Introduction
An allergic response is characterized by an
exaggerated immune reaction to certain antigens, including harmless
environmental antigens from various sources, such as food and
pollen. Approximately 25% of the population in developed countries
suffers from immunoglobulin (Ig)E-mediated type I allergy (1,2).
Interleukin (IL)-1β has been identified as an important cytokine,
which has a key role in the pathophysi-ology of allergic disorders
(3). Furthermore, IL-1β has an
essential role in the immune response to infectious agents
(4).
IL-1β can be produced by immune cells, including
blood monocytes, tissue macrophages and dendritic cells (5). IL-1β secretion is a two-step process:
Firstly, interaction of Toll-like receptors (TLRs) with their
ligands leads to an upregulation of pro-IL-1β gene transcription
via nuclear factor-κB (6).
Secondly, caspase-1 activation results in the conversion of the
immature pro-IL-1β into mature IL-1β. It is well-established that
caspase-1 activity is regulated by a cyto-solic multi-protein
complex known as the inflammasome (7). The inflammasome consists of a
nucleotide-binding domain like receptor (NLR), one or more adaptor
proteins, such as apoptosis-associated speck-like protein (ASC),
and caspase-1 (8). Phylogenetic
analysis of NLR family NACHT domains identified three distinct
subfamilies: The nucleotide-binding oligomerization domain (NOD)
subfamily [NOD1-2, NOD3/NLR family, CARD domain containing (NLRC)3,
NOD4/NLRC5, NOD5/NLRX1, and class II, major histocompatibility
complex, transactivator], the NLR family, pyrin domain containing
(NLRP) subfamily (NLRP1-14, also called NALPs), and the IL-1β
converting enzyme protease-activating factor (IPAF) subfamily,
which consists of IPAF (also known as NLRC4) and NLR family,
apoptosis inhibitory protein (NAIP) (9).
The role of the inflammasome is essential in the
maturation of IL-1β. Elevated local or systemic levels of IL-1β,
associated with inappropriate inflammasome activity, have been
reported in various diseases, particularly in allergic disorders
(10–12). Furthermore, previous studies have
reported that NLRP3, NLRP12 and absent in melanoma (AIM)2 are
involved in different models of allergy (13–15).
In addition, NLRP3 has been introduced as an important inflammasome
in contact hypersensitivity (16)
and allergic airway diseases (14,17).
Arthur et al (15)
demonstrated that the loss of NLRP12 protects against allergy in a
mouse model of contact hypersensitivity. In addition, activation of
AIM2 by DNA leads to production of IL-1β and IL-18 in patients with
psoriasis (18).
At present, inflammasome activation in allergic
diseases remains a controversial issue, and further studies are
required to elucidate the exact role of inflammasomes in the
context of allergy. Therefore, the present study was designed to
investigate which inflammasome pathway is associated with caspase-1
activation and IL-1β production in an animal model of allergy.
Materials and methods
Materials and mice
Recombinant (r)Che a 2, a major allergen in
Chenopodium album pollen, was expressed and purified from
Escherichia coli BL21 (DE3) by metal affinity
chromatography, as described in our previous study (19). The alum gel suspension was
purchased from Sigma-Aldrich (St. Louis, MO, USA). Biotinylated
monoclonal rat anti-mouse IgE antibody (cat. no 1130-08) was
purchased from SouthernBiotech, Inc. (Birmingham, AL, USA).
Horseradish peroxidase (HRP)-conjugated streptavidin was obtained
from Bio-Rad Laboratories, Inc. (Hercules, CA, USA). Female BALB/c
mice (age, 6–8 weeks) were purchased from the Razi Vaccine and
Serum Research Institute (Mashhad, Iran). The mice were housed at
21±2°C and a 12 h light-dark cycle was maintained. They were fed
with standard rodent pellet and had free access to drinking water.
Animals were maintained in the animal house according to the local
guidelines for animal care, and experiments were approved by the
Animal Ethics Committee of Babol University of Medical Sciences
(Babol, Iran).
Induction and evaluation of allergy in
BALB/c mice
Following 1 week of acclimation to the animal house
environment, the sensitization protocol was conducted. Five BALB/c
mice were sensitized by intraperitoneal (IP) injection of rChe a 2
(5 µg) adsorbed onto 2 mg alum [Al(OH)3] gel
suspension (13 mg/ml) on days 0, 7, 14 and 21. The mice in the
control group (n=5) received 20 mM phosphate-buffered saline (PBS)
absorbed onto 2 mg alum gel suspension (IP), without rChe a 2, at
the same time points (20).
Subsequently, the sensitization procedure was continued with a 20
min exposure to an aerosol challenge of 1% rChe a 2 between days 26
and 28, using an Omron CX3 nebulizer (OMRON Healthcare Europe B.V.,
Hoofddorp, Netherlands). A total of 1 week following application of
the final aerosol challenge, sensitization was complete.
Subsequently, the mice were anesthetized using ketamine and
xylazine (90 mg/kg and 10 mg/kg respectively, Sigma-Aldrich) and
blood samples were collected through cardiac puncture. Next, the
mice were scarified by cervical dislocation. In addition, weekly
blood sampling via the retro-orbital plexus was conducted during
sensitization, mice were anesthetized as aforementioned. The
collected blood samples were left for 45 min at room temperature
and then were centrifuged at 1,300 x g for 20 min at 4°C. The
obtained serum was transferred to microtubes and stored at
−80°C.
Assessment of IgE and IL-1β levels
Specific IgE reactivity in serum samples was
measured using an indirect enzyme-linked immunosorbent assay
(ELISA). Briefly, microplate wells (Nunc MaxiSorp™; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) were coated with rChe a 2 (20
µg/ml) and were incubated at 4°C overnight. Blocking was
performed using 2% bovine serum albumin (BSA; Sigma-Aldrich), and
the microplate was washed with PBS with 0.05% Tween 20 (v/v). Mouse
serum samples were diluted at 1:5 in 1% BSA and were incubated in
the microplate overnight at 4°C. Subsequently, 1:5,000 biotinylated
rat anti-mouse IgE antibody was added and incubated at 37°C for 1
h. The microplate was then incubated with HRP-conjugated
streptavidin for 1 h. Finally,
tetramethylbenzidine/H2O2 was added to each
well and color development was terminated with 3 M HCl. Optical
density was measured using an ELISA microplate reader (Stat
Fax® 2100 Microplate Reader; Awareness Technology, Inc.,
Palm City, FL, USA). At the end of the experiment, lungs were
removed and tissues were homogenized. Subsequently, IL-1β levels in
the tissue lysates were measured using the RayBio® Mouse
IL-1 beta ELISA kit (RayBiotech, Inc., Norcross, GA, USA) following
the manufacturer's protocol.
RNA extraction and cDNA synthesis
Total RNA was isolated from the lung tissue
homogenates using TriPure Isolation Reagent (Roche Diagnostics
GmbH, Mannheim, Germany), according to the manufacturer's protocol.
DNA contamination from 10 µg RNA was removed with 2 U DNase
I according to the manufacturer's protocol (Sigma-Aldrich). Total
RNA was reverse transcribed using the Easy™ cDNA Reverse
Transcription kit with oligo (dT)16 and random hexamers
as amplification primers (Pars Tous Biotechnology, Mashhad, Iran).
Incubation at 25°C and 50°C was performed for 10 and 60 min,
respectively. Finally, the reaction was stopped by heating to 70°C
for 10 min.
Semi-quantitative reverse transcription
polymerase chain reaction (RT-PCR)
Specific oligonucleotide primers for the detection
of genes involved in the inflammasome pathway, including NLRP3,
ASC, IPAF and NAIP5, were designed by Primer Premier 5 software
(PREMIER Biosoft, Palo Alto, CA, USA), according to data deposited
on the Ensembl database (http://www.ensembl.org). Primer sequences are listed
in Table I. Semi-quantitative
RT-PCR was performed using the specific primers (Bioneer
Corporation Daejeon, South Korea). The total reaction volume was 20
µl, containing 2 µl cDNA, 12.5pmol of each primer,
1.5 Mm of MgCl2, 125µM dNTPs, 2 µl 10x
reaction buffer, and 2.0 U of Taq polymerase (Bioneer Corporation).
The cycling conditions were as follows: Initial denaturation at
95°C for 5 min, then 34 cycles of denaturation at 95°C for 30 sec,
annealing temperatures for all genes were at 59°C for 45 sec,
extension for 45 sec at 72°C and the final extension for 5 min at
72°C. The PCR was performed using a peqSTAR 2X Thermocycler (Peqlab
Biotechnologie GmbH, Erlangen, Germany). Subsequently, expression
levels of the genes of interest were normalized to hypoxanthine
phosphoribosyl transferase (HPRT) expression, which was used as an
internal control. PCR products were electrophoresed on 2% agarose
gels, and the subsequent bands were visualized using ethidium
bromide (SinaClon, Tehran, Iran) staining and were documented using
G:BOX (XT4; Syngene UK, Cambridge, UK). The intensity of the
obtained bands was determined using ImageJ software (version 1.49;
National Institutes of Health, Bethesda, MD, USA).
 | Table IPrimer sequences used to evaluate the
gene expression of inflammasome pathway-associated components. |
Table I
Primer sequences used to evaluate the
gene expression of inflammasome pathway-associated components.
| Genes | Primers | Product size
(bp) |
|---|
| ASC | F:
GCAACTGCGAGAAGGCTATG | 311 |
| R:
AAGCATCCAGCACTCCGTC | |
| NALP3 | F:
GCTAAGAAGGACCAGCCAGAGT | 180 |
| R:
GAACCTGCTTCTCACATGTCGT | |
| IPAF | F:
TTACTGTGAGCCCTTGGAGCA | 395 |
| R:
TGCCAGACTCGCCTTCAATC | |
| NAIP5 | F:
TTCACATCGAGAAGTTATCCATCCA | 304 |
| R:
AGCCTGGGCAAACTTTTCTGAC | |
| HPRT | F:
CGTCGTGATTAGCGATGATGAAC | 609 |
| R:
TCACTAATGACACAAACGTGATTC | |
Caspase-1 activity assay
Mouse lung tissue was homogenized in lysis buffer
(BioVision, Inc., Milpitas, CA, USA) using a homogenizer.
Subsequently, homogenated tissue was centrifuged at 10,000 × g for
10 min in order to obtain the supernatant. Protein concentration in
the supernatant was determined according to a previously described
method for Bradford assay (21).
The protein concentration was 1.7 mg per 100 mg of tissue.
Caspase-1 activity was assessed using a Caspase-1 Fluorometric
Assay kit (BioVision Inc.). The assay was based on detection of the
cleavage of substrate YVAD-7-amino-4-trifluoromethyl coumarin
(AFC). Briefly, YVAD-AFC emits blue light, whereas upon cleavage of
the substrate by caspase-1, free AFC emits a yellow-green
fluorescence, which was quantified using a spectrofluorometer
(FP-6200; Jasco, Inc., Easton, MD, USA). Comparison of the
fluorescence of AFC from a treated sample with an untreated control
sample allows the determination of fold increase in caspase-1
activity.
Statistical analysis
All data are presented as the mean ± standard
deviation of the combined experiments. Each experiment was repeated
at least twice. Comparisons between the control and sensitized mice
were evaluated using Student's t-test for independent means. To
quantify the degree to which two variables were related in
sensitized mice, Pearson's correlation test was used. P<0.05 was
considered to indicate a statistically significant difference
between values. All data were analyzed using GraphPad
Prism® 6.0 package for Windows (GraphPad Software Inc.,
La Jolla, CA, USA).
Results
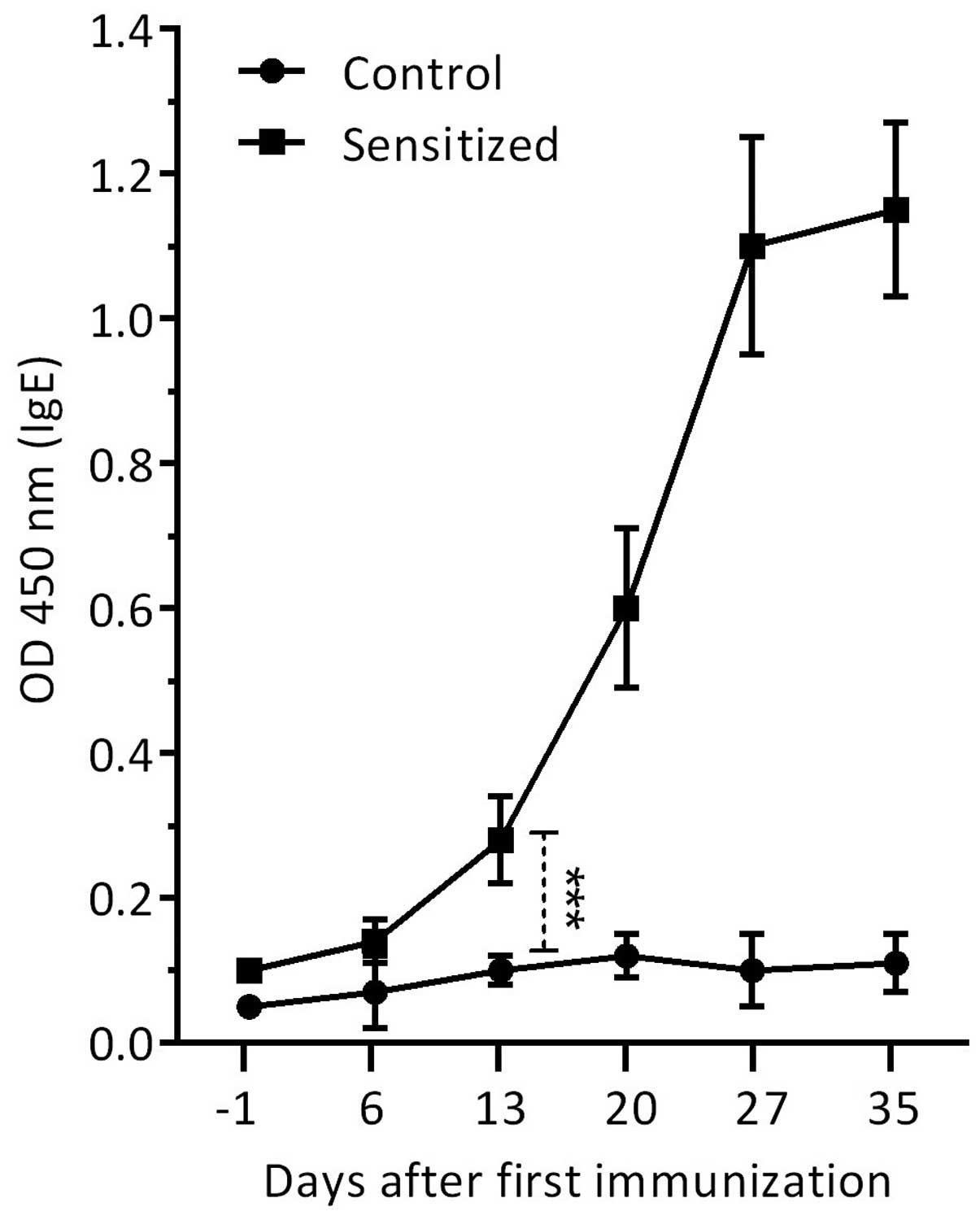
Induction of specific IgE in sensitized
mice
The allergic condition was confirmed serologically
by the presence of specific IgE to rChe a 2 in the sensitized mice
compared with in the control mice. Administration of rChe a 2 plus
alum adjuvant induced high levels of specific IgE in the sera of
sensitized mice, which was markedly increased (P=0.0001) on day 13
following initial immunization, and peaked on day 27 (Fig. 1). The increased IgE levels were
maintained at the maximum level until termination of the experiment
(day 35). No specific IgE levels were observed in the serum samples
of the control group, which received PBS absorbed onto an alum gel
suspension.
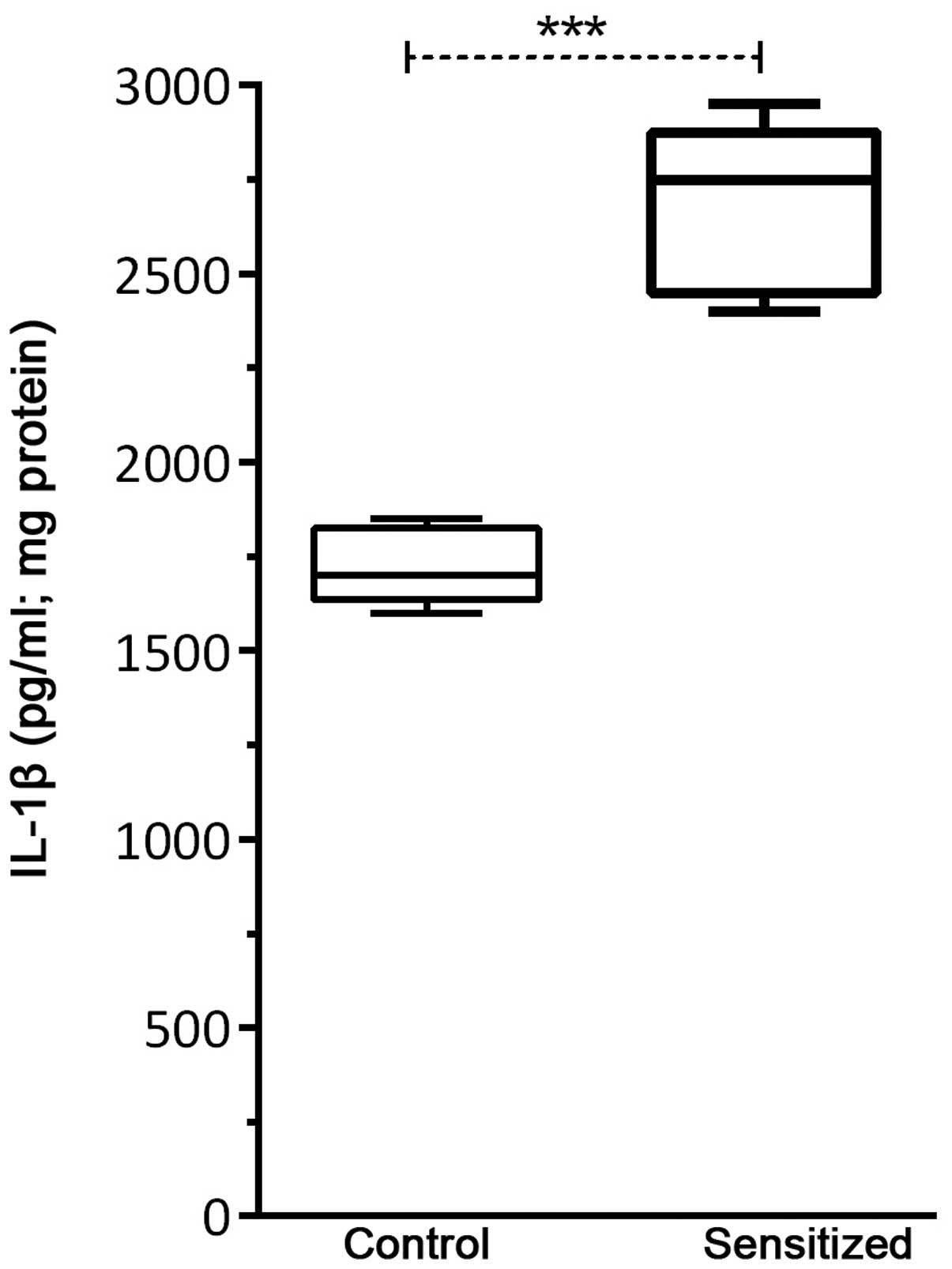
IL-1β production following
sensitization
As shown in Fig. 2,
IL-1β levels were significantly increased in the sensitized mice,
as compared with in the control mice (P<0.001). It should be
noted that IL-1β levels were calculated per 1 mg of total protein
obtained from tissue homogenates.
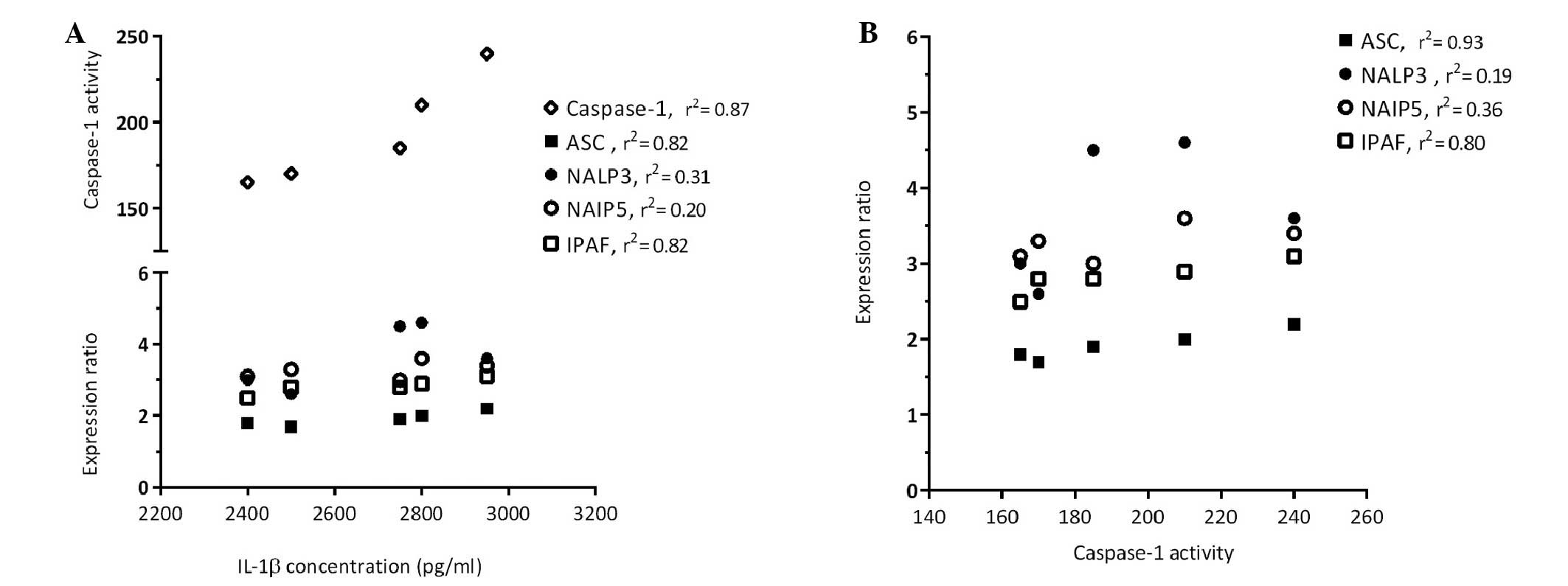
Significant association of IPAF
expression and IL-1β produc- tion in sensitized mice
In the present study, the expression levels of genes
associated with inflammasome activation were evaluated by
semi-quantitative RT-PCR in the sensitized and control mice. The
results of an RT-PCR assay for detection of the following
components of the inflammasome pathway are shown in Fig. 3: ASC, NALP3, NAIP5 and IPAF. The
relative expression levels of ASC, NALP3 and NAIP5 were normalized
to HPRT expression, and no significant differences were detected
between the groups (Fig. 3A–D). As
shown in Fig. 4, analysis of the
correlation between NALP3 and NAIP5 expression and IL-1β
concentration (Fig. 4A) and
caspase-1 activity (Fig. 4B)
demonstrated that these genes did not exert effects on IL-1β
production or caspase-1 activation following allergy induction in
sensitized mice. As shown in Fig.
3E, the mRNA expression levels of IPAF were significantly
higher (P=0.0003) in the sensitized mice compared with in the
control mice. In addition, results obtained from the correlation
analysis (Fig. 4) indicated that
the high expression of IPAF in the lung homogenates of sensitized
mice was associated with increased IL-1β production (P=0.03) and
caspase-1 activity (P=0.04).
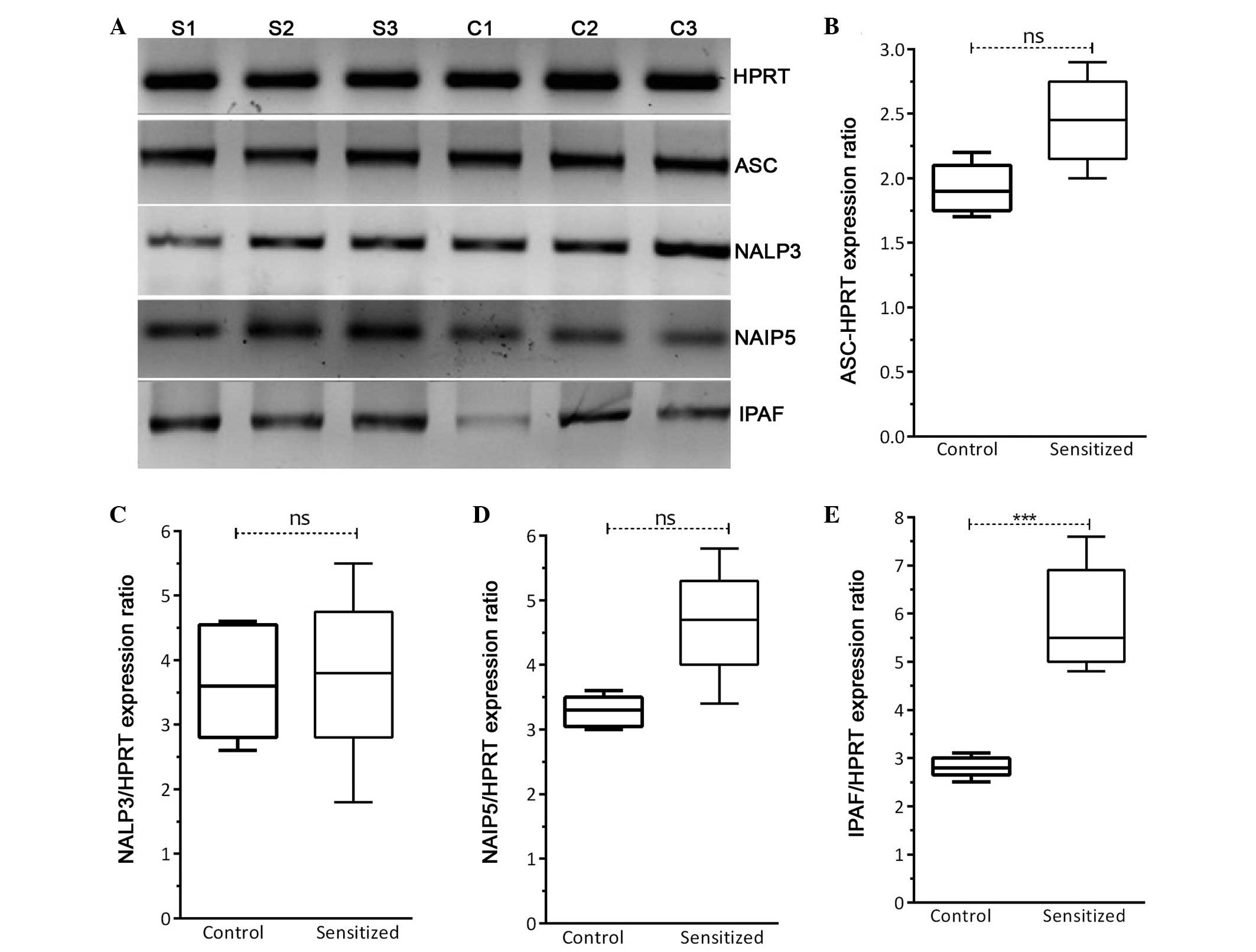 | Figure 3Expression of inflammasome-associated
genes. (A) Reverse transcription polymerase chain reaction for the
detection of components of the inflammasome pathway, including
apoptosis-associated speck-like protein (ASC), NLR family, pyrin
domain containing 3 (NALP3), NLR family, apoptosis inhibitory
protein 5 (NAIP5) and interleukin-1β converting enzyme protease
activating factor (IPAF). The gel represents the expression of
three of the five samples in each group (S, sensitized; C,
control). Relative mRNA expression levels (five samples in each
group) of (B) ASC, (C) NALP3, (D) NAIP5 and (E) IPAF following
normalization to hypoxanthine phosphoribosyl transferase (HPRT).
Data are presented as the median±interquartile range.
***P<0.001, analyzed using Student's t-test. ns, not
significant (P>0.05). |
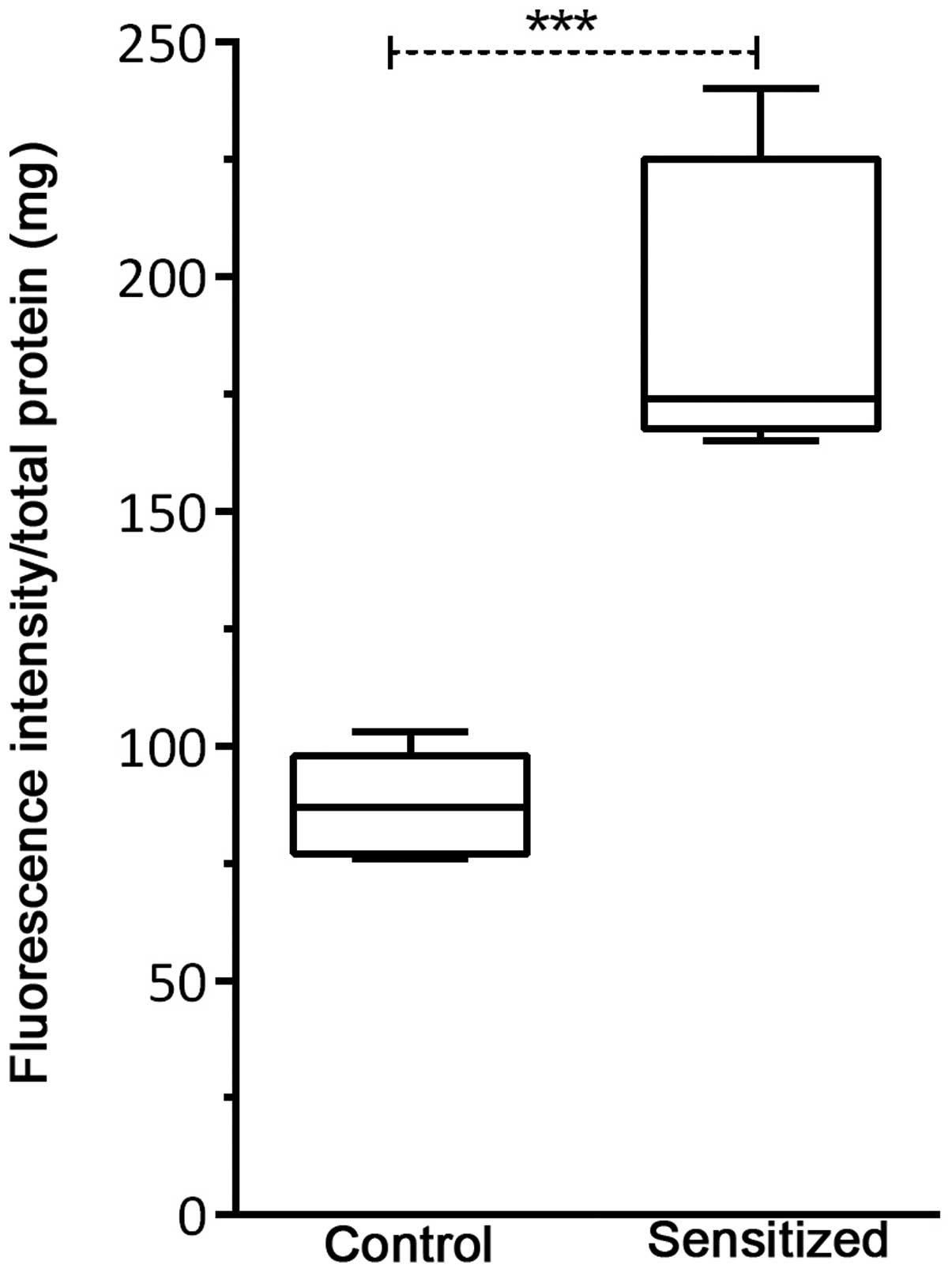
Caspase-1 activity is associated with
allergy induction
Mean adjusted caspase-1 activity per 1 mg of total
protein obtained from lung tissue homogenates was determined by
fluorometric assay (Fig. 5). A
significant increase in caspase-1 activity (P=0.001) was observed
in the sensitized mice compared with the control mice, following an
allergen challenge.
Discussion
It has been well-established that IL-1 is a critical
mediator of the inflammatory process in various diseases, including
asthma and other allergic disorders (22,23).
Allergy or hypersensitivity is one of the most important
IL-1β-associated disorders, which affects many people in developed
countries (3). Recognition of
IL-1β, and its secretion pathway via the inflammasome, may
represent a potential target for the development of novel specific
drugs for the treatment of allergy-related disorders (24,25).
Recently, the mechanisms underlying injury or
allergen-induced inflammasome activation and TLR4-dependent
allergic inflammation have been investigated (10,26).
Injury or allergen exposure may lead to the formation of specific
inflammasome pathways via caspase-1 activation, thus resulting in
IL-1β release (9,10,27).
Furthermore, animal studies have identified IL-1β as a key molecule
responsible for induction of inflammatory pathways in experimental
models of allergy (3,28,29).
The present study investigated the production of
IL-1β in an experimental model of allergy, and considered the
specific inflammasome pathway that contributes to caspase-1
activation. Previous studies have demonstrated that components of
the NLRP3 inflammasome contribute to allergic airway inflammation
via the regulation of IL-1β (17,30).
However, other publications have reported conflicting results
(31). The results of the present
study detected no significant differences in NALP3 and NAIP5
expression between the control and sensitized mice. Similar to the
present study, Kool et al (31) reported that NLRP3 does not
significantly contribute to allergic airway inflammation in mice.
In addition, Allen et al (14) indicated that NLRP3 does not
significantly contribute to the development of ovalbumin- or house
dust mite-mediated allergic airway inflammation in mice. However,
in contrast to the present results, it has been indicated that
NLRP3 inflammasome activation leads to IL-1 production, and is
important for the induction of a Th2 inflammatory allergic response
(17). The results of the present
study suggested that the NALP3 inflammasome was consistently
activated in the control and sensitized groups. This activation may
be related to the use of an alum adjuvant in the present study,
this may be controlled for in the future by adding more treatment
groups that have been sensitized to the allergen by various
methods. Other studies have confirmed that the properties of alum
adjuvant can act as a danger signal and can trigger inflammasome
activation via NALP3 (30,32).
Notably, the present study not only detected
significantly increased expression levels of IPAF in sensitized
mice compared with in control mice, but also detected increased
caspase-1 activation and IL-1β production in the lungs of
sensitized mice. Based on these obtained results, it may be
hypothesized that high levels of caspase-1 activity in sensitized
mice may be affected by a marked change in IPAF expression. In
addition, caspase-1 activation may lead to the processing and
maturation of IL-1β, thus resulting in allergic symptoms in the
sensitized mice. In accordance with these results, a previous study
reported that elevated expression of IPAF, alongside IL-1β, is
involved in the inflammation of patients with pemphigus vulgaris
(33). In addition, Liu and Chan
(34) revealed that palmitate was
able to induce the production of IL-1β via activation of the
IPAF-ASC inflammasome in primary astrocytes.
Recent studies have demonstrated that specific
pathogen-associated molecular patterns and danger-associated
molecular patterns may activate the IPAF inflammasome, which
results in recruitment of ASC and procaspase-1. Procaspase-1 is
cleaved into active caspase-1, which contributes to the processing
of pro-IL-1β into its biologically active form, IL-1β (9,35).
The IPAF inflammasome is a novel member of the NLR family, which is
a well-known sensor for bacterial flagellin during infection with
Legionella pneumophila, Salmonella and
Pseudomonas, and has been coupled with pro-IL-1β processing.
Furthermore, dysregulated IPAF expression has an essential role in
the pathogenesis of several autoimmune and inflammatory diseases,
including pemphigus vulgaris, Kawasaki and atopic dermatitis
(36,37). Composition of the IPAF inflammasome
requires ASC and pro-caspase-1. Compared with other inflammasomes,
IPAF is able to activate caspase-1 in an ASC-dependent or
-independent manner (38).
However, the exact composition of IPAF is not clear in allergy. In
an attempt to improve understanding regarding the composition of
IPAF inflammasomes, Gutierrez et al (39) demonstrated that IPAF is upregulated
by tumor necrosis factor-α in human leukemia cells. Therefore, our
next step is to identify the main stimulus of IPAF in the context
of allergy.
The results of the present study indicated that
caspase-1 activation and IL-1β expression are associated with the
IPAF inflammasome. Therefore, based on this association, the IPAF
inflammasome may be considered to have a crucial role in
experimental models of allergy. However, further experiments are
required to elucidate the exact role of the IPAF inflammasome in
allergic diseases, which may be useful for the identification of a
novel approach for the treatment and management of allergic
disorders.
In conclusion, although further analysis is required
to clarify the role of IPAF in allergy, the present study
demonstrated that IPAF inflammasome expression is associated with
caspase-1 activity and IL-1β production in an experimental model of
allergy.
Acknowledgments
The present study was supported by the Research
Administration Department of Babol University of Medical Sciences
(grant no. 9339224). The authors would like to thank Dr Evangeline
Foronda in the Research Administration Department of Babol
University of Medical Sciences for editing the manuscript.
Abbreviations:
|
ASC
|
apoptosis-associated speck-like
protein
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
IL-1β
|
interleukin-1β
|
|
IPAF
|
IL-1β converting enzyme
protease-activating factor
|
References
|
1
|
Venarske D and deShazo RD: Molecular
mechanisms of allergic disease. South Med J. 96:1049–1054. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Broide DH: Molecular and cellular
mechanisms of allergic disease. J Allergy Clin Immunol. 108(Suppl
2): S65–S71. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Krause K, Metz M, Makris M, Zuberbier T
and Maurer M: The role of interleukin-1 in allergy-related
disorders. Curr Opin Allergy Clin Immunol. 12:477–484. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hsu LC, Ali SR, McGillivray S, Tseng PH,
Mariathasan S, Humke EW, Eckmann L, Powell JJ, Nizet V, Dixit VM
and Karin M: A NOD2-NALP1 complex mediates caspase-1-dependent
IL-1beta secretion in response to Bacillus anthracis infection and
muramyl dipeptide. Proc Natl Acad Sci USA. 105:7803–7808. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dinarello CA: Immunological and
inflammatory functions of the interleukin-1 family. Annu Rev
Immunol. 27:519–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guarda G, Zenger M, Yazdi AS, Schroder K,
Ferrero I, Menu P, Tardivel A, Mattmann C and Tschopp J:
Differential expression of NLRP3 among hematopoietic cells. J
Immunol. 186:2529–2534. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Martinon F, Burns K and Tschopp J: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-beta. Mol Cell.
10:417–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schroder K and Tschopp J: The
inflammasomes. Cell. 140:821–832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Latz E: The inflammasomes: Mechanisms of
activation and function. Curr Opin Immunol. 22:28–33. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Masters SL: Specific inflammasomes in
complex diseases. Clin Immunol. 147:223–228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brusselle GG, Provoost S, Bracke KR,
Kuchmiy A and Lamkanfi M: Inflammasomes in respiratory disease:
From bench to bedside. Chest. 145:1121–1133. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jamilloux Y, Sève P and Henry T:
Inflammasomes in human diseases. Rev Med Interne. 35:730–741.
2014.In French. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dombrowski Y, Peric M, Koglin S,
Kaymakanov N, Schmezer V, Reinholz M, Ruzicka T and Schauber J:
Honey bee (Apis mellifera) venom induces AIM2 inflammasome
activation in human keratinocytes. Allergy. 67:1400–1407. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Allen IC, Jania CM, Wilson JE, Tekeppe EM,
Hua X, Brickey WJ, Kwan M, Koller BH, Tilley SL and Ting JP:
Analysis of NLRP3 in the development of allergic airway disease in
mice. J Immunol. 188:2884–2893. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arthur JC, Lich JD, Ye Z, Allen IC, Gris
D, Wilson JE, Schneider M, Roney KE, O'Connor BP, Moore CB, et al:
Cutting edge: NLRP12 controls dendritic and myeloid cell migration
to affect contact hypersensitivity. J Immunol. 185:4515–4519. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Watanabe H, Gaide O, Pétrilli V, Martinon
F, Contassot E, Roques S, Kummer JA, Tschopp J and French LE:
Activation of the IL-1beta-processing inflammasome is involved in
contact hypersensitivity. J Invest Dermatol. 127:1956–1963. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Besnard AG, Guillou N, Tschopp J, Erard F,
Couillin I, Iwakura Y, Quesniaux V, Ryffel B and Togbe D: NLRP3
inflammasome is required in murine asthma in the absence of
aluminum adjuvant. Allergy. 66:1047–1057. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dombrowski Y, Peric M, Koglin S,
Kammerbauer C, Göss C, Anz D, Simanski M, Gläser R, Harder J,
Hornung V, et al: Cytosolic DNA triggers inflammasome activation in
kerati-nocytes in psoriatic lesions. Sci Transl Med. 3:82ra382011.
View Article : Google Scholar
|
|
19
|
Nouri HR, Sankian M, Vahedi F, Afsharzadeh
D, Rouzbeh L, Moghadam M and Varasteh A: Diagnosis of Chenopodium
album allergy with a cocktail of recombinant allergens as a tool
for component-resolved diagnosis. Mol Biol Rep. 39:3169–3178. 2012.
View Article : Google Scholar
|
|
20
|
Nouri HR, Sankian M, Afsharzadeh D and
Varasteh A: Immunotherapy with a recombinant hybrid molecule
alleviates allergic responses more efficiently than an allergenic
cocktail or pollen extract in a model of Chenopodium album allergy.
Int Arch Allergy Immunol. 161:325–332. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lane T and Lachmann HJ: The emerging role
of interleukin-1β in autoinflammatory diseases. Curr Allergy Asthma
Rep. 11:361–368. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sedimbi SK, Hägglöf T and Karlsson MC:
IL-18 in inflammatory and autoimmune disease. Cell Mol Life Sci.
70:4795–4808. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
López-Castejón G and Pelegrín P: Current
status of inflam-masome blockers as anti-inflammatory drugs. Expert
Opin Investig Drugs. 21:995–1007. 2012. View Article : Google Scholar
|
|
25
|
Kontogiorgis CA and Hadjipavlou-Litina DJ:
Non steroidal anti-inflammatory and anti-allergy agents. Curr Med
Chem. 9:89–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fietta P and Delsante G: The
inflammasomes: The key regulators of inflammation. Riv Biol.
102:365–384. 2009.PubMed/NCBI
|
|
27
|
Skeldon AM, Faraj M and Saleh M: Caspases
and inflam-masomes in metabolic inflammation. Immunol Cell Biol.
92:304–313. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Blom L and Poulsen LK: IL-1 family members
IL-18 and IL-33 upregulate the inflammatory potential of
differentiated human Th1 and Th2 cultures. J Immunol.
189:4331–4337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lukens JR, Gross JM and Kanneganti TD:
IL-1 family cytokines trigger sterile inflammatory disease. Front
Immunol. 3:3152012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Eisenbarth SC, Colegio OR, O'Connor W,
Sutterwala FS and Flavell RA: Crucial role for the Nalp3
inflammasome in the immunostimulatory properties of aluminium
adjuvants. Nature. 453:1122–1126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kool M, Willart MA, van Nimwegen M, Bergen
I, Pouliot P, Virchow JC, Rogers N, Osorio F, Reise Sousa C, Hammad
H and Lambrecht BN: An unexpected role for uric acid as an inducer
of T helper 2 cell immunity to inhaled antigens and inflammatory
mediator of allergic asthma. Immunity. 34:527–540. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kool M, Pétrilli V, De Smedt T, Rolaz A,
Hammad H, van Nimwegen M, Bergen IM, Castillo R, Lambrecht BN and
Tschopp J: Cutting edge: Alum adjuvant stimulates inflammatory
dendritic cells through activation of the NALP3 inflammasome. J
Immunol. 181:3755–3759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shamsabadi RM, Basafa S, Yarahmadi R,
Goorani S, Khani M, Kamarehei M and Hossein Kiani A: Elevated
expression of NLRP1 and IPAF are related to oral pemphigus vulgaris
pathogenesis. Inflammation. 38:205–208. 2015. View Article : Google Scholar
|
|
34
|
Liu L and Chan C: IPAF inflammasome is
involved in interleukin-1β production from astrocytes, induced by
palmitate; implications for Alzheimer's disease. Neurobiol Aging.
35:309–321. 2014. View Article : Google Scholar
|
|
35
|
Mariathasan S, Newton K, Monack DM, Vucic
D, French DM, Lee WP, Roose-Girma M, Erickson S and Dixit VM:
Differential activation of the inflammasome by caspase-1 adaptors
ASC and Ipaf. Nature. 430:213–218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Abdelaziz DH, Amr K and Amer AO:
Nlrc4/Ipaf/CLAN/CARD12: More than a flagellin sensor. Int J Biochem
Cell Biol. 42:789–791. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vinzing M, Eitel J, Lippmann J, Hocke AC,
Zahlten J, Slevogt H, N'guessan PD, Günther S, Schmeck B,
Hippenstiel S, et al: NAIP and Ipaf control Legionella pneumophila
replication in human cells. J Immunol. 180:6808–6815. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu B, Elinav E, Huber S, Booth CJ, Strowig
T, Jin C, Eisenbarth SC and Flavell RA: Inflammation-induced
tumorigenesis in the colon is regulated by caspase-1 and NLRC4.
Proc Natl Acad Sci USA. 107:21635–21640. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gutierrez O, Pipaon C and Fernandez-Luna
JL: Ipaf is upregulated by tumor necrosis factor-alpha in human
leukemia cells. FEBS Lett. 568:79–82. 2004. View Article : Google Scholar : PubMed/NCBI
|