-free conditions. All experiments were approved by
the Laboratory Animal Ethics Commission of Shanghai Jiao Tong
University (Shanghai, China). Mice were allowed 1 week of
adaptation and randomly divided into six groups (n=7).
Subsequently, the animals were immunized intramuscularly in the
tibialis anterior muscle three times at 1-week intervals with
Ub-HBcAg-CTP (50 µg), HBcAg-CTP (50 µg), Ub-HBcAg (50
µg) and interferon (IFN)-α (20,000 IU; Roche Diagnostics,
Basel, Switzerland), HBcAg (50 µg; CalBioreagents, Inc., San
Mateo, CA, USA), or phosphate-buffered saline (PBS; 50 µl;
Youkang, Beijing, China). At 1 week subsequent to the last
immunization, the mice were sacrificed by cervical dislocation
following anasthesia with 3% pentobarbital sodium (Sigma-Aldrich,
St. Louis, MO, USA), via intramuscular injection, and serum
samples, splenocytes and livers were collected.
Intracellular cytokine staining in
splenic lymphocytes
HBV transgenic mouse spleens were extracted for
splenocyte collection, by grinding of the spleens. T-lymphocytes
were obtained from splenocytes using nylon wool columns
(Polysciences Europe GmbH, Eppelheim, Germany). To evaluate the
percentage of IFN-γ-secreting cells in mouse splenocytes,
single-cell suspensions of harvested T cells were analyzed by flow
cytometry (Beckman Coulter, Inc., Brea, CA, USA). Following
incubation with 10 µg/ml HBcAg for 3 h, 25 µg/ml
phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich), 1
µg/ml ionomycin (Sigma-Aldrich) and 1.7 µg/ml
monensin (Sigma-Aldrich) were added to spleen T lymphocytes for
another 3 h (21). The cells were
then washed with PBS and stained with saturating concentrations of
phycoerythrin (PE)/Cy5-conjugated anti-CD3 mouse monoclonal
antibody (0.2 mg/ml; cat. no. 15-4888; eBioscience, Inc., San
Diego, CA, USA) and fluorescein isothiocyanate-conjugated anti-CD8α
mouse monoclonal antibody (0.2 mg/ml; cat. no. 11-0081;
eBioscience, Inc.) for 30 min at 25°C. Subsequent to
fixation/permeabilization with Fix and Perm reagent A and B (BD
Biosciences, Franklin Lakes, NJ, USA), cells were incubated with
PE-labeled anti-IFN-γ mouse monoclonal antibody (0.2 mg/ml; cat.
no. 12-7311; eBioscience, Inc.) for 30 min at 25°C, washed twice
with PBS and analyzed by flow cytometry on the Epics XL Flow
Cytometer (Beckman Coulter, Inc., Brea, CA, USA) using Expo 32-ADC
software (Beckman Coulter).
ELISA
For detection of interleukin (IL)-2 and IFN-γ,
spleen T lymphocytes (2×106 cells/ml) harvested from
immunized transgenic mice were cultured in 24-well plates
(1×106 cells/well) at 37°C in the presence of 10
µg/ml HBcAg. Subsequent to 72-h incubation, the cytokine
levels in cell supernatants were assessed using commercial mouse
cytokine (IL-2 and IFN-γ) ELISA kits (R&D Systems, Inc.,
Minneapolis, MN, USA), according to the manufacturers'
instructions. Results were expressed in pg/ml.
Enzyme-linked immunospot (ELISPOT)
assay
The ELISPOT assay (22) was used to evaluate HBcAg-specific
IFN-γ secretion in splenocytes. Spleen T lymphocytes
(1×105 cells/well pulsed with 10 µg/ml HBcAg)
were seeded in triplicate and incubated at 37°C for 20 h. A
positive control (phytohemagglutinin; 15 mg/ml; Dakewe Biotech Co.,
Ltd., Shenzhen, China) and a 'non-peptidic' negative control were
included in all assays (i.e. wells containing only medium).
Following incubation, cells were removed and ImmunoSpot plates
(Dakewe Biotech Co., Ltd., Shenzhen, China) pre-coated with the
anti-IFN-γ monoclonal antibody (BD Biosciences) were processed
according to the manufacturer's instructions. The number of spots
was counted using the Bioreader 4000 PRO-X (Bio-Sys GmbH, Karben,
Germany). Exclusively brown-colored spots with 'fuzzy borders' were
scored as spot-forming cells.
CTL assay
P815/c cells (Laboratory Centre of The Sixth
Hospital affiliated to Shanghai Jiao Tong University, Shanghai,
China) used as target cells, were seeded at a density of
5×104 cells/well in 96-well plates. The spleen T
lymphocytes (5×106 cells/well) were used as effector
cells and were co-cultured with P815/c cells at effector/target
(E/T) ratios of 5:1, 10:1 or 20:1, at 37°C in a humid environment
containing 5% CO2 for 4 h. The HBcAg-specific CTL
activity was measured by lactate dehydrogenase release using a
CytoTox 96® Non-Radioactive Cytotoxicity kit (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
instructions. The absorbance values of supernatants were recorded
at 490 nm on an MK3 Multiscan (Thermo Labsystems, Waltham, MA,
USA). The cytotoxicity was calculated as follows: [(Experimental
release - effector spontaneous release - target spontaneous
release) / (target maximum release - target spontaneous release)]
×100% (23).
Serology
Venous blood was collected from transgenic mice at 1
week following the second and third immunizations, respectively.
Serum HBV DNA and HBsAg levels were assessed using quantitative PCR
(Terra PCR Direct Polymerase mix) and Abbott kits (Abbott
Diagnostics, Chicago, IL, USA) separately, as described previously
(24). The inhibitory rate of HBV
DNA and HBsAg induced by the fusion protein was assessed. In
addition, serum alanine aminotransferase (ALT) activities were
measured at 7 days subsequent to the third injection using the
ARCHITECT Automatic Biochemistry Analyzer (Abbott Diagnostics).
Histology and immunohistochemistry of
liver tissues
De-paraffinized 5-µm sections were stained
with hematoxylin-eosin (Beyotime Institute of Biotechnology,
Shanghai, China). For immunohistochemical analysis, slides were
incubated for 1 h at 60°C and de-paraffinized. Following antigen
retrieval using citrate buffer (pH=6.0; Beyotime Institute of
Biotechnology), endogenous peroxidase was quenched by 3%
H2O2 (Bioworld, Nanjing, China) in deionized
H2O for 5 min and non-specific binding blocked by 2%
normal goat serum (Novus Biologicals LLC, Littleton, CO, USA) for 3
h at room temperature. Samples were then incubated with goat
anti-HBsAg polyclonal antibody (Novus Biologicals LLC, Littleton,
CO, USA) and goat anti-HBcAg polyclonal antibody (Novus Biologicals
LLC) at 1:500 dilutions and 4°C overnight, respectively, followed
by addition of biotinyated secondary antibody (Wuhan Boster
Biological Technology, Ltd., Wuhan, China) at 37°C for 30 min with
streptavidin-biotin-peroxidase complex (Biorbyt, Cambridgeshire,
UK). Detection was conducted using diaminobenzidine (Sigma-Aldrich)
and cells were counterstained with hematoxylin (20).
Statistical analysis
Values are expressed as the mean ± standard
deviation and all analyses were performed with SPSS software,
version 16.0 (SPSS, Inc., Chicago, IL, USA). One-way analysis of
variance and the post-hoc least significant difference test were
used in order to determine statistical significance. P<0.05 was
considered to indicate a statistically significant difference.
Results
Ub-HBcAg-CTP elicits a robust and
functional specific cellular immune response
The specific CD8+ T-cell responses
induced by the fusion proteins were assessed by intracellular IFN-γ
staining of T cells isolated 1 week subsequent to the last
immunization. Ub fused with HBcAg-CTP, which induced a clear T cell
immune response, displaying the largest number of CD8+
IFN-γ+ T lymphocytes in the spleens, as detected by flow
cytometry. The CD8+ IFN-γ+ T cells
represented up to 2.2% of the total spleen T lymphocytes in mice
immunized with Ub-HBcAg-CTP (Fig.
1), compared with 1.7, 1.4, 1.1, 0.3 and 0.1% identified in
mice that received HBcAg-CTP, IFN-α, Ub-HBcAg, HBcAg and the PBS
control, respectively. These observations suggested that delivery
of ubiquitin and HBcAg was mediated via CTP-enhanced HBcAg-specific
CTL generation in HBV transgenic mice.
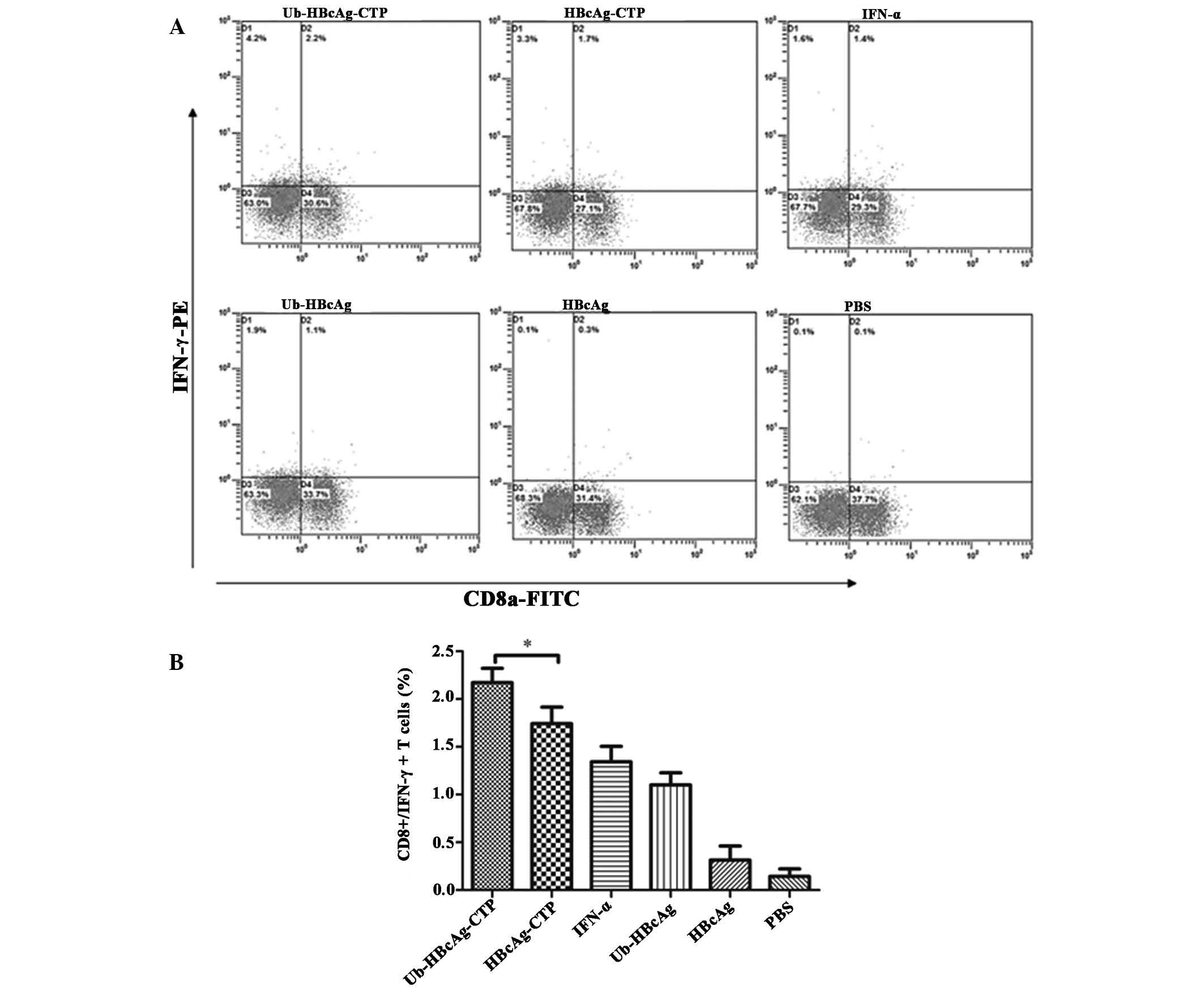 | Figure 1Intracellular cytokine expression in
splenic lymphocytes of transgenic mice. (A) One week subsequent to
the third immunization, mice were euthanized and splenic
lymphocytes were isolated and re-stimulated in vitro by
HBcAg for 3 h, followed by addition of 25 µg/ml PMA, 1
µg/ml ionomycin and 1.7 µg/ml monensin for an
additional 3 h. Re-stimulated T cells were stained with fluorescent
FITC-CD8α and PE-IFN-γ antibodies, then the doubly stained cells
were counted and analyzed by flow cytometry. (B) Values are
expressed as the mean ± standard deviation (n=7) and are
representative of a minimum of three individual experiments.
*P<0.05. HBcAg, hepatitis B core antigen; PMA,
phorbol 12-myristate 13-acetate; FITC, fluorescein isothiocyanate;
PE, phycoerythrin; IFN, interferon; Ub, ubiquitin; CTP, cytoplasmic
transduction peptide; PBS, phosphate-buffered saline. |
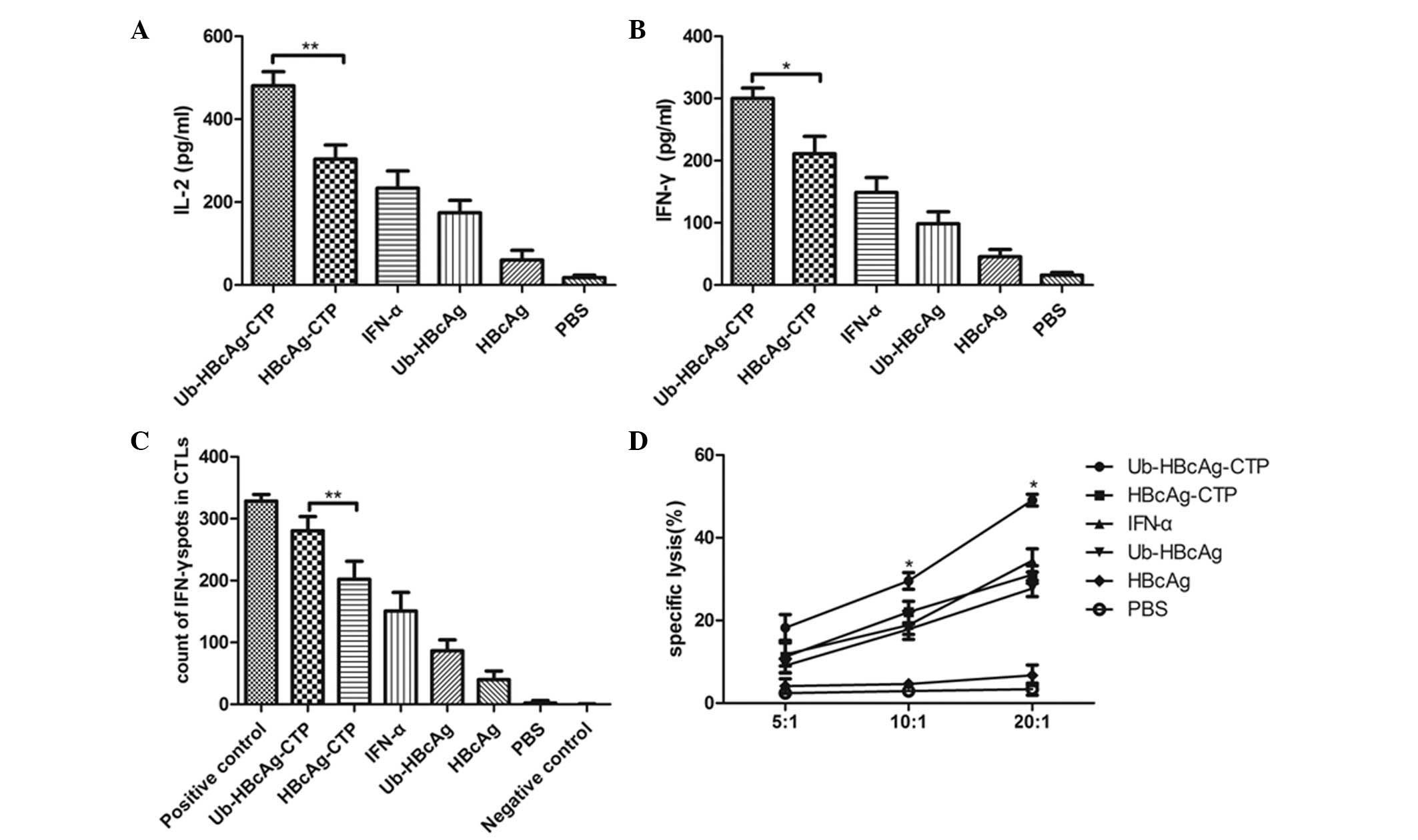
Ub-HBcAg-CTP boosts IFN-γ and IL-2
production
One week subsequent to the third intramuscular
injection of different fusion proteins, mice exhibited distinct
levels of Th1 cell-derived cytokines as detected by ELISA. Fig. 2A and B illustrates the levels of
IL-2 and IFN-γ secreted by T cells upon re-stimulation with HBcAg.
The highest production of IL-2 (480.9±33.7 pg/ml) and IFN-γ
(288.2±16.93 pg/ml) was observed in mice immunized with
Ub-HBcAg-CTP.
Ub-HBcAg-CTP enhances the specific CTL
response
ELISPOT assays were conducted to quantify the
IFN-γ-producing lymphocytes. A higher percentage of T cells
harvested from Ub-HBcAg-CTP-immunized animals produced IFN-γ
(Fig. 2C), in comparison with that
in the control group. In general, these results were in agreement
with the above flow cytometry results for IFN-γ expression in
CD8+ T cells. Overall, these observations indicated that
Ub-HBcAg-CTP enhanced HBcAg-specific CTL responses in
vivo.
To verify the role of Ub-HBcAg-CTP in cell-mediated
immune responses, the cytolytic activity of HBcAg-specific
CD8+ T cells was determined by their ability to kill
peptide-loaded target cells (P815/c). As presented in Fig. 2D, values of 49.09±1.44, 29.55±2.01
and 18.2±3.26% for specific cytolysis were obtained for
Ub-HBcAg-CTP-immunized mice at E/T ratios of 20:1, 10:1 and 5:1,
respectively. The percentage of specific lysis was significantly
higher in mice treated with Ub-HBcAg-CTP than that in animals
immunized with other control proteins (*P<0.05).
These observations suggested that Ub-HBcAg-CTP induced specific CTL
activity, in accordance with high IFN-γ levels in CD8+ T
cells.
HBsAg, HBV DNA and ALT in serum samples
from HBV transgenic mice
Subsequently, the virus clearance following
immunization was evaluated using the fusion proteins. The serum HBV
DNA load of transgenic mice was determined by quantitative PCR to
assess the curative effect of HBV-derived epitope-specific
CD8+ T cells at 1 week subsequent to the second and
third immunizations, respectively. In parallel, serum HBsAg levels
were detected at the same time-points. Of note, serum HBsAg levels
were markedly reduced in the Ub-HBcAg-CTP group compared with those
in the other groups, including HBcAg-CTP, Ub-HBcAg and IFN-α, as
presented in Fig. 3A
(*P<0.05). Likewise, serum HBV DNA titers were
significantly reduced in mice immunized with Ub-HBcAg-CTP, compared
with those in the HBcAg-CTP, Ub-HBcAg and IFN-α groups (Fig. 3B; *P<0.05). These
results suggested that specific CD8+ T cells induced by
Ub-HBcAg-CTP produced IFN-γ, mediating the inhibition of HBV
replication in HBV transgenic mice. Significant elevations in ALT
levels were observed in serum samples from mice treated with
Ub-HBcAg-CTP (Fig. 3C;
*P<0.05).
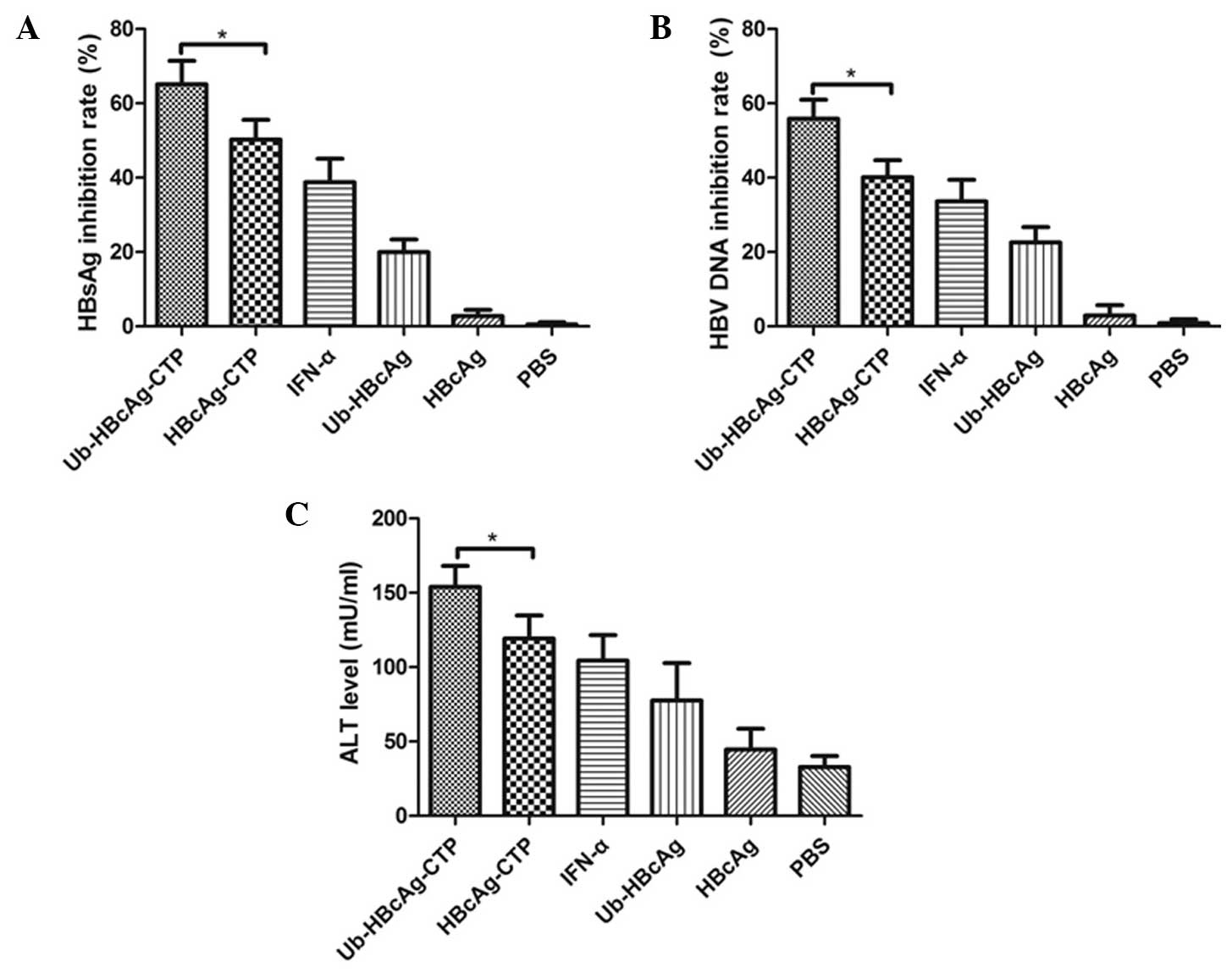 | Figure 3In vivo inhibitory effects on
HBsAg, HBV DNA and ALT in HBV transgenic mice immunized with
different fusion proteins. Mice were bled at 1 week following the
second and third immunizations, respectively, and the levels of
HBsAg and HBV DNA were quantified. (A) HBsAg and (B) HBV DNA
inhibitory rates in serum samples from mice treated with different
fusion proteins. (C) Mean levels (mU/ml) of sALT in the six groups
of mice were examined at 7 days subsequent to the third injection.
Values are expressed as the mean ± standard deviation (n=7) and are
representative of a minimum of three individual experiments.
*P<0.05. HBsAg, hepatitis B surface antigen; HBV,
hepatitis B virus; sALT, serum alanine aminotransferase; IFN-α,
interferon-α; Ub, ubiquitin; CTP, cytoplasmic transduction peptide;
PBS, phosphate-buffered saline. |
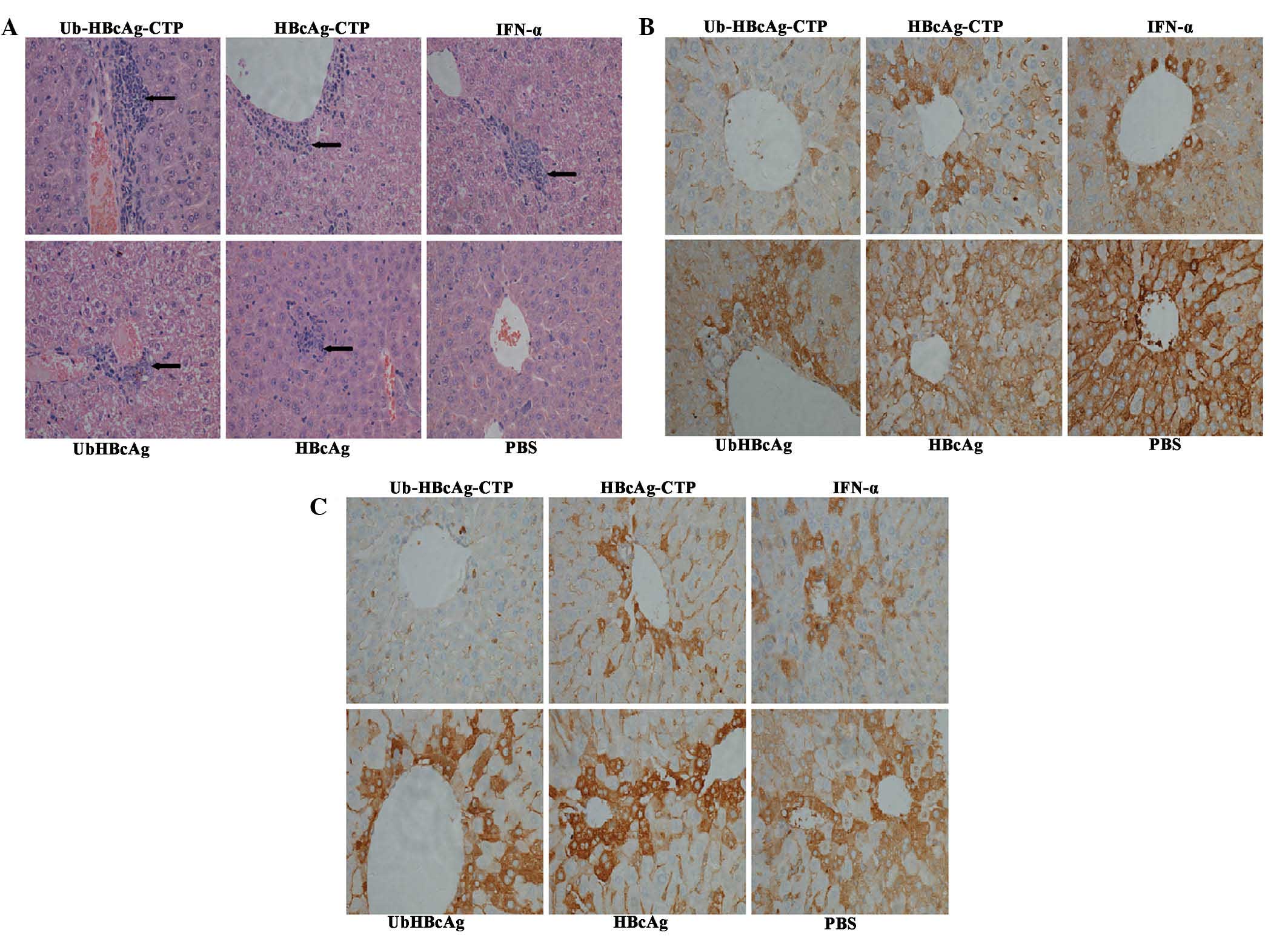
Histopathological alterations
Liver tissue sections were evaluated primarily in
terms of hepatocyte degeneration and necrosis in addition to
lymphocyte infiltration. A larger number of lymphocytes appeared in
the livers from mice immunized with Ub-HBcAg-CTP (Fig. 4A), compared with that in the other
groups. To further confirm the therapeutic effects of the fusion
proteins in transgenic mice, immunohis-tological analysis was
conducted. While high quantities of HBsAg and HBcAg were observed
(brownish yellow stain) in the cytoplasm of hepatocytes from
PBS-treated mice, they were not apparent in the
Ub-HBcAg-CTP-treated group (Fig. 4B
and C). Therefore, it was suggested that immunization with
Ub-HBcAg-CTP reduced HBsAg and HBcAg levels, to a certain
extent.
Discussion
Antigen-based immunotherapy (vaccine therapy) is
considered a promising therapeutic strategy for HBV infections,
controlling HBV replication in chronic hepatitis B by inducing Th1
immunity and enhancing HBV-specific immune responses (25,26).
Therefore, a novel therapeutic vaccine (Ub-HBcAg-CTP) was designed
for effective activation of Th1 immunity and the specific
CD8+ T cell response, which should compensate for the
deficient HBV-specific anti-viral immunity and would not be subject
to functional exhaustion during chronic HBV infections.
In the present study, HBV transgenic mice whose
hepatocytes replicated the virus at levels comparable to those
observed in the livers of patients with chronic hepatitis and who
had no evidence of cytopathology were used. This model enabled the
assessment of the effects of viral and host factors on HBV
pathogenesis and replication, and the evaluation of the anti-viral
potential of pharmacological agents and physiological processes,
including the immune response (20).
Efficient T-cell responses require CD8+ T
cells to recognize antigenic peptides presented by MHC I molecules
on the surface of APCs (predominantly DCs) (27). However, patients with chronic
hepatitis B commonly display an immunocompromised immune tolerance
with impaired DC function (28,29).
CTP, a novel transduction carrier, has been demonstrated to be
efficient in the delivery of antigens into the cytoplasm of DCs
(30). Therefore, CTP was used in
the present study as a tool for efficient delivery of Ub and HBcAg,
thus boosting DC antigen-presenting capacities and inducing a
greater number of HBV-specific CD8+ T cells.
Defects in cytokine secretion of antigen-specific
CD8+ T cells have been previously associated with
chronic HBV infection (31).
Therefore, the induction of a strong, polyclonal, potent
multifunctional HBV-specific CD8+ T-cell response is
highly desirable, due to the fact that this would be able to
trigger cytokine secretion. As mentioned above, CD8+ T
cells induced by Ub-HBcAg-CTP vaccination were highly functional.
Mice immunized with Ub-HBcAg-CTP exhibited significantly higher
IFN-γ and IL-2 (Th1-like) secretion and HBcAg-specific
CD8+/IFN-γ+ T cells in the spleen, compared
with those in the other treatment groups. These observations
indicated that the Thl dominant responses were associated with
significant enhancement of CTL activity following Ub-HBcAg-CTP
vaccination. Th1 immunity appears to be crucial for the induction
of CTL leading to cytolytic effects, which are beneficial for viral
or tumor eradication (32).
Furthermore, the in vivo cytotoxicity data from the present
study demonstrated that the higher magnitude of CTL response
induced by Ub-HBcAg-CTP was correlated with an increase in cell
death of HBcAg-derived peptide-loaded target cells (P815/c).
Therefore, a complete response to anti-viral treatment was
correlated with predominant Th1 responses accompanied with enhanced
CTL activity in patients with chronic HBV or transgenic mice
(33). This implied that
activation of Thl immunity accompanied by efficient CTL activity
subsequent to vaccination therapy is a common immune mechanism for
successful treatment of hepatitis B (34).
It has been previously demonstrated that inadequate
endogenous antigen presentation by MHC class I molecules to
CD8+ T cells is one mechanism by which the immune system
fails to eliminate HBV (35).
Therefore, efficient elimination of virus-infected target cells by
CTLs is only able to occur by rapid MHC class I antigen
presentation of viral epitopes on the cell surface (36). Ub-HBcAg-CTP, with a modification
for ubiquitin conjugation of antigen protein, exposes its
N-terminal residue ('the N-end rule') (37,38),
which further facilitates ubiquitination. In this manner, a
polyubiquitin chain is synthesized, which targets the protein for
rapid degradation by the UPS (39). It was identified in the present
study that the enhancement of Ub-antigen presentation increased the
number of HBcAg-specific CD8+/IFN-γ+ T cells
in Ub-HBcAg-CTP immunized mice. This may be due to the fact that
increased Ub-fused HBcAg is rapidly degraded by the UPS, which
results in efficient production of a variety of peptides, including
numerous CTL epitopes, which may be presented by multiple MHC class
I molecules. Additional studies have demonstrated that rapidly
ubiquitinated antigens are more rapidly presented on class I
molecules and/or highly induced (40,41).
The immunization of transgenic mice with
Ub-HBcAg-CTP induced an efficient specific immune response and led
to the control of viral replication. An increasing number of
studies have demonstrated that secreted cytokines and activated
CTLs may effectively downregulate HBV gene expression and
additionally may control HBV replication (23,42).
At 1 week subsequent to the last immunization to HBV transgenic
mice, the hepatocytes appeared to swell and exhibited hyperemia and
lymphocyte infiltration in liver tissues from Ub-HBcAg-CTP animals.
The inflammatory reactions in the livers were consistent with
specific CTL activity induced by Ub-HBcAg-CTP. In addition, the
results of the present study indicated that the fusion peptide
significantly reduced serum HBsAg and HBV DNA levels in addition to
the expression of HBcAg and HBsAg in the liver. This reduction was
closely associated with Ub-HBcAg-CTP, suggesting that the observed
therapeutic effects were associated with the enhanced immune
responses. These observations demonstrated the potential of
Ub-HBcAg-CTP to induce a stronger CTL response in transgenic mice
than control vaccinations. Akbar et al (43) reported that the strong
immunomodulatory capabilities of HBcAg may be due to an
establishment of an inflammatory hepatic micro-environment, the
induction of HBcAg-specific CTL in the liver and the activation of
host DCs. To further confirm the specific CTL activity in the
liver, the levels of ALT were measured. In contrast with control
animals, greater ALT levels were observed in Ub-HBcAg-CTP immunized
mice, indicating that infected hepatocytes were eliminated by
vaccination-induced cytotoxic T cells. Although the activation
levels of HBcAg-specific CTL in the liver were not assessed in the
present study, the histological and serological alterations
observed provided indirect support for the hypothesis that an
anti-inflammatory hepatic microenvironment may be established in
the Ub-HBcAg-CTP group. However, this remains to be confirmed in
future studies.
In conclusion, the present study demonstrated that
vaccination with Ub-HBcAg-CTP activated Th1 immunity, induced a
robust and multifunctional HBcAg-specific T-cell response and
provided a therapeutic effect in HBV transgenic mice. Therefore, a
combination of Ub-HBcAg-CTP may be used as a potential therapeutic
strategy for the treatment of chronic hepatitis B viral
infections.
Acknowledgments
The present study was supported by the grants from
the National Natural Science Foundation of China (grant no.
81270502).
References
|
1
|
Custer B, Sullivan SD, Hazlet TK, Iloeje
U, Veenstra DL and Kowdley KV: Global epidemiology of hepatitis B
virus. J Clin Gastroenterol. 38:S158–S168. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Perz JF, Armstrong GL, Farrington LA,
Hutin YJ and Bell BP: The contributions of hepatitis B virus and
hepatitis C virus infections to cirrhosis and primary liver cancer
worldwide. J Hepatol. 45:529–538. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Buchmann P, Dembek C, Kuklick L, et al: A
novel therapeutic hepatitis B vaccine induces cellular and humoral
immune responses and breaks tolerance in hepatitis B virus (HBV)
transgenic mice. Vaccine. 31:1197–1203. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rehermann B: Immune responses in hepatitis
B virus infection. Semin Liver Dis. 23:21–38. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baumert TF, Thimme R and Von Weizsacker F:
Pathogenesis of hepatitis B virus infection. World J Gastroenterol.
13:82–90. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thimme R, Wieland S, Steiger C, et al:
CD8+ T cells mediate viral clearance and disease pathogenesis
during acute hepatitis B virus infection. J Virol. 77:68–76. 2003.
View Article : Google Scholar :
|
|
7
|
Chen W, Shi M, Shi F, et al: HBcAg-pulsed
dendritic cell vaccine induces Th1 polarization and production of
hepatitis B virus-specific cytotoxic T lymphocytes. Hepatol Res.
39:355–365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang BF, Zhao HL, Xue C, et al:
Recombinant heat shock protein 65 carrying hepatitis B core antigen
induces HBcAg-specific CTL response. Vaccine. 25:4478–4486. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang Y, Chen X, Zhang Y, et al: Fusion
protein of tapasin and hepatitis B core antigen 18–27 enhances T
helper cell type 1/2 cytokine ratio and antiviral immunity by
inhibiting suppressors of cytokine signaling family members 1/3 in
hepatitis B virus transgenic mice. Mol Med Rep. 9:1171–1178.
2014.PubMed/NCBI
|
|
10
|
Kim D, Jeon C, Kim JH, et al: Cytoplasmic
transduction peptide (CTP): new approach for the delivery of
biomolecules into cytoplasm in vitro and in vivo. Exp Cell Res.
312:1277–1288. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song L, Zhuo M, Tang Y, Chen X, Yu Y, Tang
Z, et al: Ubiquitin-modified hepatitis B virus core antigen
effectively facilitates antigen presentation and enhances cytotoxic
T lymphocyte activity via the cytoplasmic transduction peptide
invitro. Mol Med Rep. 12:289–296. 2015.PubMed/NCBI
|
|
12
|
Kerscher O, Felberbaum R and Hochstrasser
M: Modification of proteins by ubiquitin and ubiquitin-like
proteins. Annu Rev Cell Dev Biol. 22:159–180. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goldberg AL: Protein degradation and
protection against misfolded or damaged proteins. Nature.
426:895–899. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bandi P, Garcia ML, Booth CJ, Chisari FV
and Robek MD: Bortezomib inhibits hepatitis B virus replication in
transgenic mice. Antimicrob Agents Chemother. 54:749–756. 2010.
View Article : Google Scholar :
|
|
15
|
Loureiro J and Ploegh HL: Antigen
Presentation and the ubiquitin-proteasome system in host-pathogen
interactions. Adv Immunol. 92:225–305. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kerscher O, Felberbaum R and Hochstrasser
M: Modification of proteins by ubiquitin and ubiquitin-like
proteins. Annu Rev Cell Dev Biol. 22:159–180. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Welchman RL, Gordon C and Mayer RJ:
Ubiquitin and ubiq-uitin-like proteins as multifunctional signals.
Nat Rev Mol Cell Biol. 6:599–609. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song L, Zhuo M, Tang Y, Chen X, Tang Z and
Zang G: Ubiquitin-hepatitis B core antigen-cytoplasmic transduction
peptide enhances HBV-specific humoral and CTL immune responses in
vivo. International Immunopharmacology. 23:1–7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen JH, Yu YS, Chen XH, Liu HH, Zang GQ
and Tang ZH: Enhancement of CTLs induced by DCs loaded with
ubiqui-tinated hepatitis B virus core antigen. World J
Gastroenterol. 18:1319–1327. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guidotti LG, Matzke B, Schaller H and
Chisari FV: High-level hepatitis B virus replication in transgenic
mice. J Virol. 69:6158–6169. 1995.PubMed/NCBI
|
|
21
|
Crawford TQ, Ndhlovu LC, Tan A, et al:
HIV-1 infection abrogates CD8+T cell mitogen-activated protein
kinase signaling responses. J Virol. 85:12343–12350. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lycke NY and Coico R: Measurement of
immunoglobulin synthesis using the ELISPOT assay. Curr Protoc
Immunol. 14:2001. View Article : Google Scholar
|
|
23
|
Chen X, Lai J, Pan Q, Tang Z, Yu Y and
Zang G: The delivery of HBcAg via Tat-PTD enhances specific immune
response and inhibits Hepatitis B virus replication in transgenic
mice. Vaccine. 28:3913–3919. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang Y, Chen Z, Jia H, Wu W, Zhong S and
Zhou C: Induction of Tc1 response and enhanced cytotoxic T
lymphocyte activity in mice by dendritic cells transduced with
adenovirus expressing HBsAg. Clin Immunol. 119:280–290. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Boni C, Fisicaro P, Valdatta C, et al:
Characterization of hepatitis B virus (HBV)-specific T-cell
dysfunction in chronic HBV infection. J Virol. 81:4215–4225. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Deng Q, Mancini-Bourgine M, Zhang X, et
al: Hepatitis B virus as a gene delivery vector activating foreign
antigenic T cell response that abrogates viral expression in mouse
models. Hepatology. 50:1380–1391. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Neefjes J, Jongsma ML, Paul P and Bakke O:
Towards a systems understanding of MHC class I and MHC class II
antigen presentation. Nat Rev Immunol. 11:823–836. 2011.PubMed/NCBI
|
|
28
|
Tavakoli S, Mederacke I, Herzog-Hauff S,
et al: Peripheral blood dendritic cells are phenotypically and
functionally intact in chronic hepatitis B virus (HBV) infection.
Clin Exp Immunol. 151:61–70. 2008. View Article : Google Scholar
|
|
29
|
Op den Brouw ML, Binda RS, Van Roosmalen
MH, et al: Hepatitis B virus surface antigen impairs myeloid
dendritic cell function: a possible immune escape mechanism of
hepatitis B virus. Immunology. 126:280–289. 2009. View Article : Google Scholar :
|
|
30
|
Chen X, Liu H, Tang Z, Yu Y and Zang G:
The modification of Tapasin enhances cytotoxic T lymphocyte
activity of intracel-lularly delivered CTL epitopes via cytoplasmic
transduction peptide. Acta Biochim Biophys Sin (Shanghai).
45:203–212. 2013. View Article : Google Scholar
|
|
31
|
Das A, Hoare M, Davies N, et al:
Functional skewing of the global CD8 T cell population in chronic
hepatitis B virus infection. J Exp Med. 205:2111–2124. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chamoto K, Kosaka A, Tsuji T, et al:
Critical role of the Th1/Tc1 circuit for the generation of
tumor-specific CTL during tumor eradication in vivo by Th1-cell
therapy. Cancer Sci. 94:924–928. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsai SL, Sheen IS, Chien RN, et al:
Activation of Th1 immunity is a common immune mechanism for the
successful treatment of hepatitis B and C: tetramer assay and
therapeutic implications. J Biomed Sci. 10:120–135. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Boni C, Bertoletti A, Penna A, et al:
Lamivudine treatment can restore T cell responsiveness in chronic
hepatitis B. J Clin Invest. 102:968–975. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kosinska AD, Johrden L, Zhang E, et al:
DNA prime-adenovirus boost immunization induces a vigorous and
multifunctional T-cell response against hepadnaviral proteins in
the mouse and woodchuck model. J Virol. 86:9297–9310. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Seifert U and Krüger E: Remodelling of the
ubiq-uitin-proteasome system in response to interferons. Biochem
Soc Trans. 36:879–884. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Varshavsky A: The N-end rule. Cell.
69:725–735. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ciechanover A and Ben-Saadon R: N-terminal
ubiquitination: more protein substrates join in. Trends Cell Biol.
14:103–106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rock KL and Goldberg AL: Degradation of
cell proteins and the generation of MHC class I-presented peptides.
Ann Rev Immunol. 17:739–779. 1999. View Article : Google Scholar
|
|
40
|
Rodriguez F, Zhang J and Whitton JL: DNA
immunization: ubiquitination of a viral protein enhances cytotoxic
T-lymphocyte induction and antiviral protection but abrogates
antibody induction. J Virol. 71:8497–8503. 1997.PubMed/NCBI
|
|
41
|
Liu Y, Testa JS, Philip R, Block TM and
Mehta AS: A ubiquitin independent degradation pathway utilized by a
hepatitis B virus envelope protein to limit antigen presentation.
PLoS One. 6:e244772011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Roh S and Kim K: Overcoming tolerance in
hepatitis B virus transgenic mice: a possible involvement of
regulatory T cells. Microbiol Immunol. 47:453–460. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Akbar SM, Chen S, Al-Mahtab M, Abe M,
Hiasa Y and Onji M: Strong and multi-antigen specific immunity by
hepatitis B core antigen (HBcAg)-based vaccines in a murine model
of chronic hepatitis B: HBcAg is a candidate for a therapeutic
vaccine against hepatitis B virus. Antiviral Res. 96:59–64. 2012.
View Article : Google Scholar : PubMed/NCBI
|