1. Introduction
Interleukin (IL-33) is a cytokine, which belongs to
the IL-1 superfamily, and induces T helper (Th) cells to produce
type 2 cytokines (1). In 1999,
Onda et al identified the Dvs27 gene in canine vasospastic
cerebral arteries following subarachnoid hemorrhage (2). Nuclear factor from high endothelial
venule (NF-HEV) was cloned in 2003 (3), and 2 years later, Schmitz et
al determined the Dvs27 and NF-HEV definitions as the same type
of molecule, which was termed IL-33 (4). IL-33 mediates its biological effects
by interacting with the receptors suppression of tumorigenicity 2
(ST2) and IL-1 receptor accessory protein (IL-1RAcP), activating
intracellular molecules in the NF-κB and mitogen-activated protein
kinase signaling pathways, which drive the production of type 2
cytokines, including IL-4, IL-5 and IL-13 from polarized Th2 cells
(5). The induction of type 2
cytokines by IL-33 in vivo is considered to induce severe
pathological changes in mucosal organs (6).
The focus of the present review is on the properties
of this cytokine in kidney diseases, as well as in renal graft
damage associated with renal transplantation. Understanding the
involvement of IL-33 in the pathogenesis of kidney diseases may
assist in identifying novel therapeutic strategies to mitigate or
prevent kidney diseases.
2. Structure of IL-33
IL-33, also termed IL-F11, is a novel member of the
IL-1 superfamily. The human IL-33 gene is located on chromosome
9p24.1, comprises 270 amino acids and the relative molecular mass
of full-length proteins is ~30 kDa. The mouse IL-33 gene can be
found on the chromosome 19qC1 region and encodes 266 amino acid
polypeptides, corresponding to full-length proteins with a
calculated mass of 29.9 kDa (4).
IL-33 has, in its amino-terminal portion, a helix-corner-helix
structural pattern, which involves a chromatin binding motif and
nuclear localization signal. In its carboxy-terminal portion is an
IL-1-like β-trefoil domain, which binds with the orphan receptor,
ST2 (7). Roussel et al
showed that IL-33 combines to chromatin by an acidic pocket area of
histone H2A-H2B (8).
3. Distribution of IL-33
In humans, the constitutive and widespread
expression of IL-33 can be detected in several normal tissues. For
example, IL-33 is constitutively expressed in human secondary
lymphoid tissues, including the lymph nodes and appendix (9), and widespread expression is observed
along the vascular tree, including large and small blood vessels
from normal tissues, including the liver, skeletal muscle, kidney
and prostate, despite the microcirculation of the brain and kidney
glomeruli (9). In certain tissues
exposed to the external environment, high levels of IL-33 have also
been found, including the skin, mucosal surfaces and gastric glands
in the stomach, as well as in tonsillar crypts and salivary glands
(9). Furthermore, the accumulation
of IL-33 has been reported in adenocarcinoma of the kidney,
stomach, liver, pancreas, lung, breast and colon (9). In addition, IL-33 is substantially
elevated in lymphoid tissues, the synovium in chronically inflamed
rheumatoid arthritis and the intestines in Crohn's disease
(10). Together, these results
indicate that IL-33 is broadly expressed in normal, tumor and
chronically inflamed human tissues. High mRNA expression levels of
IL-33 have been found in stomach, lung, spinal cord, brain and skin
in the mouse, whereas low mRNA expression levels of IL-33 have been
detected in mouse lymph tissue, the spleen, pancreas, kidney and
heart (3).
Compared with its expression in tissue, IL-33 mRNA
is more restricted at the cellular level. Activated dendritic cells
and macrophages are the only hematopoietic cells to exhibit low
mRNA expression levels of human IL-33. By contrast, IL-33 mRNA has
been found in resting dendritic cells and activated macrophages in
mice (11). Human smooth muscle
cells (SMCs) of various tissues, as well as epithelial cells
forming the bronchus or small airways exhibit constitutive
expression of IL-33 mRNA (11). In
addition, high expression levels of IL-33 have been confirmed in
activated dermal fibroblasts, activated and resting bronchial SMCs,
resting pulmonary artery SMCs, resting coronary artery SMCs and
bronchial epithelial cells (4).
The distributions of IL-33 in humans in mice are presented in
Table I.
 | Table IDistribution of interleukin-33 in
humans and mice. |
Table I
Distribution of interleukin-33 in
humans and mice.
| Site | Human | Mouse | Reference |
|---|
| Tissue | | | (3,9,10) |
| Appendix | + | NA | |
| Brain | − | + | |
| Colon | + | − | |
| Kidney | + | + | |
| Liver | + | + | |
| Lung | + | + | |
| Lymph nodes | + | + | |
| Pancreas | + | + | |
| Prostate | + | NA | |
| Salivary
glands | + | NA | |
| Skeletal
muscle | + | NA | |
| Skin | + | + | |
| Spinal cord | NA | + | |
| Stomach | + | + | |
| Tonsillar
crypts | + | NA | |
| Cells | | | (4,8–11) |
| Activated dermal
fibroblasts | + | NA | |
| Activated
macrophages | + | + | |
| Epithelial
cells | + | + | |
| Resting dendritic
cells | − | + | |
| Smooth muscle
cells | + | NA | |
4. Cleavage of IL-33
It has been suggested that IL-33 is produced as a
pro-form IL-33 (pro-IL-33), and is digested into a mature form with
a lower molecular weight when it is secreted from the cells
(12). Mature IL-33 was initially
considered to be the active form, however, subsequent reports have
shown that the active pro-form of IL-33 is digested into an
inactive mature form (4,13). The initial investigation of IL-33
suggested that it is activated via caspase-1-dependent proteolysis,
similar to the proinflammatory cytokines, IL-1β and IL-18 (14). By contrast, Cayrol and Girard
reported that full-length IL-33 (1-270) is active, and that
processing by caspase-1 results in IL-33 inactivation, rather than
activation (13). Another previous
report independently arrived at the conclusion that the executioner
caspase-3 and caspase-7 inactivate IL-33 by cleaving the
carboxy-terminal IL-1-like structure to prevent an inappropriate
immune response during apoptosis, but not in necrosis (15).
5. Secretion of IL-33
In addition to the conventional secretion approaches
for cytokines, including autocrine, paracrine, intracrine,
juxtacrine and retrocrine pathways, full-length IL-33, as with high
mobility group protein-1, can also be released into the
extracellular space following cell damage or mechanical injury
(13). The release of IL-33 by
necrotic cells is another recognized mechanism for a cytokine to
exert its function, termed a 'necrocrine' pathway (16). The necrocrine pathway can be
deleted by endogenous apoptotic caspases in cells undergoing
apoptosis (15,17). Therefore, IL-33 functions as an
extracellular 'danger signal' in a necrocrine manner, to alert the
immune system during infectious and autoimmune diseases.
6. Receptors of IL-33
IL-33R is a heterodimer comprised of IL-1RL1, also
ST2, and IL-1RAcP (18). ST2,
which exhibits marked homology to the ligand-binding subunits of
the IL-1 and IL-18 receptor complexes, was identified in 1989,
prior the identification of IL-33, and has been termed an 'orphan
receptor' (19). The human ST2
gene is located on chromosome 2, and its germline sequence is
conserved (20). IL-33 mediates
signal transduction through ST2, which is expressed on mast cells
and Th2 cella, but not Th1 cells (21,22).
Enhancing the expression of ST2 is associated with an increased
risk of developing atopic dermatitis (23).
ST2 has two major forms: soluble (s)ST2 and
membrane-bound ST2 (ST2 L), which are produced from the IL-1RL1
gene as a result of alternative splicing under the control of two
distinct promoters (24–26). sST2 is a soluble ST2, which has no
transmembrane sequence, therefore, it can be excreted outside
cells. Increased levels of soluble ST2 have been associated with
several human diseases, including acute myocardial infarction,
asthma with acute exacerbation, eosinophilic pneumonia, sepsis and
trauma, and exacerbated idiopathic pulmonary fibrosis (27–32).
ST2L is the trans-membrane ST2, possessing a transmembrane
sequence, and is considered to be a functional component of IL-33R,
whereas sST2 is regarded as a decoy receptor for IL-33 (33). T cell-associated ST2L augments Th2
immune responses, however, macrophage-associated ST2L has been
reported to exhibit anti-inflammatory activity (34). There are two splice variants of
ST2: ST2V and ST2LV, produced via loss of the third immunoglobulin
motif and alternative splicing in the C-terminal portion of ST2
(35).
7. IL-33/ST2 signaling pathway
IL-33 can transfer extracellular information through
binding of a receptor complex comprised of ST2 and IL-1RAcP as a
cytokine-alarmin (9). ST2L and
IL-1RAcP are necessary for IL-33 action (36,37).
Extracellular IL-33 signals results in recruitment of myeloid
differentiation primary response gene 88, IL-1 receptor-associated
kinase 1 and tumor necrosis factor (TNF) receptor-associated factor
6, leading to activation of the transcription factor NF-kB, c-Jun
N-terminal kinase1/2 and extracellular regulated protein kinase
1/2, and finally causing inflammatory responses (4,36–38).
IL-33-mediated signalling can be inhibited by single Ig
IL-1-related molecule, also known as Toll IL-1R8, through
interactions with the IL-33 receptor complex (39) (Fig.
1).
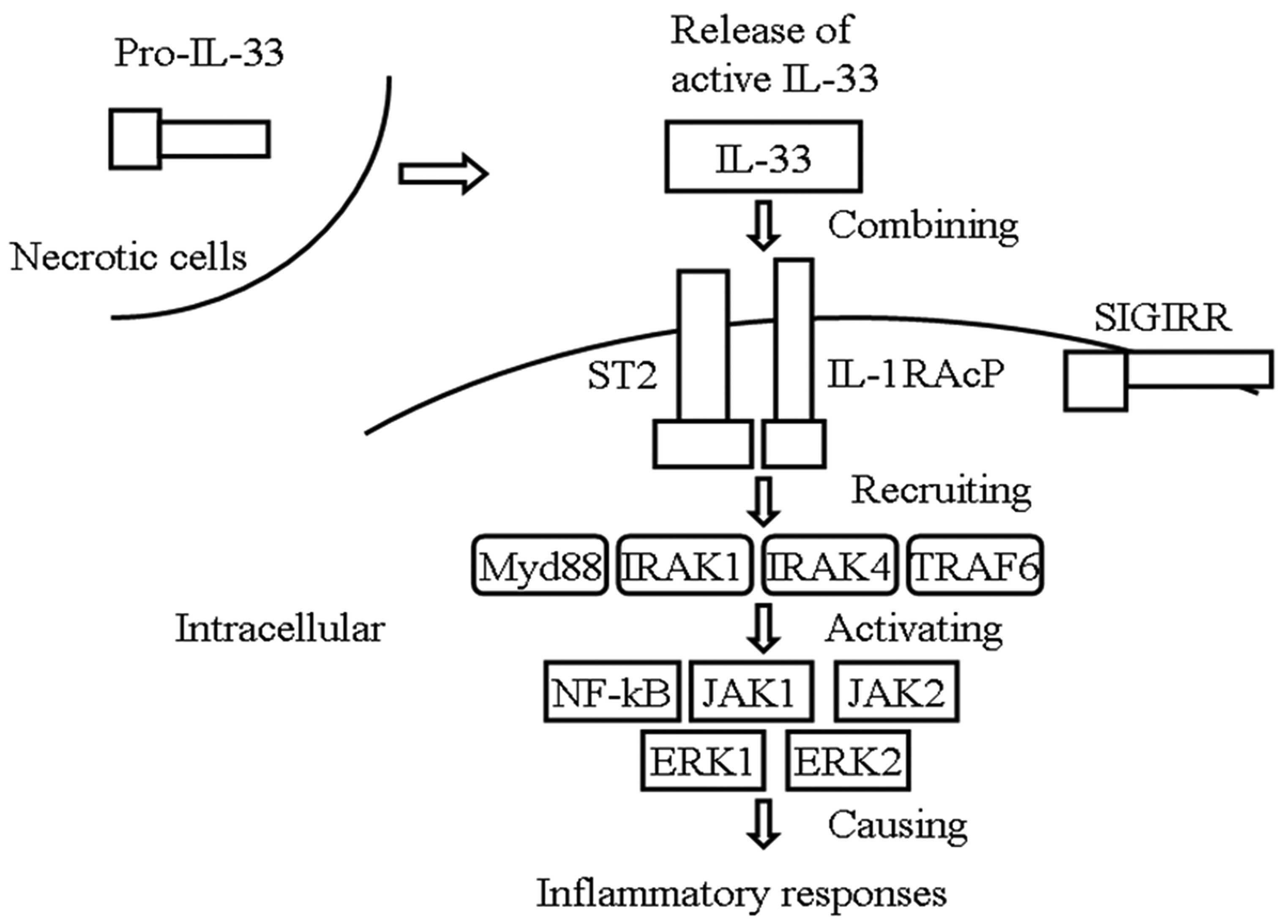 | Figure 1IL-33/ST2 signaling pathway. IL-33
can transfer extracellular information through the binding of the
receptor complex. comprised of ST2 and IL-1RAcP, following release
into the extracellular space during cell damage or mechanical
injury. Extracellular IL-33 signals result in the recruitment of
Myd88, IRAK1/4 and TRAF6, leading to the activation of
transcription factor NF-kB, JNK1/2 and ERK1/2, causing inflammatory
responses. IL-33-mediated signaling can be inhibited by SIGIRR
through interactions with the IL-33 receptor complex. IL,
interleukin; ST2, suppression of tumorigenicity 2; IL-1RAcP, IL-1
receptor accessory protein; Myd88, myeloid differentiation primary
response gene 88; IRAK1/4, interleukin-1 receptor-associated kinase
1; TRAF6, tumor necrosis factor receptor-associated factor 6;
JNK1/2, c-Jun N-terminal kinase1/2; ERK1/2, extracellular regulated
protein kinase 1/2; SIGIRR, single IgIL-1-related molecule. |
8. Function of IL-33
Similar to the chromatin-associated cytokine,
high-mobility group box 1 (HMGB1), IL-33 acts as a dual-function
molecule, as a nuclear factor and cytokine. As a nuclear factor,
the transcriptional repressor function of IL-33 may be involved in
the nucleus. Küchler et al showed that nuclear IL-33 is
rapidly downregulated during wound healing and is lost in tumor
endothelium. In addition, activation of endothelial cell cultures
with either TNF-α or vascular endothelial growth factor and
subcutaneous injection of these cytokines also leads to a
downregulation in vascular IL-33 (40). The above evidence supports the
hypothesis that the transcriptional repressor function of IL-33 may
be involved in the control of endothelial cell activation.
As a cytokine, IL-33 promotes the polarization of T
cells towards a Th2 cell phenotype and is involved in Th2-type
responses through stimulating the production of IL-5, IL-6, IL-13
and granulocyte-macrophage colony-stimulating factor in vivo
(41). In addition, IL-33 has been
referred to as an 'endogenous danger signal' or 'alarmin', similar
to HMGB1, in order to alert the immune system of tissue damage and
infection, and to promote the initiation of a healing responses
(42).
9. IL-33 and kidney disease
IL-33 and chronic kidney disease
(CKD)
A study by Bao et al (43) on CKD aimed to examine the
association between serum levels of IL-33 and sST2, and disease
severity. This involved comparing the serum concentrations of IL-33
and sST2 between patients with CKD and healthy individuals. The
results showed no difference in the serum concentration of IL-33
between the patients with CKD and healthy individuals, whereas a
higher serum level of sST2 was found in the patients with CKD.
Therefore, the results revealed a significant correlation between
the serum level of sST2 and disease severity. In addition, higher
levels of sST2 correlated with elevated parathyroid hormone, serum
phosphorus and serum calcium (43). However, the expression of IL-33 was
observed to be increased in aortic endothelial cells from a mouse
model of CKD (44). Additionally,
higher concentrations of sST2 appeared to be associated with
impaired kidney function in a study involving participants with
cardiovascular disease (45).
Together, these findings indicate that the levels of IL-33 and sST2
are relevant to the progressive deterioration of kidney
function.
IL-33 and systemic lupus erythematosus
(SLE) nephropathy (LN)
Renal fibrosis is the common pathway of chronic
kidney disease eventually lead to kidney failure, which is one of
the most serious complications of SLE (46). Fibrotic disease is characterized by
the excess accumulation of extracellular matrix components,
including colla gen, and re quires eosinophils and RAG-dependent
lymphocytes (47). Of note, IL-33
mediates the regulation of several extracellular matrix-associated
genes, including collagen VI, collagen III and tissue inhibitor of
metalloproteases-1. In addition, the administration of IL-33
resultes in IL-33R-dependent accumulation of eosinophils,
RAG-dependent lymphocytes and CD3+ lymphocytes (48). The levels of IL-33 in patients with
SLE have been found to be greater than those in a healthy control
group, and were correlated with elevated erythrocyte sedimentation
rate and C-reactive protein, suggesting that the abnormal increase
in serum IL-33 is closely associated with the development of SLE
and may be involved in the acute phase reaction of SLE (49). Shui-Lian et al indicated
that the IL-33/ST2 axis has a detrimental effect in the
pathogenesis of renal fibrosis associated with LN (46). Therefore, IL-33 may involved in
renal fibrosis associated with SLE. Further mechanistic
investigations examining the precise physiological and
pathophysiological roles of IL-33 in SLE are required.
IL-33 and diabetic nephropathy
The principle of the pathogeny of type 2 diabetes
and its complications, including diabetic nephropathy, remain to be
fully elucidated, however predicting the potential complications of
diabetic patients can assist in early treatment. Whether IL-33 can
be used for predicting the early stage of kidney injury in diabetic
patients remains to be elucidated. In a study by Caner et al
identified three groups: Healthy group; diabetes mellitus (DM)
group without any known kidney disease; and DM+microalbuminuria
(MA) group, assumed to have nephropathy. Following assessment of
the concentrations of IL-33 in the three groups, it was found that
the level of IL-33 in the DM group was greater than that in the
healthy group; and the level of IL-33 in the DM+MA group was
greater than that in the healthy group; although no difference was
observed between the DM and DM+MA group. Therefore, IL-33 cannot be
used in the early recognition of diabetic nephropathy (50). A study by Miller et al
showed that the levels of sST2 in individuals largely without
vascular disease, are associated principally with markers
associated with diabetes, and support a role for sST2 in diabetes
(51). These findings support the
hypothesis that the increase in IL-33 levels in diabetic
nephropathy is not associated with kidney injury, but that the
increase may be a result of diabetes. Further investigations are
required to clarify the value of IL-33 in DM and the early stage of
kidney injury.
IL-33 and renal transplantation
Ischemia-reperfusion injury (IRI) contributes to the
develop ment of renal graft damage associated with renal
transplantation (52).
Inflammatory and immune responses are involved in kidney IRI
(52). IL-33 has been identified
as an alarmin, capable of mediating danger signals during tissue
damage (53). Thierry et al
addressed the role of IL-33 in IRI following human kidney
transplantation (54). This
involved analysis of the levels of IL-33 in a cohort of 26 deceased
renal transplant recipients, and revealed that the level of IL-33
was significantly increased as soon as 30 min post-reperfusion,
which supported the potential role of IL-33 as an immune mediator
following transplantation during kidney IRI in humans (54). Consistent with this, invariant
natural killer T cells, which have been suggested to be crucial in
IRI and targeted by IL-33, exhibited a state of early activation in
kidney transplant recipients (54). In addition, a significant
correlation was found between serum and urinary levels of IL-33
levels and cold ischemia duration, between 30 min and 3 days
post-tranplantation (55). In
conclusion, these results emphasize the possible role of IL-33 as
an innate-immune mediator during IRI in humans.
IL-33 and acute kidney injury (AKI)
AKI contributes to significant morbidity and
mortality rates in intensive care units (56). Alterations in renal hemodynamics,
inflammation, endothelial dysfunction, tubular obstruction and
glomerular thrombosis are involved in the pathogenesis of AKI
(57). To determine whether IL-33
promotes AKI, a study by Akcay et al examined the protein
expression of IL-33 in the kidney using an AKI mouse model.
Following neutralizing IL-33 activity with sST2, these mice had
fewer CD4 T cells infiltrating the kidney, lower levels of serum
creatinine, and reduced acute tubular necrosis and apoptosis,
compared with cisplatin-induced AKI in the untreated mice. By
contrast, the administration of recombinant IL-33 exacerbated
cisplatin-induced AKI (58). Of
note, IL-33 mediates cisplatin-induced AKI by acting as an
proinflammatory cytokine, whereas IL-10 protects against
cisplatin-induced AKI by acting as an anti-inflammatory cytokine
(59). In addition, high
expression levels of IL-33 have been observed in
lipopolysaccharide-induced acute glomerular injury (57). The inhibition of IL-33 may provide
a novel strategy in the treatment of AKI (Table II).
| Table IIRole of IL-33 in kidney diseases. |
Table II
Role of IL-33 in kidney diseases.
| Kidney disease | Role of IL-33 | Reference |
|---|
| CKD | Serum levels of
IL-33 not correlated with disease severity in CKD. | (44,45) |
| Serum levels of
sST2 are significantly correlated with disease severity in
CKD. | |
| Systemic lupus
erythematosus nephropathy | Mediates regulation
of several extracellular matrix-associated genes, resulting in
IL-33R-dependent accumulation of eosinophils, RAG-dependent
lymphocytes and CD3+ lymphocytes. | (47–49) |
| Diabetic
nephropathy | Expression of IL-33
is not associated with kidney injury, but the increase may be a
result of diabetes. | (50,51) |
| Renal
transplantation | Immune mediator
following transplantation during kidney IRI in humans; correlated
with cold ischemia duration; activates invariant natural killer T
cells in kidney transplant recipients. | (53–55) |
| Acute kidney
injury | Stimulates
CD4+ T cell infiltration in the kidney, induces higher
levels of serum creatinine, acute tubular necrosis and
apoptosis. | (57–59) |
10. Conclusion
In conclusion, increasing evidence indicates that
the IL-33/ST2 axis has a significant effect in the pathogenesis of
kidney disease. Although the mechanism underlying the effects of
IL-33 in kidney disease remains to be fully elucidated,
accumulating evidence links IL-33 to the nephropathies, indicating
that the antagonism of IL-33 may be a novel strategy for the
treatment of kidney disease. Further detailed investigations of the
association between IL-33 and kidney disease are required in the
future.
Acknowledgments
This study was supported by the Natural Science
Foundation of Hubei province (grant. no. 2012FFB03708) and the
Natural Science Foundation of Yichang City (grant. no.
A13301-020).
References
|
1
|
Louten J, Rankin AL, Li Y, Murphy EE,
Beaumont M, Moon C, Bourne P, McClanahan TK, Pflanz S and de Waal
Malefyt R: Endogenous IL-33 enhances Th2 cytokine production and
T-cell responses during allergic airway inflammation. Int Immunol.
23:307–315. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Onda H, Kasuya H, Takakura K, et al:
Identification of genes differentially expressed in canine
vasospastic cerebral arteries after subarachnoid hemorrhage. J
Cereb Blood Flow Metab. 19:1279–1288. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baekkevold ES, Roussigné M, Yamanaka T,
Johansen FE, Jahnsen FL, Amalric F, Brandtzaeg P, Erard M,
Haraldsen G and Girard JP: Molecular characterization of NF-HEV, a
nuclear factor preferentially expressed in human high endothelial
venules. Am J Pathol. 163:69–79. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schmitz J, Owyang A, Oldham E, Song Y,
Murphy E, McClanahan TK, Zurawski G, Moshrefi M, Qin J, Li X,
Gorman DM, et al: IL-33, an interleukin-1-like cytokine that
signals via the IL-1 receptor-related protein ST2 and induces T
helper type 2-associated cytokines. Immunity. 23:479–490. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marian AJ: Pathogenesis of diverse
clinical and pathological phenotypes in hypertrophic
cardiomyopathy. Lancet. 355:58–60. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartemes KR, Iijima K, Kobayashi T,
Kephart GM, McKenzie AN and Kita H: IL-33-responsive lineage- CD25+
CD44(hi) lymphoid cells mediate innate type 2 immunity and allergic
inflammation in the lungs. J Immunol. 188:1503–1513. 2012.
View Article : Google Scholar :
|
|
7
|
Miller AM and Liew FY: The IL-33/ST2
pathway–A new therapeutic target in cardiovascular disease.
Pharmacol Ther. 131:179–186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roussel L, Erard M, Cayrol C and Girard
JP: Molecular mimicry between IL-33 and KSHV for attachment to
chromatin through the H2A-H2B acidic pocket. EMBO Rep. 9:1006–1012.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moussion C, Ortega N and Girard JP: The
IL-1-like cytokine IL-33 is constitutively expressed in the nucleus
of endothelial cells and epithelial cells in vivo: A novel
'alarmin'? PloS one. 3:e33312008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carriere V, Roussel L, Ortega N, Lacorre
DA, Americh L, Aguilar L, Bouche G and Girard JP: IL-33, the
IL-1-like cytokine ligand for ST2 receptor, is a
chromatin-associated nuclear factor in vivo. Proc Natl Acad Sci
USA. 104:282–287. 2007. View Article : Google Scholar :
|
|
11
|
Nakajima A: Application of cellular gene
therapy for rheumatoid arthritis. Mod Rheumatol. 16:269–275. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tsuda H, Komine M, Karakawa M, Etoh T,
Tominaga S and Ohtsuki M: Novel splice variants of IL-33:
Differential expression in normal and transformed cells. J Invest
Dermatol. 132:2661–2664. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cayrol C and Girard JP: The IL-1-like
cytokine IL-33 is inactivated after maturation by caspase-1. Proc
Natl Acad Sci USA. 106:9021–9026. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Creagh EM, Conroy H and Martin SJ:
Caspase-activation pathways in apoptosis and immunity. Immunol Rev.
193:10–21. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luthi AU, Cullen SP, McNeela EA, Duriez
PJ, Afonina IS, Sheridan C, Brumatti G, Taylor RC, Kersse K,
Vandenabeele P, et al: Suppression of interleukin-33 bioactivity
through proteolysis by apoptotic caspases. Immunity. 31:84–98.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao W and Hu Z: The enigmatic processing
and secretion of interleukin-33. Cell Mol Immunol. 7:260–262. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lamkanfi M and Dixit VM: IL-33 raises
alarm. Immunity. 31:5–7. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakae S, Morita H, Ohno T, Arae K,
Matsumoto K and Saito H: Role of interleukin-33 in innate-type
immune cells in allergy. Allergol Int. 62:13–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tominaga S: A putative protein of a growth
specific cDNA from BALB/c-3T3 cells is highly similar to the
extracellular portion of mouse interleukin 1 receptor. FEBS Lett.
258:301–304. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luzina IG, Pickering EM, Kopach P, Kang
PH, Lockatell V, Todd NW, Papadimitriou JC, McKenzie AN and Atamas
SP: Full-length IL-33 promotes inflammation but not Th2 response in
vivo in an ST2-independent fashion. J Immunol. 189:403–410. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Löhning M, Stroehmann A, Coyle AJ, Grogan
JL, Lin S, Gutierrez-Ramos JC, Levinson D, Radbruch A and Kamradt
T: T1/ST2 is preferentially expressed on murine Th2 cells,
independent of interleukin 4, interleukin 5 and interleukin 10 and
important for Th2 effector function. Proc Natl Acad Sci USA.
95:6930–6935. 1998. View Article : Google Scholar
|
|
22
|
Yanagisawa K, Naito Y, Kuroiwa K, Arai T,
Furukawa Y, Tomizuka H, Miura Y, Kasahara T, Tetsuka T and Tominaga
S: The expression of ST2 gene in helper T cells and the binding of
ST2 protein to myeloma-derived RPMI8226 cells. J Biochm.
121:95–103. 1997. View Article : Google Scholar
|
|
23
|
Shimizu M, Matsuda A, Yanagisawa K, Hirota
T, Akahoshi M, Inomata N, Ebe K, Tanaka K, Sugiura H, Nakashima K,
et al: Functional SNPs in the distal promoter of the ST2 gene are
associated with atopic dermatitis. Hum Mol Genet. 14:2919–2927.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin J, Zhang L, Zhao G, Su Z, Deng R,
Pflugfelder SC and Li DQ: A novel interleukin 33/ST2 signaling
regulates inflammatory response in human corneal epithelium. PloS
One. 8:e609632013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Oboki K, Ohno T, Kajiwara N, Saito H and
Nakae S: IL-33 and IL-33 receptors in host defense and diseases.
Allergol Int. 59:143–160. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Smith DE: IL-33: A tissue derived cytokine
pathway involved in allergic inflammation and asthma. Clin Exp
Allergy. 40:200–208. 2010. View Article : Google Scholar
|
|
27
|
Oshikawa K, Kuroiwa K, Tago K, Iwahana H,
Yanagisawa K, Ohno S, Tominaga SI and Sugiyama Y: Elevated soluble
ST2 protein levels in sera of patients with asthma with an acute
exacerbation. Am J Respir Crit Care Med. 164:277–281. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Oshikawa K, Kuroiwa K, Tokunaga T, Kato T,
Hagihara SI, Tominaga SI and Sugiyama Y: Acute eosinophilic
pneumonia with increased soluble ST2 in serum and bronchoalveolar
lavage fluid. Respir Med. 95:532–533. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Weinberg EO, Shimpo M, De Keulenaer GW,
MacGillivray C, Tominaga S, Solomon SD, Rouleau JL and Lee RT:
Expression and regulation of ST2, an interleukin-1 receptor family
member, in cardiomyocytes and myocardial infarction. Circulation.
106:2961–2966. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tajima S, Oshikawa K, Tominaga S and
Sugiyama Y: The increase in serum soluble ST2 protein upon acute
exacerbation of idiopathic pulmonary fibrosis. Chest.
124:1206–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brunner M, Krenn C, Roth G, Moser B,
Dworschak M, Jensen-Jarolim E, Spittler A, Sautner T, Bonaros N,
Wolner E, et al: Increased levels of soluble ST2 protein and IgG1
production in patients with sepsis and trauma. Intensive Care Med.
30:1468–1473. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shimpo M, Morrow DA, Weinberg EO, Sabatine
MS, Murphy SA, Antman EM and Lee RT: Serum levels of the
interleukin-1 receptor family member ST2 predict mortality and
clinical outcome in acute myocardial infarction. Circulation.
109:2186–2190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ohno T, Morita H, Arae K, Matsumoto K and
Nakae S: Interleukin-33 in allergy. Allergy. 67:1203–1214. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Brint EK, Xu D, Liu H, Dunne A, McKenzie
AN, O'Neill LA and Liew FY: ST2 is an inhibitor of interleukin 1
receptor and toll-like receptor 4 signaling and maintains endotoxin
tolerance. Nat Immunol. 5:373–379. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kakkar R and Lee RT: The IL-33/ST2
pathway: Therapeutic target and novel biomarker. Nat Rev Drug
Discov. 7:827–840. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kurowska-Stolarska M, Kewin P, Murphy G,
Russo RC, Stolarski B, Garcia CC, Komai-Koma M, Pitman N, Li Y,
Niedbala W, et al: IL-33 induces antigen-specific IL-5+ T cells and
promotes allergic-induced airway inflammation independent of IL-4.
J Immunol. 181:4780–4790. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chackerian AA, Oldham ER, Murphy EE,
Schmitz J, Pflanz S and Kastelein RA: IL-1 receptor accessory
protein and ST2 comprise the IL-33 receptor complex. J Immunol.
179:2551–2555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kurowska-Stolarska M, Hueber A, Stolarski
B and McInnes IB: Interleukin-33: A novel mediator with a role in
distinct disease pathologies. J Internal Med. 269:29–35. 2011.
View Article : Google Scholar
|
|
39
|
Bulek K, Swaidani S, Qin J, Lu Y, Gulen
MF, Herjan T, Min B, Kastelein RA, Aronica M, Kosz-Vnenchak M and
Li X: The essential role of single Ig IL-1 receptor-related
molecule/Toll IL-1R8 in regulation of Th2 immune response. J
Immunol. 182:2601–2609. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Küchler AM, Pollheimer J, Balogh J,
Sponheim J, Manley L, Sorensen DR, De Angelis PM, Scott H and
Haraldsen G: Nuclear interleukin-33 is generally expressed in
resting endothelium but rapidly lost upon angiogenic or
proinflammatory activation. Am J Pathol. 173:1229–1242. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Smithgall MD, Comeau MR, Yoon BR, Kaufman
D, Armitage R and Smith DE: IL-33 amplifies both Th1- and Th2-type
responses through its activity on human basophils,
allergen-reactive Th2 cells, iNKT and NK cells. Int Immunol.
20:1019–1030. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bianchi ME: DAMPs, PAMPs and alarmins: All
we need to know about danger. J Leukoc Biol. 81:1–5. 2007.
View Article : Google Scholar
|
|
43
|
Bao YS, Na SP, Zhang P, Jia XB, Liu RC, Yu
CY, Mu SH and Xie RJ: Characterization of interleukin-33 and
soluble ST2 in serum and their association with disease severity in
patients with chronic kidney disease. J Clin Immunol. 32:587–594.
2012. View Article : Google Scholar
|
|
44
|
Wiese CB, Toth CL, Tabet F, Taylor RC,
Landstreet SR, Rye KA, Hofmeister LH, Harrison DG, Kon V and
Vickers KC: HDL-microRNA-92a and interleukin-33 axis underlies
endothelial dysfunction associated with atherosclerosis and chronic
kidney disease. Arterioscler Thromb Vasc Biol. 34:A3. 2014.
|
|
45
|
Januzzi JL Jr, Peacock WF, Maisel AS, Chae
CU, Jesse RL, Baggish AL, O'Donoghue M, Sakhuja R, Chen AA, van
Kimmenade RR, et al: Measurement of the interleukin family member
ST2 in patients with acute dyspnea: Results from the PRIDE
(Pro-Brain natriuretic peptide investigation of dyspnea in the
emergency department) study. J Am Coll Cardiol. 50:607–613. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yu SL, Wong CK and Tam LS: The alarmin
functions of high-mobility group box-1 and IL-33 in the
pathogenesis of systemic lupus erythematosus. Expert Rev Clin
Immunol. 9:739–749. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wynn TA: Cellular and molecular mechanisms
of fibrosis. J Pathol. 214:199–210. 2008. View Article : Google Scholar
|
|
48
|
Rankin AL, Mumm JB, Murphy E, Turner S, Yu
N, McClanahan TK, Bourne PA, Pierce RH, Kastelein R and Pflanz S:
IL-33 induces IL-13-dependent cutaneous fibrosis. J Immunol.
184:1526–1535. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang Z, Liang Y, Xi W, Li C and Zhong R:
Association of increased serum IL-33 levels with clinical and
laboratory characteristics of systemic lupus erythematosus in
Chinese population. Clin Exp Med. 11:75–80. 2011. View Article : Google Scholar
|
|
50
|
Caner S, Usluoğullari CA, Balkan F,
Büyükcam F, Kaya C, Saçıkara M, Koca C, Ersoy R and Çakır B: Is
IL-33 useful to detect early stage of renal failure? Ren Fail.
36:78–80. 2014. View Article : Google Scholar
|
|
51
|
Miller AM, Purves D, McConnachie A,
Asquith DL, Batty GD, Burns H, Cavanagh J, Ford I, McLean JS,
Packard CJ, et al: Soluble ST2 associates with diabetes but not
established cardiovascular risk factors: A new inflammatory pathway
of relevance to diabetes? PloS One. 7:e478302012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Basile DP, Anderson MD and Sutton TA:
Pathophysiology of acute kidney injury. Compr Physiol. 2:1303–1353.
2012.PubMed/NCBI
|
|
53
|
Galli SJ, Nakae S and Tsai M: Mast cells
in the development of adaptive immune responses. Nat Immunol.
6:135–142. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
54
|
Thierry A, Giraud S, Robin A, Barra A,
Bridoux F, Ameteau V, Hauet T, Girard JP, Touchard G, Gombert JM
and Herbelin A: The alarmin concept applied to human renal
transplantation: Evidence for a differential implication of HMGB1
and IL-33. PloS One. 9:e887422014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gombert JM, Thierry A, Robin A, Barra A,
Thierry H, Touchard G and Herbelin A: Study of alarmin release
during ischemia reperfusion injury after human renal
transplantation (P2214). J Immunol. 190:69.452013.
|
|
56
|
Singbartl K and Kellum JA: AKI in the ICU:
Definition, epidemiology, risk stratification, and outcomes. Kidney
Int. 81:819–825. 2012. View Article : Google Scholar
|
|
57
|
Lee SJ, Borsting E, Declèves AE, Singh P
and Cunard R: Podocytes express IL-6 and lipocalin 2/neutrophil
gelatinase-associated lipocalin in lipopolysaccharide-induced acute
glomerular injury. Nephron Exp Nephrol. 121:e86–96. 2012.
View Article : Google Scholar
|
|
58
|
Akcay A, Nguyen Q, He Z, Turkmen K, Won D,
Hernando AA, Altmann C, Toker A, Pacic A, Ljubanovic DG, et al:
IL-33 exacerbates acute kidney injury. J Am Soc Nephrol.
22:2057–2067. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ozkok A and Edelstein CL: Pathophysiology
of cisplatin-induced acute kidney injury. Biomed Res Int.
2014:9678262014. View Article : Google Scholar : PubMed/NCBI
|














