Introduction
Curcumin (diferuloylmethane), which is the main
component extracted from turmeric (Curcuma longa), is a
traditional medicinal plant that exerts various biological
functions (1–3), including anti-inflammatory,
antioxidant, anticancer and cardio-protective effects. The exact
mechanism by which curcumin exerts these effects remains to be
elucidated; however, the antioxidant activity of the hydrophobic
polyphenol appears to be the essential component underlying its
pleiotropic biological effects (4). Previous studies have demonstrated
that curcumin is able to ameliorate the production of reactive
oxygen species (ROS) and lipid peroxidation in various models of
oxidative damage in cardiac tissue (5,6).
Endothelial cell injury is a critical step in the development of
atherosclerosis and hypertension (7). In addition, it has previously been
reported that curcumin may attenuate oxidative damage in
endothelial cells (8); however,
the underlying mechanism remains unclear.
Apoptosis, which is the process of programmed cell
death, leads to the rapid degradation of cellular structures and
organelles (9). Furthermore,
autophagy is a highly conserved cellular process that comprises
bulk degradation and recycling of cytoplasmic components, including
long-lived proteins and organelles (10). The functional relationship between
apoptosis and autophagy is complex. Autophagy-induced
cytoprotection is the basic cellular function of autophagy in
eukaryotic cells; however, some stressors, including oxidative
stress, induce excessive autophagy that may result in heart disease
(11). Active autophagy can be
either pro-survival (adaptive) or anti-survival (maladaptive).
Complete abrogation of cardiomyocyte autophagy is detrimental to
cardiac homeostasis under basal conditions, whereas upregulation of
autophagy in failing heart tissue is an adaptive response that
protects cells from hemodynamic stress (12). Autophagy has distinct roles in
various types of heart disease, in certain conditions, the response
is beneficial, in other cases, it can promote disease progression.
Targeting autophagy in the cardiovascular system may be
therapeutically relevant. It has previously been hypothesized that
induction of autophagy and inhibition of apoptosis may be the
mechanism underlying the protective effects of curcumin against
oxidative stress.
The phosphoinositide 3-kinase/Akt/mammalian target
of rapamycin (PI3K/Akt/mTOR) pathway is closely associated with the
regulation of autophagy for its role in cell survival,
proliferation and differentiation (13). mTOR, which is an amino acid, ATP
and hormone receptor, may inhibit autophagy. Conversely, inhibition
of mTOR by nutritional deficiency or the direct use of rapamycin
may activate autophagy-related 1, and thus promote autophagy
(14,15). By modulating the mTOR signaling
pathway, cell apoptosis and autophagy can be adjusted in numerous
cells.
The aim of the present study was to determine the
molecular mechanism of action of curcumin in
H2O2-treated EA.hy926 cells. The EA.hy926
human umbilical vein endothelial cell line was pretreated with
various concentrations of curcumin prior to hydrogen peroxide
(H2O2) stimulation, in order to explore the
potential underlying mechanism. Alterations in the expression of
autophagy and apoptosis-related proteins, cell viability, and
activation of the Akt/mTOR pathway in curcumin-pretreated EA.hy926
cells were determined following H2O2
stimulation.
Materials and methods
Cell culture and induction of oxidative
stress
The EA.hy926 cell line was purchased from the Cell
Bank of the Chinese Academy of Science (Shanghai, China). The cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.),
penicillin (10,000 units/l; Wisent Inc., Quebec, Canada) and
streptomycin (100,000 µg/l) at 37°C in an atmosphere
containing 95% air/5% CO2. The media were changed every
2 days. The cells were allowed to grow to 80% confluence within 24
h prior to drug treatment. The cells were pretreated with curcumin
(5–20 µmol/l; Sigma-Aldrich, St. Louis, MO, USA) for 4 h,
after which the medium was removed and replaced with medium
containing various concentrations of curcumin alongside 200
µmol/l H2O2 (Nanjing Chemical Reagent,
Co., Ltd., Nanjing, China). The medium of the control group was
changed at 4 h and cells were not treated with curcumin or
H2O2. Following an additional 4 h incubation
at 37°C, the cells were assessed.
Cytotoxicity assays
Cell viability was determined using the Cell
Counting kit-8 (CCK-8) colorimetric assay (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan). Briefly, the cells were
seeded into a 96-well cell culture plate (1×104
cells/well), and were pretreated with various concentrations of
curcumin and H2O2. To measure cell viability,
10 µl CCK-8 assay solution was added to each well, which
contained 100 µl medium, and the cells were incubated at
37°C for a further 4 h. Subsequently the optical densities of the
wells were measured at 540 nm using a microplate reader (ELx800;
BioTek Instruments, Inc., Winooski, VT, USA).
Measurement of apoptosis using Hoechst
33258 staining
Chromatin condensation was detected by nuclear
staining using Hoechst 33258. After pretreatment, the medium was
removed and the cells were fixed with 500 µl methyl hydrate
at room temperature for 15 min, before being washed three times
with phosphate-buffered saline (PBS). The cells were then stained
with 1 µl Hoechst 33258 (5 mg/ml; Sigma-Aldrich) in 1 ml
basal medium and incubated at room temperature in the dark for 20
min. Stained cells were visualized under a fluorescent microscope
(excitation, 350 nm; emission, 460 nm; BX51; Olympus, Tokyo,
Japan).
Determination of autophagosome
formation
Autophagy is controlled by autophagosome formation
(input) and autophagosome degradation (output), and the speed of
autophagosome turnover is defined as autophagic flux. A fluorescein
isothiocyanate (FITC)-labeled-microtubule-associated protein
1A/1B-light chain 3 (LC3) antibody (CYTO-ID® Autophagy
Detection kit) was used to detect autophagosome formation using the
CYTO-ID® Autophagy Detection kit (Enzo Life Sciences,
Inc., Farmingdale, NY, USA), according to the manufacturers'
protocol. The autophagy inducer rapamycin (500 nmol/l;
CYTO-ID® Autophagy Detection kit), which is often used
as a positive control of autophagy, was added to the cells for 18 h
at 37°C. The nuclei were then stained using
4′,6-diamidino-2-phenylindole (Beyotime Institute of Biotechnology,
Haimen, China). Subsequently, the cells were washed twice with 1X
Assay Buffer provided in the kit. Images of autophagic cells were
captured using a fluorescent microscope (BX51; Olympus) with a FITC
filter (excitation, 480 nm; emission, 530 nm).
Western blot analysis
For whole cell lysate preparations, cultured cells
were washed twice with cold PBS and immersed in
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). The cell lysate was harvested and centrifuged at
12,000 x g for 10 min at 4°C, and the supernatant was subsequently
collected. The protein concentration was determined using the
bicinchoninic acid protein assay (Beyotime Institute of
Biotechnology). Identical protein samples (20 µg/µl)
were separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred to a polyvinylidene difluo-ride
membrane (EMD Millipore, Billerica, MA, USA). The membrane was
blocked with 5% fat-free milk at room temperature for 1 h, and was
then incubated with the following primary antibodies: Rabbit
anti-B-cell lymphoma 2 (Bcl-2; 1:800; cat. no. sc-492; Bioworld
Technology Inc., St. Louis Park, MN, USA), rabbit
anti-Bcl-2-associated X protein (Bax; 1:800; cat. no. BS1030;
Bioworld Technology Inc.), rabbit anti-caspase-3 (1:1,000; cat. no.
ab52314; Abcam, Cambridge, MA, USA), rabbit monoclonal anti-human
cleaved caspase-3 (1:1,000; Abcam; cat. no. ab184787), anti-LC3
(1:1,000, cat. no. 3868; Cell Signaling Technology, Inc., Danvers,
MA, USA), rabbit anti-Akt (1:1,000; cat. no. 4691; Cell Signaling
Technology, Inc.), rabbit anti-phosphorylated (p)-Akt (1:1,000;
cat. no. 4060; Cell Signaling Technology, Inc.), rabbit anti-mTOR
(1:1,000; Abcam; cat. no. ab2732), rabbit anti-p-mTOR (1:1,000;
cat. no. ab84400; Abcam) and anti-β-actin (1:800; cat. no. AP0733;
Bioworld Technology, Inc.) at 4°C overnight. The membrane was
subsequently washed three times for 15 min with Tris-buffered
saline containing 0.1% Tween, and incubated with horseradish
peroxidase-conjugated secondary antibodies (1:500; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 1 h at room temperature.
The membrane was visualized using a western blotting detection
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The digital
image was analyzed for densitometry using ImageJ (version 1.49;
National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
GraphPad Prism 5 (GraphPad Software, Inc., La Jolla,
CA, USA) and SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA) were
used for statistical analysis and graphing. All data are presented
as the mean ± standard deviation. Statistical differences among
groups were analyzed by one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference.
Results
Curcumin protects against
H2O2-induced cytotoxicity in EA.hy926
cells
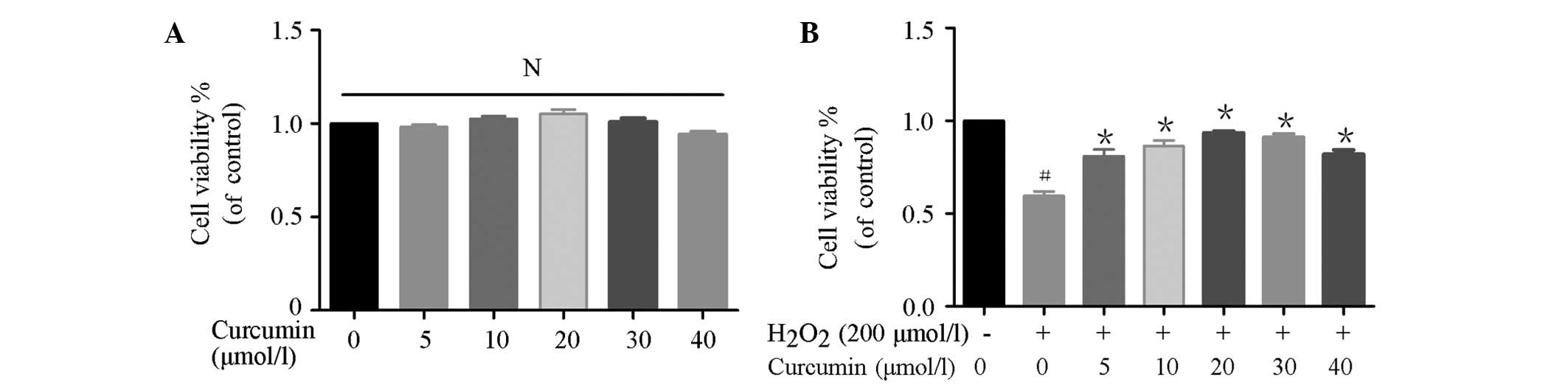
EA.hy926 cells were pretreated with curcumin (0, 5,
10, 20, 30 or 40 µmol/l) for 4 h, and were then co-incubated
with 200 µmol/l H2O2 for an additional
4 h, in order to determine the effects of curcumin on
H2O2-induced cell death. As shown in Fig. 1A, curcumin (0–40 µmol/l) did
not significantly affect viability of the EA.hy926 cells. In
addition, curcumin (5–40 µmol/l) prevented
H2O2-induced cytotoxicity (Fig. 1B).
Curcumin mitigates
H2O2-induced apoptosis in EA.hy926 cells
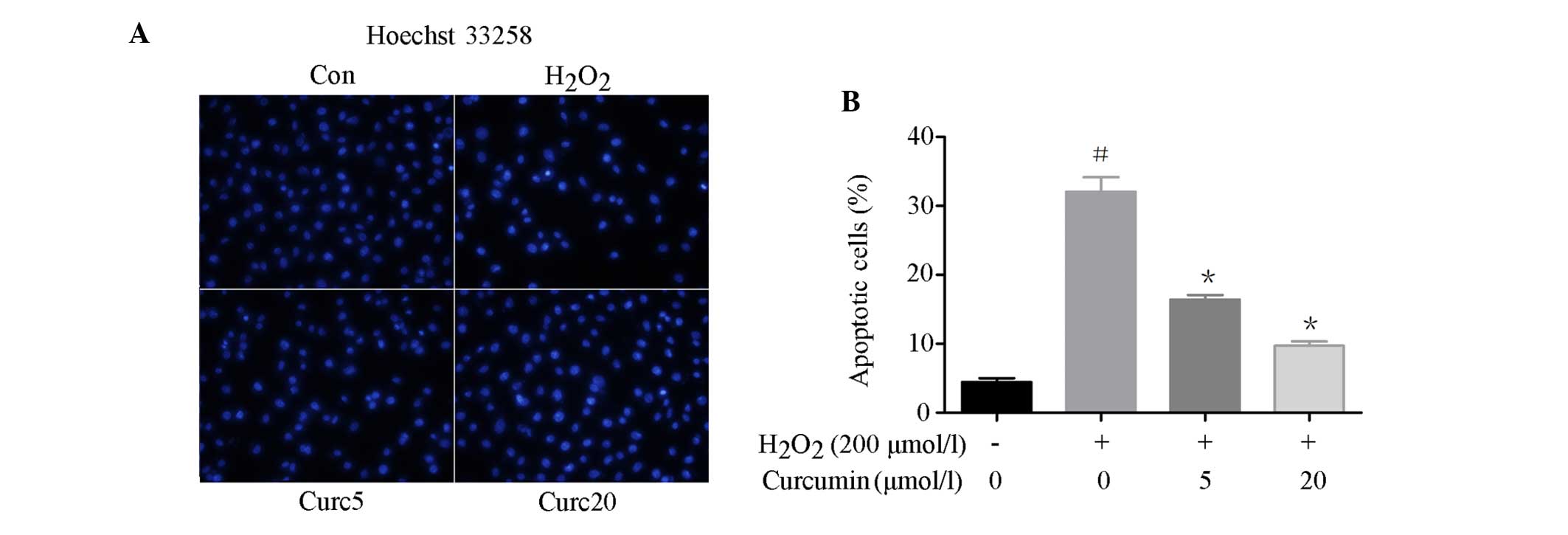
To examine the effects of curcumin on the apoptosis
of EA.hy926 cells, Hoechst 33258 staining was conducted and the
expression levels of apoptosis-associated proteins were detected
following curcumin pretreatment. Hoechst 33258 staining was used to
assess DNA fragmentation. As shown in Fig. 2A and B, the percentage of apoptotic
cells was significantly reduced in the curcumin-pretreated cells in
a concentration-dependent manner, as compared with the
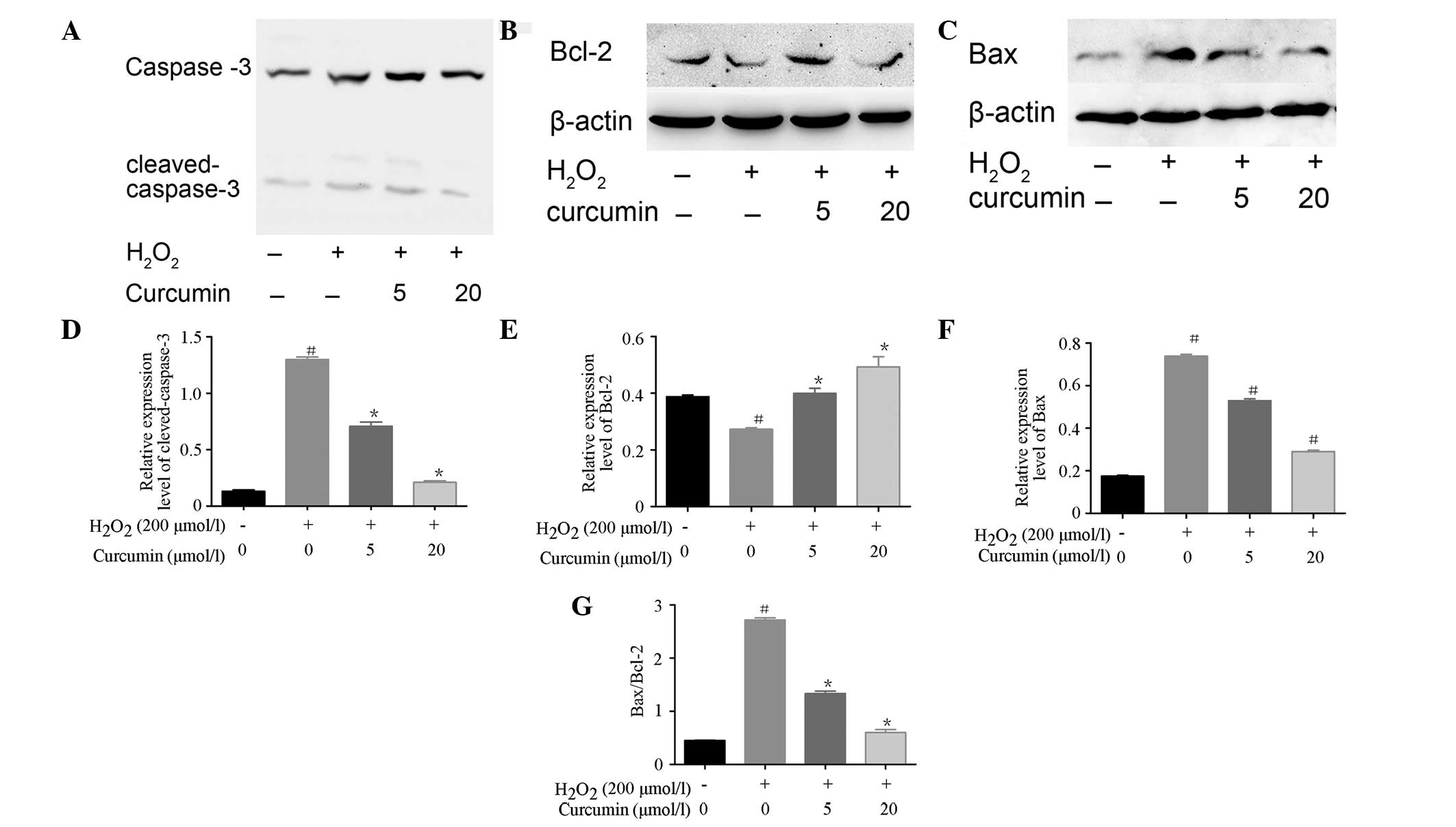
H2O2-treated cells. The expression levels of
apoptosis regulatory proteins, including caspase-3, Bax and Bcl-2
were detected, in order to confirm the anti-apoptotic effect of
curcumin on H2O2-induced cells. As shown in
Fig. 3A–G, the expression levels
of cleaved caspase-3 and Bax were increased following treatment
with H2O2, as compared with the control.
Curcumin pretreatment significantly decreased the expression levels
of H2O2-induced cleaved caspase-3 and Bax in
EA.hy926 cells. In addition, the expression levels of the
anti-apoptotic protein Bcl-2 were decreased following
H2O2 treatment, and were increased in a
concentration-dependent manner following curcumin pretreatment. The
increase in Bax/Bcl-2 ratio induced by H2O2
was inhibited by curcumin.
Curcumin promotes autophagy in
H2O2-treated EA.hy926 cells
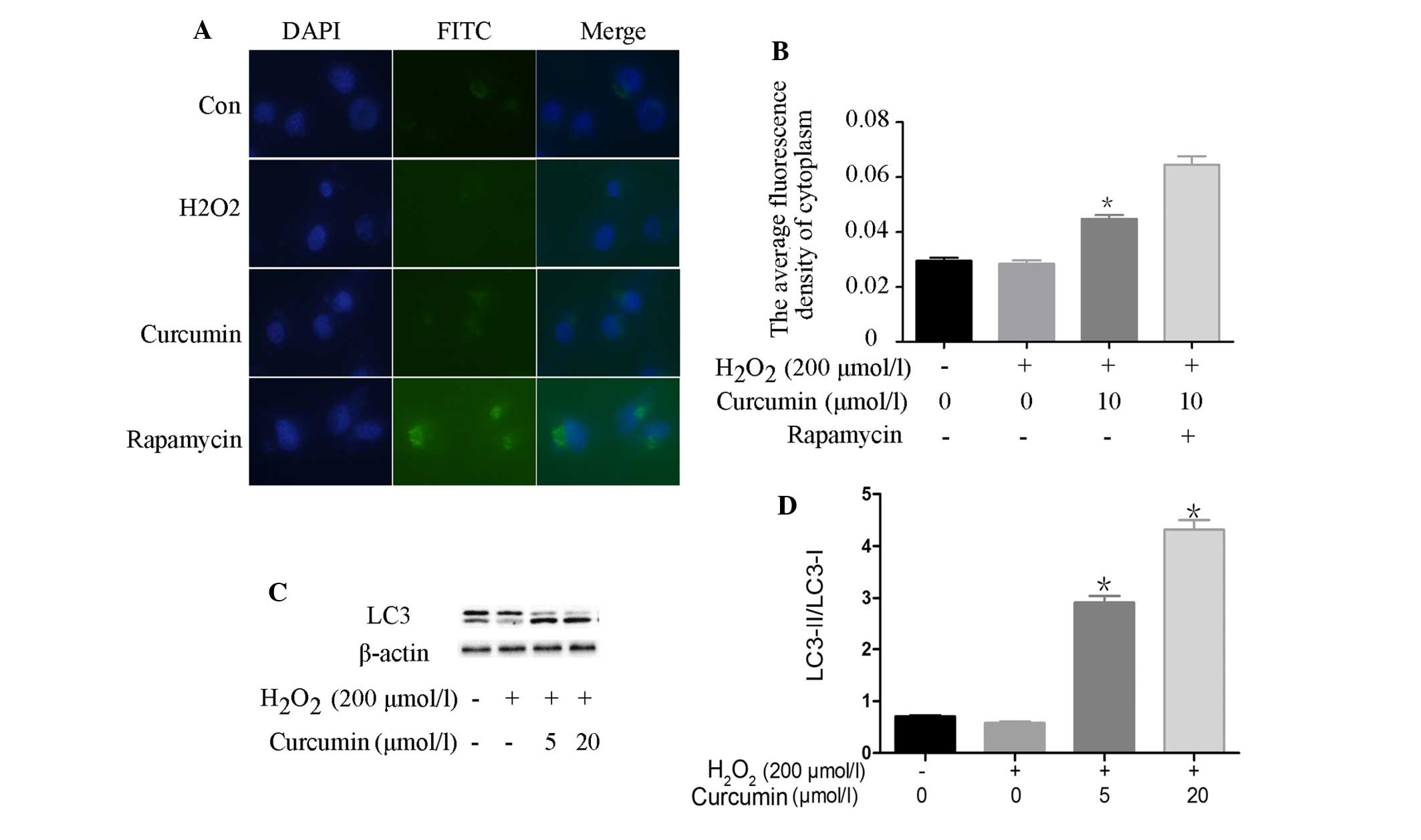
In order to detect autophagy and evaluate the extent
of autophagosome formation in H2O2-treated
cells, a FITC-labeled LC3 antibody from the CYTO-ID®
Autophagy Detection kit (Enzo Life Sciences, Inc.) was used. When
autophagy takes place in mammalian cells, LC3 content, particularly
LC3-II content, is significantly increased. LC3-II is considered an
appropriate marker of autophagy activation, since it is a vital
protein in autophagosome formation (16). As shown in Fig. 4A and B, as compared with the
control group, there was no significant difference in LC3-II
content in the H2O2-treated cells; however,
there was a significant increase in LC3 expression in the
rapamycin-treated cells, and a slight increase in expression in the
curcumin-pretreated cells. Lipidation of LC3 is essential for
autophagy to proceed. As shown in Fig.
4C and D, following an increase in autophagy, the protein
expression levels of LC3-II were significantly higher in the
curcumin-pretreated cells compared with in the control and
H2O2-treated cells. Furthermore, the results
of western blotting indicated that the protein expression levels of
LC3-II were significantly higher in the curcumin-pretreated cells,
as compared with in the control and
H2O2-treated cells.
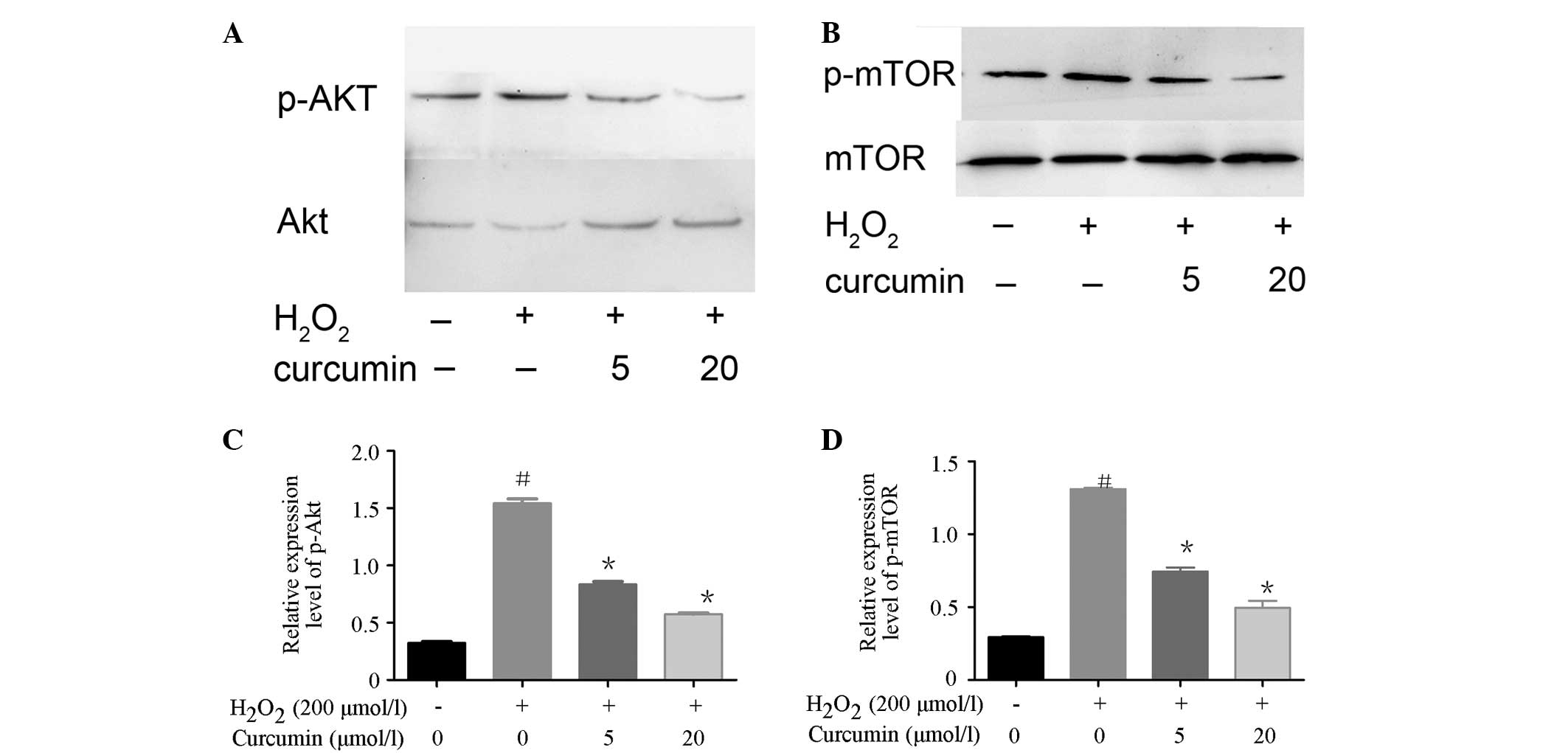
Curcumin downregulates the Akt/mTOR
signaling pathway in H2O2-treated EA.hy926
cells
To further evaluate the mechanisms and signaling
pathways underlying curcumin-induced autophagy, the activation
status of mTOR and its upstream regulator Akt were determined by
western blotting. As shown in Fig.
5, the expression levels of p-Akt and p-mTOR were increased in
the H2O2-treated cells. However, the
expression levels of p-Akt and p-mTOR were significantly decreased
in the curcumin-pretreated cells in a concentration-dependent
manner. These results indicate that curcumin was able to inhibit
the expression of the autophagy suppressor mTOR in EA.hy926 cells,
and this was associated with the cell-specific modulation of the
mTOR upstream regulator, Akt.
Discussion
The results of the present study demonstrated that
pretreatment with curcumin alleviated
H2O2-induced cytotoxicity in EA.hy926 cells.
In addition, pretreatment with curcumin inhibited oxidative
stress-induced apoptosis and activated adaptive autophagy in
EA.hy926 cells. These results indicated that the protective effects
of curcumin were dependent on regulation of the Akt/mTOR signaling
pathway.
The EA.hy926 human umbilical vein cell line is
produced by hybridizing human umbilical endothelial cells (HUVECs)
with the A549 epithelial cell line. EA.hy926 cells are widely used
as a replacement for HUVECs in in vitro experiments, and
H2O2 is widely used to generate models of
oxidative stress. Oxidative stress is capable of activating the
mitochondrial signaling pathway and has a pivotal role in the
pathogenesis of cardiovascular disease, including ischemic heart
disease, hypertension and heart failure (17). Therefore, the elimination of
excessive intracellular ROS and prevention of oxidative stress may
be an effective intervention for the treatment of these
diseases.
Curcumin, which is a major effective ingredient
extracted from a traditional Chinese herbal medicine, exerts potent
antioxidative effects and has been widely used to prevent and treat
cardiovascular disease, including atherosclerosis (18,19).
However, the mechanisms underlying the protective effects of
curcumin vary considerably between studies. Previously, Notch,
Toll-like receptor (TLR)2, ROS-relative TLR4-mitogen-activated
protein kinases/nuclear factor-κB, glycogen synthase kinase-3β,
peroxisome proliferator-activated receptor γ, and Sirtuin 1
(2,20–24)
have been reported to have a role in the protective effects of
curcumin on oxidative stress-injured endothelial cells. In our
previous study, curcumin was shown to mitigate
H2O2-induced myocardial damage by inhibiting
the expression of monocyte chemoattractant protein-1 (25); however, the role of autophagy in
this process has not been reported. To the best of our knowledge,
no studies have yet reported that curcumin activates adaptive
autophagy in oxidative stress-injured endothelial cells; however,
considerable evidence has demonstrated that curcumin induces cancer
cell apoptosis via autophagy activation (26–31).
Curcumin, or its analogue, have previously been reported to induce
cell death in colon cancer, uterine leiomyosarcoma, astrocytoma,
ovarian cancer, lung adenocarcinoma and cutaneous T-cell lymphoma
cells (26–31) by activating autophagy. In the
present study, curcumin attenuated
H2O2-induced cell death by activating
adaptive autophagy in EA.hy926 cells.
Autophagy and apoptosis are two closely regulated
pathways through which superfluous, damaged, or aged cells or
organelles are eliminated. In addition, autophagy is a process by
which cells adapt their metabolism to environmental or
intracellular stress conditions, including starvation, endoplasmic
reticulum stress, hypoxia, ischemia/reperfusion injury, pathogens
and oxidative stress. Autophagy generates metabolic substrates that
meet the bioenergetic needs of cells, and thereby allows for
adaptive protein synthesis by the catabolism of macromolecules
(32–34). Autophagy enables cells to adapt to
stress, in order to avoid cell death; however, autophagy is also an
alternative pathway that may lead to cell death (35–37).
Autophagy and apoptosis can be initiated in response to similar
stimuli, whereas in some situations autophagy and apoptosis are
initiated in a mutually exclusive manner. The relationship between
autophagy and apoptosis is complex. There are several examples in
which the induction of autophagy promotes the activation of
apoptosis; however, autophagy also arises from the inhibition of
apoptosis and protects cells from cell death (38), and sometimes autophagy inhibits the
induction of apoptosis. High dose H2O2 can
induce cellular damage due to oxidative stress. Oxidative stress
increases the permeability of the lysosomal membrane, and induces
the uncoupling of oxidation and phosphorylation reactions in the
mitochondria, resulting in the activation of various cell death
programs (39). At a low level of
oxidative stress, autophagy protects the cell against major harm by
degrading damaged mitochondria prior to cytochrome c release
(40). In the present study, a
significant decrease in cell viability was detected in the
H2O2-treated (200 µmol/l) EA.hy926
cells. In addition, the levels of apoptosis were increased, whereas
autophagy exhibited no significant change. When pretreated with
curcumin, H2O2-induced EA.hy926 cell death
was reduced, the rate of apoptosis was reduced and autophagy was
increased. Therefore, these results suggested that
H2O2 (200 µmol/l) was able to induce
EA.hy926 cell apoptosis but had no effect on autophagy, whereas
curcumin pretreatment attenuated apoptosis in
H2O2-treated EA.hy926 cells by activating
autophagy. These data demonstrated that curcumin may induce
autophagy, in order to protect endothelial cells.
The Bcl-2 protein family, which is comprised of
anti- and pro-apoptotic members, consists of important
mitochondrial regulators during cell apoptosis. As compared with
Bax, Bcl-2 blocks cytochrome c release, inhibits caspase
activity and suppresses cell apoptosis (41). Therefore, alterations to the
Bcl-2/Bax ratio influences apoptotic balance. In the present study,
curcumin significantly inhibited Bax expression, and increased
Bcl-2 expression in cells undergoing oxidative stress, resulting in
a reduced Bax/Bcl-2 ratio and increased cell viability.
The regulation of autophagy is complex and involves
numerous pathways. The mTOR pathway is the most extensively studied
network with regards to autophagy regulation due to its ability to
sense nutrient state, growth factor availability and stress
(42). Upstream of mTOR complex 1
(mTORC1) is the tuberous sclerosis complex (TSC)1-TSC2 inhibitory
complex, which functions as an upstream activator of mTOR. The
TSC1-TSC2 complex inactivates Rheb, which inhibits mTOR signaling,
leading to the subsequent activation of autophagy. mTORC1 is able
to directly sense amino acid concentration and energy state,
whereas the PI3K/Akt axis can sense growth factor status. mTOR
complex 2 is an inhibitor that decreases the extent of Akt-induced
mTORC1 activation (43). The
present study demonstrated that H2O2 induced
the phosphorylation of mTOR, and this activation was inhibited by
curcumin. These findings are consistent with the view that mTOR
regulates autophagy by controlling phosphorylation (44). Indeed, curcumin pretreatment led to
decreased Akt phosphorylation, which was associated with mTOR
inhibition and autophagy. These results indicated that Akt is
involved in curcumin-induced mTOR suppression and autophagy.
However, the underlying mechanisms of action require further
study.
In conclusion, the results of the present study
demonstrated that oxidative stress may promote cell death in
EA.hy926 cells, and pre-treatment with curcumin suppresses cell
death by inducing autophagy via regulation of the Akt/mTOR
pathway.
Acknowledgments
The present study was supported by grants from the
Fourth Period Project '333' of Jiangsu Province (grant no.
BRA2012207), the Priority Academic Program Development of Jiangsu
Higher Education Institutions (grant no. BL2012011) and the
National Natural Science Foundation of China (grant no.
81170102/H0203).
References
|
1
|
Kim YS, Ahn Y, Hong MH, Joo SY, Kim KH,
Sohn IS, Park HW, Hong YJ, Kim JH, Kim W, et al: Curcumin
attenuates inflammatory responses of TNF-alpha-stimulated human
endothelial cells. J Cardiovasc Pharmacol. 50:41–49. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lubbad A, Oriowo MA and Khan I: Curcumin
attenuates inflammation through inhibition of TLR-4 receptor in
experimental colitis. Mol Cell Biochem. 322:127–135. 2009.
View Article : Google Scholar
|
|
3
|
Miriyala S, Panchatcharam M and
Rengarajulu P: Cardioprotective effects of curcumin. Adv Exp Med
Biol. 595:359–377. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aggarwal BB and Harikumar KB: Potential
therapeutic effects of curcumin, the anti-inflammatory agent,
against neurodegenerative, cardiovascular, pulmonary, metabolic,
autoimmune and neoplastic diseases. Int J Biochem Cell Biol.
41:40–59. 2009. View Article : Google Scholar :
|
|
5
|
Nazam Ansari M, Bhandari U and Pillai KK:
Protective role of curcumin in myocardial oxidative damage induced
by isoproterenol in rats. Hum Exp Toxicol. 26:933–938. 2007.
View Article : Google Scholar
|
|
6
|
Naik SR, Thakare VN and Patil SR:
Protective effect of curcumin on experimentally induced
inflammation, hepatotoxicity and cardiotoxicity in rats: Evidence
of its antioxidant property. Exp Toxicol Pathol. 63:419–431. 2011.
View Article : Google Scholar
|
|
7
|
Zanchetti A, Hennig M, Hollweck R, Bond G,
Tang R, Cuspidi C, Parati G, Facchetti R and Mancia G: Baseline
values but not treatment-induced changes in carotid intimamedia
thickness predict incident cardiovascular events in treated
hypertensive patients: Findings in the European Lacidipine Study on
Atherosclerosis (ELSA). Circulation. 120:1084–1090. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Motterlini R, Foresti R, Bassi R and Green
CJ: Curcumin, an antioxidant and anti-inflammatory agent, induces
heme oxygenase-1 and protects endothelial cells against oxidative
stress. Free Radic Biol Med. 28:1303–1312. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Green DR: Apoptotic pathways: Ten minutes
to dead. Cell. 121:671–674. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takagi H, Matsui Y, Hirotani S, Sakoda H,
Asano T and Sadoshima J: AMPK mediates autophagy during myocardial
ischemia in vivo. Autophagy. 3:405–407. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao DJ, Wang ZV, Battiprolu PK, Jiang N,
Morales CR, Kong Y, Rothermel BA, Gillette TG and Hill JA: Histone
deacetylase (HDAC) inhibitors attenuate cardiac hypertrophy by
suppressing autophagy. Proc Natl Acad Sci USA. 108:4123–4128. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sinnberg T, Lasithiotakis K, Niessner H,
Schittek B, Flaherty KT, Kulms D, Maczey E, Campos M, Gogel J,
Garbe C and Meier F: Inhibition of PI3K-AKT-mTOR signaling
sensitizes melanoma cells to cisplatin and temozolomide. J Invest
Dermatol. 129:1500–1515. 2009. View Article : Google Scholar
|
|
14
|
Hietakangas V and Cohen SM: Regulation of
tissue growth through nutrient sensing. Annu Rev Genet. 43:389–410.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kamada Y, Yoshino K, Kondo C, Kawamata T,
Oshiro N, Yonezawa K and Ohsumi Y: Tor directly controls the Atg1
kinase complex to regulate autophagy. Mol Cell Biol. 30:1049–1058.
2010. View Article : Google Scholar :
|
|
16
|
Tanida I, Ueno T and Kominami E: LC3
conjugation system in mammalian autophagy. Int J Biochem Cell Biol.
36:2503–2518. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ong SB and Gustafsson AB: New roles for
mitochondria in cell death in the reperfused myocardium. Cardiovasc
Res. 94:190–196. 2012. View Article : Google Scholar :
|
|
18
|
Kowluru RA and Kanwar M: Effects of
curcumin on retinal oxidative stress and inflammation in diabetes.
Nutr Metab (Lond). 4:82007. View Article : Google Scholar
|
|
19
|
Quiles JL, Mesa MD, Ramírez-Tortosa CL,
Aguilera CM, Battino M, Gil Á and Ramírez-Tortosaet MC: Curcuma
longa extract supplementation reduces oxidative stress and
attenuates aortic fatty streak development in rabbits. Arterioscler
Thromb Vasc Biol. 22:1225–1231. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang Y, Duan W, Liang Z, Yi W, Yan J, Wang
N, Li Y, Chen W, Yu S, Jin Z and Yi D: Curcumin attenuates
endothelial cell oxidative stress injury through Notch signaling
inhibition. Cell Signal. 25:615–629. 2013. View Article : Google Scholar
|
|
21
|
Meng Z, Yan C, Deng Q, Gao DF and Niu XL:
Curcumin inhibits LPS-induced inflammation in rat vascular smooth
muscle cells in vitro via ROS-relative TLR4-MAPK/NF-κB pathways.
Acta Pharmacol Sin. 34:901–911. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chanoit G, Lee S, Xi J, Zhu M, Mclntosh
RA, Mueller RA and Xu Z: Exogenous zinc protects cardiac cells from
reperfusion injury by targeting mitochondrial permeability
transition pore through inactivation of glycogen syntheses
kinase-3beta. Am J Physiol Heart Circ Physiol. 295:H1227–H1233.
2008. View Article : Google Scholar
|
|
23
|
Blanquicett C, Kang BY, Ritzenthaler JD,
Jones DP and Hart CM: Oxidative stress modulates PPAR gamma in
vascular endothelial cells. Free Radic Biol Med. 48:1618–1625.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chung S, Yao H, Caito S, Hwang JW,
Arunachalam G and Rahman I: Regulation of SIRT1 in cellular
functions: Role of polyphenols. Arch Biochem Biophys. 501:79–90.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo SY and Long MZ: Effects of curcumin on
expression of MCP-1 induced by H2O2 in
myocardial cell. Nanjing Yi Ke Da Xue Xue Bao. 4:442–444. 2010.In
Chinese.
|
|
26
|
Basile V, Belluti S, Ferrari E, Gozzoli C,
Ganassi S, Quaglino D, Saladini M and Imbriano C:
bis-Dehydroxy-Curcumin triggers mitochondrial-associated cell death
in human colon cancer cells through ER-stress induced autophagy.
PLoS One. 8:e536642013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li B, Takeda T, Tsuiji K, Wong TF,
Tadakawa M, Kondo A, Nagase S and Yaegashi N: Curcumin induces
cross-regulation between autophagy and apoptosis in uterine
leiomyosarcoma cells. Int J Gynecol Cancer. 23:803–808. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Romero-Hernández MA, Eguía-Aguilar P,
Perézpeña-DiazConti M, Rodríguez-Leviz A, Sadowinski-Pine S,
Velasco-Rodríguez LA, Cáceres-Cortés JR and Arenas-Huertero F:
Toxic effects induced by curcumin in human astrocytoma cell lines.
Toxicol Mech Methods. 23:650–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qu W, Xiao J, Zhang H, Chen Q, Wang Z, Shi
H, Gong L, Chen J, Liu Y, Cao R and Lv J: B19, a novel monocarbonyl
analogue of curcumin, induces human ovarian cancer cell apoptosis
via activation of endoplasmic reticulum stress and the autophagy
signaling pathway. Int J Biol Sci. 9:766–777. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiao K, Jiang J, Guan C, Dong C, Wang G,
Bai L, Sun J, Hu C and Bai C: Curcumin induces autophagy via
activating the AMPK signaling pathway in lung adenocarcinoma cells.
J Pharmacol Sci. 123:102–109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yosifov DY, Kaloyanov KA, Guenova ML,
Prisadashka K, Balabanova MB, Berger MR and Konstantinov SM:
Alkylphosphocholines and curcumin induce programmed cell death in
cutaneous T-cell lymphoma cell lines. Leuk Res. 38:49–56. 2014.
View Article : Google Scholar
|
|
32
|
Rubinsztein DC, Gestwicki JE, Murphy LO
and Klionsky DJ: Potential therapeutic applications of autophagy.
Nat Rev Drug Discov. 6:304–312. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Meléndez A and Neufeld TP: The cell
biology of autophagy in metazoans: A developing story. Development.
135:2347–2360. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang Z and Klionsky DJ: Eaten alive: A
history of macroautophagy. Nat Cell Biol. 12:814–822. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Baehrecke EH: Autophagy: Dual roles in
life and death? Nat Rev Mol Cell Biol. 6:505–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mariño G, Niso-Santano M, Baehrecke EH and
Kroemer G: Self-consumption: The interplay of autophagy and
apoptosis. Nat Rev Mol Cell Biol. 15:81–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kroemer G and Jäättelä M: Lysosomes and
autophagy in cell death control. Nat Rev Cancer. 5:886–897. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lum JJ, DeBerardinis RJ and Thompson CB:
Autophagy in metazoans: Cell survival in the land of plenty. Nat
Rev Mol Cell Biol. 6:439–448. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kubota C, Tor S, Hou N, Saito N, Yoshimoto
Y, Imai H and Takeuchi T: Constitutive reactive oxygen species
generation from autophagosome/lysosome in neuronal oxidative
toxicity. J Biol Chem. 285:667–674. 2010. View Article : Google Scholar :
|
|
40
|
Kiffin R, Bandyopadhyay U and Cuervo AM:
Oxidative stress and autophagy. Antioxid Redox Signal. 8:152–162.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fischer U and Schulze-Osthoff K: New
approaches and therapeutics targeting apoptosis in disease.
Pharmacol Rev. 57:187–215. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jung CH, Ro SH, Cao J, Otto NM and Kim DH:
mTOR regulation of autophagy. FEBS Lett. 584:1287–1295. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hay N and Sonenberg N: Upstream and
downstream of mTOR. Genes Dev. 18:1926–1945. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Alers S, Löffler AS, Wesselborg S and
Stork B: Role of AMPK-mTOR-Ulk1/2 in the regulation of autophagy:
Cross talk, shortcuts, and feedbacks. Mol Cell Biol. 32:2–11. 2012.
View Article : Google Scholar :
|