Introduction
Alzheimer's disease (AD), a progressive
neurodegenerative disease of the central nervous system, is the
most common cause of senile dementia (1). AD is characterized by irreversible
loss of neurons and a clinically gradual deterioration in
intellectual abilities including cognition and memory (1). The AD brain is characterized by the
presence of neuritic plaques, neurofibrillary tangles, and the loss
of cortical neurons and synapses (2). β-amyloid (Aβ), generated from a
sequent cleavage of amyloid precursor protein (APP) by β-site APP
cleavage enzyme 1 and γ-secretase, is the major component of
extracellular neuritic plaques (3). The accumulation of Aβ contributes to
AD pathogenesis in that the accumulation of Aβ induces an increase
in intracellular reactive oxygen species (ROS) and neuronal cell
apoptosis (4–7). Previous findings emphasize the
critical role of oxidative stress as a major cause in the
pathogenesis of AD (8,9). Therefore, antioxidants may attenuate
Aβ-induced neurotoxicity and cell death, leading to the
amelioration of AD-induced impairment in cognition and memory
(10,11).
Flavonoids have a 15-carbon skeleton composed of two
fused 6-membered rings (an aromatic ring and a heterocyclic ring)
connected through a carbon-carbon bridge to an aromatic ring
(12). In the human diet,
flavonoids constitute the most common group of polyphenolic
compounds that are synthesized in plant cells. The phenolic
hydroxyl groups attached to ring structures of flavonoids allow
them to act as antioxidants (13,14),
as radical-scavenging agents (15,16),
or as agents of anti-inflammation (17,18).
Accumulating evidence showed that the consumption of flavonoid-rich
foods is associated with lower rates of dementia and has some
beneficial effects on memory and learning (19,20).
Luteolin, a flavonoid compound, has been suggested
to have neuroprotective properties. Findings of in vivo
studies examining rat models have shown that luteolin protects
against cognitive dysfunction induced by chronic cerebral
hypoperfusion (21). Luteolin also
protects against high-fat diet-induced cognitive defects in obesity
mice (22). Fu et al
(21) recently reported that
luteolin can protect against cognitive dysfunction induced by
chronic cerebral hypoperfusion in rats. In the context of AD,
Rezai-Zadeh et al (23)
showed that luteolin treatment of murine N2a cells transfected with
SweAPP and primary neuronal cells derived from
SweAPP-overexpressing mice resulted in a significant reduction in
Aβ generation. The mechanism may involve selective inactivation of
the GSK-3α isoform, which increases the phosphorylation of PS1, the
catalytic core of the γ-secretase complex, thereby reducing PS1-APP
interaction and Aβ generation. In a later study, Zhou et al
(24) reported that luteolin
attenuated zinc-induced hyperphosphorylation of the protein τ in
SH-SY5Y cells through its antioxidant action. Luteolin also
inhibited the τ kinase p7056K but established recovery of total
phosphatase activity. More recently, it was found that luteolin
reduced AD pathologies induced by traumatic brain injury (25). However, the overall neuroprotective
effect of luteolin in drug-induced Alzheimer's rat models remains
to be investigated. The present study aimed to investigate the
effect of luteolin on spatial learning and the structure of CA1
pyramidal layer thickness in a streptozotocin (STZ)-induced AD
model.
Materials and methods
Animals
A total of 60 male Wistar rats, weighing 200–230 g,
and aged 3 months, were housed in cages with a temperature of
24–26°C and a 12-h dark/light cycle with food and water ad
libitum. Behavioral experiments were performed in the morning.
The experiment protocols were approved by the Animal Ethics
Committee of the Xinxiang Central Hospital (Henan, China).
Establishment of the animal model
Animals were anesthetized with ketamine (80 mg/kg)
and xylazine (15 mg/kg) intraperitoneally, and their heads were
fixed into a stereotaxic instrument (Narishige, Tokyo, Japan). Two
stainless 23-gauge guide cannula were implanted in the lateral
ventricles bilaterally. Stereotaxic coordinates were based on
Paxinos and Watson atlas of the rat brain. Following surgery, the
animals were kept in cages for 6 days to recover.
Experimental protocol
The 60 rats were divided into 6 groups with 10 rats
in each group. Apart from the control group in which the rats did
not receive any treatment or surgery, the rats in the remaining 5
groups received surgery and recovery treatments. In the saline
group, rats received normal saline after recovery; in the sham
group, rats received 10% DMSO after recovery; in the STZ group
(Alzheimer's model), rats received STZ (3 mg/kg) on the fourth and
sixth days after recovery; and in the L10 and L20 groups, STZ was
injected on days 4 and 6 and the rats were treated with luteolin at
doses of 10 and 20 mg/kg, respectively, on the 1st, 2nd, 3rd and
5th days after recovery.
Drugs were injected via the intracerebroventricular
(ICV) route in a total volume of 10 μl at the rate of 1
μl/min. The 27-gauge injection needle was inserted into the
guide cannula. The injection needle was attached to a 10 μl
syringe by a polyethylene tube.
Morris water maze test
The Morris water maze consists of a circular water
tank with 160 cm diameter and 60 cm height, filled with non-toxic
water (25±2°C) to a depth of 25 cm. The pool was divided into 4
quadrants (North, South, East and West) which were used as start
points. An escape platform (10 cm in diameter) made of plexiglass
was placed in the middle of one of the randomly selected quadrants
of the pool, 1 cm below the surface of water and kept in the same
position throughout the entire experiment (North-West for this
study). Spatial learning of animals was tested 14 days after STZ
infusion in the Morris water maze. The rats in each group were
tested (one at a time). The rats were trained for four days prior
to the formal experiments. Each rat was subjected to 4 consecutive
trials on each day with an interval of 1 min.
Each trial was initiated by placing the rat randomly
at 1 of the 4 starting points. The rats were allowed to swim in the
pool during a period of 90 sec to locate the hidden platform. If a
rat did not locate the hidden platform within this period, it was
manually guided to the platform by the investigator. The rats were
allowed to remain on the platform for 30 sec. All the trials were
performed at 9 a.m.
Directions of the rats were recorded by a video
camera (Fuji, Tokyo, Japan) above the center of the maze that was
linked to a computer. Spatial acquisition was evaluated by
measuring escape latency (time to find the platform), traveled
distance (path length to reach the platform), and swimming speed
using the EthoVision tracking system (Noldus Information
Technology, Wageningen, The Netherlands), as described previously
(26,27). The data obtained from rats with
visual impairment were excluded.
Probe test
To assess memory consolidation, a probe test was
performed 24 h after the Morris water maze test. For the probe
test, the platform was removed and the rats were allowed to swim
freely. The swimming pattern of every rat was recorded with a
camera. Consolidated spatial memory was estimated by the time spent
in the target quadrant area.
Tissue preparation
The rats were anesthetized with Zoletil 50 (10
mg/kg, i.m.) and perfused intra-cardiac infusion with 100 mmol/l
phosphate-buffered saline (PBS) followed by ice-cold 4%
paraformaldehyde. The brains of the rats were isolated and
post-fixed in 50 mmol/l PBS containing 4% paraformaldehyde
overnight, immersed in a solution containing 30% sucrose in 50
mmol/l PBS and stored at 4°C until sectioning. The frozen brains
were coronally sectioned on paraffin at 3 μm and then stored
in a storage solution at 4°C.
Immunohistochemistry
For the evaluation of dendritic damage, the sections
were immunostained with a rabbit antibody against rat MAP2
(Millipore Corp., Billerica, MA, USA) at a 1:250 dilution. Three
sections from each animal were assessed for scoring.
Statistical analysis
SPSS 13.0 software (SPSS Inc., Chicago, IL, USA) was
used for data analysis. Analysis of variance was used for
comparison of the behavioral and histological data. A Tukey
multiple comparison post-test was performed to assess differences
between groups. P<0.05 was considered to indicate a
statistically significant.
Results
Luteolin improved the cognitive function
of rats with STZ in Morris water maze test
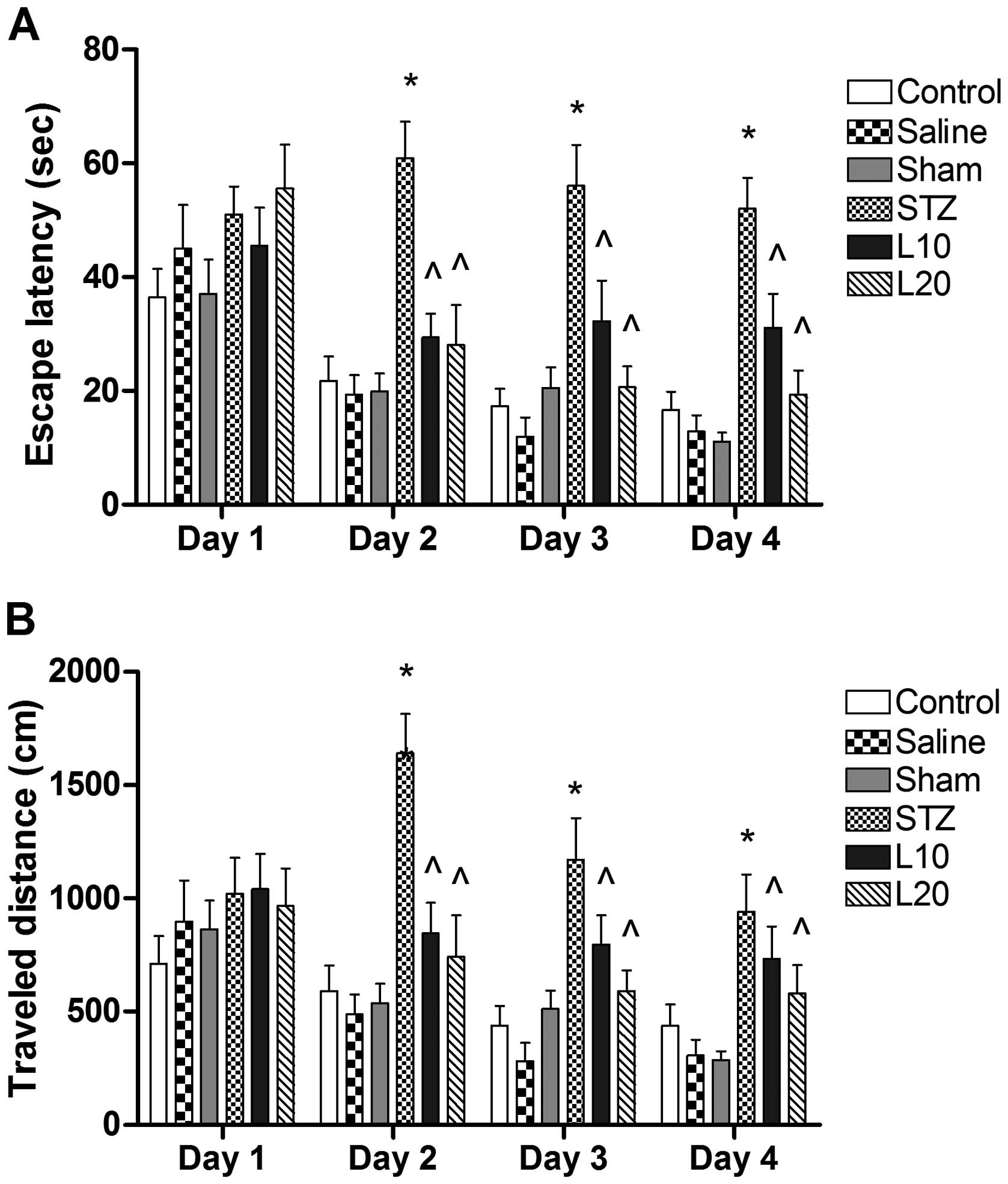
To evaluate the cognitive function of rats, the
Morris water maze test was performed. The mean escape latency of
rats in different groups did not show a significant difference for
the first day (P>0.05). However, there was a significant
difference in escape latency from the second day (P<0.05). The
rats treated with STZ showed a significantly decreased ability to
locate the platform and to learn its location on the fifth day of
training (Fig. 1A). By contrast,
this poor performance was significantly mitigated by the treatment
with luteolin (10 and 20 mg/kg) as shown by the decreased latency
to locate the platform from the second day of training. The effect
of treatment with luteolin at 10 and 20 mg/kg was not significantly
different (P>0.05).
Luteolin decreased the travel distance to
reach the hidden platform (path length)
Progressive decreases in the path length to reach
the hidden platform on subsequent days in the water maze task were
associated with intact memory of animals. Therefore, the traveled
distance was measured to evaluate the memory impairment. As shown
in Fig. 1B, the total distance
traveled to reach the hidden platform did not differ between any of
the groups on the first day of testing in the Morris water maze.
However, there was a significant difference in the traveled
distance of STZ-treated rats as compared to the sham animals from
the second day onwards (P<0.05). Chronic treatment with luteolin
(10 and 20 mg/kg) significantly (P<0.05) decreased the total
distance traveled to reach the platform in the STZ-injected rats,
which suggested improvement in memory associated with luteolin
treatment. When the effect of luteolin at different doses was
compared, the reduction in traveled distance with luteolin at doses
of 10 and 20 mg/kg was not significantly different (P>0.05).
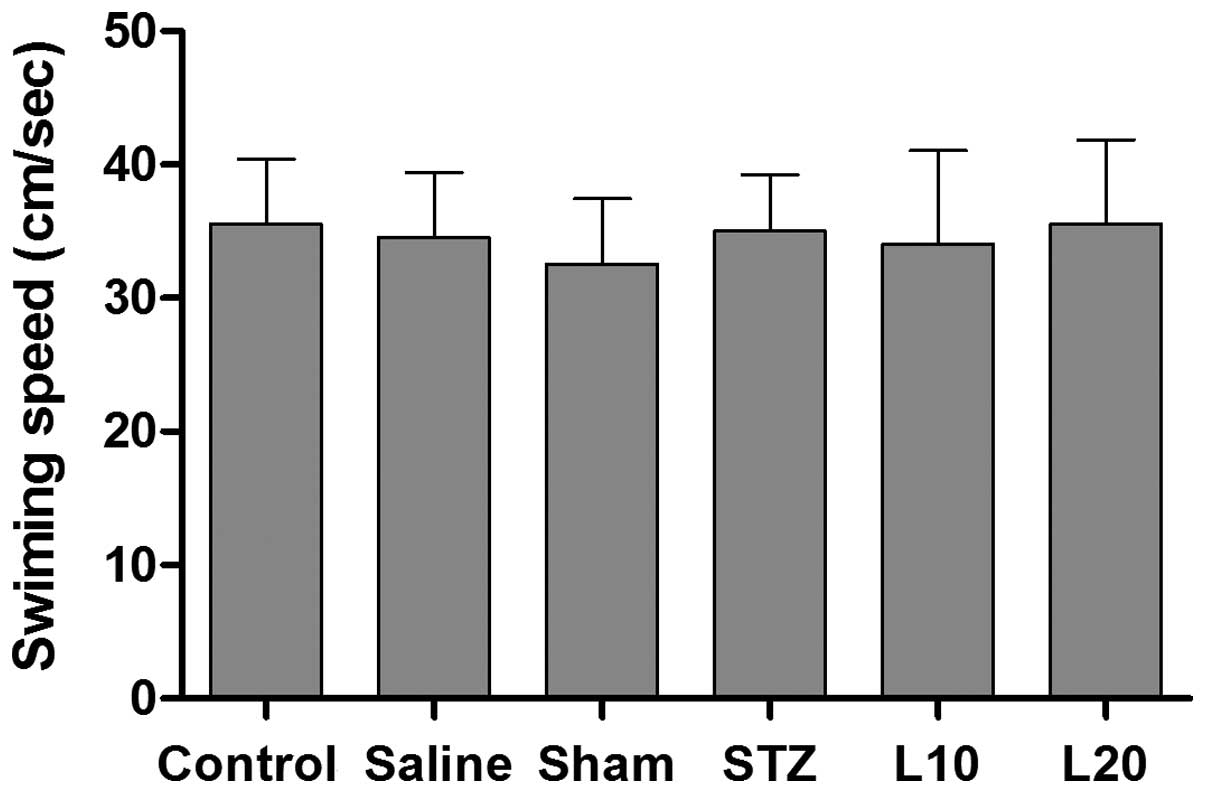
Luteolin has no effect on swimming speed
of ICV-STZ-injected rats
As shown in Fig. 2,
no significant difference in swimming speed was observed between
any groups in the probe trial. The mean values in control, saline,
sham, STZ, and luteolin (10 and 20 mg/kg) groups were 35.5±3.7,
34.6±5.1, 32.7±3.6, 35.6±4.6, 34.5±3.9 and 35.7±4.9 cm/sec,
respectively (P>0.05), indicating that there was no motor
activity disturbances in tested animals.
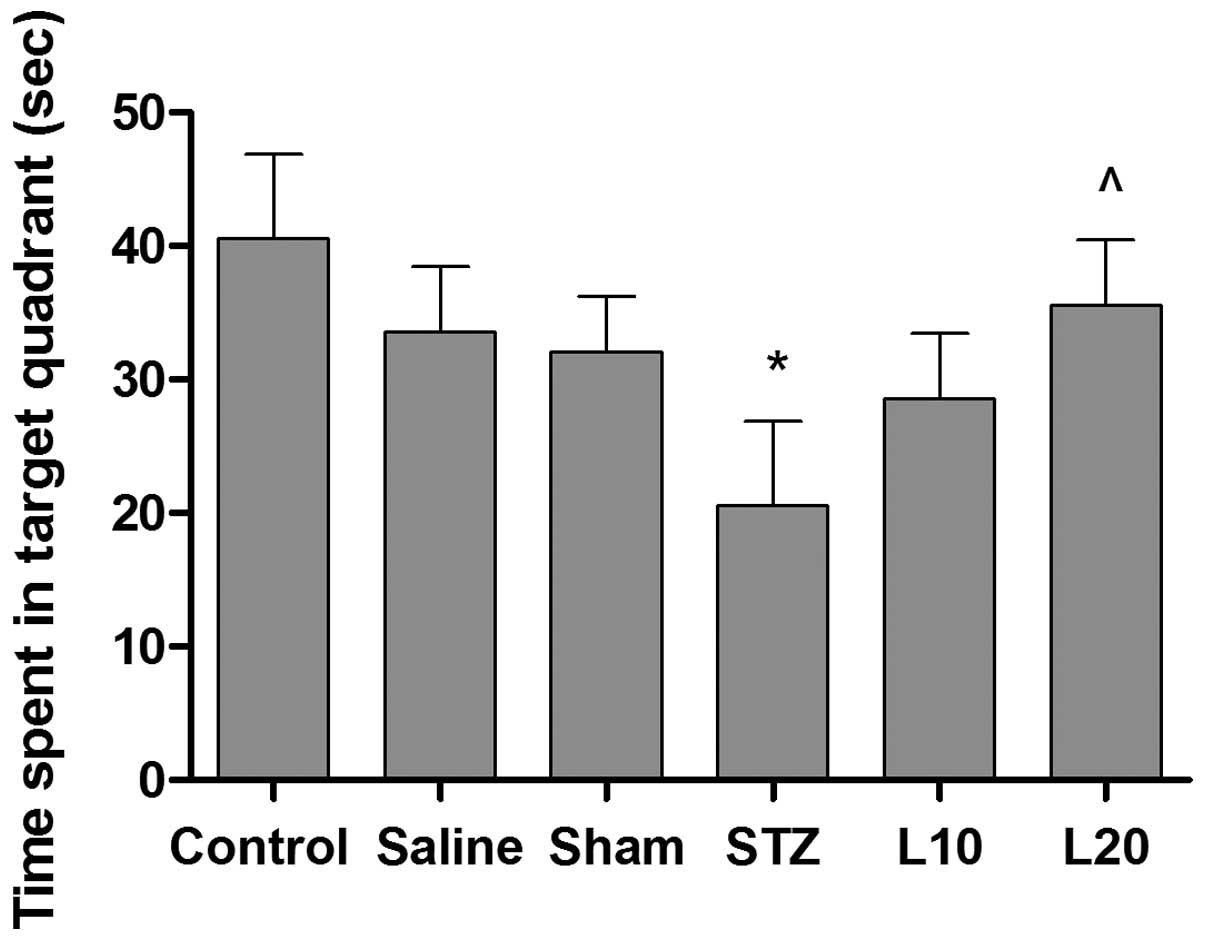
Effect of luteolin on time spent in
target quadrant in ICV-STZ-injected rats
To examine the learning ability and consolidation of
the platform location during the training, a probe test was
performed. As shown in Fig. 3,
animals in different groups showed a significant different
performance. The time spent in the target quadrant was
significantly lower in the STZ rats as compared to the sham group.
STZ rats treated with luteolin (10 and 20 mg/kg) spent more time in
the target quadrant than the monotherapy group in the probe test.
In particular, STZ-treated rats with luteolin at a dose of 20 mg/kg
spent significantly more time in the target quadrant compared with
STZ-treated rats (P<0.05).
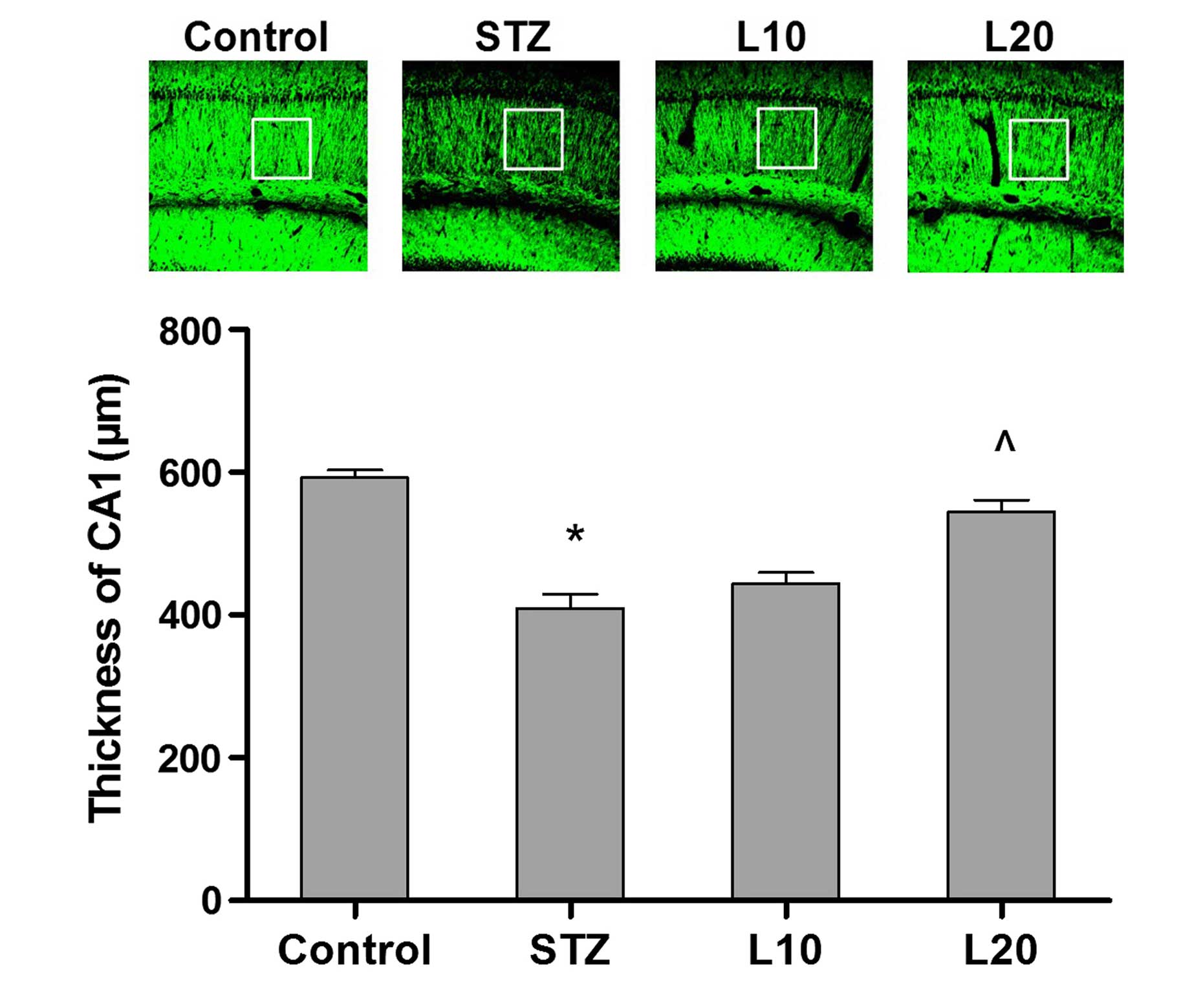
Luteolin increased the thickness of CA1
pyramidal layer
The thickness of the hippocampal CA1 pyramidal layer
in the tested groups was measured to test the effect of luteolin
treatment. The mean CA1 pyramidal layer thicknesses were 592.5±27.2
μm in the control group and 409.3±23.6 μm in the STZ
group (P<0.05). The mean CA1 pyramidal layer thickness in the
L20 group was 544.3±32.6 μm showing a significant increase
compared to 409.3±23.6 μm of the STZ group (P<0.05,
Fig. 4).
Discussion
Flos Chrysanthemi, the flower of Dendranthema
morifolium Ramat Tzvel (Chrysanthemum morifolium
Ramat.), is known as Ju-Hua in Chinese. This herb is listed in Shen
Nong's Herbal (a historical book of Chinese herbs) as a non-toxic,
top-grade herb that has been used as an agent for the treatment of
headache, vertigo, and sore throat (28). Flos Chrysanthemi is an edible
medicinal herb, and one of the major active ingredients of this
plant is luteolin, a flavonoid compound. The pharmacological
activity of luteolin is thought to be associated in part with its
antioxidant potential (28,29),
anti-tumorigenic effects (30,31),
and anti-inflammatory/anti-allergic activities (29). In addition, it has been reported
that luteolin can act as an inhibitor of protein kinase C and
lipoxygenase (32). In
vitro and in vivo studies have also shown that luteolin
reduces high blood cholesterol through the inhibition of
cholesterol uptake in Caco-2 cells and affects cholesterol
transport (33). Luteolin also has
a radio-protective and a protective effect on doxorubicin-induced
cardiotoxicity in mice (28,34).
The results of the present study indicate that luteolin reduced the
escape latency and traveled distance parameters in the Morris water
maze while increasing the time spent in the target quadrant in the
animal model of AD. These results indicate that luteolin can
improve spatial learning and memory impairment in this experimental
model. It also prevents the thickness reduction of the CA1
pyramidal cell layer. This finding along with the previous in
vitro studies revealed the neuroprotective effect of triazine
derivatives in the experimental model (23,24).
The ICV STZ model produces cognitive defects similar
to those observed in the sporadic dementia of Alzheimer's type
(35). STZ administration in rats
induces oxidative stress in the brain, Aβ plaques aggregation, τ
protein hyperphosphorylation, neuroinflammation, and apoptosis,
which cooperate to repair memory and learning in the AD animal
models (35,36). Aβ plaque is the proteolytic product
of APP, which can generate the ROS, especially hydrogen peroxides,
leading to cell death in the neuronal cultures and toxicity in
hippocampal neurons (37–39). On the other hand, STZ (ICV-STZ) in
sub-diabetogenic dose reduces energy metabolism, leading to
cognitive dysfunction by inhibiting the synthesis of adenosine
triphosphate. The repair of glucose and energy impairment caused by
STZ is a potential source for this oxidative stress (40). Oxidative stress is the most
important hypothesis involved in the pathophysiology of AD in that
excess free radicals of oxygen lead to cell damage, a progressive
cognition and memory loss (38,39,41).
Additionally, the reciprocal effects of oxidative stress and Aβ
aggregation intensify the impairment of neurological function
(5). In the present study, the
results showed that STZ injection induced significant defects of
memory, indicating the success of AD model establishment.
Given the critical role of ROS in the pathogenesis
of AD, numerous ROS scavengers have been employed to investigate
whether they can protect against memory defects in an experimental
model of AD (42,43). A number of ROS modulators including
flavonoid compounds such as EGCG attenuate cell injury and
neurotoxicity during Aβ exposure (44). Accumulating evidence has shown that
luteolin can act as a ROS scavenger agent in a number of tissues or
cells. Yu et al (45)
reported that luteolin can protect myocardial ischemia/reperfusion
injury by inhibiting the ROS-MAPK-mediated mechanism. Another study
showed that luteolin inhibits Cr (VI)-induced malignant cell
transformation of human lung epithelial cells by targeting
ROS-mediated multiple cell signaling pathways (46). In human umbilical vein endothelial
cells, luteolin suppressed the TNF-α-activated ROS generation, and
the expression of Nox4, p22phox, and ICAM-1 and VCAM-1 (47). The present findings suggest that
luteolin improved memory impairment and prevents the reduction of
CA1 pyramidal cell layer in an AD model. Thus, the neuroprotective
effect of luteolin against AD nay be due to the antioxidative
characteristic of this agent by inhibiting free radical products
and dispersing Aβ plaques. Therefore, luteolin is potentially a
suitable therapeutic candidate for neural disorders such as AD,
although further studies including clinical trials, are needed to
confirm this hypothesis.
References
|
1
|
Zhu Z, Yan J, Jiang W, Yao XG, Chen J,
Chen L, Li C, Hu L, Jiang H and Shen X: Arctigenin effectively
ameliorates memory impairment in Alzheimer's disease model mice
targeting both β-amyloid production and clearance. J Neurosci.
33:13138–13149. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Terry RD, Masliah E, Salmon DP, Butters N,
DeTeresa R, Hill R, Hansen LA and Katzman R: Physical basis of
cognitive alterations in Alzheimer's disease: synapse loss is the
major correlate of cognitive impairment. Ann Neurol. 30:572–580.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Choi SM, Kim BC, Cho YH, Choi KH, Chang J,
Park MS, Kim MK, Cho KH and Kim JK: Effects of flavonoid compounds
on β-amyloid-peptide-induced neuronal death in cultured mouse
cortical neurons. Chonnam Med J. 50:45–51. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marcus DL, Thomas C, Rodriguez C,
Simberkoff K, Tsai JS, Strafaci JA and Freedman ML: Increased
peroxidation and reduced antioxidant enzyme activity in Alzheimer's
disease. Exp Neurol. 150:40–44. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zheng L, Kågedal K, Dehvari N, Benedikz E,
Cowburn R, Marcusson J and Terman A: Oxidative stress induces
macroautophagy of amyloid beta-protein and ensuing apoptosis. Free
Radic Biol Med. 46:422–429. 2009. View Article : Google Scholar
|
|
6
|
Behl C and Holsboer F: Oxidative stress in
the pathogenesis of Alzheimer's disease and antioxidant
neuroprotection. Fortschr Neurol Psychiatr. 66:113–121. 1998.In
German. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng QL, Buz'Zard AR and Lau BH:
Pycnogenol protects neurons from amyloid-beta peptide-induced
apoptosis. Brain Res Mol Brain Res. 104:55–65. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Andersen JK: Oxidative stress in
neurodegeneration: cause or consequence? Nat Med. 10(Suppl):
S18–S25. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Butterfield DA, Perluigi M and Sultana R:
Oxidative stress in Alzheimer's disease brain: New insights from
redox proteomics. Eur J Pharmacol. 545:39–50. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aliev G, Obrenovich ME, Reddy VP, Shenk
JC, Moreira PI, Nunomura A, Zhu X, Smith MA and Perry G:
Antioxidant therapy in Alzheimer's disease: theory and practice.
Mini Rev Med Chem. 8:1395–1406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin YH, Liu AH, Wu HL, Westenbroek C, Song
QL, Yu HM, Ter Horst GJ and Li XJ: Salvianolic acid B, an
antioxidant from Salvia miltiorrhiza, prevents Abeta(25–35)-induced
reduction in BPRP in PC12 cells. Biochem Biophys Res Commun.
348:593–599. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Merken HM and Beecher GR: Measurement of
food flavonoids by high-performance liquid chromatography: A
review. J Agric Food Chem. 48:577–599. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rice-Evans CA, Miller NJ and Paganga G:
Structure-antioxidant activity relationships of flavonoids and
phenolic acids. Free Radic Biol Med. 20:933–956. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zeng LH, Zhang HD, Xu CJ, Bian YJ, Xu XJ,
Xie QM and Zhang RH: Neuroprotective effects of flavonoids
extracted from licorice on kainate-induced seizure in mice through
their antioxidant properties. J Zhejiang Univ Sci B. 14:1004–1012.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dugas AJ Jr, Castañeda-Acosta J, Bonin GC,
Price KL, Fischer NH and Winston GW: Evaluation of the total
peroxyl radical-scavenging capacity of flavonoids:
Structure-activity relationships. J Nat Prod. 63:327–331. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cho JG, Song NY, Nam TG, Shrestha S, Park
HJ, Lyu HN, Kim DO, Lee G, Woo YM, Jeong TS, et al: Flavonoids from
the grains of C1/R-S transgenic rice, the transgenic Oryza sativa
spp. japonica, and their radical scavenging activities. J Agric
Food Chem. 61:10354–10359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nijveldt RJ, van Nood E, van Hoorn DE,
Boelens PG, van Norren K and van Leeuwen PA: Flavonoids: a review
of probable mechanisms of action and potential applications. Am J
Clin Nutr. 74:418–425. 2001.PubMed/NCBI
|
|
18
|
Thilakarathna SH and Rupasinghe HP:
Flavonoid bioavailability and attempts for bioavailability
enhancement. Nutrients. 5:3367–3387. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Beking K and Vieira A: Flavonoid intake
and disability-adjusted life years due to Alzheimer's and related
dementias: a population-based study involving twenty-three
developed countries. Public Health Nutr. 13:1403–1409. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Letenneur L, Proust-Lima C, Le Gouge A,
Dartigues JF and Barberger-Gateau P: Flavonoid intake and cognitive
decline over a 10-year period. Am J Epidemiol. 165:1364–1371. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fu X, Zhang J, Guo L, Xu Y, Sun L, Wang S,
Feng Y, Gou L, Zhang L and Liu Y: Protective role of luteolin
against cognitive dysfunction induced by chronic cerebral
hypoperfusion in rats. Pharmacol Biochem Behav. 126:122–130. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Y, Fu X, Lan N, Li S, Zhang J, Wang S,
Li C, Shang Y, Huang T and Zhang L: Luteolin protects against high
fat diet-induced cognitive deficits in obesity mice. Behav Brain
Res. 267:178–188. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rezai-Zadeh K, Douglas Shytle R, Bai Y,
Tian J, Hou H, Mori T, Zeng J, Obregon D, Town T and Tan J:
Flavonoid-mediated presenilin-1 phosphorylation reduces Alzheimer's
disease beta-amyloid production. J Cell Mol Med. 13:574–588. 2009.
View Article : Google Scholar :
|
|
24
|
Zhou F, Chen S, Xiong J, Li Y and Qu L:
Luteolin reduces zinc-induced tau phosphorylation at Ser262/356 in
an ROS-dependent manner in SH-SY5Y cells. Biol Trace Elem Res.
149:273–279. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sawmiller D, Li S, Shahaduzzaman M, Smith
AJ, Obregon D, Giunta B, Borlongan CV, Sanberg PR and Tan J:
Luteolin reduces Alzheimer's disease pathologies induced by
traumatic brain injury. Int J Mol Sci. 15:895–904. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sharifzadeh M, Naghdi N, Khosrovani S,
Ostad SN, Sharifzadeh K and Roghani A: Post-training
intrahippocampal infusion of the COX-2 inhibitor celecoxib impaired
spatial memory retention in rats. Eur J Pharmacol. 511:159–166.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tabrizian K, Najafi S, Belaran M,
Hosseini-Sharifabad A, Azami K, Hosseini A, Soodi M, Kazemi A,
Abbas A and Sharifzadeh M: Effects of selective iNOS inhibitor on
spatial memory in recovered and non-recovered ketamine
induced-anesthesia in wistar rats. Iran J Pharm Res. 9:313–320.
2010.PubMed/NCBI
|
|
28
|
Shimoi K, Masuda S, Furugori M, Esaki S
and Kinae N: Radioprotective effect of antioxidative flavonoids in
gamma-ray irradiated mice. Carcinogenesis. 15:2669–2672. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Seelinger G, Merfort I and Schempp CM:
Anti-oxidant, anti-inflammatory and anti-allergic activities of
luteolin. Planta Med. 74:1667–1677. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yasukawa K, Takido M, Takeuchi M and
Nakagawa S: Effect of chemical constituents from plants on
12-O-tetradecanoylphorbol-13-acetate-induced inflammation in mice.
Chem Pharm Bull (Tokyo). 37:1071–1073. 1989. View Article : Google Scholar
|
|
31
|
Majumdar D, Jung KH, Zhang H, Nannapaneni
S, Wang X, Amin AR, Chen Z, Chen ZG and Shin DM: Luteolin
nanoparticle in chemoprevention: in vitro and in vivo anticancer
activity. Cancer Prev Res (Phila). 7:65–73. 2014. View Article : Google Scholar
|
|
32
|
Ferriola PC, Cody V and Middleton E Jr:
Protein kinase C inhibition by plant flavonoids. Kinetic mechanisms
and structure-activity relationships. Biochem Pharmacol.
38:1617–1624. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nekohashi M, Ogawa M, Ogihara T, Nakazawa
K, Kato H, Misaka T, Abe K and Kobayashi S: Luteolin and quercetin
affect the cholesterol absorption mediated by epithelial
cholesterol transporter niemann-pick c1-like 1 in caco-2 cells and
rats. PLoS One. 9:e979012014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bast A, Kaiserová H, den Hartog GJ, Haenen
GR and van der Vijgh WJ: Protectors against doxorubicin-induced
cardiotoxicity: Flavonoids. Cell Biol Toxicol. 23:39–47. 2007.
View Article : Google Scholar
|
|
35
|
Salkovic-Petrisic M and Hoyer S: Central
insulin resistance as a trigger for sporadic Alzheimer-like
pathology: an experimental approach. J Neural Transm Suppl.
72:217–233. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Grünblatt E, Salkovic-Petrisic M,
Osmanovic J, Riederer P and Hoyer S: Brain insulin system
dysfunction in streptozotocin intracerebroventricularly treated
rats generates hyperphosphorylated tau protein. J Neurochem.
101:757–770. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Maiese K and Chong ZZ: Insights into
oxidative stress and potential novel therapeutic targets for
Alzheimer disease. Restor Neurol Neurosci. 22:87–104.
2004.PubMed/NCBI
|
|
38
|
Behl C, Davis JB, Lesley R and Schubert D:
Hydrogen peroxide mediates amyloid beta protein toxicity. Cell.
77:817–827. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Varadarajan S, Yatin S, Kanski J,
Jahanshahi F and Butterfield DA: Methionine residue 35 is important
in amyloid beta-peptide-associated free radical oxidative stress.
Brain Res Bull. 50:133–141. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sharma M and Gupta YK:
Intracerebroventricular injection of streptozotocin in rats
produces both oxidative stress in the brain and cognitive
impairment. Life Sci. 68:1021–1029. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Selkoe DJ: Alzheimer's disease: Genes,
proteins, and therapy. Physiol Rev. 81:741–766. 2001.PubMed/NCBI
|
|
42
|
Rasoolijazi H, Joghataie MT, Roghani M and
Nobakht M: The beneficial effect of (−)-epigallocatechin-3-gallate
in an experimental model of Alzheimer's disease in rat: a
behavioral analysis. Iran Biomed J. 11:237–243. 2007.
|
|
43
|
Gilgun-Sherki Y, Rosenbaum Z, Melamed E
and Offen D: Antioxidant therapy in acute central nervous system
injury: current state. Pharmacol Rev. 54:271–284. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Subramaniam R, Koppal T, Green M, Yatin S,
Jordan B, Drake J and Butterfield DA: The free radical antioxidant
vitamin E protects cortical synaptosomal membranes from amyloid
beta-peptide (25–35)toxicity but not from hydroxynonenal toxicity:
relevance to the free radical hypothesis of Alzheimer's disease.
Neurochem Res. 23:1403–1410. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu D, Li M, Tian Y, Liu J and Shang J:
Luteolin inhibits ROS-activated MAPK pathway in myocardial
ischemia/reperfusion injury. Life Sci. 122:15–25. 2015. View Article : Google Scholar
|
|
46
|
Pratheeshkumar P, Son YO, Divya SP, Roy
RV, Hitron JA, Wang L, Kim D, Dai J, Asha P, Zhang Z, et al:
Luteolin inhibits Cr(VI)-induced malignant cell transformation of
human lung epithelial cells by targeting ROS mediated multiple cell
signaling pathways. Toxicol Appl Pharmacol. 281:230–241. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xia F, Wang C, Jin Y, Liu Q, Meng Q, Liu K
and Sun H: Luteolin protects HUVECs from TNF-α-induced oxidative
stress and inflammation via its effects on the Nox4/ROS-NF-κB and
MAPK pathways. J Atheroscler Thromb. 21:768–783. 2014. View Article : Google Scholar
|