Introduction
Gliomas are a common type of primary tumor of the
central nervous system and account for 45.2% of all intracranial
types of tumor (1). The annual
incidence of brain glioma ranges between 0.003 and 0.005% (2). Brain gliomas exhibit diffuse and
invasive characteristics, and traditional resection has a poor
prognosis (3,4). In addition, drug resistance in brain
gliomas leads to unsatisfactory radiotherapeutic and
chemotherapeutic outcomes and relatively short survival rates
(5). Therefore, investigations on
the mechanisms underlying brain glioma metastasis and invasion are
important for the development of anti-glioma drugs.
The multi-domain signaling molecule girdin, which
was discovered by a Japanese group in 2005, acts as a signaling
transduction bridge and link (6).
Girdin is an actin-binding protein involved in the regulation of
the actin cytoskeleton, thus affecting pseudopodia extension
(7) and regulating the migration
of endothelial, smooth muscle and neural stem cells (8). Girdin also promotes vascular
endothelial growth factor (VEGF)-mediated endothelial cell
migration and regulates neointimal formation following vascular
injury (9,10). Girdin is expressed in various types
of cancer, including esophageal cancer, gastric cancer, colon
cancer, breast cancer and lung cancer, and its expression is
particularly high in invasive tumor cells (11). Girdin is also involved in the
regulation of tumor metastasis (11,12),
angiogenesis (9,10) and autophagy (13) and has a close association with
tumorigenesis and development (12). High expression levels of girdin
also correlate closely with poor tumor prognosis (11).
Girdin is expressed at relatively high levels in
brain gliomas. A previous study reported an association between the
expression of girdin and glioma metastasis (14), and the expression levels of girdin
correlate with the degree of malignancy in brain gliomas. However,
the role of girdin in brain gliomas remains to be elucidated. The
present study aimed to determine the effects of short hairpin
(sh)RNA-induced girdin silencing on the proliferation, migration
and invasion of glioma cells.
Materials and methods
Cell culture
Human glioma cell lines U251, A172, U87-MG and
SHG-44 were purchased from Shanghai Institutes for Biological
Sciences, Chinese Academy of Sciences (Shanghai, China). U373 was
purchased from Bioleaf Biotech Co., Ltd (Shanghai, China). U251,
U373 and A172 cells were cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco Life Sciences, Grand Island, NY, USA)
supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT,
USA). U87-MG cells were cultured in minimum essential medium (Gibco
Life Sciences) supplemented with 15% FBS. SHG-44 cells were
cultured in RPMI-1640 (Gibco Life Sciences) supplemented with 10%
FBS. All cells were incubated in a 37°C incubator with 5%
CO2.
Construction of girdin RNA interference
vectors and transfection
The girdin short hairpin (sh)RNA sequences and
unrelated sequences were designed and synthesized by Sangon Biotech
(Shanghai) Co., Ltd. (Shanghai, China). The sequences were as
follows: Interference sequence, forward 5′-GAT CCC CGT CAA TAA TGA
TGC CTC ACT TCA AGA GAG TGA GGC ATC ATT ATT GAC TTTTT-3′ and
reverse 5′-AGC TAA AAA GTC AAT AAT GAT GCC TCA CTCTCT TGA AGT GAG
GCA TCA TTA TTG ACGGG-3; and unrelated sequence, forward 5′-GAT CCC
CTT CTC CGA ACG TGT CAC GTT TCA AGA GAA CGT GAC ACG TTC GGA GAA
TTTTT-3′ and reverse 5′-AGC TAA AAA TTC TCC GAA CGT GTC ACG TTC TCT
TGA AAC GTG ACA CGT TCG GAG AAGGG-3′. The annealed interference
sequences or unrelated sequences were inserted into
pGCsi-H1/Neo/GFP vectors via digestion with FastDigest BamHI
and FastDigest Hind III (Thermo Fisher Scientific, Pittsburgh, PA,
USA), and ligation with T4 DNA Ligase (Thermo Fisher Scientific).
The sequencing-confirmed plasmids were termed girdin shRNA and
negative control (NC) respectively. The girdin shRNA or NC plasmids
were subsequently transfected into the U251 cells using
Lipofectamine 2000 (Invitrogen Life Technologies, Carlsbad, CA,
USA), according to the manufacturer's instructions. At 24 h
post-transfection, the culture medium was replaced with medium
containing 400 µg/ml G418 (Invitrogen Life Technologies) for
the screening of stably transfected cells. Positive clones were
identified using quantitative polymerase chain reaction (qPCR) and
western blot analysis.
Drug treatment
When the cells transfected with girdin shRNA or NC
grew to 70–80% confluency, the LY294002 PI3K inhibitor (20
µM; Beyotime Instutute of Biotechnology, Shanghai, China) or
dimethylsulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO, USA) was
added to the cells. Following incubation for an additional 24 h, a
Transwell invasion assay and western blot analysis were
performed.
Analysis of cell viability
Cell viability was examined using a
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide
(MTT) assay. The cells transfected with girdin shRNA or NC, as well
as untreated U251 cells, were seeded into 96-well plates
(2×103 cells per well) and incubated at 37°C. MTT
solution (0.2 mg/ml, Sigma-Aldrich) was added after 0, 24, 48, 72
or 96 h incubation. Following incubation for an additional 4 h at
37°C, the supernatants were removed and 200 µl DMSO was
added to each well to dissolve the formazan crystals. The 96-well
plates were placed in a microplate reader (ELX-800; Biotek,
Winooski, VT, USA) to measure the absorbance at 490 nm, and growth
curves were plotted.
Wound healing assay
The cells were seeded into 6-well plates
(1×105 cells per well). When the cells reached 80–90%
confluency, scratches were introduced onto the monolayer cell
surfaces using 200 µl pipette tips. The cells were
subsequently washed with serum-free medium, and then incubated with
serum-free DMEM at 37°C. At 0, 12, and 24 h, images of each sample
were captured and the data were recorded to calculate the relative
cell mobility, based on the following formula: Relative mobility =
(1 − distance between the edges of migrated scratches / distance
between the edges of initial scratches) ×100%.
Transwell invasion assay
The cells were suspended in serum-free medium at a
density of 1×105 cells/ml. Subsequently, 200 µl
cell suspension was added to the upper chamber of Transwell
chambers (Corning Life Sciences, Tewksbury, MA, USA) pretreated
with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA), and 800
µl DMEM media supplemented with 30% FBS was added to each
lower chamber. Following incubation in the incubator for another 24
h at 37°C, the cells on the upper surface of the microporous
membrane were removed with cotton swabs, whereas the cells on the
lower surface of the membrane were fixed in 4% paraformaldehyde
solution (Sinopharm, Shanghai, China) for 20 min at room
temperature and subsequently stained with 0.5% crystal violet
solution (Amresco, Solon, OH, USA). Images of the stained cells
from five selected views were captured under a light microscope
(AE31; Motic Incoporation, Ltd., Xiamen, China) at 200×
magnification, and the number of cells, which migrated through the
micro-porous membranes was calculated.
Western blot analysis
The cells were collected via centrifugation and the
protein in the cells was extracted using NP-40 lysis buffer
(Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. The protein concentrations were
measured using a BCA Protein Concentration Detection kit (Beyotime
Institute of Biotechnology). Subsequently, 40 µg of the
protein was separated via 10% sodium dodecyl sulfate polyacrylamide
gel electrophoresis (SDS-PAGE), followed by transferring onto
polyvinylidene fluoride (PVDF) membranes (EMD Millipore, Bedford,
MA, USA). The PVDF membranes containing the transferred proteins
were blocked in 5% skim milk or 5% bovine serum albumin (Biosharp,
Hefei, China). Following washing with Tris-buffered saline with
0.05% Tween-20 (TBST), the membranes were incubated with the
following primary antibodies at 4°C overnight: Rabbit anti-human
polyclonal antibody against girdin (1:500 diluted; cat. no.
bs-5150R; Bioss, Beijing, China); rabbit anti-human polyclonal
antibody against MMP-2 (1:1,000 diluted; cat. no. WL0657); rabbit
anti-human polyclonal antibody against MMP-9 (1:1,000 diluted; cat.
no. WL0884); rabbit anti-human polyclonal antibody against P85α
(1:1,000 diluted; cat. no. WL0191); rabbit anti-human polyclonal
antibody against P110α (1:1,000 diluted; cat. no. WL0339); rabbit
anti-human polyclonal antibody against AKT (1:1,000 diluted; cat.
no. WL0003); rabbit anti-human polyclonal antibody against p-AKT
(1:1,000 diluted; cat. no. WLP001). All antibodies were purchased
from Wanleibio (Shenyang, China) unless stated. Following washing
with TBST, the membranes were subsequently incubated with
horseradish peroxidase-linked goat anti-rabbit IgG (1:5,000,
Beyotime Institute of Biotechnology) at 37°C for 45 min. Enhanced
chemiluminescence solution was added for luminescent image
development, and the target protein levels were analyzed using a
Gel-Pro-Analyzer (Liuyi, Beijing, China), with β-actin as a
reference.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the cells using a Total
RNA Extraction kit (Tiangen Biotech, Co., Ltd., Beijing, China)
according to the manufacturer's instructions. The extracted RNA
from each group was reverse transcribed into cDNA using Super M-MLV
reverse transcriptase (BioTeke Corporation, Beijing, China) and
oligo (dT)15. The mRNA expression levels of girdin,
MMP-2 and MMP-9 were detected by RT-qPCR on an Exicycler TM 96
Real-Time Quantitative PCR instrument (Bioneer Corporation,
Daejeon, Korea) using cDNA as a template. The PCR reaction
conditions were as follows: 95°C for 10 min; 95°C for 10 sec, 60°C
for 20 sec, 72°C for 30 sec (40 cycles); 4°C for 5 min. The
relative mRNA levels in each sample were calculated using the
2−ΔΔCt method (15),
using β-actin as a reference. The SYBR Green qRT-PCR Master mix was
purchased from Beijing Solarbio Science & Technology Co., Ltd
(Beijing, China). The following primers were used: Girdin, forward
CTC CAG GCA TGA AGC GAACA and reverse 5′-TGG CAG AGC GAG CAT
CCGA-3′; MMP-2, forward 5′-TGC TGA AGG ACA CAC TAAAG-3′ and
reverse, 5′-GTA GCC AAT GAT CCT GTA TGT-3′; MMP-9 forward, GCT ACG
TGA CCT ATG ACA TCCT-3′ and reverse 5′-TCC TCC AGA ACA GAA TAC
CAGT-3′; and β-actin forward CTT AGT TGC GTT ACA CCC TTT CTTG-3′
and reverse 5′-CTG TCA CCT TCA CCG TTC CAG TTT-3′.
Gelatin zymogram
The glioma cells (1×106 cells/ml) from
each treatment group were collected via centrifugation at 112 × g
for 3 min at room temperature, suspended in phosphate-buffered
saline (PBS) containing 1% phenylmethyl sulfonyl fluoride (PMSF;
Beyotime Institute of biotechnology), and homogenized on ice; the
samples were subsequently subjected to liquid nitrogen freeze-thaw
three times. Following centrifugation, the proteins from the
samples were harvested in the supernatant fraction. Following
measurement of the protein concentrations using a BCA Protein
Concentration Detection kit, equal quantities of protein were
separated on an SDS-PAGE gel containing 1 mg/ml gelatin
(Sigma-Aldrich). Following electrophoresis, the gel was washed in
an eluent solution (5% Triton X-100, 50 mM Tris-HCl, 5 mM
CaCl2 and 1 µM ZnCl2; pH 7.6), rinsed
in a rinsing solution (50 mM Tris-HCl, 5 mM CaCl2and 1
µM ZnCl2; pH 7.6), incubated in an incubating
solution (50 mM Tris-HCl, 5 mM CaCl2, 1 µM
ZnCl2, 0.02% Brij, and 0.2 M NaCl), and stained with dye
(0.05% Coomassic brilliant blue G-250, 30% methanol and 10% acetic
acid; Amresco). The stained gel was then decolorized sequentially
in destaining solutions A, B and C (30, 20 and 10% methanol
solutions, and 10, 10 and 5% ethanol solutions, respectively).
Images of the gels were captured using a gel imaging system, and
the optical density values were analyzed.
Statistical analysis
Experimental results are presented as the mean ±
standard deviations. Differences between groups were analyzed using
one-way analysis of variance and Bonferroni's multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Identification of stable girdin
shRNA-transfected cell lines
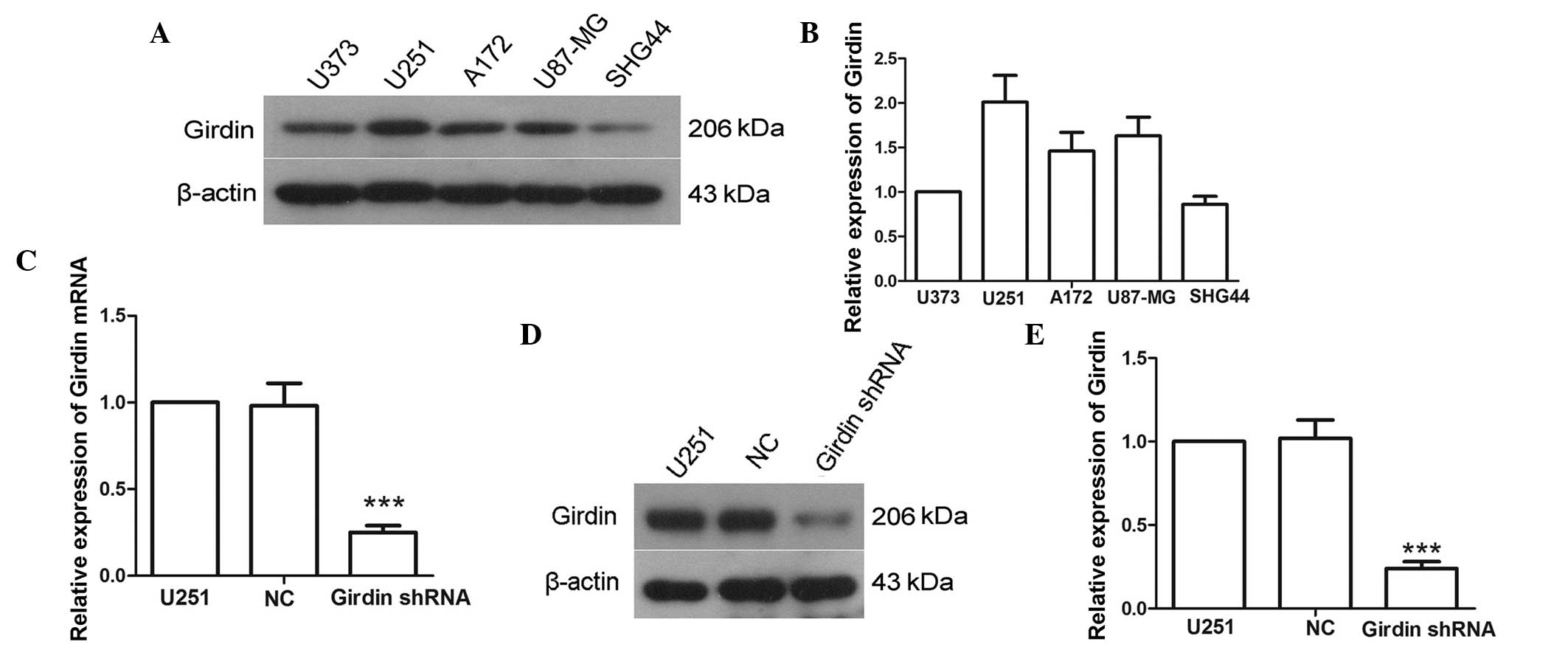
To select the appropriate cell line for the present
study, the expression levels of girdin in U373, U251, A172, U87-MG
and SHG-44 cell lines were assessed using western blot analysis. As
the U251 cell line exhibited the highest expression level of girdin
(Fig. 1A and B), the U251 cells
were selected for use in the subsequent experiments. Following
transfection with girdin shRNA, the expression of girdin was
detected using RT-qPCR and western blot analysis. The RT-qPCR
results indicated that the mRNA levels of girdin were reduced to
25±4% following transfection with girdin shRNA (Fig. 1C). Similarly, the western blot
analysis revealed that the protein expression of girdin was
significantly reduced following transfection with girdin shRNA
(P<0.001; Fig. 1D and E). These
results suggested that the expression of girdin was effectively
silenced following transfection with girdin shRNA.
Girdin silencing inhibits the
proliferation, migration and invasion of glioma cells
To evaluate the effect of girdin silencing on glioma
proliferation, migration and invasion, an MTT assay was used to
determine the changes in cell viability following transfection with
girdin shRNA. The MTT assay results demonstrated that girdin
silencing significantly reduced the proliferation of glioma cells
(P<0.01; Fig. 2A). A
wound-healing assay was subsequently performed to detect changes in
migration capacity of the cells. The results of the wound-healing
assay demonstrated that the relative mobility of the glioma cells
was significantly lower following girdin silencing, compared with
that of the cells transfected with NC (P<0.05; Fig. 2B and C), suggesting that girdin
silencing may impede cell migration. In addition, Transwell
invasion assays were also performed to evaluate the effect of
girdin silencing on glioma cell invasion. The results revealed that
fewer cells passed through the microporous membrane following
transfection with girdin shRNA (P<0.001; Fig. 2D and E), indicating that girdin
silencing significantly reduced glioma cell invasion. The above
results suggested that girdin silencing suppressed glioma cell
proliferation, migration and invasion.
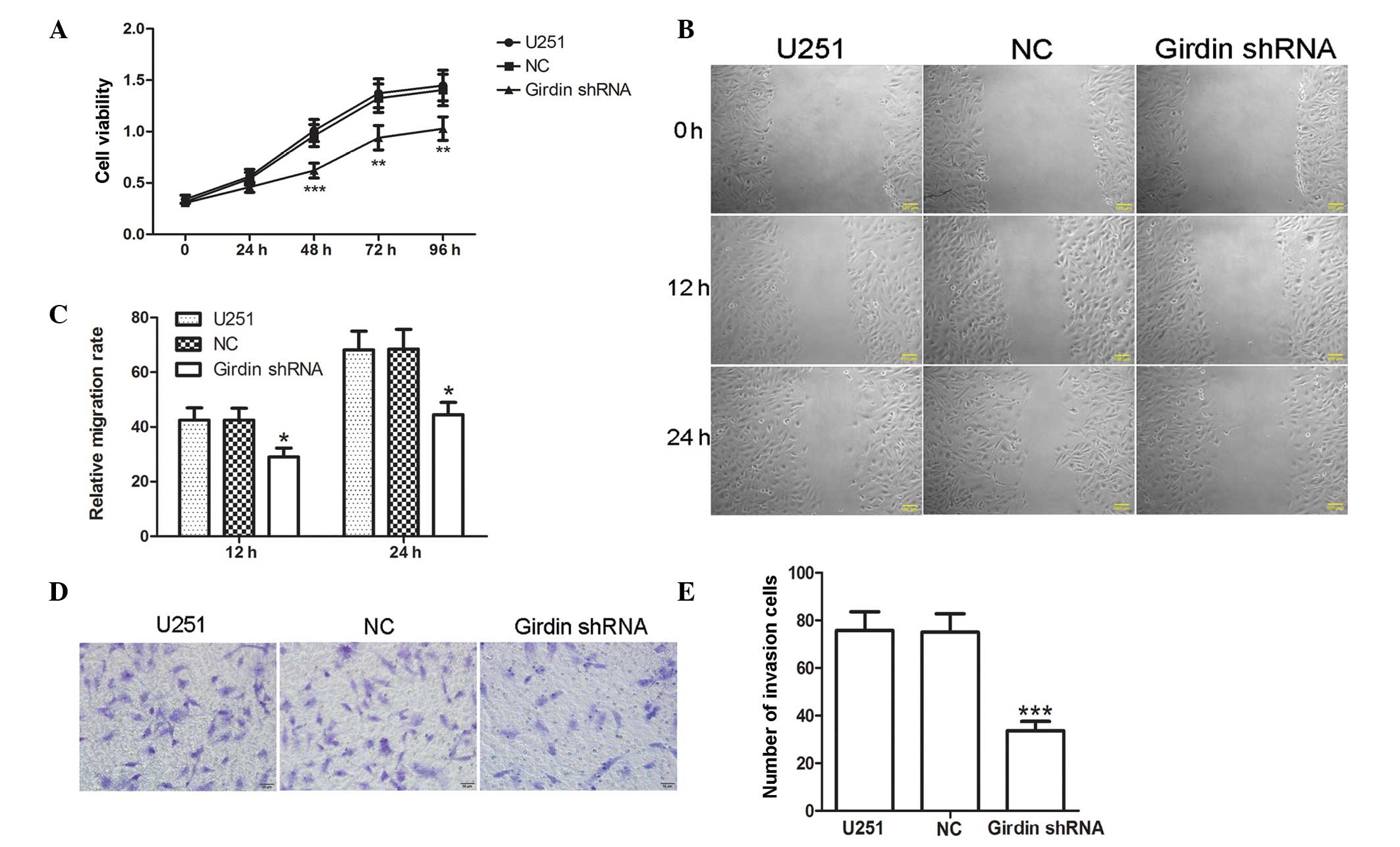 | Figure 2Girdin silencing inhibits the
proliferation, migration and invasion of glioma cells. (A) Changes
of cell viability were detected using a 3-(4,5-dimethy
-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide assay following
transfection. (B and C) Following transfection, a wound-healing
assay was performed to investigate changes in migration capacity
(scale bar, 100 µm). (D and E) A Transwell invasion assay
was performed to detect cell invasion capacity following
transfection. Images were captured under a light microscope
(magnification, ×200). Each experiment was repeated three times.
The experimental results are presented as the means ± standard
deviation. *P<0.05, **P<0.01 and
***P<0.001, compared with the NC group. shRNA, short
hairpin RNA; NC, negative control. |
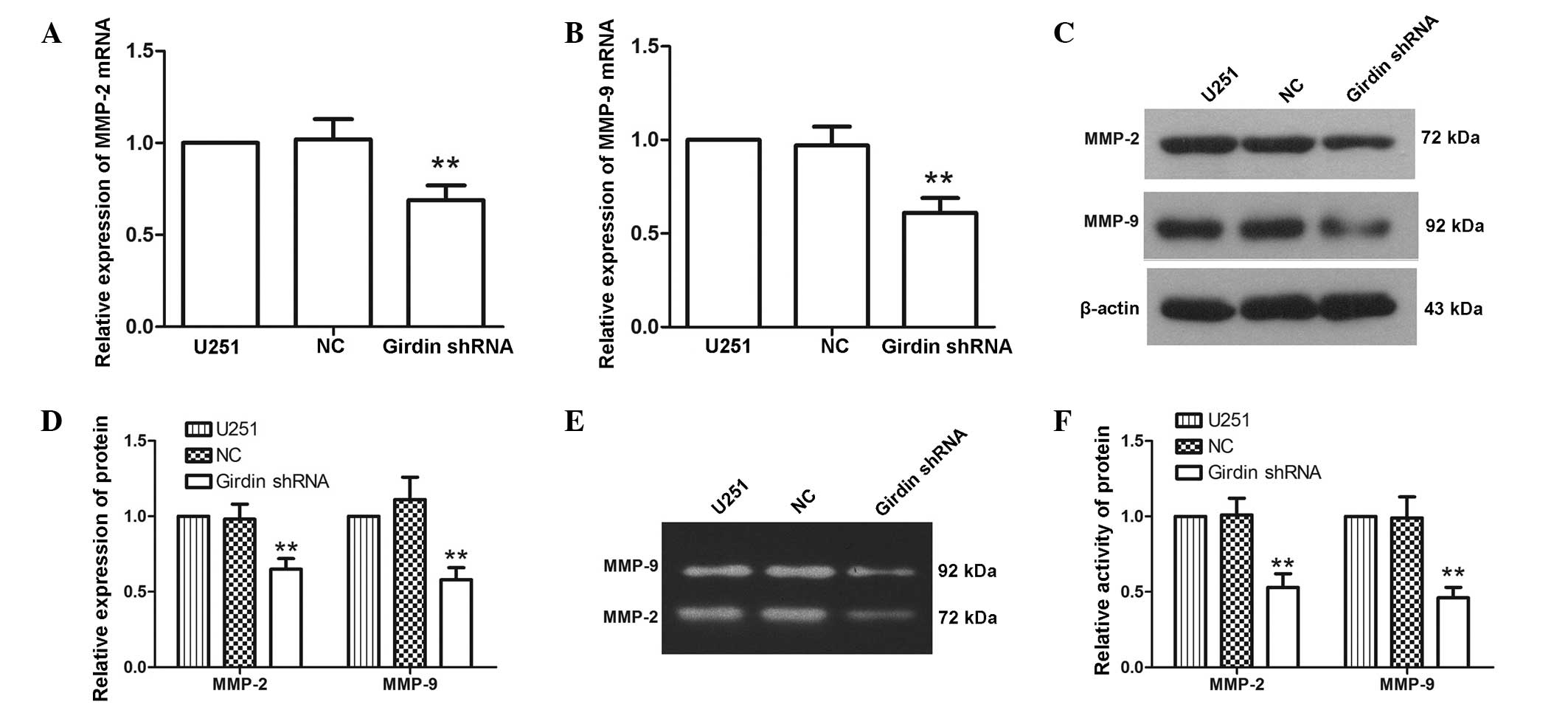
Girdin silencing inhibits the expression
and activity levels of MMP-2 and MMP-9
MMP-2 and MMP-9 are closely associated with cell
migration and invasion. To further investigate the effect of girdin
silencing on glioma cell proliferation, migration and invasion,
RT-qPCR and western blot analyses were performed to detect changes
in the expression levels of MMP-2 and MMP-9 following transfection
with girdin shRNA. The RT-qPCR results demonstrated that the mRNA
levels of MMP-2 and MMP-9 were reduced to 69±8 and 61±8%,
respectively, following transfection with girdin shRNA (Fig. 3A and B). Similarly, western blot
analysis revealed that the protein expression levels of MMP-2 and
MMP-9 were decreased to 65±7 and 58±8% following transfection with
girdin shRNA (Fig. 3C and D),
which was consistent with the results of the RT-qPCR. As the
activities of MMP-2 and MMP-9 are important for their biological
functions, gelatin zymography was used to characterize the
activities of MMP-2 and MMP-9. The results of the gelatin zymogram
revealed that the activities of MMP-2 and MMP-9 were significantly
reduced following transfection with girdin shRNA (P<0.01;
Fig. 3E and F). These results
indicated that girdin silencing inhibited the expression levels and
activities of MMP-2 and MMP-9.
Girdin regulates glioma cell migration
and invasion via the PI3K-Akt signaling pathway
To further investigate how girdin affects glioma
cell migration and invasion, western blotting was used to detect
the protein levels of p85α, p110α, Akt and p-Akt following
transfection with girdin shRNA. The results demonstrated that the
protein levels of p85α, p110α and p-Akt were reduced to 55±9, 58±7
and 59±8%, respectively; however, no significant changes were
detected in the expression of Akt (Fig. 4A and B). These results indicated
that girdin silencing affected the activation of the PI3K-Akt
signaling pathway. The LY294002 PI3K signaling pathway inhibitor
was used to inhibit the PI3K-Akt pathway, followed by western blot
analysis to detect changes in the levels of Akt, p-Akt and MMP-9.
The results revealed that the levels of p-Akt and MMP-9 were
significantly reduced following girdin silencing. In addition, the
LY294002-mediated PI3K-Akt signaling pathway inhibition yielded the
same results as girdin silencing. The combined application of
LY294002 with girdin silencing further enhanced the reductions of
p-Akt and MMP-9 (Fig. 4C and D). A
Transwell invasion assay, which was used to detect the effect of
LY294002 on glioma cell invasion, also demonstrated that LY294002
further enhanced the inhibitory effect of girdin silencing on
glioma cell invasion (Fig. 4E and
F). These results suggested that girdin silencing may affect
glioma cell migration and invasion by regulating the PI3K-Akt
signaling pathway.
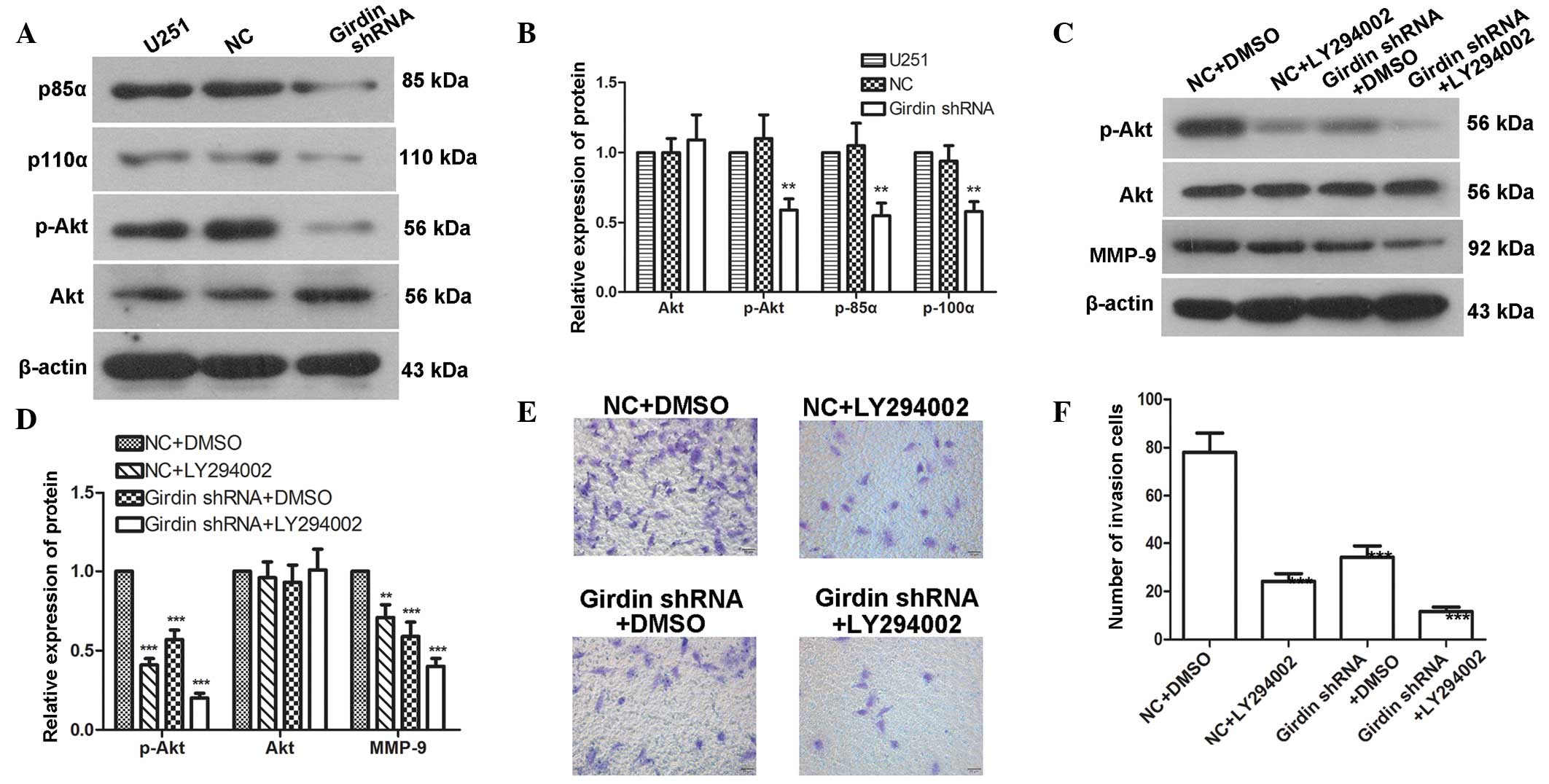 | Figure 4Girdin regulates glioma cell migration
and invasion via the PI3K-Akt signaling pathway. (A and B) Western
blot analysis was performed to detect changes in the levels of
p85α, p110α, p-Akt and Akt following transfection with girdin
shRNA. (C and D) Western blot analysis was performed to detect the
effects of girdin silencing on the protein levels of p-Akt, Akt and
MMP-9 following treatment with LY294002. (E and F) A Transwell
invasion assay was performed to detect the effect of girdin
silencing on cell invasion following treatment with LY294002.
Images were captured under a light microscope (magnification,
×200). Each experiment was repeated three times. The experimental
results are presented as the mean ± standard deviation.
**P<0.01, compared with the negative control. shRNA,
short hairpin RNA; NC, negative control; DMSO, dimethyl sulfoxide;
MMP, matrix metalloproteinase; Akt, protein kinase B; p-,
phosphorylated. |
Discussion
Glioma is a common type of brain tumor and poses a
serious threat to human health. Girdin is expressed at high levels
in glioma cells and is closely associated with glioma development.
The present study focused on the effects of girdin silencing on
glioma cell proliferation, migration and invasion, as well as
investigating the underlying mechanism. The results of the present
study suggested that girdin silencing may affect glioma cell
migration and invasion by regulating the PI3K-Akt signaling
pathway.
Girdin is an actin-binding protein and is activated
by girdin phosphorylation via the PI3K-Akt signaling pathway.
Activated girdin migrates to the pseudopodia at the front edges of
migrating cells. At this location, girdin regulates the front-edge
actin cytoskeletal structure and promotes cyto-skeletal
rearrangement, thereby facilitating cell motility and being
important in tumor invasion and metastasis (7). Girdin is expressed at high levels in
glioma cells, and the expression level of girdin is associated with
tumor metastasis (14). In the
present study, the proliferation, migration and invasion of glioma
cells were significantly reduced following girdin silencing.
Consistent with these findings, Natsume et al (16) reported that genetic girdin knockout
promoted glioma stem cell differentiation, but inhibited cell
motility, invasion, metastasis and proliferation in vivo.
Cao et al (17)
demonstrated that girdin knockout reduced esophageal cancer cell
proliferation, migration and invasion, which was also similar to
the findings of the present study. Girdin deprivation has also been
observed to inhibit vascular smooth muscle cell (VSMC)
proliferation and to affect actin cytoskeletal rearrangement,
resulting the in impaired migration of VSMCs and altered neointimal
formation following vascular injury (10). Therefore, girdin is important in
the processes of tumor cell proliferation, migration and
invasion.
In the present study, girdin silencing inhibited the
expression levels and activities of MMP-2 and MMP-9. MMP-2 and
MMP-9 exerted certain effects on cell migration and invasion, and
inhibition of the expression and activities of MMP-2 and MMP-9 by
girdin silencing demonstrated the regulatory effects of girdin on
glioma cell migration and invasion at the molecular level. Similar
to the findings of the present study, Gu et al (18) reported that girdin silencing
inhibits the in vivo and in vitro expression of MMP-2
and MMP-9, and reduces cell migration and invasion. In addition,
girdin silencing affects the phosphorylation of integrin β1 and
focal adhesion kinase adhesion molecules, suggesting an effect on
cell adhesion (18).
The PI3K-Akt signaling pathway is involved in the
regulation of various cellular processes and is important in tumor
proliferation, invasion and metastasis (19,20).
Studies have revealed that Akt knock down inhibits brain glioma
invasion and metastasis (19,21).
Girdin is an important downstream target of the Akt signaling
pathway. This protein enhances PI3K-Akt signaling pathway activity
and regulates cell proliferation and apoptosis (7). Girdin can also be activated via
phosphorylation by Akt, and can bind and activate Gαi3 to further
activate the PI3K-Akt signaling pathway (22). In the present study, activation of
PI3K-Akt signaling pathway was suppressed by girdin silencing. In
addition, treatment with a PI3K-Akt signaling pathway inhibitor
enhanced the inhibitory effects of girdin silencing on glioma cell
migration and invasion. These results suggested that girdin may
regulate glioma cell migration and invasion through the PI3K-Akt
signaling pathway. Similar to these findings, Lin et al
(23) reported that, in breast
cancer cells, girdin binds to the PI3K regulatory subunit p85α and
promotes the phosphorylation of p85α and activation of the PI3K-Akt
signaling pathway, regulating breast cancer cell migration.
In the present study, shRNA silencing technology was
used to evaluate the effects of girdin on the proliferation,
migration and invasion of glioma cells. The results demonstrated
that girdin silencing decreased the proliferation, migration and
invasion of glioma cells, and subsequent mechanistic investigation
indicated that girdin may regulate glioma cell migration and
invasion via the PI3K-Akt signaling pathway. The results of the
present study provide a theoretical basis for the development of
anti-glioma drugs.
Acknowledgments
This study was supported by grants from the National
Nature Science Foundation of China (grant. no. 81300601), the
Social Development Project of Department of Science and Technology,
Liaoning Province (grant. no. 2013225049) and the Nature Science
Foundation of Liaoning Province (grant. no. 2013022025).
References
|
1
|
Ostrom QT, Gittleman H, Farah P, Ondracek
A, Chen Y, Wolinsky Y, Stroup NE, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol.
15(Suppl 2): ii1–ii56. 2013. View Article : Google Scholar :
|
|
2
|
Stupp R, Brada M, van den Bent MJ, Tonn JC
and Pentheroudakis G: High-grade glioma: ESMO clinical practice
guidelines for diagnosis, treatment and follow-up. Ann Oncol.
25(Suppl 3): iii93–iii101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Tonn JC, Brada M and
Pentheroudakis G: High-grade malignant glioma: ESMO clinical
practice guidelines for diagnosis, treatment and follow-up. Ann
Oncol. 21(Suppl 5): v190–v193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Demuth T and Berens ME: Molecular
mechanisms of glioma cell migration and invasion. J Neurooncol.
70:217–228. 2004. View Article : Google Scholar
|
|
5
|
Mrugala MM, Adair J and Kiem HP:
Temozolomide: Expanding its role in brain cancer. Drugs Today
(Barc). 46:833–846. 2010. View Article : Google Scholar
|
|
6
|
Anai M, Shojima N, Katagiri H, Ogihara T,
Sakoda H, Onishi Y, Ono H, Fujishiro M, Fukushima Y, Horike N, et
al: A novel protein kinase B (PKB)/AKT-binding protein enhances PKB
kinase activity and regulates DNA synthesis. J Biol Chem.
280:18525–18535. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Enomoto A, Murakami H, Asai N, Morone N,
Watanabe T, Kawai K, Murakumo Y, Usukura J, Kaibuchi K, Takahashi
M, et al: Akt/PKB regulates actin organization and cell motility
via Girdin/APE. Dev Cell. 9:389–402. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ohara K, Enomoto A, Kato T, Hashimoto T,
Isotani-Sakakibara M, Asai N, Ishida-Takagishi M, Weng L, Nakayama
M, Watanabe T, et al: Involvement of Girdin in the determination of
cell polarity during cell migration. PLoS One. 7:e366812012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kitamura T, Asai N, Enomoto A, Maeda K,
Kato T, Ishida M, Jiang P, Watanabe T, Usukura J, Kondo T, et al:
Regulation of VEGF-mediated angiogenesis by the Akt/PKB substrate
Girdin. Nat Cell Biol. 10:329–337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miyake H, Maeda K, Asai N, Shibata R,
Ichimiya H, Isotani-Sakakibara M, Yamamura Y, Kato K, Enomoto A,
Takahashi M and Murohara T: The actin-binding protein Girdin and
its Akt-mediated phosphorylation regulate neointima formation after
vascular injury. Circ Res. 108:1170–1179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garcia-Marcos M, Jung BH, Ear J, Cabrera
B, Carethers JM and Ghosh P: Expression of GIV/Girdin, a
metastasis-related protein, predicts patient survival in colon
cancer. FASEB J. 25:590–599. 2011. View Article : Google Scholar :
|
|
12
|
Jiang P, Enomoto A, Jijiwa M, Kato T,
Hasegawa T, Ishida M, Sato T, Asai N, Murakumo Y and Takahashi M:
An actin-binding protein Girdin regulates the motility of breast
cancer cells. Cancer Res. 68:1310–1318. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Garcia-Marcos M, Ear J, Farquhar MG and
Ghosh P: A GDI (AGS3) and a GEF (GIV) regulate autophagy by
balancing G protein activity and growth factor signals. Mol Biol
Cell. 22:673–686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao L, Ma S, Liu Q and Liang P: Clinical
implications of Girdin protein expression in glioma.
ScientificWorldJournal. 2013:9860732013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
16
|
Natsume A, Kato T, Kinjo S, Kato T,
Hasegawa T, Ishida M, Sato T, Asai N, Murakumo Y, Takahashi M and
Wakabayashi T: Girdin maintains the stemness of glioblastoma stem
cells. Oncogene. 31:2715–2724. 2012. View Article : Google Scholar
|
|
17
|
Cao K, Jiang W, Cao P, Zou Q, Xiao S, Zhou
J and Huang C: Talen-mediated girdin knockout downregulates cell
proliferation, migration and invasion in human esophageal carcinoma
ECA109 cells. Mol Med Rep. 10:848–854. 2014.PubMed/NCBI
|
|
18
|
Gu F, Wang L, He J, Liu X, Zhang H, Li W,
Fu L and Ma Y: Girdin, an actin-binding protein, is critical for
migration, adhesion and invasion of human glioblastoma cells. J
Neurochem. 131:457–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang B, Gu F, She C, Guo H, Li W, Niu R,
Fu L, Zhang N and Ma Y: Reduction of Akt2 inhibits migration and
invasion of glioma cells. Int J Cancer. 125:585–595. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu W, Bagaitkar J and Watabe K: Roles of
AKT signal in breast cancer. Front Biosci. 12:4011–4019. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nicholson KM and Anderson NG: The protein
kinase B/Akt signalling pathway in human malignancy. Cell Signal.
14:381–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ghosh P, Garcia-Marcos M, Bornheimer SJ
and Farquhar MG: Activation of Galphai3 triggers cell migration via
regulation of GIV. J Cell Biol. 182:381–393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin C, Ear J, Pavlova Y, Mittal Y,
Kufareva I, Ghassemian M, Abagyan R, Garcia-Marcos M and Ghosh P:
Tyrosine phosphorylation of the Galpha-interacting protein GIV
promotes activation of phosphoinositide 3-kinase during cell
migration. Sci Signal. 4:ra642011.
|