Introduction
When dietary nutrient intake chronically exceeds
energy expenditure, obesity occurs due to an increase in the size
and/or number of white adipocytes (1). Obesity is closely associated with
type 2 diabetes, fatty liver, hyperlipidemia, cardiovascular
diseases and tumorigenesis (2,3).
Thus, investigating the molecular and cellular mechanisms
regulating adipocyte differentiation is crucial for designing
potential therapies to prevent obesity and the associated
diseases.
Decades of studies have identified a number of
transcription factors involved in adipocyte differentiation,
including peroxisome proliferator-activated receptor γ (PPARγ) and
members of the C/EBP family (4,5).
Additionally, a number of other factors or signaling molecules are
involved in the control of adipocyte differentiation, such as
Wnt/β-catenin, cAMP/CREB, and the family of kruppel-like factor
(KLF) proteins (6–8). Furthermore, recent studies have
indicated that modification of these proteins has an important role
in adipogenesis. It has been shown that phosphorylation,
acetylation and small ubiquitin-like modifier (SUMO)ylation of
PPARγ affects adipocyte differentiation, though regulation of its
transcriptional activity (9–11).
In this regard, the process of adipogenesis is rather complex,
suggesting that identification of the additional factors may
provide novel insights into the mechanisms regulating the
initiation of fat cell differentiation and obesity.
In the present study, polymerase chain reaction
(PCR)-based microarrays were performed in obese and lean mice to
screen potential genes involved in adipogenesis.
Materials and methods
Mice
Male C57BL/6, ob/ob and db/db mice,
aged 10–12 weeks, were purchased from the Shanghai Laboratory
Animal Company (Shanghai, China). All mice were housed and bred
according to international standard conditions, with a 12-h
dark/light cycle. For induction of obesity, mice aged 10 weeks were
placed on a high-fat-diet (50 kcal% fat, 25 kcal% protein and 25
kcal% carbohydrate) for 12 weeks. The normal diet consisted of 10
kcal% fat, 15 kcal% protein and 75 kcal% carbohydrate. The animal
protocol was reviewed and approved by the Animal Care Committee of
Chongqing Ninth People's Hospital (Chongqing, China).
Human tissue samples
Human subcutaneous fat tissues from obese
(BMI>25; n=25) and lean (BMI<23; n=20) adults were collected
at the Department of Endocrinology at Chongqing Ninth People's
Hospital. All subjects had a stable weight, defined as the absence
of fluctuations of >1% of body weight for at least 2 months
prior to surgery. Adipose tissue was immediately frozen in liquid
nitrogen following extraction. All samples were obtained with
informed consent and the study was approved by the Chongqing Ninth
People's Hospital Institutional Review Board.
Cell culture
3T3-L1 fibroblasts were obtained from The Cell Bank
of Type Culture Collection of Chinese Academy of Sciences
(Shanghai, China), and cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco-BRL, Carlsbad, CA, USA) supplemented with 10%
fetal bovine serum (FBS; Gibco-BRL), 100 IU/ml penicillin and 100
µg/ml streptomycin (Gibco-BRL). Adipocyte differentiation
was induced by treating the cells at 80–90% confluency with
standard differentiation inducers [100 µM IBMX (Beyotime
Institute of Biotechnology, Nantong, China), 100 nM dexamethasone
(Beyotime Institute of Biotechnology) and 10 µg/ml insulin
(Eli Lilly & Co., Indianapolis, IN, USA)] for 48 h (from day 0
to day 2). Cells were treated with DMEM supplemented with 10
µg/ml insulin and 10% FBS for another 48 h (from day 2 to
day 4). Subsequently, the medium was replaced by DMEM and 10% FBS,
which was changed daily, and the cells were cultured for a further
4–6 days.
RNA extraction, cDNA microarrays, and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR)
Total RNA was isolated from tissues or cells using
the TRIzol reagent (Invitrogen Life Technologies, Shanghai, China),
and reverse transcription was performed using the RT kit from
Promega Corporation (Madison, WI, USA), according to the
manufacturer's instructions. Affymetrix arrays were performed using
Mouse Genome 2.0 chips from Gene Tech Company Limited (Shanghai,
China). In order to quantify the transcripts of the interest genes,
qPCR was performed using a SYBR Green Premix Ex Taq kit (Takara Bio
Inc., Dalian, China) on a Light Cycler 480 (Roche, Basel,
Switzerland) with 5 mg RNA. The following primer sequences form
Bioyare Biotechnology Company (Shanghai, China) were used: Mouse
Grem2, forward 5′-TGTGCTGGTAAAGGTAGCTGA-3′ and reverse
5′-CCACCTCTCTGAGTTGTTGCT-3′; human Grem2, forward
5′-ATCCCCTCGCCTTACAAGGA-3′ and reverse 5′-TCTTGCACCAGTCACTCTTGA-3′;
C/EBPa, forward 5′-CAAGAACAGCAACGAGTACCG-3′ and reverse
5′-GTCACTGGTCAACTCCAGCAC-3′; PPARg, forward
5′-CTCCAAGAATACCAAAGTGCGA-3′ and reverse
5′-GCCTGATGCTTTATCCCCACA-3′; LPL, forward
5′-ATGGATGGACGGTAACGGGAA-3′ and reverse
5′-CCCGATACAACCAGTCTACTACA-3′; PEPCK, forward
5′-ATGAGCCGCAAACTGGGTC-3′ and reverse 5′-AGAGCCGAACTCCACAATCTC-3′;
CD36, forward 5′-AGCATTCAACGCCAGGTTC-3′ and reverse
5′-CGAGTCTGTCAGTTCAATACCAA-3′; aP2, forward
5′-AAGGTGAAGAGCATCATAACCCT-3′ and reverse
5′-TCACGCCTTTCATAACACATTCC-3′; mouse HPRT1, forward
5′-TCAGTCAACGGGGGACATAAA-3′ and reverse
5′-GGGGCTGTACTGCTTAACCAG-3′; human HPRT1, forward
5′-CCTGGCGTCGTGATTAGTGAT-3′ and reverse
5′-AGACGTTCAGTCCTGTCCATAA-3′. The PCR cycling conditions were as
follows: Initial period at 94°C for 3 min, followed by a two-step
PCR program consisting of 95°C for 5 sec and 60°C for 30 sec for 55
cycles. Gene expression was calculated relative to the mRNA
expression of HPRT1, and relative quantitation analysis of gene
expression data was performed using the 2−ΔΔCt
method.
Retrovirus, small interfering (si)RNA and
transfection
Retroviruses containing empty vector or Grem2 cDNA
were constructed by Invitrogen Life Technologies. siRNA oligos
targeting Grem2 or β-catenin were purchased from Gene Pharma
(Shanghai, China) and transfected into 3T3-L1 cells using
Lipofectamine 2000 (Invitrogen Life Technologies), according to the
manufacturer's instructions. Histological analysis was conducted
using Oil Red O staining. Cells were fixed with 4% paraformaldehyde
for 30 min, followed by incubation with Oil Red O (Sigma-Aldrich,
St. Louis, MO, USA) for 45 min.
Western blot analysis
Cells or tissues were harvested and lysed with lysis
buffer (150 mM NaCl, 50 mM Tris-HCl and 1% NP-40; pH 7.5). Proteins
were quantified and separated by 10% SDS-PAGE and transferred to
polyvinylidine difluoride membranes (Amersham Bioscience,
Buckinghamshire, UK). After blocking with 10% non-fat milk in
phosphate-buffered saline, membranes were immunoblotted with the
following rabbit antibodies from Abcam (Cambridge, MA, USA):
Polyclonal anti-Grem2 (ab102563; 1:1,000), monoclonal C/EBPα
(ab40764; 1:1,000), polyclonal PPARγ (ab19481; 1:1,000), monoclonal
P-P38 (ab178867; 1:2,000), polyclonal P-AKT (ab66138; 1:1,000),
monoclonal P-CREB (ab32096; 1:2,000), monoclonal CREB (ab32515;
1:2,000), polyclonal β-catenin (ab6302; 1:1,000), polyclonal lamin
B (ab16048; 1:2,000) and monoclonal GAPDH (ab181602; 1:1,000). The
blots were then incubated with horseradish peroxidase-linked
secondary antibodies (Cell Signaling Technology, Inc. Danvers, MA,
USA). The signals were detected using the SuperSignal West Pico
Chemiluminescent Substrate kit (Pierce, Rockford, IL, USA)
according to manufacturer's instructions.
Statistical analysis
The results are expressed as the mean ± standard
error of the mean and analyzed using Student's t-test or one way
analysis of variance with GraphPad Prism software, version 5.0.1
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
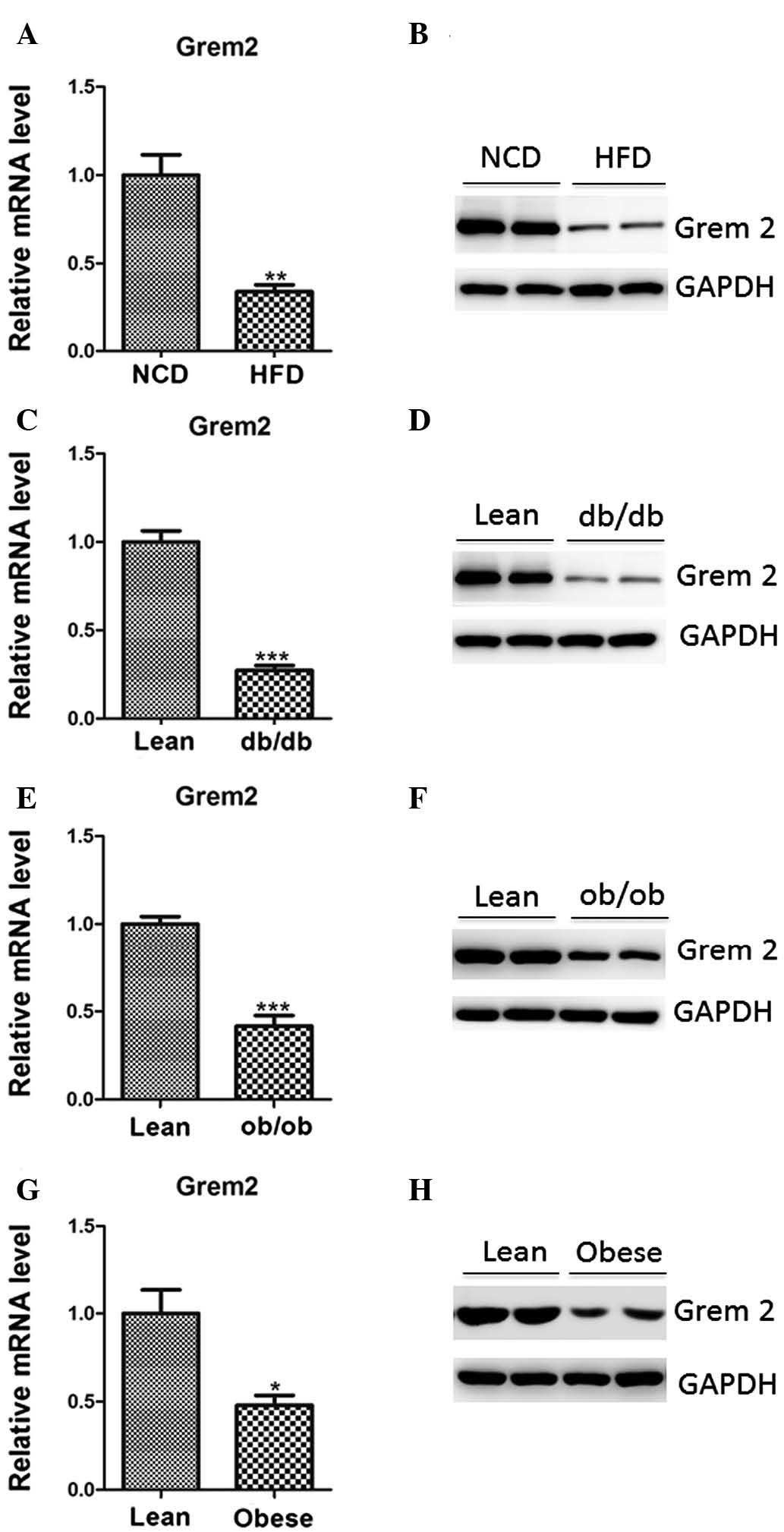
Downregulation of Grem2 in obese mice and
humans
To identify potential genes involved in adipogenesis
and obesity, male C57BL/6 mice aged 10 weeks were placed on
high-fat or normal diets for 12 weeks. The mice were sacrificed by
cervical dislocation and gene expression arrays were performed
using epididymal adipose tissues. The clustering analysis of the
Affymetrix arrays revealed that 1,174 genes were upregulated and
1,572 genes were downregulated in the adipose tissues of mice fed a
high-fat-diet compared with those fed a normal diet (P<0.05;
data not shown). Among these genes, it was determined that Grem2
was significantly downregulated (P<0.001). Its downregulation
was further confirmed by RT-qPCR and western blot analysis
(Fig. 1A and B).
To determine whether the inhibition of Grem2
represents a common feature of obesity, db/db and
ob/ob mice were employed as models for monogenic-induced
obesity. Indeed, Grem2 mRNA and protein levels were found to be
reduced in the epididymal adipose tissues of these mice, as
compared with lean controls (Fig.
1C–F). Notably, Grem2 mRNA and protein levels were
significantly reduced in subcutaneous fat from obese humans
(Fig. 1G and H), further
suggesting that the downregulation of Grem2 expression may be a
conserved feature in obese mice and humans.
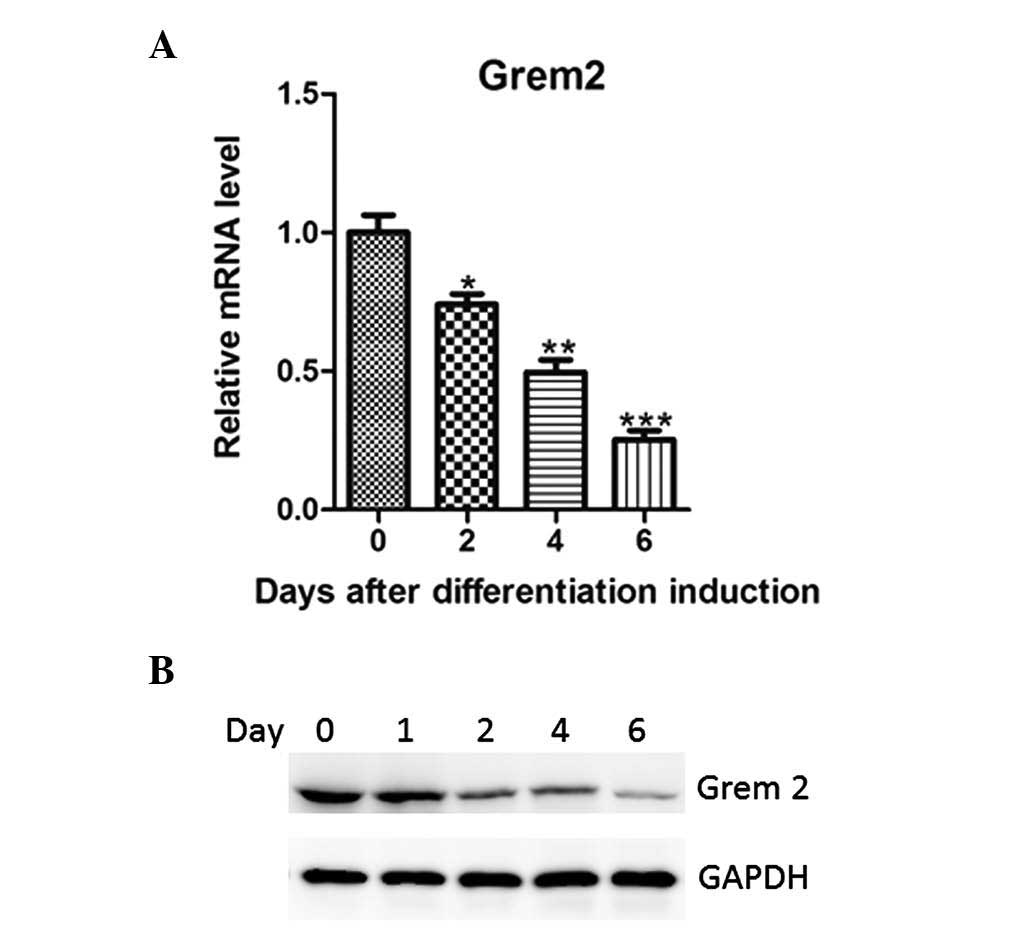
Grem2 suppresses the differentiation of
3T3-L1 preadipocytes into adipocytes
To explore the functions of Grem2 in adipogenesis,
its gene expression was investigated at numerous time points during
3T3-L1 differentiation. The RT-qPCR results revealed that Grem2
mRNA gradually reduced from day 0 post-induction (Fig. 2A). The protein levels for Grem2
were consistent with its mRNA levels (Fig. 2B).
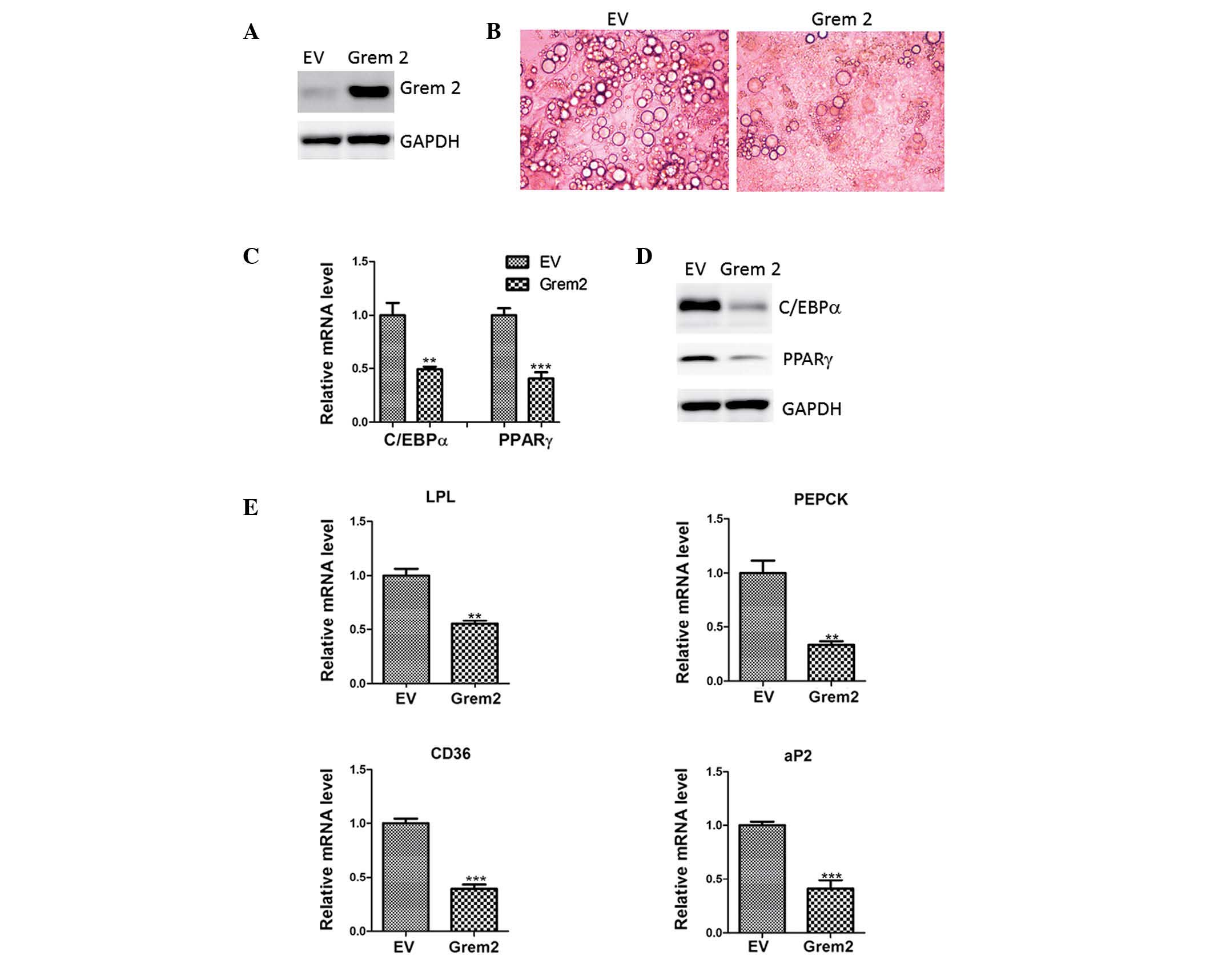
The role of Grem2 in adipogenesis was tested using
retroviruses carrying Grem2 cDNA or empty vector, which resulted in
forced overexpression of Grem2 in 3T3-L1 preadipocytes (Fig. 3A). These cells were cultured to
confluence and differentiated using the standard induction cocktail
for 8 days. Cell morphology indicated that cells overexpressing
Grem2 accumulated significantly less lipids than the control cells
(Fig. 3B). In addition, the
expression levels of adipogenic regulators such as C/EBPα and PPARγ
were reduced (Fig. 3C and D),
which was consistent with the Oil Red O staining. Furthermore,
markers of mature adipocytes LPL, PEPCK, CD36 and aP2, which are
downstream targets of C/EBPα and PPARγ, were repressed by Grem2
overexpression (Fig. 3E).
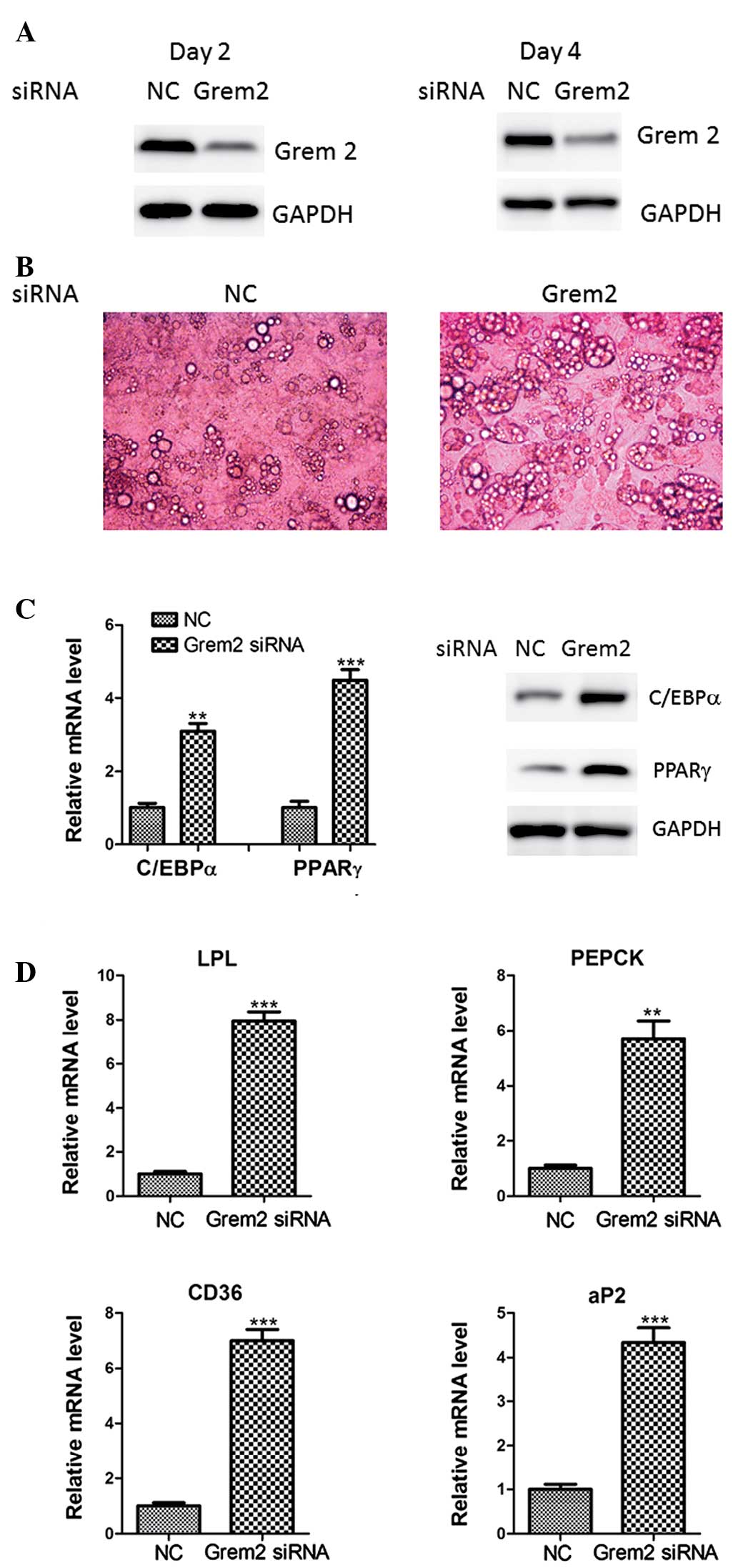
Grem2 knockdown promotes adipocyte
differentiation
To further examine the roles of Grem2, its specific
siRNA was transfected into 3T3-L1 cells (Fig. 4A). As a result, knockdown of Grem2
levels showed a clear increase in the adipogenic potential of the
3T3-L1 cells compared with that of the cells treated with the
negative control siRNA, as illustrated by cell morphology (Fig. 4B). Furthermore, knockdown of Grem2
increased expression levels of the adipocyte markers including
C/EBPα, PPARγ, LPL, PEPCK, CD36 and aP2 (Fig. 4C and D). Together, these results
indicate that Grem2 is a critical negative regulator in the
adipogenesis.
Grem2 activates Wnt/β-catenin signaling
in adipocytes
Finally, the molecular basis for the observed
phenotypic changes in 3T3-L1 cells with Grem2 overexpression was
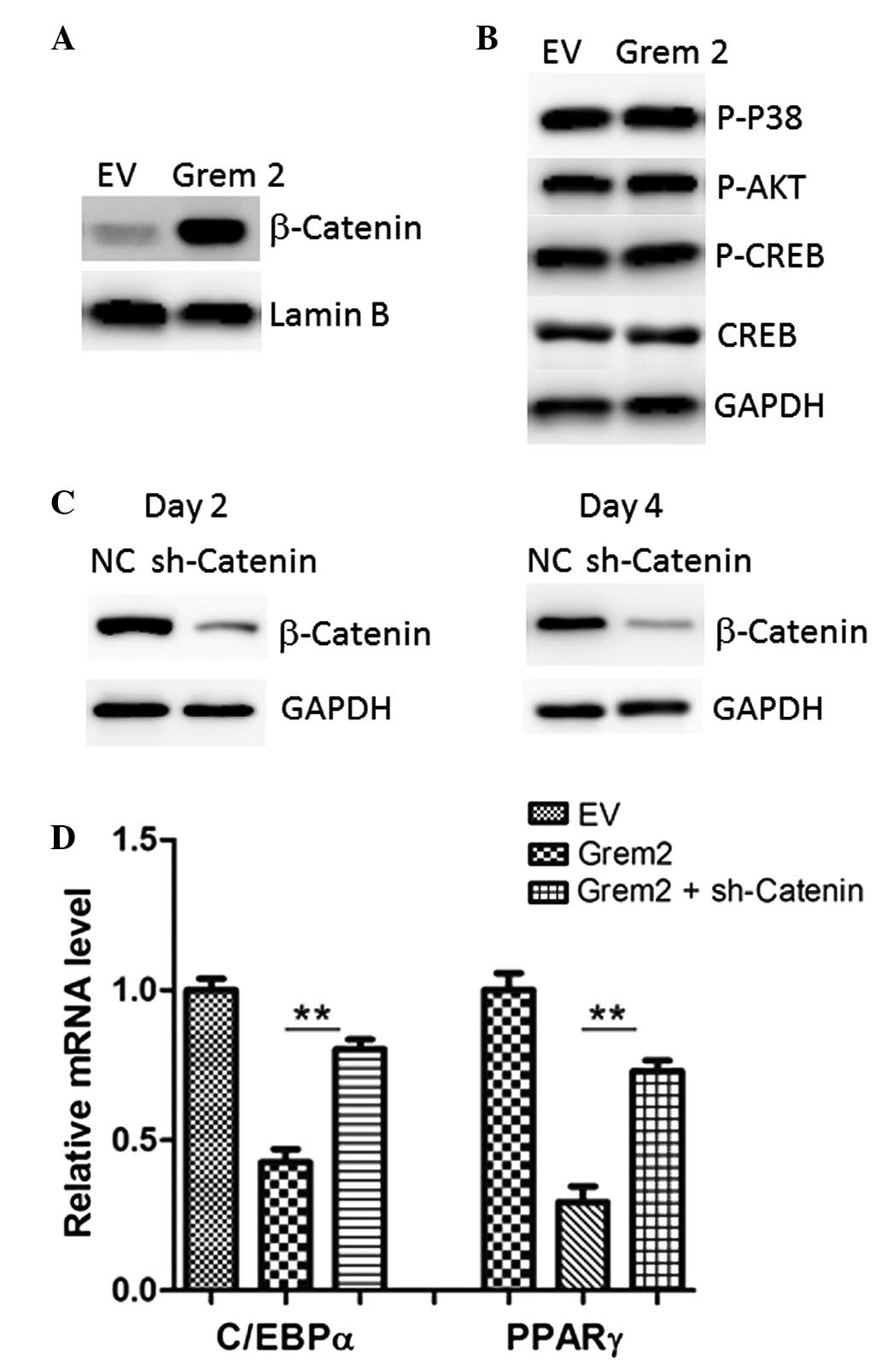
investigated. As shown in Fig. 5A,
Wnt signaling was highly activated by Grem2 overexpression, as
evidenced by nuclear accumulation of β-catenin, while other
signaling molecules, including p38, AKT and cAMP/CREB, remained
unaffected (Fig. 5B).
Subsequently, it was investigated whether ablation of β-catenin
reversed the roles of Grem2. A retrovirus targeting a β-catenin
coding region (sh-catenin) was generated using pAD_BLOCK_IT_DEST
vectors (Invitrogen Life Technologies, Grand Island, NY, USA) to
knockdown endogenous β-catenin expression in 3T3-L1 cells (Fig. 5C). As a result, sh-catenin largely
attenuated the inhibitory roles of Grem2 on the adipogenesis, as
evidenced by gene expression of adipogenic markers (Fig. 5D), suggesting that the role of
Grem2 in the adipogenesis, at least in part, relies on
Wnt/β-catenin activation.
Discussion
In the present study, it was determined that the
Grem2 gene was significantly downregulated in white adipose tissue
from obese mice and humans. Using 3T3-L1 cells cultured in
vitro, it was demonstrated that Grem2 inhibited adipocyte
differentiation. Thus, for the first time to the best of our
knowledge, these results identify Grem2 as a key regulator in
adipose formation and obesity.
Members of the Gremlin family, including Grem2, have
been shown to be bone morphogenetic protein (BMP) antagonists
(12). Indeed, Grem2 regulates BMP
signaling in cardiomyocyte and osteoblast differentiation (13,14).
Furthermore, a recent human study revealed that genetic variants in
Grem2 are associated with bone mineral density (15). Since certain members of the family
of BMPs support white and brown adipocyte differentiation (16,17),
whether Grem2 regulates BMP-induced adipogenesis requires further
investigation.
At the molecular level, it was revealed that Grem2
activates Wnt/β-catenin signaling, as evidenced by its nuclear
accumulation. Initial studies have demonstrated that ectopic
expression of Wnt1, an activator of Wnt/β-catenin signaling, may
inhibit adipogenesis (6,18,19).
Concurrently, pharmacological compounds that activate Wnt/β-catenin
signaling or transfection of stable β-catenin block preadipocyte
differentiation (18). However,
inhibition of Wnt signaling promotes cell differentiation (18,20),
indicating that Wnt signaling has a negative role in the adipocyte
differentiation. In accordance with this conclusion in
vitro, transgenic mice overexpressing Wnt10b, an
endogenous factor for Wnt signaling, have a 50% reduction in
adiposity under a normal chow diet (21). These mice resist expansion of
adipose tissue under conditions of high-fat-diet-induced and
genetic-induced obesity (21).
Furthermore, recent genetic research indicated the importance of
Wnt/β-catenin signaling in human obesity. Polymorphisms of the
Wnt10b gene may be associated with obesity in human subjects of
European origin (22). Together,
these data suggest that modulators of Wnt/β-catenin pathway may
represent a useful therapeutic target for obesity (23).
In conclusion, the results of the current study
highlight a pivotal role for Grem2 in white fat-cell
differentiation by controlling Wnt/β-catenin signaling. Further
studies are required to investigate the functions and mechanisms of
Grem2 in vivo, using knockout or transgenic animals.
References
|
1
|
Rueda-Clausen CF, Padwal RS and Sharma AM:
New pharmacological approaches for obesity management. Nat Rev
Endocrinol. 9:467–478. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fenske W, Athanasiou T, Harling L,
Drechsler C, Darzi A and Ashafian H: Obesity-related cardiorenal
disease: the benefits of bariatric surgery. Nat Rev Nephol.
9:539–551. 2013. View Article : Google Scholar
|
|
3
|
Tao W and Lagergren J: Clinical management
of obese patients with cancer. Nat Rev Clin Oncol. 10:519–533.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rosen ED and MacDougald OA: Adipocyte
differentiation from the inside out. Nat Rev Mol Cell Biol.
7:885–896. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kawai M and Rosen CJ: PPARy: A circadian
transcription factor in adipogenesis and osteogenesis. Nat Rev
Endocrinol. 6:629–636. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cristancho AG and Lazar MA: Forming
functional fat: a growing understanding of adipocyte
differentiation. Nat Rev Mol Cell Biol. 12:722–734. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oishi Y, Manabe I, Tobe K, et al:
Kruppel-like transcription factor klf5 is a key regulator of
adipocyte differentiation. Cell Metab. 1:27–39. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Birsoy K, Chen Z and Friedman J:
Transcriptional regulation of adipogenesis by klf4. Cell Metab.
7:339–347. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choi JH, Banks AS, Estall JL, et al:
Anti-diabetic drugs inhibit obesity-linked phosphorylation of
PPARgamma by cdk5. Nature. 466:451–456. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qiang L, Wang L, Kon N, et al: Brown
remodeling of white adipose tissue by sirt1-dependent deacetylation
of ppargamma. Cell. 150:620–632. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dutchak PA, Katafuchi T, Bookout AL, et
al: Fibroblast growth factor-21 regulates PPARgamma activity and
the antidiabetic actions of thiazolidinediones. Cell. 148:556–567.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zuniga E, Rippen M, Alexander C, Schilling
TF and Crump JG: Gremlin 2 regulates distinct roles of bmp and
endothelin 1 signaling in dorsoventral patterning of the facial
skeleton. Development. 138:5147–5156. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Suzuki D, Yamada A, Aizawa R, et al: bmp2
differentially regulates the expression of gremlin1 and gremlin2,
the negative regulators of bmp function, during osteoblast
differentiation. Calcif Tissue Int. 91:88–96. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Muller II, Melville DB, Tanwar V, et al:
Functional modeling in zebrafish demonstrates that the
atrial-fibrillation-associated gene GREM2 regulates cardiac
laterality, cardiomyocyte differentiation and atrial rhythm. Dis
Models Mech. 6:332–341. 2013. View Article : Google Scholar
|
|
15
|
Paternoster L, Lorentzon M, Lehtimaki T,
et al: Genetic determinants of trabecular and cortical volumetric
bone mineral densities and bone microstructure. PLoS Genet.
9:e10032472013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Macotela Y, Emanuelli B, Mori MA, et al:
Intrinsic differences in adipocyte precursor cells from different
white fat depots. Diabetes. 61:1691–1699. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takahashi A, Morita M, Yokoyama K, Suzuki
T and Yamamoto T: Tob2 inhibits peroxisome proliferator-activated
receptor gamma2 expression by sequestering smads and c/ebpalpha
during adipocyte differentiation. Mol Cell Biol. 32:5067–5077.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ross SE, Hemati N, Longo KA, et al:
Inhibition of adipogenesis by wnt signaling. Science. 289:950–953.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Laudes M: Role of WNT signalling in the
determination of human mesenchymal stem cells into preadipocytes. J
Mol Endocrinol. 46:R65–R72. 2011.PubMed/NCBI
|
|
20
|
Prestwich TC and Macdougald OA:
Wnt/beta-catenin signaling in adipogenesis and metabolism. Curr
Opin Cell Biol. 19:612–617. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wright WS, Longo KA, Dolinsky VW, et al:
WNT10b inhibits obesity in ob/ob and agouti mice. Diabetes.
56:295–303. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chistodoulides C, Scarda A, Granzotto M,
et al: WNT10B mutations in human obesity. Diabetologia. 49:678–684.
2006. View Article : Google Scholar
|
|
23
|
Takada I, Kouzmenko AP and Kato S: Wnt and
PPARgamma signaling in osteoblastogenesis and adipogenesis. Nat Rev
Rheumatol. 5:442–447. 2009. View Article : Google Scholar : PubMed/NCBI
|