Introduction
Diffuse large B-cell lymphoma (DLBCL) is the most
common subtype of adult non-Hodgkin's lymphoma, a group of highly
invasive and heterogeneous cancers, accounting for 30–40% of cases
(1). According to the immune
phenotype, DLBCL is divided into three subtypes, namely, germinal
center B cell-like subtype (GCB), activated B cell-like subtype
(ABC), and type III diffuse large B-cell lymphoma (2). Immunological therapy using rituximab,
a monoclonal antibody against B cell surface protein cluster of
differentiation (CD)20 (3),
combined with traditional therapeutic strategies, have notably
improved the rate of complete remission and disease-free survival
in patients with DLBCL (4,5). However, a marked number of patients
are resistant to these advanced therapies for reasons that remain
to be elucidated (6,7). Previous studies have suggested that
the prognosis of GCB subtype is better compared with that of the
other two subtypes (8,9).
Omacetaxine mepesuccinate is a plant alkaloid
extracted from the total alkaloids of Cephalotaxaceae.
Omacetaxine mepesuccinate has been used as an antitumor therapeutic
agent to treat acute myelogenous leukemia (AML), chronic
myelogenous leukemia (CML) and myelodysplastic syndrome (10–12).
Furthermore, previous studies have reported that omacetaxine
mepesuccinate may be useful in treating lymphoma (13–15);
however, it remains to be elucidated how omacetaxine mepesuccinate
exerts its therapeutic effects on this type of cancer. The present
study examined the cellular effect of omacetaxine mepesuccinate on
two human DLBCL cell lines and demonstrated that omacetaxine
mepesuccinate induces apoptosis and regulates cell cycling,
differentiation and telomerase activity in the ABC and GCB subtype
of human DLBCL cells. Notably, the efficacy of omacetaxine
mepesuccinate was higher in the ABC compared with in the GCB
subtype. The present study provides evidence regarding the
development of omacetaxine mepesuccinate into a potential
therapeutic agent for the treatment of DLBCL.
Materials and methods
Cell lines and reagents
Human DLBCL cell lines SU-DHL-4 (SU-4; GCB subtype)
and OCI-LY3 (LY3; ABC subtype) were provided by the Shanghai
Institute of Hematology, Ruijin Hospital (Shanghai, China). These
cell lines together with Kasumi-1 human AML cell line, K562 human
CML cell line, MCF-7 human breast cancer cell line, and SGC-7901
human gastric cancer cell line (provided by Tianjin Institute of
Hematology, Tianjin, China) were maintained for use in the present
study. SU-DHL-4, OCI-LY3, K562, SGC-7901 and Kasumi-1 cells were
cultured in RPMI 1640 medium (Hyclone; GE Healthcare Life Sciences,
Chalfont, UK or Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher
Scientific., Inc.). MCF-7 cells were cultured in Dulbecco's
modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% FBS. These cultures were maintained at 37°C in a
humidified incubator supplied with 5% CO2.
Omacetaxine mepesuccinate (Hangzhou Minsheng
Pharmaceutical Group Co., Ltd., Hangzhou, China) was dissolved at a
concentration of 4 ng/µl in culture medium without FBS
according to the cell type and maintained at 4°C prior to use.
Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit
and Cell Cycle Detection kit were purchased from BestBio Company
(Shanghai, China). Telomeric repeat amplification protocol
(TRAP)-silver staining telomerase detection kit was purchased from
Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China). CD19
allophycocyanin (APC) antibodies (cat. no. 302211), CD138
phycoerythrin (PE) antibodies (cat. no. 352305), immunoglobulin
(Ig) D FITC antibodies (cat. no. 348205), IgM APC antibodies (cat.
no. 314509) and mouse IgG antibodies (cat. no. 409305) were
purchased from BioLegend, Inc. (San Diego, CA, USA). Protein
standards were purchased from Biomed Mechnikov (Moscow, Russia) and
an RNA inhibitor was purchased from Promega Corporation (Madison,
WI, USA).
Omacetaxine mepesuccinate treatment
To determine whether omacetaxine
mepesuccinate-induced apoptosis was dose-dependent, LY3 cells and
SU-4 cells were incubated with omacetaxine mepesuccinate at various
concentrations (5, 10, 20, 40 and 100 ng/ml), or with a vehicle (0
ng/ml) serving as a control, for 48 h. To determine the
time-dependent effects of omacetaxine mepesuccinate, LY3 and SU-4
cells were incubated with 40 ng/ml omacetaxine mepesuccinate for 0,
4, 8, 24, 48 or 72 h. To compare the differences between the
various cancer cells, MCF-7, SGC-7901, Kasumi-1 and K562 cells were
incubated with 40 ng/ml omacetaxine mepesuccinate for 0, 24 or 48
h. For cellular morphological analysis, LY3 and SU-4 cells were
treated with 40 ng/ml omacetaxine mepesuccinate for 0 or 24 h. To
determine the stage of the cell cycle, cell differentiation status,
and telomerase activity, LY3 cells and SU-4 cells were exposed to
20 ng/ml omacetaxine mepesuccinate for 0, 4, 8, 12, 24 and 48 h. To
determine cell differentiation status, LY3 cells and SU-4 cells
were exposed to 20 ng/ml omacetaxine mepesuccinate for 0, 12, 24
and 48 h. All experiments were repeated at least three times.
Cell apoptosis analysis
Cells were plated in 12-well plates at a density of
5×105 cells/well and exposed to omacetaxine
mepesuccinate or a vehicle for the designated period of time. Cells
were then collected and apoptosis analysis was conducted using
Annexin V-FITC Apoptosis Detection kit, according to the
manufacturer's protocol. In apoptotic cells, the phospholipid
phosphatidylserine (PS) is translocated from the inner to the outer
surface of the plasma membrane. While exposed to the external
cellular space, PS is labeled by FITC-conjugated Annexin V, which
binds PS with high affinity. Annexin V/FITC staining is used in
conjunction with propidium iodide (PI), which is a vital dye that
permeates damaged membranes of dead cells but is excluded by the
intact membrane of healthy cells, for identification of early and
late apoptotic cells. Viable cells are Annexin V and PI-negative,
whereas early apoptotic cells are Annexin V-positive and
PI-negative, and late apoptotic or dead cells are Annexin V and
PI-positive. The number of cells with single or double staining was
counted using a FACSCalibur™ flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). For cell morphological analysis, images
of the cells were captured using a microscope (BX51; Olympus
Corporation, Tokyo, Japan) following conventional Wright staining
(Shanghai Hengyuan Biological Technology Co., Ltd., Shanghai,
China).
Cell cycle detection
Cells were plated at 5×105 cells/well in
12-well plates. Each well contained ~2 ml culture medium. In each
well, omacetaxine mepesuccinate was first diluted in 10 µl
culture medium and then added into the well. Cultures were
incubated at 37°C in an atmosphere containing 5% CO2
until experimentation. Cells were collected and prepared for cell
cycle detection using the Cell Cycle Detection kit, according to
manufacturer's protocol. Cells were counted using a FACSCalibur™
flow cytometer. FCS Express 4 Plus Research version 4.0 (De Novo
Software, Los Angeles, CA, USA) was used to analyze flow cytometry
results.
Cell surface antigen detection
Cells were cultured and treated with omacetaxine
mepesuccinate as described previously. The cells were collected,
washed twice with cold 1X phosphate-buffered saline (PBS), and were
adjusted to a concentration of 3–6×104 cells/10
µl. For each sample, four 10 µl aliquots were taken.
One aliquot was used as a blank control, whereas the other three
aliquots were incubated with CD19 APC/CD138 PE, IgD FITC/IgM APC,
or mouse IgG antibodies (all diluted 1:200). The four aliquots were
incubated at 4°C for 30 min in the dark. Cells were subsequently
washed twice with cold PBS and subjected to analysis using a
FACS-Calibur™ flow cytometer.
Cell telomerase activity measurement
Cells were maintained and treated with omacetaxine
mepesuccinate as described previously, until ready for
experimentation. Telomerase activity was measured using the
TRAP-silver staining telomerase detection kit, according to the
manufacturer's protocol. Briefly, pellets were resuspended in
ice-cold lysis buffer containing 200 U/ml RNase inhibitor, 0.1 mM
benzamidine and 10 mM β-mercaptoethanol, and incubated on ice for
30 min with gentle rocking. After centrifugation at 12,000 × g for
30 min, the supernatant containing telomerase was collected,
aliquoted into a small volume to avoid freeze-thaw cycles, measured
for total protein concentration and stored at −70°C. The protein
concentration was adjusted to 10–750 ng/µl prior to
telomerase extension and PCR amplification. A master mix was
prepared in an RNase-free PCR tube by mixing the following: 39.5
µl diethylpyrocarbonate H2O, 5 µl 10X TRAP
buffer, 1 µl dNTP, 1 µl TS primer, 1 µl TRAP
primer mix, 0.5 µl Taq-DNA polymerase and 2 µl
telomerase extract. PCR amplification was performed using the
following thermal cycling conditions: 30°C For 30 min (telomerase
extension reaction), 95°C for 5 min, and 30–33 cycles at 94°C for
30 sec and 59°C for 30 sec. The PCR products then underwent
polymerase chain reaction amplification and separation by 14%
non-denaturing polyacrylamide gel electrophoresis. Immediately
after the completion of gel electrophoresis, the gel was placed in
500 ml fixative solution containing 10% ethanol and 0.5% acetic
acid in deionized water for 10–20 min. The gel was then transferred
into staining solution containing 0.2% silver nitrate, 10% ethanol
and 0.5% acetic acid, and stained for 20–30 min. Following rinsing
with water for 5–10 sec, the gel was developed for 15–30 min,
rinsed in water for 5–10 min and photographed for analysis.
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Statistical analyses were conducted using SPSS 17.0
(SPSS, Inc., Chicago, IL, USA). Statistical significance was
determined by Student's t-test for pairwise comparisons or by
analysis of variance (ANOVA) with Bonferroni's multiple comparisons
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Induction of apoptosis by omacetaxine
mepesuccinate
Apoptotic cells were detected by Annexin V/FITC and
PI double staining. Early apoptotic cells were labeled with Annexin
V-FITC but not PI, whereas late apoptotic or dead cells were
indicated by double staining with Annexin V-FITC and PI. The number
of apoptotic cells was detected using flow cytometry. The number of
total apoptotic cells was calculated as the sum of early apoptotic
cells and late apoptotic or dead cells. As presented in Fig. 1, LY3 and SU-4 cells exhibited
increased rates of apoptosis following treatment with omacetaxine
mepesuccinate. Increased apoptosis was particularly noticeable in
the cells treated with omacetaxine mepesuccinate at 20 ng/ml or
higher for 24 h, in which the apoptotic rates were >60%
(Fig. 1A). Notably, the apoptotic
rates were significantly higher (P<0.004 and P<0.044,
respectively) in LY3 cells compared with in SU-4 cells following
exposure to 5 or 10 ng/ml omacetaxine mepesuccinate for 24 h
(Fig. 1A). Consistently, the
apoptotic rates were significantly higher in LY3 cells compared
with in SU-4 cells when these cells were treated with 40 ng/ml
omacetaxine mepesuccinate for 8 and 24 h (P<0.007 and
P<0.013, respectively; Fig.
1B). These results suggest that omacetaxine mepesuccinate
induces apoptosis in DLBCL cells in a dose- and time-dependent
manner, and that ABC subtype cells (LY3) are more vulnerable than
GCB subtype cells (SU-4).
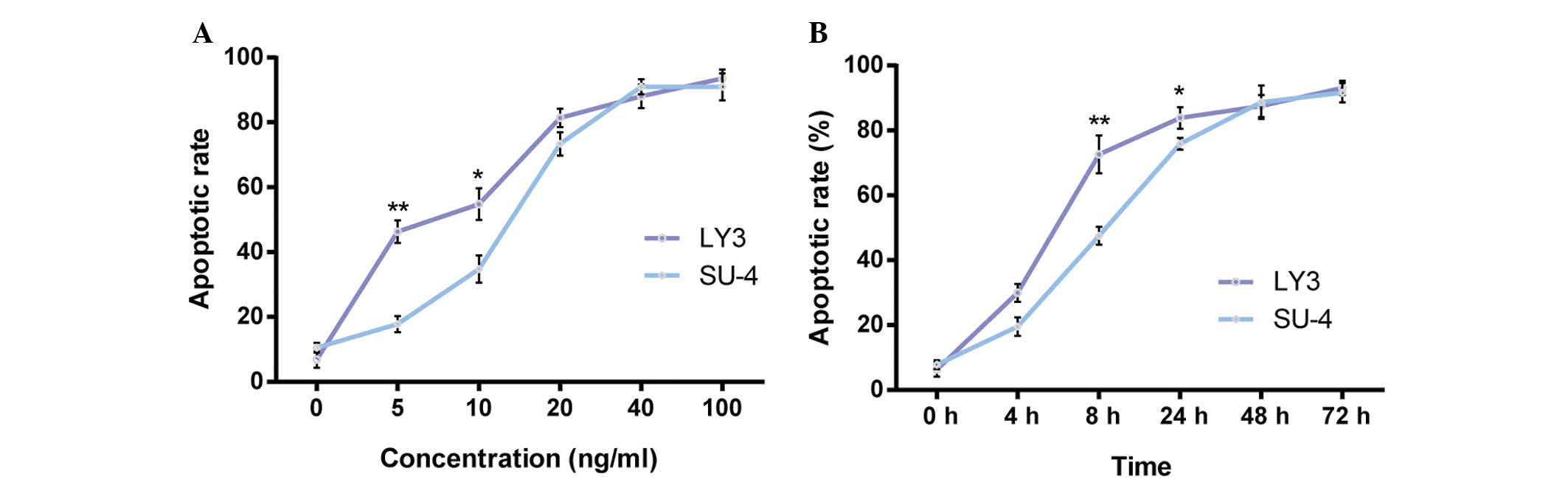 | Figure 1Omacetaxine mepesuccinate induced
apoptosis in a dose- and time-dependent manner in DLBCL cells. (A)
Graph presenting increased apoptotic rates in the LY3 and SU-4
DLBCL cells treated with omacetaxine mepesuccinate at various
concentrations (0, 5, 10, 20, 40, or 100 ng/ml) for 24 h. (B) Graph
presenting increased apoptotic rates in the two types of cells
exposed to 40 ng/ml omacetaxine mepesuccinate for 0, 4, 8, 24, 48,
or 72 h. The data are presented as the mean ± standard error of the
mean and were analyzed using Student's t-test; n=3.
*P<0.05, **P<0.01 vs. SU-4 cells. LY3,
OCI-LY3; SU-4, SU-DHL-4; DLBCL, diffuse large B-cell lymphoma. |
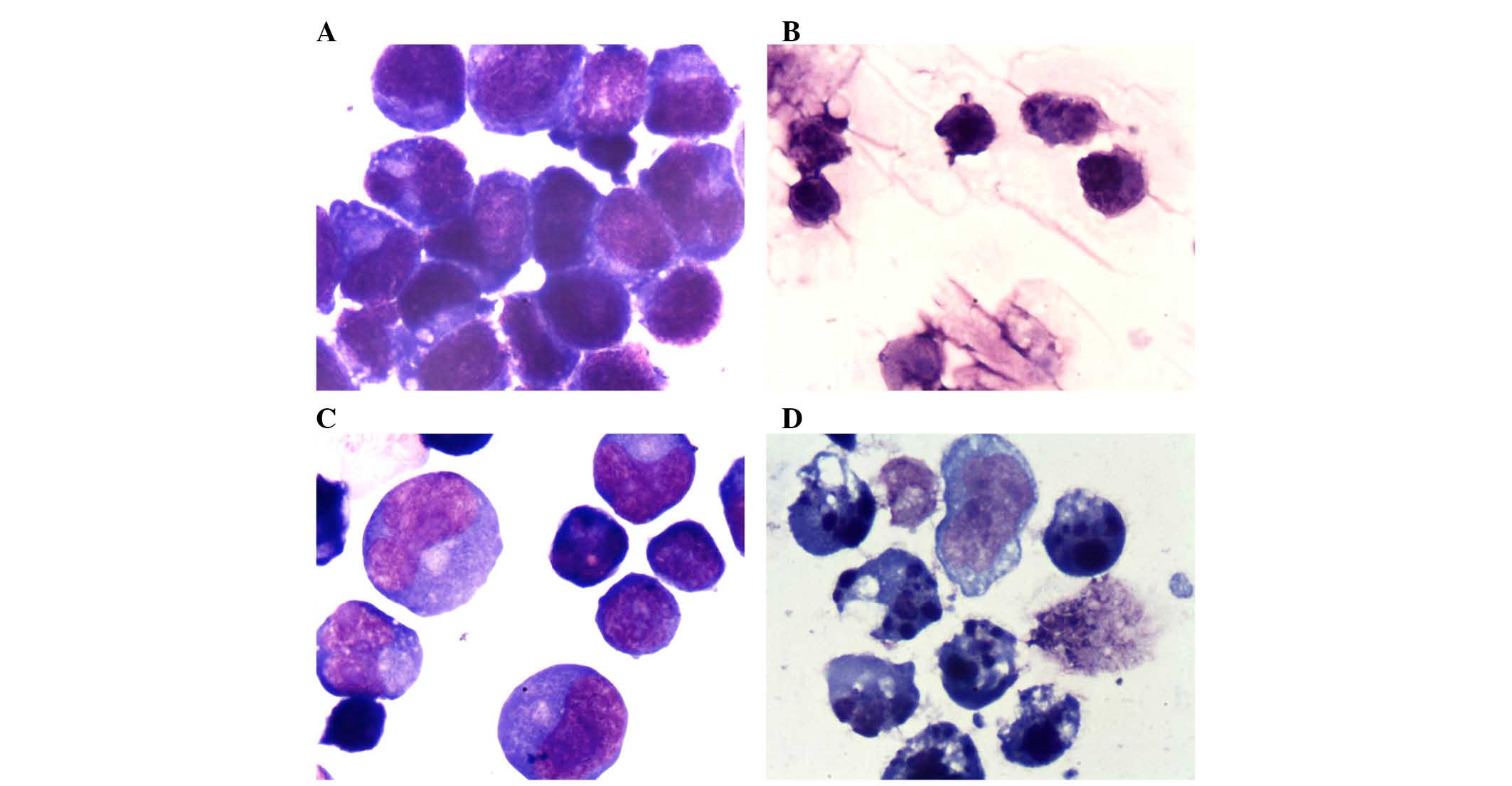
Consistent with omacetaxine mepesuccinate-mediated
apoptotic effects, cell morphology was altered following exposure
to 40 ng/ml omacetaxine mepesuccinate for 24 h. Compared with
vehicle treatment, omacetaxine mepesuccinate induced shrinkage,
karyopyknosis and an increased number of apoptotic bodies (Fig. 2). A greater number of
morphologically irregular apoptotic cells were observed in LY3
cells than in SU-4 cells (data not shown), which is consistent with
the flow cytometry results demonstrating that the ABC subtype
exhibits increased sensitivity to omacetaxine mepesuccinate than
the GCB subtype.
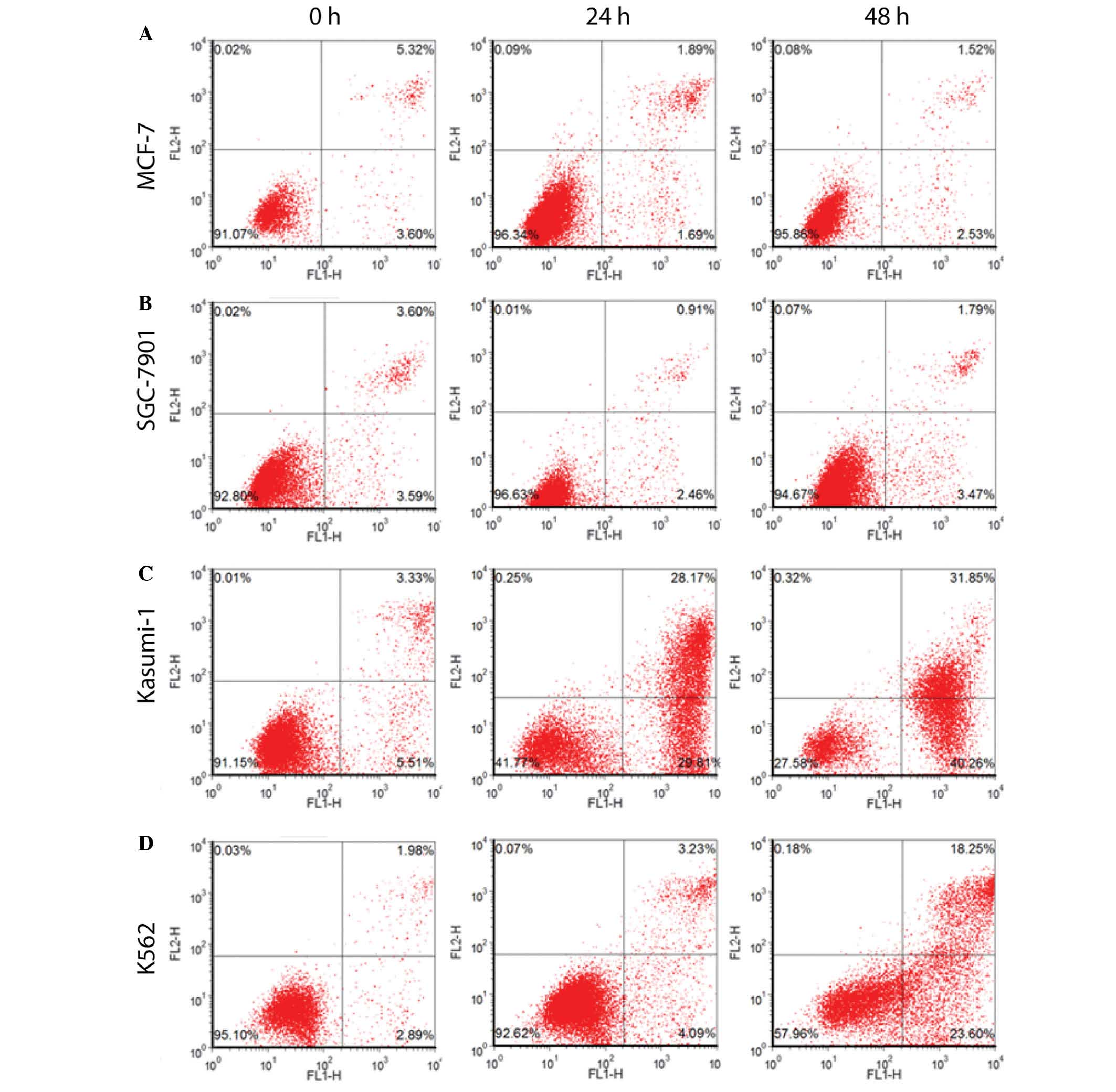
To determine whether omacetaxine mepesuccinate
results in a different apoptotic effect in different types of
cancer, omacetaxine mepesuccinate (40 ng/ml) was administered to
different cell lines, including MCF-7 human breast cancer cell
line, SGC-7901 human gastric cancer cell line, Kasumi-1 AML cell
line, and K562 human CML cells for 0, 24 or 48 h. As presented in
Table I and Fig. 3, omacetaxine mepesuccinate
application resulted in a significant increase in the apoptotic
rates of Kasumi-1 AML cells and K562 human CML cells, but not in
the MCF-7 human breast cancer and SGC-7901 human gastric cancer
cells. These results indicate that omacetaxine
mepesuccinate-induced apoptosis is specific to the type of
tumor.
 | Table IOmacetaxine mepesuccinate-induced
apoptosis in various cancer cell lines. |
Table I
Omacetaxine mepesuccinate-induced
apoptosis in various cancer cell lines.
| Cell line | Time (h)
|
|---|
0
| 24
| 48
|
|---|
| Apoptotic rate | P-value | Apoptotic rate | P-value | Apoptotic rate | P-value |
|---|
| MCF-7 | 6.4967±2.3099 | 0.934 | 4.3200±0.8007 | 0.001 | 4.6500±1.1805 | 0.001 |
| SGC-7901 | 6.4667±0.6623 | 0.915 | 4.6567±1.1621 | 0.001 | 5.9833±0.7514 | 0.001 |
| Kasumi-1 | 7.3267±1.3540 | 0.657 | 59.4867±1.4009 | 0.011 | 72.9900±3.8071 | 0.071 |
| K562 | 4.5867±1.0151 | 0.305 | 9.0333±1.5047 | 0.001 | 46.0767±3.7792 | 0.009 |
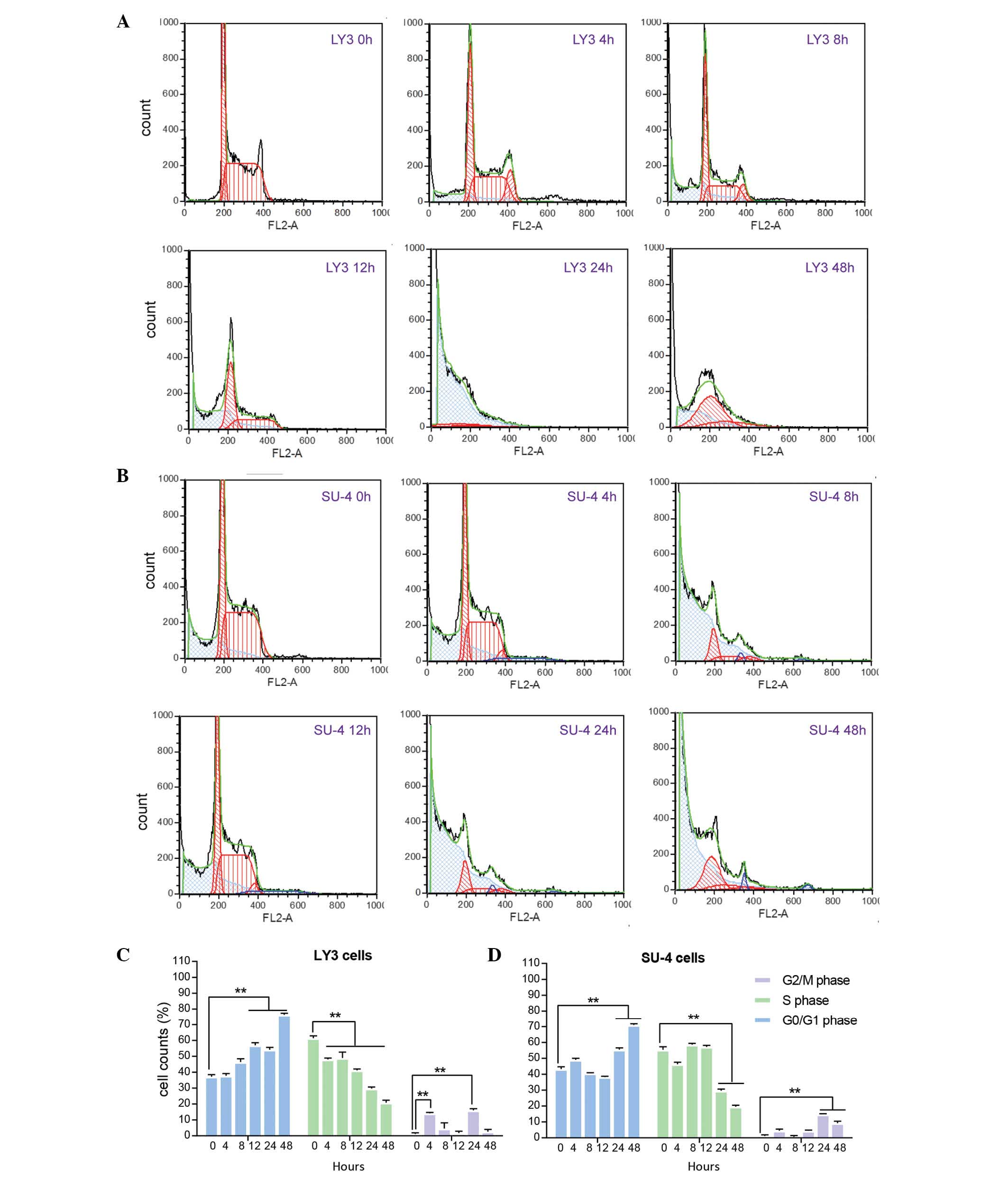
Cell cycle detection
The present study demonstrated that cell cycle
distribution of LY3 and SU-4 cells was altered by omacetaxine
mepesuccinate exposure for a selected period of time. LY3 and SU-4
cells exposed to omacetaxine mepesuccinate demonstrated a typical
subdiploid apoptotic peak prior to G0/G1
phase (Fig. 4A and B). Prior to
omacetaxine mepesuccinate treatment, there were more cells in S
phase than in G0/G1 phase in the LY3 and SU-4
cells (Fig. 4C and D). Upon
omacetaxine mepesuccinate exposure, the majority of LY3 and SU-4
cells were in G0/G1 phase. Following
treatment with omacetaxine mepesuccinate for 24 h or longer, more
than half of the cells were arrested in G0/G1
phase in the two cell lines (Fig. 4C
and D).
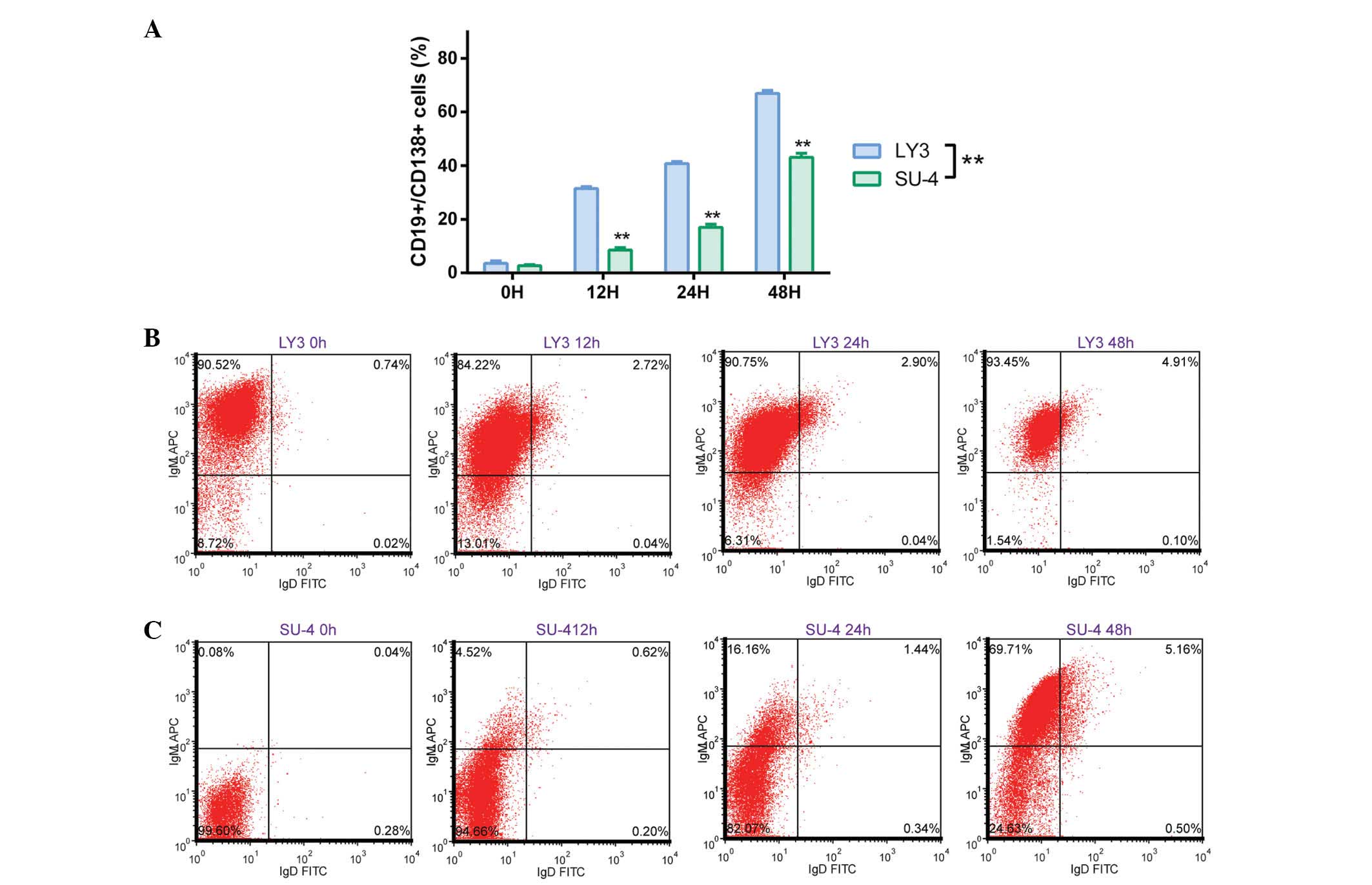
Cell surface antigen detection
The effect of omacetaxine mepesuccinate on cell
differentiation in DLBCL cells was investigated by detecting
changes in the expression of B lymphocyte antigen, CD19 and plasma
cell antigen, CD138. As presented in Fig. 5A, the percentage of
CD19+/CD138+ cells increased gradually in the
LY3 and SU-4 cells following treatment with omacetaxine
mepesuccinate. Omacetaxine mepesuccinate-mediated differentiation
into B lymphocytes was more effective in LY3 cells than in SU-4
cells (P<0.01, according to two-way ANOVA). Similarly, upon
omacetaxine mepesuccinate exposure, the percentage of LY3 cells
expressing IgM and IgD (IgM+/IgD+, mature B
lymphocytes) increased steadily, whereas most of the untreated LY3
control cells expressed IgM but not IgD
(IgM+/IgD−; Fig.
5B). Conversely, the majority of untreated SU-4 control cells
were IgM−/IgD−. However, the percentage of
IgM+ SU-4 cells was significantly increased (P<0.01
according to one-way ANOVA; data not shown) upon omacetaxine
mepesuccinate exposure (Fig. 5C).
These results demonstrate that omacetaxine mepesuccinate
application in DLBCL cells promotes cell differentiation and
maturation.
Omacetaxine mepesuccinate reduces cell
telomerase activity in DLBCL cells
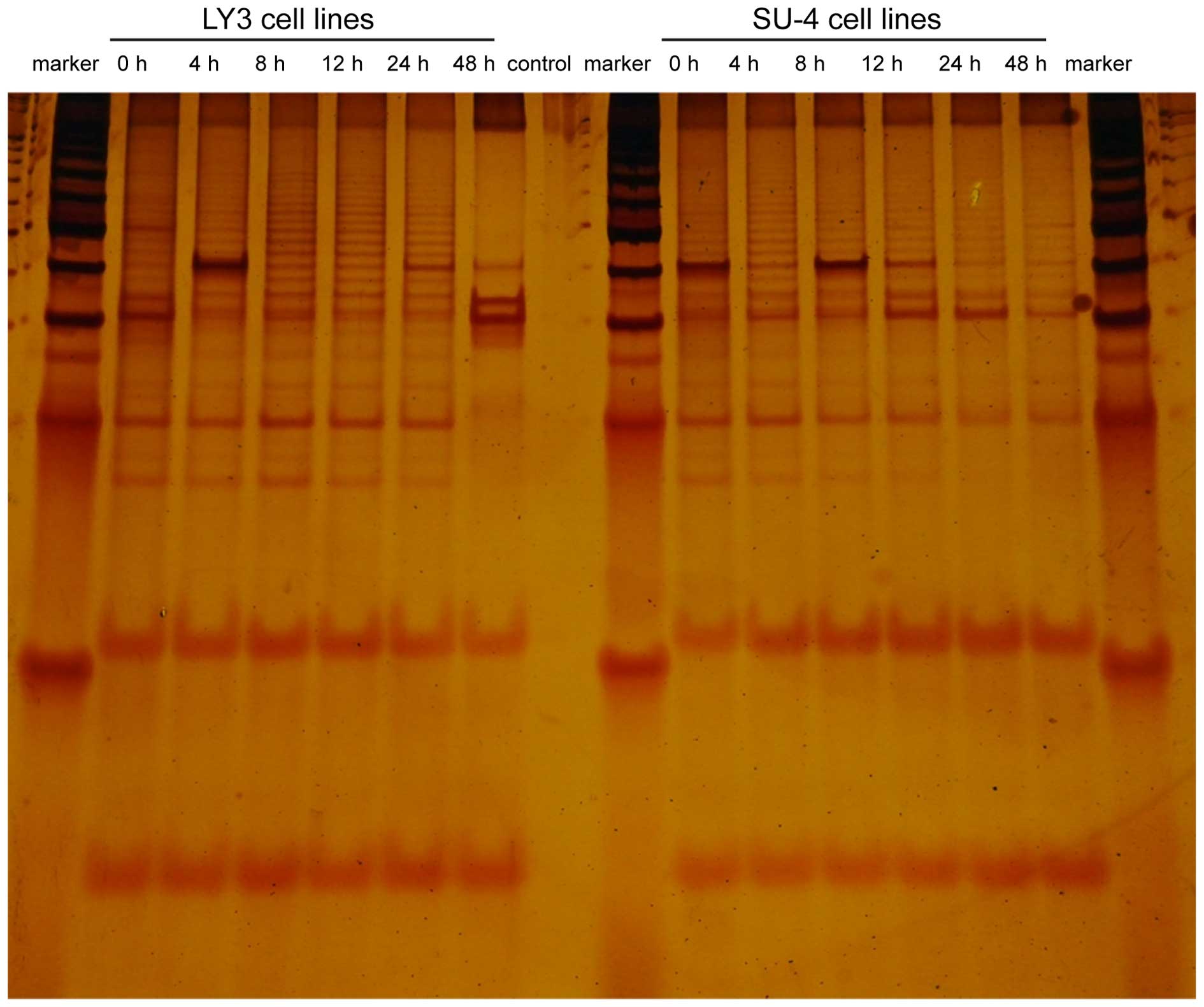
LY3 cells and SU-4 cells were incubated in 20 ng/ml
omacetaxine mepesuccinate for 4, 8, 12, 24 or 48 h, and were then
subjected to measurement of telomerase activity using TRAP-silver
staining. DNA strips indicated a 6 bp ladder gap in non-denaturing
polyacrylamide gel electrophoresis, where the number and depth of
the DNA strips represented telomerase activity. Untreated control
cells (0 h) served as positive controls. As presented in Fig. 6, telomerase activity in the LY3 and
SU-4 cells was not significantly altered following treatment with
omacetaxine mepesuccinate for 4, 8, 12 and 24 h compared with the
activity of the control. However, the two types of cells
demonstrated a marked decrease in telomerase activity following
exposure to omacetaxine mepesuccinate for 48 h. This change is
particularly clear in LY3 cells, in which telomerase activity was
almost completely absent.
Discussion
It has previously been demonstrated that omacetaxine
mepesuccinate results in the apoptosis of various types of cancer
cells, including leukemia cells (16,17),
certain types of solid carcinoma cells (18–21),
and selected types of lymphocytes (13–15).
However, omacetaxine mepesuccinate-mediated cell death has not yet,
to the best of our knowledge, been reported in DLBCL cells. The
present study is the first, to the best of our knowledge, to
indicate that omacetaxine mepesuccinate induces apoptosis in DLBCL
cells in a dose- and time-dependent manner. The ABC subtype was
demonstrated to be more sensitive to omacetaxine
mepesuccinate-induced apoptosis than the GCB subtype. In addition,
omacetaxine mepesuccinate was shown to induce cell cycle arrest,
promote cell differentiation and maturation, and reduce telomerase
activity. Consistent with a previous study (18), omacetaxine mepesuccinate
application also induced apoptosis in K562 cells (CML) and Kasumi-1
cells (AML), but not in MCF-7 cells (breast cancer) or SGC-7901
cells (gastric cancer). These results suggested that omacetaxine
mepesuccinate-mediated apoptosis is cancer type-specific.
An efficient anticancer therapeutic agent is often
evaluated for its ability to induce apoptosis and cell cycle
arrest. It has been demonstrated that omacetaxine mepesuccinate
increases the expression levels of cyclin-dependent kinase (CDK)
inhibitors, p27 and p21, which in turn bind to CDK or cyclin/CDK
complexes to inhibit enzymatic activity, leading to the arrest of
the cell cycle at G1 phase (22,23).
Consistently, the present study observed that more than half of
DLBCL cells were arrested at G0/G1 phase
following Omacetaxine mepesuccinate application for 24 and 48 h.
Accordingly, the percentage of cells at S phase and G2/M
phase was significantly decreased. Consistent with this result,
omacetaxine mepesuccinate promotes cell differentiation into mature
B lymphocytes, as indicated by the increased number of cells
expressing CD19/CD138 in addition to IgD/IgM, which are signs of
cell differentiation and maturation of B lymphocytes.
Telomeres are considered the biological clock of
cell ageing and cell life span/survival (24,25).
Telomerase activity, which caps the ends of chromosomes to
facilitate chromosome duplication and cell division, is inactivated
or undetectable in normal somatic cells but present in germ cells
with proliferative potential, embryonic stem cells and certain
lymphocyte cells. However, during cancer development, telomerase
activity is aberrantly increased, which allows cancer cells to
divide continuously. Increased telomerase activity has been
detected in human colon, lung, liver and breast cancer, in addition
to leukemia and lymphoma (26–29).
Therefore, telomerase may be used as a tumor marker, and may be
investigated as a target for anticancer therapeutic agents in
somatic tissues (30–32). Reducing telomerase activity may
result in cell senescence, inhibition of cell proliferation, and
programmed cell death, suggesting it may be an effective approach
to treat cancer (33–37). Consistent with the results of a
previous study (38), the present
study demonstrated that the GCB and ABC subtype DLBCL cells
exhibited high telomerase activity, as demonstrated using
TRAP-silver staining. In addition, the present study demonstrated
that the increased telomerase activity was suppressed by
omacetaxine mepesuccinate application for 24 and 48 h, and that
omacetaxine mepesuccinate-mediated inhibition is more effective in
ABC subtype cells, as telomerase activity in LY3 cells was reduced
to close to nothing upon omacetaxine mepesuccinate exposure for 48
h. Notably, the results of the present study indicated that
omacetaxine mepesuccinate-induced apoptosis occurred earlier than
omacetaxine mepesuccinate-mediated telomerase activity reduction,
which has also been reported by previously published studies
(29,39).
In conclusion, omacetaxine mepesuccinate induces
apoptosis in DLBCL cells in a dose- and time-dependent manner. The
effect of omacetaxine mepesuccinate is more effective in ABC
subtype (LY3 cells) than in GCB subtype (SU-4 cells) cells.
Furthermore, omacetaxine mepesuccinate was able to arrest the cell
cycle, promote cell differentiation/maturation, and reduce
telomerase activity. The findings of the present study may provide
valuable insight into the molecular mechanism underlying
omacetaxine mepesuccinate-mediated apoptosis. Since it is already
in use for the treatment of AML and CML, omacetaxine mepesuccinate
may be further investigated for its therapeutic effects in
DLBCL.
Abbreviations:
|
Omacetaxine mepesuccinate
|
homoharringtonine
|
|
DLBCL
|
diffuse large B-cell lymphoma
|
|
GCB
|
germinal center B cell-like
subtype
|
|
ABC
|
activated B cell-like subtype
|
|
AML
|
acute myelogenous leukemia
|
|
CML
|
chronic myelogenous leukemia
|
|
SU-4
|
SU-DHL-4
|
|
LY3
|
OCI-LY3
|
|
CD19
|
B-lymphocyte antigen CD19
|
|
CDK
|
cyclin-dependent kinase
|
References
|
1
|
Cultrera JL and Dalia SM: Diffuse large
B-cell lymphoma: Current strategies and future directions. Cancer
Control. 19:204–213. 2012.PubMed/NCBI
|
|
2
|
Rosenwald A, Wright G, Chan WC, Connors
JM, Campo E, Fisher RI, Gascoyne RD, Muller-Hermelink HK, Smeland
EB, Giltnane JM, et al Lymphoma/Leukemia Molecular Profiling
Project: The use of molecular profiling to predict survival after
chemotherapy for diffuse large-B-cell lymphoma. N Engl J Med.
346:1937–1947. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grillo-López AJ, White CA, Dallaire BK,
Varns CL, Shen CD, Wei A, Leonard JE, McClure A, Weaver R, Cairelli
S and Rosenberg J: Rituximab: The first monoclonal antibody
approved for the treatment of lymphoma. Curr Pharm Biotechnol.
1:1–9. 2000. View Article : Google Scholar
|
|
4
|
Pfreundschuh M, Trümper L, Osterborg A,
Pettengell R, Trneny M, Imrie K, Ma D, Gill D, Walewski J, Zinzani
PL, et al MabThera International Trial Group: CHOP-like
chemotherapy plus rituximab versus CHOP-like chemotherapy alone in
young patients with good-prognosis diffuse large-B-cell lymphoma: A
randomised controlled trial by the MabThera International Trial
(MInT) Group. Lancet Oncol. 7:379–391. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martelli M, Ferreri AJ, Agostinelli C, Di
Rocco A, Pfreundschuh M and Pileri SA: Diffuse large B-cell
lymphoma. Crit Rev Oncol Hematol. 87:146–171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Colomo L, López-Guillermo A, Perales M,
Rives S, Martínez A, Bosch F, Colomer D, Falini B, Montserrat E and
Campo E: Clinical impact of the differentiation profile assessed by
immunophenotyping in patients with diffuse large B-cell lymphoma.
Blood. 101:78–84. 2003. View Article : Google Scholar
|
|
7
|
Meyer PN, Fu K, Greiner TC, Smith LM,
Delabie J, Gascoyne RD, Ott G, Rosenwald A, Braziel RM, Campo E, et
al: Immunohistochemical methods for predicting cell of origin and
survival in patients with diffuse large B-cell lymphoma treated
with rituximab. J Clin Oncol. 29:200–207. 2011. View Article : Google Scholar :
|
|
8
|
Saad AA, Awed NM, Abdel-Hafeez ZM, Kamal
GM, Elsallaly HM and Alloub AI: Prognostic value of
immunohistochemical classification of diffuse large B-cell lymphoma
into germinal center B-cell and non-germinal center B-cell
subtypes. Saudi Med J. 31:135–141. 2010.PubMed/NCBI
|
|
9
|
Li M, Liu CL, Yin WJ, He YX, Xue XM, Duan
ZJ and Gao ZF: The clinical significance of a new classification
algorithm in Chinese DLBCL cases. Zhonghua Xue Ye Xue Za Zhi.
33:801–804. 2012.In Chinese.
|
|
10
|
Zhou XJ, Zhou YH, Chen XH and Qian WB:
Homoharringtonine combined arsenic trioxide induced apoptosis in
human multiple myeloma cell line RPMI 8226: An experimental
research. Zhongguo Zhong Xi Yi Jie He Za Zhi. 33:834–839. 2013.In
Chinese. PubMed/NCBI
|
|
11
|
Daver N, Vega-Ruiz A, Kantarjian HM,
Estrov Z, Ferrajoli A, Kornblau S, Verstovsek S, Garcia-Manero G
and Cortes JE: A phase II open-label study of the intravenous
administration of homoharringtonine in the treatment of
myelodysplastic syndrome. Eur J Cancer Care (Engl). 22:605–611.
2013. View Article : Google Scholar
|
|
12
|
Cao H, Cheng Y, You L, Qian J and Qian W:
Homohar-ringtonine and SAHA synergistically enhance apoptosis in
human acute myeloid leukemia cells through upregulation of TRAIL
and death receptors. Mol Med Rep. 7:1838–1844. 2013.PubMed/NCBI
|
|
13
|
Chen R, Guo L, Chen Y, Jiang Y, Wierda WG
and Plunkett W: Homoharringtonine reduced Mcl-1 expression and
induced apoptosis in chronic lymphocytic leukemia. Blood.
117:156–164. 2011. View Article : Google Scholar :
|
|
14
|
Cai Z, Lin M, Ludwig WD and Karawajew L:
Involvement of mitochondrial membrane potential in the
homoharringtonine induced apoptosis of leukemic T-cells. Zhonghua
Xue Ye Xue Za Zhi. 22:238–240. 2001.In Chinese.
|
|
15
|
Wang L and Jin J: Study on inhibition of
telomerase activity of human T lymphocyte Jurkat cells by
homoharringtonine and its mechanism. Shiyong Zhongliu Zazhi.
20:391–394. 2005.In Chinese.
|
|
16
|
Li YF, Liu X, Liu DS, Din BH and Zhu JB:
The effect of homoharringtonine in patients with chronic myeloid
leukemia who have failed or responded suboptimally to imatinib
therapy. Leuk Lymphoma. 50:1889–1891. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xie WZ, Lin MF, Huang H and Cai Z:
Homoharringtonine-induced apoptosis of human leukemia HL-60 cells
is associated with downregulation of telomerase. Am J Chin Med.
34:233–244. 2006. View Article : Google Scholar
|
|
18
|
Huang HJ, He XZ and Jiao F: Induction of
apoptosis on SGC-7901 cells by harringtonine. Dongnan Daxue Xuebao
(Yixueban). 32:206–209. 2013.In Chinese.
|
|
19
|
Liu X and Ji YB: Study on anticancer
effects of homoharringtonine to HepG2 by MTT in vitro. J Harbin
Univ Commer (Nat Sci Ed. 28:393–395. 2012.
|
|
20
|
Jin Y, Lu Z, Cao K, Zhu Y, Chen Q, Zhu F,
Qian C and Pan J: The antitumor activity of homoharringtonine
against human mast cells harboring the KIT D816V mutation. Mol
Cancer Ther. 9:211–223. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Beranova L, Pombinho AR, Spegarova J, Koc
M, Klanova M, Molinsky J, Klener P, Bartunek P and Andera L: The
plant alkaloid and anti-leukemia drug homoharringtonine sensitizes
resistant human colorectal carcinoma cells to TRAIL-induced
apoptosis via multiple mechanisms. Apoptosis. 18:739–750. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Belletti B, Nicoloso MS, Schiappacassi M,
Chimienti E, Berton S, Lovat F, Colombatti A and Baldassarre G: p27
(kip1) functional regulation in human cancer: A potential target
for therapeutic designs. Curr Med Chem. 12:1589–1605. 2005.
View Article : Google Scholar
|
|
23
|
Liu JN, Bi GF, Wen PE, Yang WH, Ren X,
Tang TH, Xie C, Dong W, Jiang GS and Lin RX: Study on variation of
CD44 expression and its role in differentiation of HL-60 cells
induced by HHT. Chinese Journal of Cancer Prevention and Treatment.
15:1361–1364. 2008.
|
|
24
|
Mengual Gómez DL, Armando RG, Farina HG
and Gómez DE: Telomerase and telomere: Their structure and dynamics
in health and disease. Medicina (B Aires). 74:69–76. 2014.In
Spanish.
|
|
25
|
Autexier C and Lue NF: The structure and
function of telomerase reverse transcriptase. Annu Rev Biochem.
75:493–517. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu YF, Zhang Y, Shen N, Zhang RY and Lu
XQ: Effect of VEGF, P53 and telomerase on angiogenesis of gastric
carcinoma tissue. Asian Pac J Trop Med. 7:293–296. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu D, Wang Q, Gruber A, Björkholm M, Chen
Z, Zaid A, Selivanova G, Peterson C, Wiman KG and Pisa P:
Downregulation of telomerase reverse transcriptase mRNA expression
by wild type p53 in human tumor cells. Oncogene. 19:5123–5133.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sakurai S, Fukayama M, Kaizaki Y, Saito K,
Kanazawa K, Kitamura M, Iwasaki Y, Hishima T, Hayashi Y and Koike
M: Telomerase activity in gastrointestinal stromal tumors. Cancer.
83:2060–2066. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Akiyama M, Yamada O, Kanda N, Akita S,
Kawano T, Ohno T, Mizoguchi H, Eto Y, Anderson KC and Yamada H:
Telomerase overexpression in K562 leukemia cells protects against
apoptosis by serum deprivation and double-stranded DNA break
inducing agents, but not against DNA synthesis inhibitors. Cancer
Lett. 178:187–197. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo M and Wang J: Advances in study on
telomerase of esophageal carcinoma. Chinese Journal of
Gastroenterology. 18:122–124. 2013.
|
|
31
|
Lin G, Chen Q, Yu S, Lin S, Yao H, Ding Z,
Chen S, Lin MC and Wang X: Overexpression of human telomerase
reverse transcriptase C-terminal polypeptide sensitizes HeLa cells
to 5-fluorouracilinduced growth inhibition and apoptosis. Mol Med
Rep. 9:279–284. 2014.
|
|
32
|
Park ES, Lee J, Kang SY, Lee EJ, Lee MH,
Yoon N, Oh YL and Kim KM: A comparative study of telomerase
activity and cytologic diagnosis in malignant ascites. Anal Quant
Cytopathol Histpathol. 35:146–151. 2013.PubMed/NCBI
|
|
33
|
Li HJ, Wang JM, Tian YT, Bai ML, Zhang LX
and Zhao XX: Effect of matrine on Fas, VEGF, and activities of
telomerase of MCF-7 cells. Zhongguo Zhong Xi Yi Jie He Za Zhi.
33:1247–1251. 2013.In Chinese. PubMed/NCBI
|
|
34
|
Lin G, Chen Q, Yu S, Lin S, Yao H, Ding Z,
Chen S, Lin MC and Wang X: Overexpression of human telomerase
reverse transcriptase C-terminal polypeptide sensitizes HeLa cells
to 5-fluorouracilinduced growth inhibition and apoptosis. Mol Med
Rep. 9:279–284. 2014.
|
|
35
|
Tomizawa A, Kanno SI, Osanai Y, Yomogida S
and Ishikawa M: Cytotoxic effects of caffeic acid undecyl ester are
involved in the inhibition of telomerase activity in NALM-6 human
B-cell leukemia cells. Oncol Lett. 6:875–877. 2013.PubMed/NCBI
|
|
36
|
Halacli SO, Canpinar H, Cimen E and
Sunguroglu A: Effects of gamma irradiation on cell cycle, apoptosis
and telomerase activity in p53 wild-type and deficient HCT116 colon
cancer cell lines. Oncol Lett. 6:807–810. 2013.PubMed/NCBI
|
|
37
|
Nakashima M, Nandakumar J, Sullivan KD,
Espinosa JM and Cech TR: Inhibition of telomerase recruitment and
cancer cell death. J Biol Chem. 288:33171–33180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Norrback KF, Enblad G, Erlanson M,
Sundström C and Roos G: Telomerase activity in Hodgkin's disease.
Blood. 92:567–573. 1998.PubMed/NCBI
|
|
39
|
Armitage JO and Weisenburger DD: New
approach to classifying non-Hodgkin's lymphomas: Clinical features
of the major histologic subtypes. Non-Hodgkin's lymphoma
classification project. J Clin Oncol. 16:2780–2795. 1998.PubMed/NCBI
|