Introduction
Lung cancer is at present the number one cause of
cancer-associated mortality of men and women. The 88%-mortality
rate for non-small cell lung cancer (NSCLC) has remained unchanged
since 1985 despite advances in cytotoxic drug development,
radiotherapy and patient management (1). An important step toward deciphering
key intervention points for this disease is a clear understanding
of its genetic pathobiology. Little information exists regarding
the sequence of genetic events leading to the genesis of lung
cancer, particularly for tumors such as adeno-carcinomas, which
occur in the peripheral airways of the lung. The human tumor
suppressor gene differentially expressed in adenocarcinoma of the
lung (DAL-1)/4.1B was identified using differential display
polymerase chain reaction (DDPCR) as a gene whose expression was
lacking in NSCLC when compared with matched normal tissue (2). This gene was determined to be a novel
member of the Protein 4.1 superfamily by virtue of the presence of
a 336 amino-acid N-terminal region with significant homology with
the 4.1/Ezrin/Radixin/Moesin domain present in all 4.1 family
proteins (3). Frequent loss of
4.1B in cervical cancer (4),
laryngeal squamous cell carcinoma (5), breast cancer (6), esophageal squamous cell carcinoma
(7) and lung adenocarcinoma
(8) suggested that 4.1B is a
potential tumor suppressor (9,10).
4.1B is a member of the 4.1-family of proteins together with 4.1R,
4.1N and 4.1G and shows signifi-cant homology with ezrin, radixin
and moesin as well as merlin, which is the specific gene product of
neurofibromatosis type 2. It has also been reported that loss of
4.1B expression and methylation of the 4.1B promoter are involved
in the development and progression of NSCLC, providing a possible
indicator of poor prognosis. Moreover, re-expression of protein
4.1B or a smaller fragment of the entire protein, termed DAL-1,
resulted in growth suppression of meningioma cells (11,12).
The epithelial-mesenchymal transition (EMT) is a
biological phenomenon responsible for the formation of various
tissues and organs during normal metazoan development. Due to the
association of the EMT with the pathogenesis of cancer, the
attention of the scientific community has been directed towards the
search for and identification of effective therapeutic targets to
inhibit EMT-associated phenotypic changes and tumoral progression
(13,14). In order to explore the role of
DAL-1/4.1B in the process of EMT progression, the present study
used RNA interference technology to knockdown DAL-1 expression in
H460 cells and to then determine the effects on cellular
proliferation and invasion. The present study attempted to
elucidate the molecular mechanisms associated with the malignant
progression of NSCLC, which may represent a basis for adjuvant
chemotherapeutic strategies for this disease.
Materials and methods
Cell culture and reagents
The human non-small cell lung cancer cell lines NCI
H460 and A549 cells were obtained from the American Type Culture
Collection (Manassas, VA, USA) were cultured in RPMI-1640 medium
(Gibco-BRL, Invitrogen Life Technologies, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT,
USA) in the absence of antibiotics at 37°C in a humidified
atmosphere containing 5% CO2. The medium of actively
growing cells was replenished with medium containing 10 ng/ml TGF-β
(R&D Systems, Minneapolis, MN, USA).
Plasmid pcDNA3-DAL-1 containing a
full-length DAL-1 coding region (gene ID: 23136)
This plasmid was verified by DNA sequencing. The
control vector pcDNA3 was purchased from Invitrogen Life
Technologies (Grand Island, NY, USA). Lipofectamine® LTX
& Plus Reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA) was used for transfection. To obtain stable transfectants,
cells seeded in 6-well plates were transfected with 2.5
µg/well plasmids using 10 µl/well
Lipofectamine® LTX Reagent and 2.5 µl/well Plus
Reagent. After 6 h incubation in serum and antibiotic free
conditions, the medium was replaced with RPMI-1640 containing 10%
FBS, and the cells were cultured for 48 h prior to submitting to a
2-week selection in medium containing G418 (600 µg/ml).
DAL-1 shRNA and transient transfection
procedure
A commercially available vector, pGPU6/GFP/Neo
(Shanghai Sangon Biotech Co, Shanghai, China), was used to generate
short hairpin (sh)RNA specific for DAL-1. The human DAL-1
gene-coding sequence was obtained from GenBank (http://www.ncbi.nlm.nih.gov/genbank/),
where three sets of shRNA sequences targeting the human DAL-1 gene
are listed: DAL-1-sh710 (5′-GCAGTGCAAAGTGATACTTCT-3′); DAL-1-sh1329
(5′-GCAAGTGGTCTGTTGATATAT-3′) and DAL-1-sh1436
(5′-GCCGGGAGAGTTTGAACAATT-3′). A non-specific shRNA was designed as
a negative control (DAL-1-shNC):
(5′-CACCGTTCTCCGAACGTGTCACGTCAAGAGATTACGTGACACGTTCGGAGAATTTTTTG-3′).
All shRNAs were synthesized by Shanghai ShengGong Biotechnology Co.
(Shanghai, China). These oligonucleotides were synthesized and
sub-cloned into the restriction sites of the vector at 22°C
for 1h. H460 or A549 cells were plated in six-well plates at a
density of 6×104 cells per well and incubated overnight.
Cells were then transfected with DAL-1-shRNAs (2 mg plasmid in 250
ml RPMI-1640 medium) using 5 ml Lipofectamine™ 2000 (Invitrogen
Life Technologies) according to the manufacturer's instructions.
Untransfected H460 cells were included as a blank control group.
The success of the transfection was determined 48 h later by
inverted fluorescence and phase-contrast microscopy (LEICA
DMIL-PH1; Leica Microsystems, Wetzlar, Germany) using three
randomly-selected fields of vision.
Reverse transcription quantitative
(RT-q)PCR assays
Total RNA was isolated from H460 or A549 cells
(1×107 cells per well) using TRIzol reagent (Invitrogen
Life Technologies). cDNA was synthesized using 2xTaq PCR MasterMix
(Tiangen Biotech Co., Ltd., Beijing, China). The 25-µl
reaction mixture was composed of 1 µl cDNA, 2 µl of
the upstream and downstream primers (MBI Fermentas, Vilnius,
Lithuania), 12.5 µl PCR-2X master mix (Tiangen Biotech Co.,
Ltd.) and 9.5 µl RNase-free water (Amresco LLC, Solon, OH,
USA). PCR was performed according to the manufacturer's
instructions. PCR was performed using the ABI 7500 real time system
(Applied Biosystems Life Technologies, Foster City, CA, USA) using
SYBR Green® Premix Ex Taq (Takara Bio Inc., Tokyo,
Japan). The cycling conditions included a holding step at
94°C for 3 min and 29 cycles of 94°C for 30 sec,
55°C for 30 sec and 72°C for 1 min. β-actin was used
as internal control. Relative quantification was performed using
the ΔΔCT method. Primer sequences of the target genes are listed in
Table I.
 | Table IPrimers used in quantitative
polymerase chain reaction. |
Table I
Primers used in quantitative
polymerase chain reaction.
| Target gene | Forward primer | Reverse primer |
|---|
| E-cadherin |
5′-CAATGCCGCCATCGCTTAC-3′ |
5′-ATGACTCCTGTGTTCCTGTTAATG-3′ |
| Vimentin |
5′-GAGAACTTTGCCGTTGAAGC-3′ |
5′-TCCAGCAGCTTCCTGTAGGT-3′ |
| Snail1 |
5′-GCTCCACAAGCACCAAGAGT-3′ |
5′-ATTCCATGGCAGTGAGAAGG-3′ |
| Snail2 |
5′-CTTTTTCTTGCCCTCACTGC-3′ |
5′-ACAGCAGCCAGATTCCTCAT-3′ |
| Twist1 |
5′-TGCATGCATTCTCAAGAGGT-3′ |
5′-GTTTTGCAGGCCAGTTTGAT-3′ |
| Fibronectin |
5′-ACCAACCTACGGATGACTCG-3′ |
5′-GCTCATCATCTGGCCATTTT-3′ |
| β-catenin |
5′-GTACGTCCATGGGTGGGACA-3′ |
5′-GGCTCCGGTACAACCTTCAACTA-3′ |
| DAL-1 |
5′-GAGCTGCCAAGCGTTTATGGA-3′ |
5′-CCTGCCACTATAACGAAACTTGGAA-3′ |
| β-actin |
5′-AGGTCGGAGTCAACGGATTTGGTCG-3′ |
5′-TGGCCAGGGGTGCTAAGCAGT-3 |
Antibodies and western blot analysis
The following antibodies were used in the present
study: Mouse monoclonal anti-4.1B (1:200; cat. no. 514386; Santa
Cruz Biotechnology, Dallas, TX, USA.); mouse monoclonal
anti-vimentin (1:1,000; cat. no. 3390; Cell Signaling Technology,
Inc., Danvers, MA, USA); mouse monoclonal anti-β-catenin (1:1,000;
cat. no. 2698; Cell Signaling Technology, Inc.); mouse monoclonal
anti-Snail (1:1,000; cat. no. 3895; Cell Signaling Technology,
Inc.); mouse monoclonal anti-fibronectin (1:1,000; cat. no. F7387;
Sigma-Aldrich, St Louis, MO, USA); mouse monoclonal anti-β-actin
(1:10,000; cat. no. A5441; Sigma-Aldrich).
Cells (1×107 cells per well) were lysed
in pre-cooled lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 1 mM
phenylmethylsulphonyl fluoride, 1 mM EDTA, 1% NP-40, 1% sodium
deoxycholate and 0.1% SDS, pH 7.4) and the protein content of the
lysates was assessed using a Bicinchoninic Protein Assay kit
(Beyotime Institute of Biotechnology, Shanghai, China). Briefly,
cells were lysed using lysis buffer [20 mM Tris/HCl, pH 7.5, 137 mM
NaCl, 1% (v/v) Triton X-100, protease inhibitor cocktail
(Complete™, EDTA-free; Roche Applied Science Indianapolis, IN, USA)
and a phosphatase inhibitor cocktail (PhosSTOP; Roche Applied
Science)] and sonicated on ice. Bradford protein assay was used to
determine the protein concentration of lysates (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Lysates were heated at
70°C with lithium dodecyl sulfate sample buffer (Invitrogen
Life Technologies) and dithiothreitol (Sigma-Aldrich) for 10 min.
Equal amounts of cell lysate (20 mg) were separated by 10% SDS-PAGE
and were electroblotted onto polyvinylidene fluoride membranes
(Millipore, Billerica, MA, USA). Blotted membranes were blocked
with 5% skimmed milk in Tris-buffered saline containing Tween
(TBST; pH 7.6) at room temperature for 1 h. Blots were then
incubated overnight at 4°C with the primary antibodies
followed by incubation for 1 h at room temperature with a 1:1,000
dilution of horseradish peroxidase-conjugated goat anti-mouse
monoclonal antibody (Santa Cruz Biotechnology). After incubation,
membranes were washed three times each for 15 min using 0.05% TBST.
Protein bands were visualized using a diaminobenzidine coloration
kit and the Fusion FX7 electrochemiluminescence analysis system
(Wuhan Boster Biological Technology, Wuhan, China) according to the
manufacturer's instructions. Relative band intensities were
detected using ImageJ 1.48 software (National Institutes of Health,
Bethesda, MD, USA). The density of each band was normalized against
that of β-actin.
Cell counting kit-8 (CCK8) assay for
assessment of cell proliferation
Exponentially growing H460 cells were seeded into
96-well microtiter plates at a concentration of 5×104
cells per well and allowed to attach overnight prior to DAL-1 shRNA
transfection. Following 24, 48, 72 and 96 h of culture, cells were
incubated with 20 µl CCK8 (5 mg/ml; Sigma-Aldrich) at
37°C for 4 h after which the medium was decanted and the
reaction stopped by addition of 150 ml dimethylsulfoxide
(Sigma-Aldrich). The spectrometric absorbance of each sample was
measured at 490 nm using an automatic micro-plate reader (iMark™
microplate absorbance reader; Bio-Rad Laboratories, Inc.).
Wound-healing assays
Cells were seeded into six-well plates and cultured
until >90% confluent. Three straight wounds were scratched in
each well using a sterile 200-µl pipette tip. Cells were
rinsed gently with phosphate-buffered saline (PBS) and 2 ml media
(without FBS or containing 10% FBS) was added. Images were captured
at 40× magnification immediately after scratching and again after
24 h and 48 h. All assays were performed in triplicate and all
experiments repeated three times.
Matrigel™ invasion assay
Transwell chambers with 8-mm pore size polycarbonate
membranes (Corning-Costar, Corning, NY, USA) were used to perform
the cell invasion assay. Following 24 h of transfection,
2×104 cells were suspended in 200 µl serum-free
medium and inoculated into each extracellular matrix-coated upper
compartment of the 24-well plates that were pre-coated with 50
µl 1 µg/µl Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA). The lower compartment of each chamber was filled
with 10,000 µl RPMI-1640 with 30% FBS and incubated for 48 h
at 37°C with 5% CO2. The cells on the upper
surface were then removed using cotton tips and the cells that had
migrated to the lower side of the membrane were fixed with methanol
for 30 min, stained with 0.1% crystal violet (Tianjin Yixin Hengxin
Chemical Co., Ltd., Tianjin, China) for 30 min and washed with PBS
three times. The number of invaded tumor cells was calculated in
five random fields at a magnification of ×200, using an inverted
microscope (DP70; Olympus, Tokyo, Japan) and expressed as the
average number of cells/field of view.
Statistical analysis
Each experiment was performed in triplicate.
Statistical analyses were performed using SPSS statistical
software, version 19.0 (SPSS, Inc. Chicago, IL, USA) for Windows.
Values are expressed as the mean ± standard deviation. Analysis of
variance (ANOVA) experiments (one-way, factorial and
repeated-measures ANOVA), followed by the Student-Newman-Keuls test
were performed, with P<0.05 considered to indicate a
statistically significant difference between values.
Results
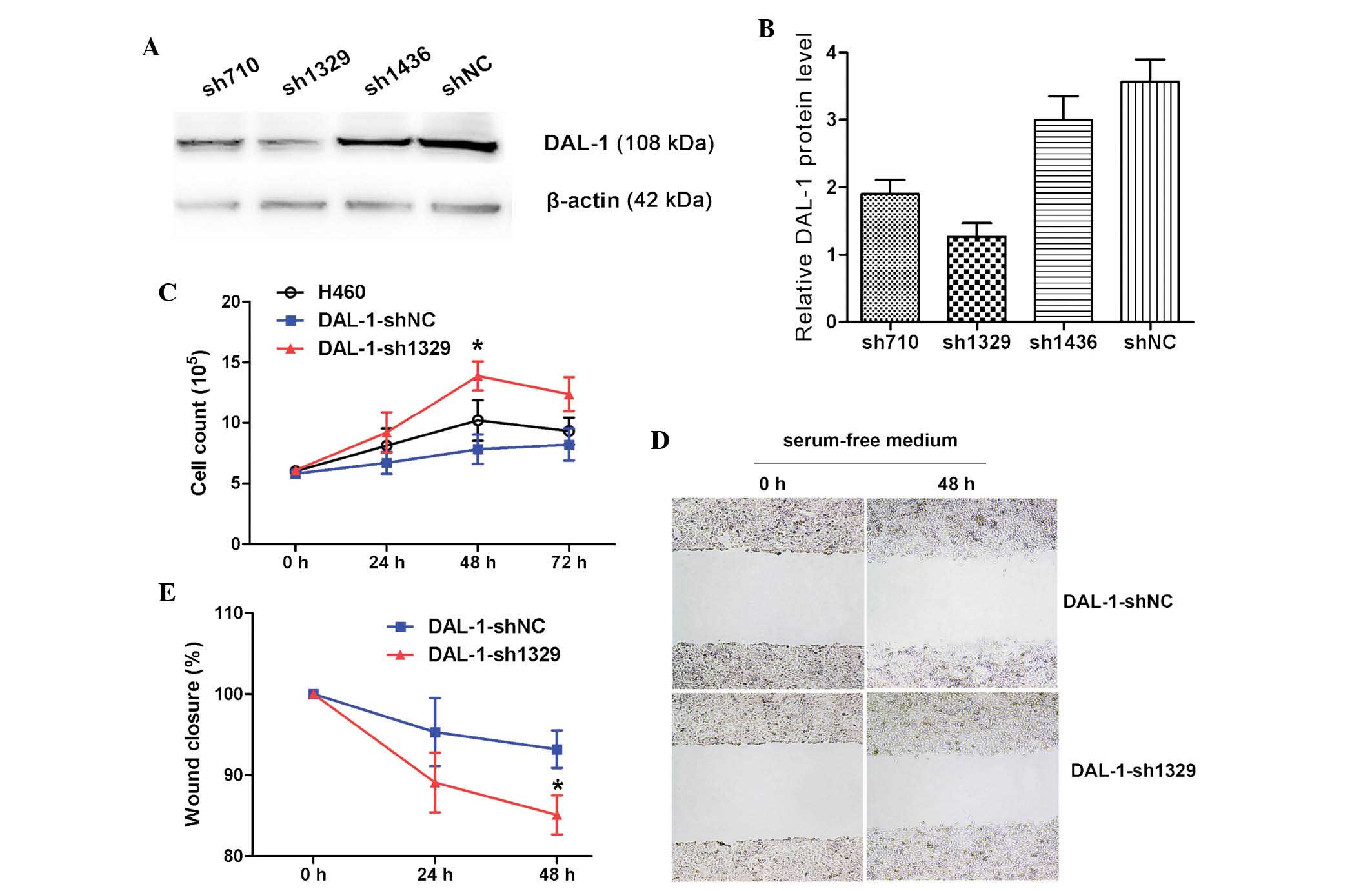
Downregulation of DAL-1/4.1B expression
effectively suppresses DAL-1/4.1B protein expression in lung cancer
cells
To examine the possible roles of DAL-1/4.1B in lung
cancer cells, DAL-1/4.1B was knocked down using small interfering
(si)RNA. Following transfection with DAL-1/4.1B siRNA, the levels
of DAL-1/4.1B mRNA were reduced compared with those of the blank
control group. At 48 h after transfection, DAL-1/4.1B mRNA
expression was effectively knocked down by sh1329, while sh710 and
sh1436 had no obvious effect (Fig.
1A). In addition, western blot analysis demonstrated that
DAL-1/4.1B shRNA exerted a silencing effect on DAL-1/4.1B
expression in vitro (Fig.
1B). These results confirmed that sh1329 effectively interfered
with DAL-1/4.1B expression.
DAL-1/4.1B shRNA promotes cell
proliferation
Compared with untransfected H460 cells (blank
control group), the cell proliferation significantly increased in
cells transfected with DAL-1sh1329 (Fig. 1C) (P<0.05 at 48 h). There were
no significant differences in cell proliferation between the
control groups (P>0.05; blank controls and shNC-transfected
cells).
Downregulation of DAL-1/4.1B decreases
the migration of lung cancer cells in vitro
The role of DAL-1/4.1B in cell migration and
invasion was evaluated by a wound-healing assay and the
Matrigel-based Transwell invasion assay. Cell migration is a
critical step in metastasis, and the results demonstrated that
DAL-1/4.1B has a critical role in the metastasic behavior of cancer
cells. The effects of DAL-1/4.1B shRNA on the characteristics of
lung cancer cells were examined. Transfection of DAL-1/4.1B shRNA
into H460 cells resulted in increased migration capacity compared
with that of the blank control, as was evident from the migration
assay (Fig. 1D and E). Culture
media without FBS was used to exclude the contribution of cell
proliferation in the determination of migration of H460 cells. The
average number of migrated cells in the experimental group
(DAL-1-sh1329) was significantly lower than that in the blank
control group (DAL-1-shNC) (P<0.05).
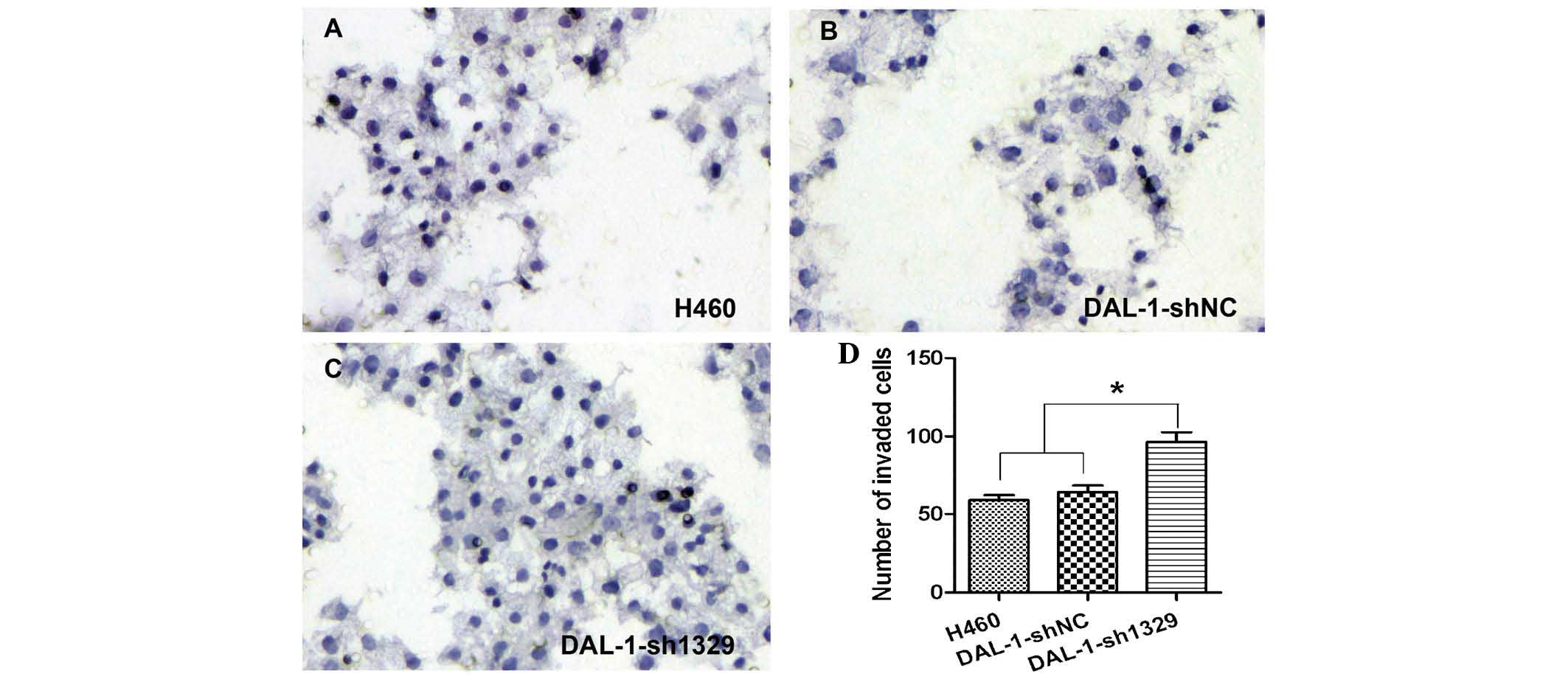
The numbers of cells which transgressed the
Matrigel™ and adhered to the lower side of the membrane in the
blank control (Fig. 2A),
DAL-1-shNC (Fig. 2B) and
DAL-1-sh1329 (Fig. 2C) groups were
59.5±6.52, 64.0±8.16, 96.67±10.33, respectively. An increase in the
number of invaded cells in the DAL-1-sh1329 group was observed
compared with that in the control groups (Fig. 2D; P<0.05), while there was no
significant difference in the invasive potential between the two
control groups.
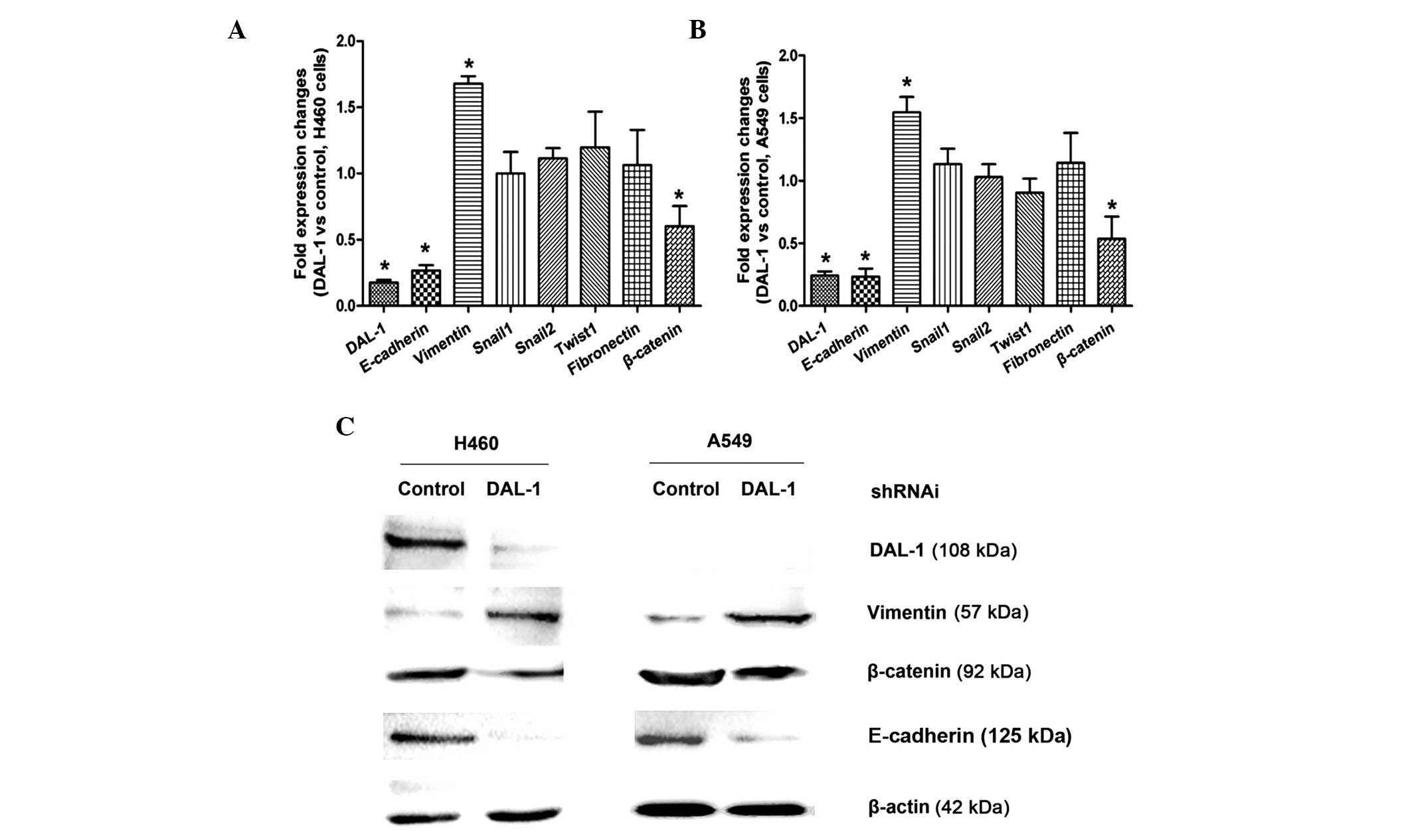
DAL-1/4.1B deficiency alters the
expression of EMT markers
To address the role of DAL-1/4.1B in H460 and A549
non-small cell lung cancer cell lines, control and
DAL-1/4.1B-knockdown cells were generated by transfection with
scrambled shRNA as a control and shRNA targeting DAL-1/4.1B.
RT-qPCR and western blot analysis were used to examine the
expression of EMT markers in control and DAL-1-knockdown H460 and
A549 cells. As shown in Fig. 3A and
B, E-cadherin and β-catenin mRNA were decreased, while vimentin
was increased in DAL-1/4.1B-knockdown cells (P<0.05). However,
changes in the expression of fibronectin, Snail and Twist1
following DAL-1/4.1B knockdown were not significant (P>0.05).
Western blot analysis confirmed that DAL-1/4.1B knockdown resulted
in a decrease in E-cadherin and β-catenin expression and an
increase in vimentin expression at the protein level (Fig. 3C).
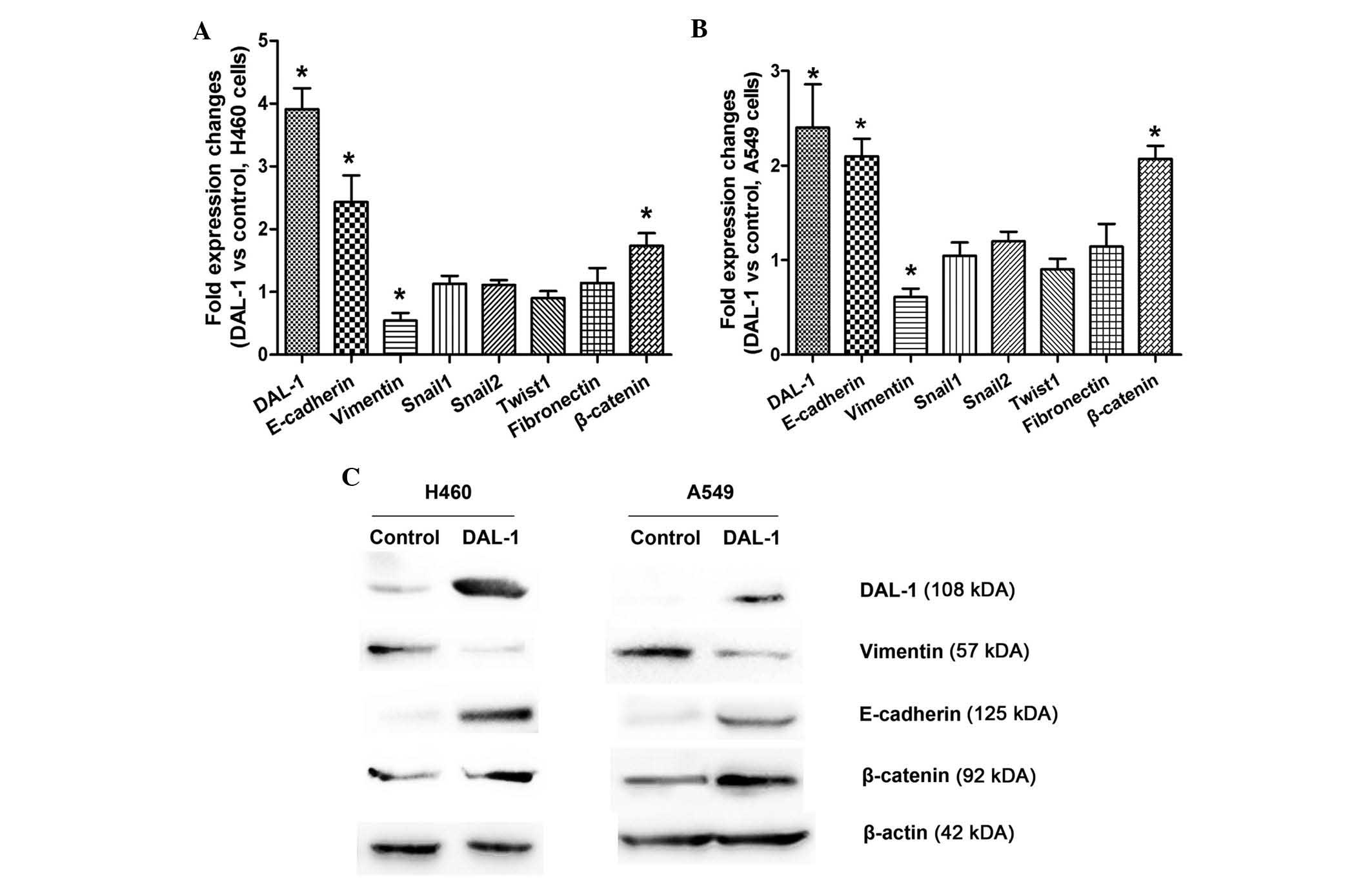
DAL-1/4.1B overexpression alters the
expression of EMT markers
To assess the effect of DAL-1/4.1B overexpression on
EMT marker expression, A549 and H460 cells stably expressing DAL-1
were generated along with control cells transfected with empty
vector. As expected, DAL-1 overexpression resulted in increased
E-cadherin and β-catenin expression levels, whereas vimentin levels
were reduced at the mRNA as well as at the protein level (Fig. 4). Collectively, these results
suggested that DAL-1 is involved in the EMT process in lung cancer
cell lines.
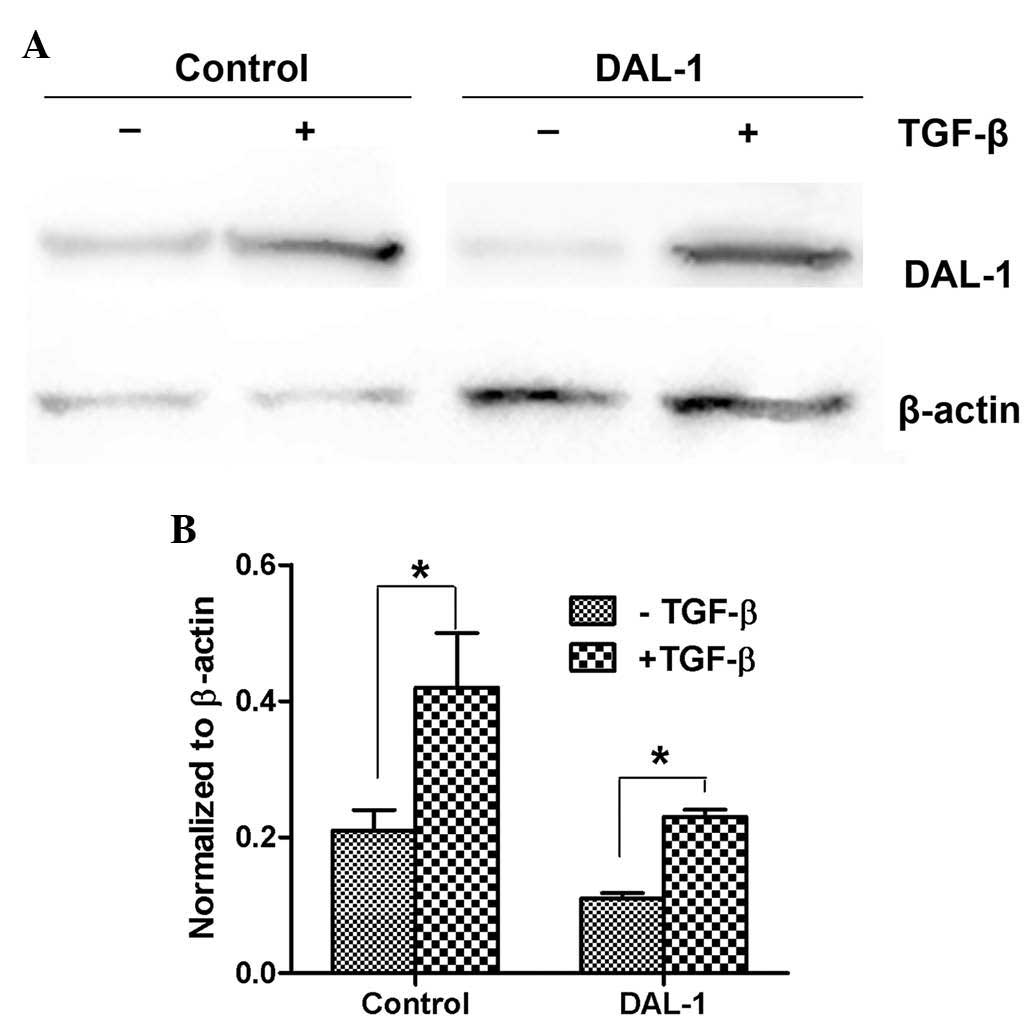
DAL-1/4.1B expression is induced by
transforming growth factor (TGF)-β
It is known that TGF-β signaling is increased in a
variety of cancer types, including lung cancer (15). To determine whether TGF-β and
DAL-1/4.1B are associated in lung cancer, the non-small cell lung
cancer cell line H460, which is known to undergo EMT upon TGF-β
treatment, was utilized as a model. DAL-1/4.1B-knockdown H460 cells
and scrambled control-transfected cells were incubated with TGF-β.
As shown in Fig. 5A, DAL-1/4.1B
protein levels were increased upon TGF-β treatment in H460 cells.
Although the basal levels of DAL-1/4.1B were markedly low in
DAL-1/4.1B-knockdown cells, its expression was induced following
incubation with TGF-β. RT-qPCR showed a similar effect on the mRNA
levels of DAL-1/4.1B (Fig.
5B).
Discussion
Lung cancer is one of the most common malignant
tumor types, and their morbidity and mortality have increased in
recent years. Lung cancer has the highest mortality rate in males,
and the second highest mortality rate in females amongst all tumor
types. According to a recent study (16), the global number of newly diagnosed
lung cancer cases was as high as 120 million people per year, and
100 million people succumbed to this disease. Lung cancer has
become one of the most significant threats to human health.
Although the early diagnosis and treatment of non-small cell lung
cancer have significantly improved in recent years, the five-year
survival rate remains at 10–20% only (17).
The tumor suppressor gene DAL-1/4.1B was shown to be
located on chromosomal fragment 18p11.3 and was inactivated in
certain types of tumor (18,19).
The protein it encodes, 4.1B, belongs to the protein 4.1
superfamily of scaffold proteins. DAL-1/4.1B is normally expressed
at high levels in the brain, while its expression is low in the
kidneys, intestine and testes. As other 4.1 family member proteins,
DAL-1/4.1B has been identified to be localized in regions of the
plasma membrane at points of cell-to-cell contact by
immunocytochemistry (20).
A previous study suggested that aberrant 4.1B
expression is involved in progression of breast cancer,
particularly in invasion into the stroma and metastasis (3). The 4.1B protein is downregulated in
several carcinoma types, including prostate cancer (21). Cavanna et al (22) identified a sub-set of genes with
significantly altered expression levels between non-metastasizing
and metastasizing cells in tissue culture and in primary tumors.
Cells with reduced 4.1B expression displayed an altered F-actin
morphology, with significantly fewer stress fibres.
DAL-1/4.1B-knockdown cells migrated at twice the speed of the
untreated cells. Examination of the expression of the 4.1B protein
in human intestinal mucosa showed that DAL-1 was also expressed in
matured epithelial cells in human colons, with a definite
expression gradient along the crypt axis (23).
The EMT is characterized by a loss of cell-cell
adhesion and polarity, downregulation of epithelial markers, as
well as acquisition of mesenchymal markers and phenotype (24). E-cadherin, another tumor suppressor
of the transmembrane adhesion molecule, also interacts with the
cytoskeleton and is involved in the invasion or metastasis of
gastric cancer as well as several other cancer types (25,26).
Accumulating evidence from studies on the EMT have indicated the
involvement of numerous signaling pathways, including TGF-β, Notch,
Wnt, epidermal growth factor and fibroblast growth factor (27,28).
Among these, TGF-β efficiently induces EMT in a variety of model
cell lines and in vivo (29,30).
In conclusion, the present study showed that shRNA
targeting DAL-1/4.1B significantly downregulated DAL-1/4.1B mRNA
and protein expression in lung cancer cells, and inhibited cell
proliferation as well as migratory and invasive potential. The
results also indicated that downregulation of DAL-1/4.1B decreased
the expression of E-cadherin and β-catenin in H460 and A549 cells.
The findings of the present study provided novel insight into the
underlying molecular mechanisms of NSCLC associated with the EMT,
indicating that the tumor suppressor gene DAL-1/4.1B may be a
potential target for anti-tumour drugs as well as gene therapy for
treating lung cancer.
Acknowledgments
This study was supported by a scientific research
project fund of Jiangxi Provincial Education Department (no.
GJJ11374).
References
|
1
|
Sculier JP, Berghmans T and Meert AP:
Small cell lung cancer: What are the treatment results in routine
management? Lung Cancer. 84:101–102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tran YK, Bogler O, Gorse KM, Wieland I,
Green MR and Newsham IF: A novel member of the NF2/ERM/4.1
superfamily with growth suppressing properties in lung cancer.
Cancer Res. 59:35–43. 1999.PubMed/NCBI
|
|
3
|
Takakuwa Y: Protein 4.1, a multifunctional
protein of the erythrocyte membrane skeleton: Structure and
functions in erythrocytes and nonerythroid cells. Int J Hematol.
72:298–309. 2000.
|
|
4
|
Steenbergen RD, Kramer D, Braakhuis BJ,
Stern PL, Verheijen RH, Meijer CJ and Snijders PJ: TSLC1 gene
silencing in cervical cancer cell lines and cervical neoplasia. J
Natl Cancer Inst. 96:294–305. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu B, Di W, Wang H, Ma H, Li J and Zhang
Q: Tumor suppressor TSLC1 is implicated in cell proliferation,
invasion and apoptosis in laryngeal squamous cell carcinoma by
regulating Akt signaling pathway. Tumour Biol. 33:2007–2017. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Heller G, Geradts J, Ziegler B, Newsham I,
Filipits M, Markis-Ritzinger EM, Kandioler D, Berger W, Stiglbauer
W, Depisch D, et al: Downregulation of TSLC1 and DAL-1 expression
occurs frequently in breast cancer. Breast Cancer Res Treat.
103:283–291. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ito T, Shimada Y, Hashimoto Y, Kaganoi J,
Kan T, Watanabe G, Murakami Y and Imamura M: Involvement of TSLC1
in progression of esophageal squamous cell carcinoma. Cancer Res.
63:6320–6326. 2003.PubMed/NCBI
|
|
8
|
Mao X, Seidlitz E, Truant R, Hitt M and
Ghosh HP: Re-expression of TSLC1 in a non-small-cell lung cancer
cell line induces apoptosis and inhibits tumor growth. Oncogene.
23:5632–5642. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kikuchi S, Yamada D, Fukami T, Masuda M,
Sakurai-Yageta M, Williams YN, Maruyama T, Asamura H, Matsuno Y,
Onizuka M and Murakami Y: Promoter methylation of DAL-1/4.1B
predicts poor prognosis in non-small cell lung cancer. Clin Cancer
Res. 11:2954–2961. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goto A, Niki T, Chi-Pin L, Matsubara D,
Murakami Y, Funata N and Fukayama M: Loss of TSLC1 expression in
lung adenocarcinoma: Relationships with histological subtypes, sex
and prognostic significance. Cancer Sci. 96:480–486. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gutmann DH, Hirbe AC, Huang ZY and Haipek
CA: The protein 4.1 tumor suppressor, DAL-1, impairs cell motility,
but regulates proliferation in a cell-type-specific fashion.
Neurobiol Dis. 8:266–278. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Robb VA, Gerber MA, Hart-Mahon EK and
Gutmann DH: Membrane localization of the U2 domain of Protein 4.1B
is necessary and sufficient for meningioma growth suppression.
Oncogene. 24:1946–1957. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Franco-Chuaire ML, Magda Carolina SC and
Chuaire-Noack L: Epithelial-mesenchymal transition (EMT):
Principles and clinical impact in cancer therapy. Invest Clin.
54:186–205. 2013.PubMed/NCBI
|
|
14
|
Nieto MA: Epithelial-Mesenchymal
Transitions in development and disease: Old views and new
perspectives. Int J Dev Biol. 53:1541–1547. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elliott RL and Blobe GC: Role of
transforming growth factor Beta in human cancer. J Clin Oncol.
23:2078–2093. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goel A, Chhabra R, Ahmad S, Prasad AK,
Parmar VS, Ghosh B and Saini N: DAMTC regulates cytoskeletal
reorganization and cell motility in human lung adenocarcinoma cell
line: An integrated proteomics and transcriptomics approach. Cell
Death Dis. 3:e4022012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tran Y, Benbatoul K, Gorse K, Rempel S,
Futreal A, Green M and Newsham I: Novel regions of allelic deletion
on chromosome 18p in tumors of the lung, brain and breast.
Oncogene. 17:3499–505. 1998. View Article : Google Scholar
|
|
19
|
Yageta M, Kuramochi M, Masuda M, Fukami T,
Fukuhara H, Maruyama T, Shibuya M and Murakami Y: Direct
association of TSLC1 and DAL-1, two distinct tumor suppressor
proteins in lung cancer. Cancer Res. 62:5129–5133. 2002.PubMed/NCBI
|
|
20
|
Zhang Y, Xu R, Li G, Xie X, Long J and
Wang H: Loss of expression of the differentially expressed in
adenocarcinoma of the lung (DAL-1) protein is associated with
metastasis of non-small cell lung carcinoma cells. Tumour Biol.
33:1915–1925. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bernkopf DB and Williams ED: Potential
role of EPB41L3 (protein 4.1B/Dal-1) as a target for treatment of
advanced prostate cancer. Expert Opin Ther Targets. 12:845–853.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cavanna T, Pokorna E, Vesely P, Gray C and
Zicha D: Evidence for protein 4.1B acting as a metastasis
suppressor. J Cell Sci. 120:606–616. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ohno N, Terada N, Murata S, Yamakawa H,
Newsham IF, Katoh R, Ohara O and Ohno S: Immunolocalization of
protein 4.1B/DAL-1 during neoplastic transformation of mouse and
human intestinal epithelium. Histochem Cell Biol. 122:579–586.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Christofori G and Semb H: The role of the
cell-adhesion molecule E-cadherin as a tumour-suppressor gene.
Trends Biochem Sci. 24:73–76. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Q, Mao H, Nie J, Chen W, Yang Q, Dong
X and Yu X: Transforming growth factor {beta}1 induces
epithelial-mesenchymal transition by activating the JNK-Smad3
pathway in rat peritoneal mesothelial cells. Perit Dial Int.
28(Suppl 3): S88–S95. 2008.PubMed/NCBI
|
|
26
|
Xu XL, Ling ZQ, Chen SZ, Li B, Ji WH and
Mao WM: The impact of E-cadherin expression on the prognosis of
esophageal cancer: A meta-analysis. Dis Esophagus. 27:79–86. 2014.
View Article : Google Scholar
|
|
27
|
Nakaya Y and Sheng G: EMT in developmental
morphogenesis. Cancer Lett. 341:9–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saito A: EMT and EndMT: Regulated in
similar ways? J Biochem. 153:493–495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Willis BC and Borok Z: TGF-beta-induced
EMT: Mechanisms and implications for fibrotic lung disease. Am J
Physiol Lung Cell Mol Physiol. 293:L525–L534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Duangkumpha K, Techasen A, Loilome W,
Namwat N, Thanan R, Khuntikeo N and Yongvanit P: BMP-7 blocks the
effects of TGF-beta-induced EMT in cholangiocarcinoma. Tumour Biol.
35:9667–9676. 2014. View Article : Google Scholar : PubMed/NCBI
|