Introduction
Gastric cancer (GC) is one of the most common types
of cancer worldwide. Since the improvement of neoadjuvant treatment
for GC, which combines surgery with neoadjuvant chemotherapy, the
survival rate of GC has increased; however, in patients with
metastatic disease the outcome is worse, with <30% survival
(1). The etiology of GC is
complex, thus resulting in the lack of an internationally accepted
standard early prevention regimen (2). Since distant metastasis is the
predominant pattern of GC recurrence, and is the most common cause
of cancer-associated mortality, it is important to clarify its
pathogenesis and to investigate the genes responsible for this
progress.
Epithelial-mesenchymal transition (EMT) is an
embryonic development program that is associated with changes in
cell morphology and increased expression of EMT-associated genes.
In cancer, EMT has been reported to confer motility and
invasiveness onto cancer cells, leading them to acquire the ability
to metastasize to distant sites (3,4).
Previous studies have demonstrated that aberrant EMT activation has
a crucial role in the genesis, invasion and metastasis of various
types of cancer (5,6), including GC (7). Ryu et al (8) analyzed numerous GC specimens and
reported that the majority of primary GC tumors, and even
premalignant lesions, exhibit a mesenchymal phenotype as
characterized by downregulation of CDH1 (E-cadherin), and
upregulation of zinc-finger E-box-binding homeobox factor-1
(ZEB1) and SNAI1 (Snail-1). In addition, ZEB1
and Snail-1 expression levels are positively associated with
expression of the cancer stem cell marker CD44 (8). Furthermore, a previous study
demonstrated that the expression of CDH2 (N-cadherin), which
is normally expressed in mesenchymal cells, was associated with the
invasive phenotype of GC, further suggesting the important role of
EMT in the initiation and progression of GC (9).
In addition to traditional transcriptional genes,
the role of non-coding microRNAs (miRNAs) on the regulation of EMT
has been widely studied (10).
miR-205 is a highly conserved miRNA among various species, which
has been reported to be closely associated with metastasis in
numerous types of cancer (11,12).
miR-205 is located in the second intron of the LOC642587 locus in
chromosome 1, and has been reported to have an important role in
orchestrating the morphogenesis of epithelium during embryogenesis
(13). miR-205 has been reported
to exhibit consistent overexpression in epidermis (14), whereas, in cells that have
undergone EMT progression, its expression is downregulated,
alongside a marked downregulation in E-cadherin, and an
upregulation in N-cadherin and fibronectin (15). Notably, a previous study reported
that by counteracting EMT, miR-205 expression is inversely
associated with the aggressive behavior of malignant mesothelioma
and suppresses its tumor proliferation and invasion (16). These results suggest that miR-205
may act as a suppressor of EMT, as well as a tumor suppressor in
cancer. A previous study reported that the expression levels of
miR-205 were significantly downregulated in GC tissue, as compared
with in normal gastric tissue, and was negatively associated with
the clinical and pathological characteristics of patients (17). However, the effects of miR-205 on
the metastasis and EMT progression of GC cells, and the underlying
molecular mechanisms, remain largely unknown.
The present study aimed to determine whether there
was a correlation between miR-205 expression and metastasis of
human GC cells. The results demonstrated that restored miR-205
expression resulted in a marked inhibition in the growth, migration
and invasion of GC cells. In addition, miR-205 suppressed the EMT
progression of GC cells, which may be due to the targeting of the
EMT-related transcriptional gene ZEB1.
Materials and methods
Cell culture and cell transfection
The NCI-H87 human GC cell line was obtained from the
American Type Culture Collection (Manassas, VA, USA). The cells
were cultured in Dulbecco's modified Eagle medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), supplemented
with 10% fetal bovine serum (FBS; PAA Laboratories GmbH, Pasching,
Austria), streptomycin (100 μg/ml; Sigma-Aldrich, St. Louis,
MO, USA) and penicillin (100 U/ml; Sigma-Aldrich) at 37°C in a
humidified atmosphere containing 5% CO2. miR-205 and
scramble mimic were purchased from GE Dharmacon (Lafayette, CO,
USA), and were transfected into the cells (1×105) at a
final concentration of 50 nM using DharmaFECT 1 (GE Dharmacon),
according to the manufacturer's protocol. The sequences were as
follows: miR-205 5′-GAUUUCAGUGGAGUGAAGUUC-3′; and scramble
5′-UCCUUCAUUCCACCGGAGUCUG-3′.
Cell Counting kit (CCK)-8 assay
In order to analyze cell proliferation, NCI-H87
cells were seeded into 24-well plates at 5×103
cells/well. The cells were incubated in 10% CCK-8 reagent (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) diluted in normal
culture medium at 37°C, until visual color conversion occurred. The
absorbance in each well was measured at 450 and 630 nm using a
microplate reader (Varioskan Flash; Thermo Fisher Scientific, Inc.)
at 0, 24, 48 and 72 h post-transfection.
Cell migration and invasion assays
A Transwell device containing 8 μm
microporous membranes (Corning, Inc., Corning, NY) was placed into
a 24-well plate. Normal NCI-H87 cells, or NCI-H87 cells
(4×105) transfected with miR-205 or scramble mimic were
seeded in the upper chamber alongside DMEM supplemented with 0.1%
bovine serum albumin (Sigma-Aldrich). DMEM supplemented with 10%
FBS served as chemoattractant. For the invasive assays, the upper
and lower chambers of the basal membrane were coated with 5 mg/ml
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). The rate of
migration/invasion was measured after 24 h. The cells adhering to
the lower surface were fixed and stained with 0.1% crystal violet
(Sigma-Aldrich), and transferred to a microscope slide. The total
number of invading cells was counted in six representative fields
under a microscope (BX51; Olympus Corporation, Tokyo, Japan)
(magnification, 200×).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Total RNA was then
reverse transcribed using the First-Strand cDNA Synthesis kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The specific primers
used for reverse transcription were as follows: miR-205,
5′-TTATTGCTTAAGAATACGCGTAG-3′; ZEB1,
5′-TTTTTTTTTTTTTTTTTT-3′; and U6, 5′-AAAATATGGAACGCTTCACGAATTTG-3′
(Tsingke Biotechnology, Co., Ltd., Beijing, China). Subsequently,
qPCR was performed using the QuantiTect SYBR Green PCR mixture
(Invitrogen; Thermo Fisher Scientific, Inc.) on an ABI PRISM 7900
Sequence Detection system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The primers used for qPCR were as follows:
miR-205, sense 5′-GCGCTTATTGCTTAAGAATAC-3′, anti-sense
5′-CAGTGCAGGGTCCGAGGT-3′; ZEB1, sense
5′-AAACTCGAGTACTTCAATTCCTCGGTATTG-3′, anti-sense
5′-AAATCTAGACACACTGTTCTACAGTCCAAGGC-3′; U6, sense
5′-CTCGCTTCGGCAGCACATATACT-3′, anti-sense
5′-ACGCTTCACGAATTTGCGTGTC-3′; and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH), sense 5′-TCAACGACCACTTTGTCAAGCTCA-3′,
and anti-sense 5′-GCTGGTGGTCCAGGGGTCTTACT-3′. The primers were
synthesized by Tsingke Biotechnology, Co., Ltd. The expression
levels of U6 and GAPDH were used as an internal control for
miRNA and mRNA expression, respectively. The PCR cycling conditions
were as follows: 94°C for 5 sec, followed by 40 cycles at 94°C for
5 sec and 60°C for 34 sec, and a final extension step at 72°C for
45 sec. PCR efficiency was calculated using a relative standard
curve derived from a cDNA (1 μl; 20 ng/μl) mixture,
and gave regression coefficients >0.95. The relative expression
levels were evaluated using the 2−∆∆Cq method (18). All experiments were repeated three
times, in order to reduce curve-derived variance.
Luciferase reporter assay
The whole 3′-untranslated region (UTR) of
ZEB1 was amplified in 293T cells from genomic DNA and cloned
into the pGL-3-vector (Promega Corporation, Madison, WI, USA)
immediately downstream of the Renilla luciferase gene. A
mutated 3′-UTR of ZEB1, in which the miR-205 target site was
deleted (Mut), was generated using the QuickChange Site-Directed
Mutagenesis kit (Agilent Technologies, Inc., Santa Clara, CA, USA).
NCI-H87 cells (1×105/well) were seeded into 24-well
plates 24 h prior to transfection. The cells were co-transfected
with 50 ng pGL-3 firefly luciferase reporter, 10 ng pRL-TK
Renilla luciferase reporter and 50 nM miR-205 or scramble
mimic using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). Cell lysates were prepared using Passive
Lysis Buffer (Promega Corporation) 48 h post-transfection, and
luciferase activity was measured using the Dual-Luciferase Reporter
Assay (Promega Corporation). Results were normalized to
Renilla luciferase.
Rescue assay
The full length ZEB1 gene open reading frame
(ORF) was amplified by PCR and cloned into a pCDNA-3.1 construct
(Promega Corporation), in order to generate the
pCDNA-3.1-ZEB1 construct. Briefly, ZEB1 ORF was
extracted for EcoRV and Xbal double digestion (New
England Biolabs, Inc., Ipswich, MA, USA), and purified gene
fragments were recovered. EcoRV and Xbal double
digestion of the pcDNA-3.1 expression vector was then performed.
The recovered target gene fragments were ligated into digested
pcDNA-3.1 expression vectors and identified using agarose gel
electrophoresis. An empty pCDNA-3.1 construct was used as the
control. The NCI-H87 cells were initially transfected with miR-205
or scramble mimic (60 nM) in 6-well plates. Following a 24 h
culture, the NCI-H87 cells were co-transfected with miR-205 mimic
(30 nM) and 2.0 μg of either pcDNA-3.1-ZEB1 or
pcDNA-3.1 constructs. The cells were harvested at predetermined
intervals and assessed as necessary.
Western blot analysis
For western blotting, the cells were harvested in
ice-cold phosphate-buffered saline 48 h post-transfection, and
lysed on ice in cold modified radioimmunoprecipitation buffer
(Beyotime Institute of Biotechnology, Haimen, China) supplemented
with protease inhibitors (Roche Diagnostics, Basel, Switzerland).
Protein concentration was determined using the Bicinchoninic Acid
Protein Assay kit (Vigorous Biotechnology Beijing Co., Ltd.,
Beijing, China) and equal amounts of protein (30 μg) were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. The gels were electroblotted onto nitrocellulose
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
subsequently blocked for 2 h with 5% non-fat dry milk in
Tris-buffered saline containing 0.1% Tween-20, and incubated at 4°C
overnight with primary antibody. Detection was performed using
alkaline phosphatase-conjugated anti-mouse (cat. no. 7056) and
anti-rabbit (cat. no. 7054) immunoglobulin G secondary antibodies
(1:5,000) and the blots were visualized using an enhanced
chemiluminescence system (EMD Millipore). The results of western
blotting were analyzed using Quantity One v4.6.2 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The primary antibodies used
were as follows: Rabbit monoclonal anti-human ZEB (1:1,000; cat.
no. 3396), mouse monoclonal anti-human E-cadherin (1:1,000; cat.
no. 14472), rabbit monoclonal anti-human vimentin (1:1,000; cat.
no. 5741), rabbit monoclonal anti-human N-cadherin (1:1,000; cat.
no. 13116) and rabbit monoclonal anti-human GAPDH (1:10,000; cat.
no. 5174), which was used as a negative control. All primary and
secondary antibodies were purchased from Cell Signaling Technology,
Inc. (Danvers, MA, USA).
Statistical analysis
All experiments were repeated at least three times.
Data are presented as the mean ± standard deviation of repeated
experiments. Statistical analysis was carried out using SPSS 15.0
software (SPSS Inc., Chicago, IL, USA). Two-tailed Student's t-test
was used to analyze the data. P<0.05 was considered to indicate
a statistically significant difference.
Results
miR-205 suppresses the proliferation,
migration and invasion of NCI-H87 cells
A previous study reported a downregulation of
miR-205 in GC tissue samples (17); however, its biological significance
on cancer progression remains unclear. To further explore the
effects of miR-205 on the malignant phenotype of GC cells, NCI-H87
cells in which miR-205 was underexpressed (17), were exogenously transfected with
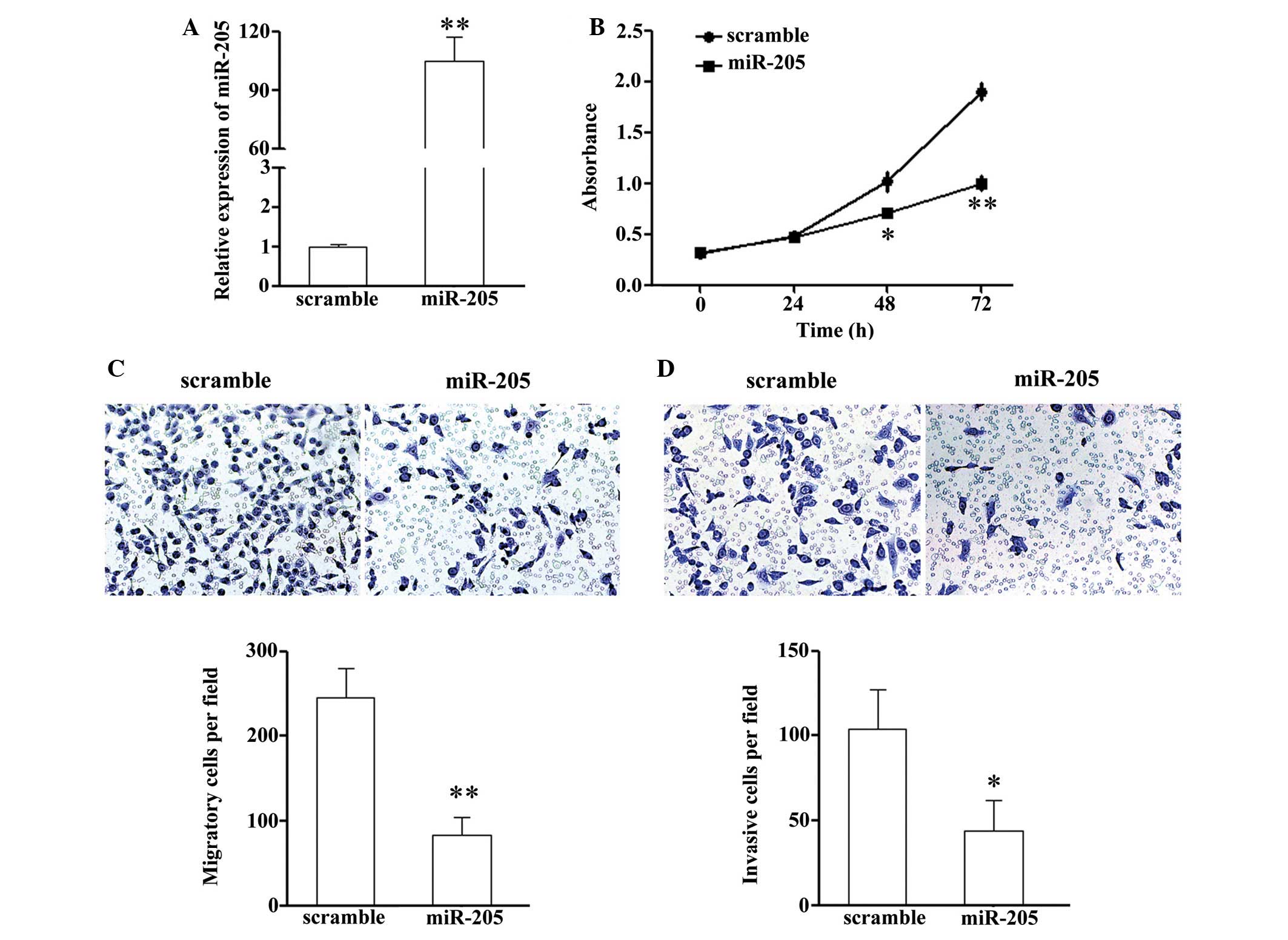
miR-205 or a scramble mimic. Upon transfection, the intracellular
expression levels of miR-205 were ~100-fold higher the NCI-H87
cells transfected with the miR-205 mimic, as compared with the
scramble control group (Fig. 1A).
In addition, the effects of miR-205 on cell proliferation were
determined using a CCK-8 assay. As shown in Fig. 1B, treatment with miR-205
significantly suppressed the cell growth rate of the NCI-H87
cells.
Since miR-205 is closely associated with tumor
metastasis, the present study hypothesized that miR-205 may have an
important role in GC cell migration and invasion, which promote
tumor metastasis, giving rise to GC-associated mortality.
Therefore, the present study explored the effects of miR-205 on the
migration and invasion of NCI-H87 cells using a Transwell assay. A
Transwell assay without Matrigel demonstrated that overexpression
of miR-205 in NCI-H87 cells resulted in a significant reduction in
the number of cells that passed through the chambers, as compared
with the scramble control group (240±45 vs. 85±15 cells)
(P<0.05; Fig. 1C).
Subsequently, the chambers were coated with Matrigel, which mimics
the extracellular matrix, prior to experimentation. The invasion
assay exhibited similar results to the migration assay. As shown in
Fig. 1D, miR-205 overexpression
significantly reduced the number of NCI-H87 cells that passed
through the chambers (104±26 vs. 42±18 cells) (P<0.05). These
results indicate that miR-205 may efficiently suppress the motility
and invasiveness of GC cells in vitro.
miR-205 promotes an epithelial phenotype
in GC cells
Since miR-205 can inhibit gastric cancer cell
migration and invasion, the present study hypothesized that it may
be associated with the inhibition of EMT progression in GC cells.
Since EMT is often associated with a decrease or loss of epithelial
markers, including E-cadherin, and a gain of mesenchymal markers,
including vimentin and N-cadherin, the present study detected the
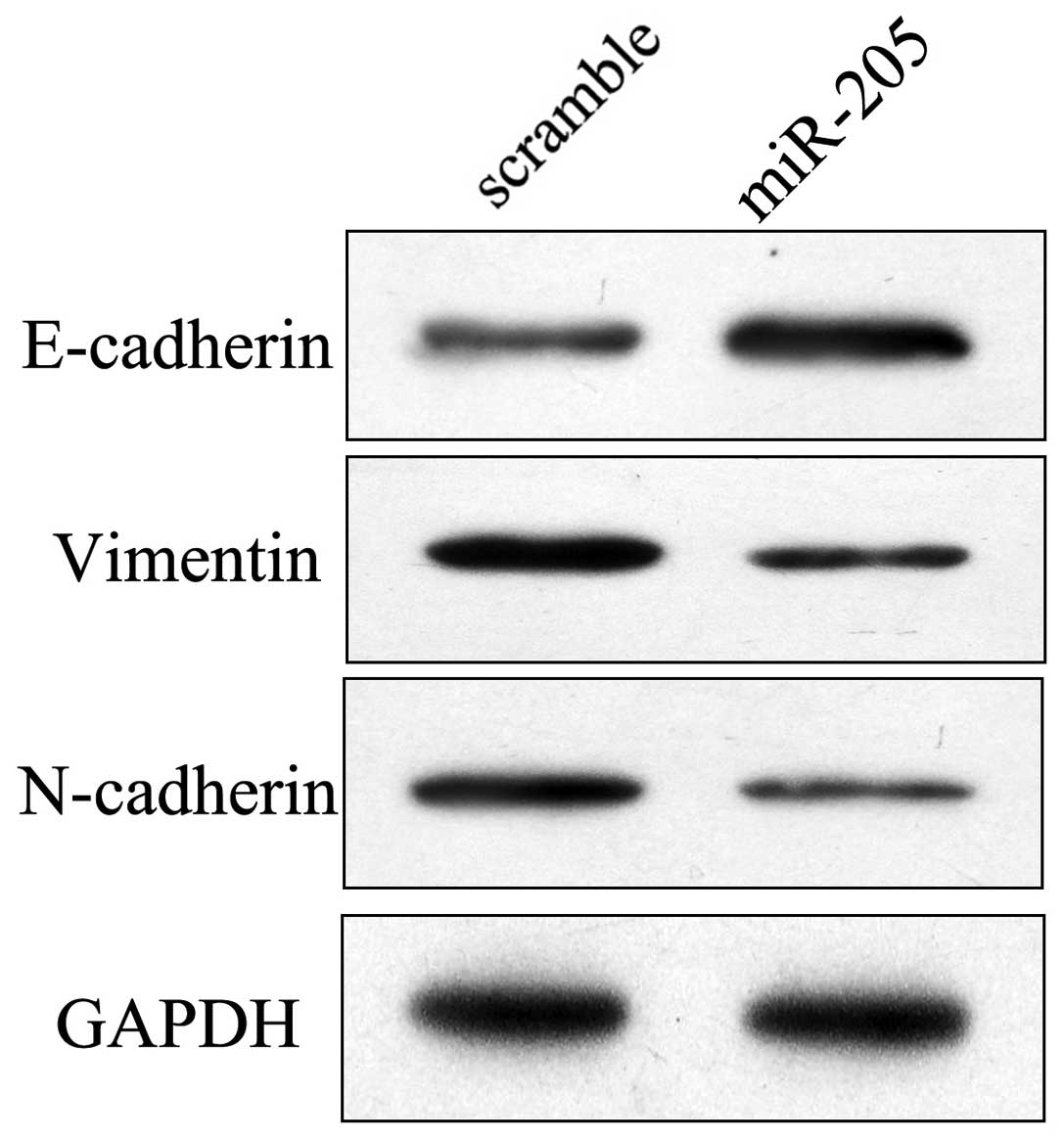
molecular alterations in cells overexpressing miR-205. The protein
expression levels of mesenchymal and epithelial markers were
detected in the NCI-H87 cells. As shown in Fig. 2, the mesenchymal markers vimentin
and N-cadherin were consistently suppressed in NCI-H87 cells
treated with the miR-205 mimic. However, the epithelial marker
E-cadherin was markedly upregulated following transfection with
miR-205. These results suggest that overexpression of miR-205 may
induce an epithelial phenotype in GC cells.
ZEB1 is a putative target gene of miR-205
in GC cells
To explore the target genes associated with GC tumor
progression triggered by miR-205, putative targets of miR-205 were
searched using prediction programs (Targetscan, http://genes.mit.edu/targetscan/ and miRanda,
http://www.microrna.org/microrna/home.do). Among the
common predicted targets of miR-205, ZEB1 was selected as an
ideal candidate due to its important role in EMT (15). ZEB1 has previously been
reported as a target of miR-205 in breast cancer (19); however, the interaction between
miR-205 and ZEB-1 has not been experimentally validated in
GC.
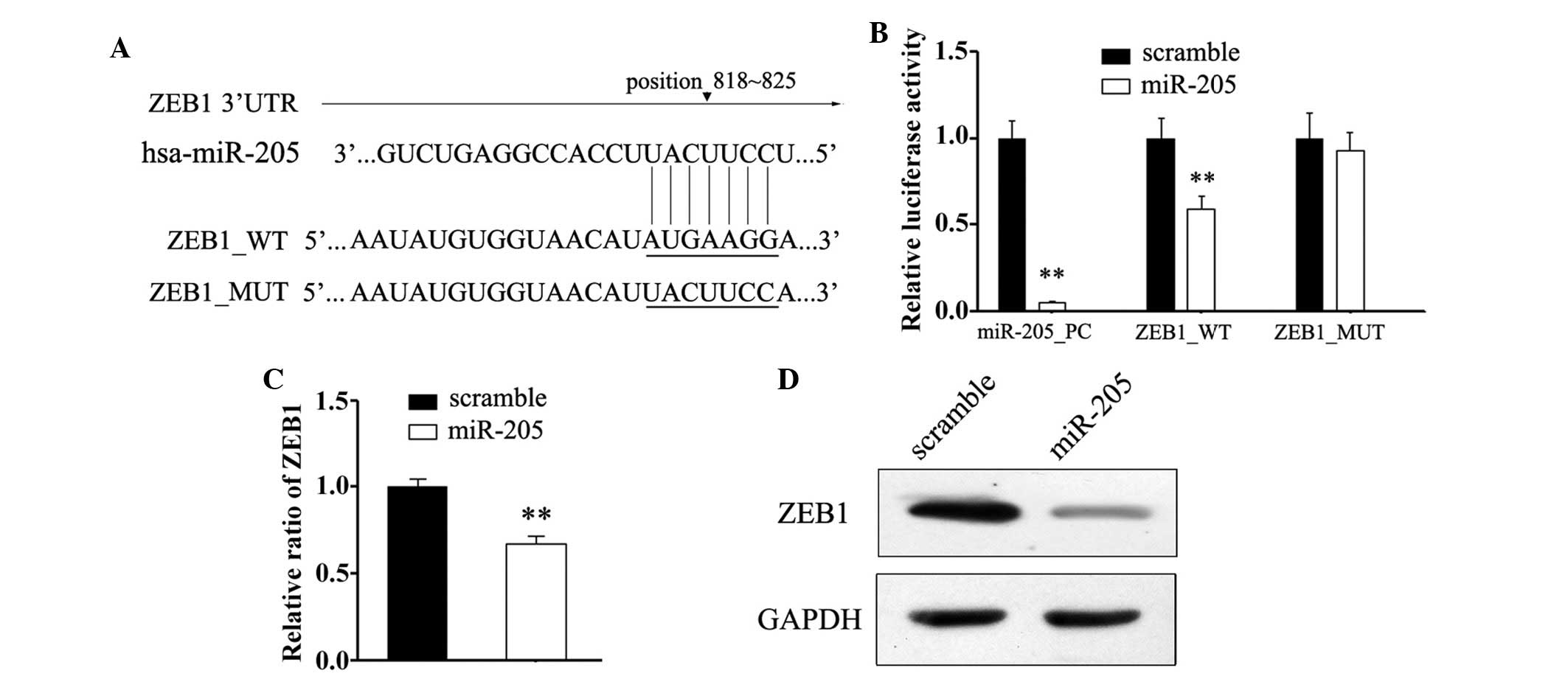
To confirm miR-205 binding within the 3′-UTR of
ZEB1, a mutated 3′-UTR of ZEB1, in which the miR-205
target site was deleted, was generated (Fig. 3A). Subsequently, the effects of Mut
and wild type 3′UTR constructs on NCI-H87 cells overexpressing
miR-205 were determined using a dual-luciferase detection system.
As a result, significant suppression of luciferase activities were
observed in the NCI-H87 cells co-transfected with the wild type
3′UTR construct and miR-205 mimic, as compared with the Mut
construct groups (Fig. 3B). These
results suggest that miR-205 may suppress the transcriptional
activity of the ZEB1 gene by targeting the binding site in
the 3′UTR of ZEB1 mRNA. Consistent with the reporter assays,
transfection with miR-205 decreased the mRNA expression levels of
ZEB1, as compared with in the scramble group (Fig. 3C). In addition, according to
immunoblotting results, changes were detected in the protein
expression levels of ZEB1 post-transfection with a miR-205 mimic.
These results indicate that miR-205 may directly target the
expression of ZEB1 in GC cells.
ZEB1 is associated with the
miR-205-mediated suppression of proliferation and EMT
progression
ZEB1 is a transcriptional inducer of EMT in
cancer of epithelial origin, and has been reported to have a key
role in tumor metastasis (20).
However, whether it is involved in miR-205-mediated suppression of
migration and EMT progression in NCI-H87 cells remains unclear.
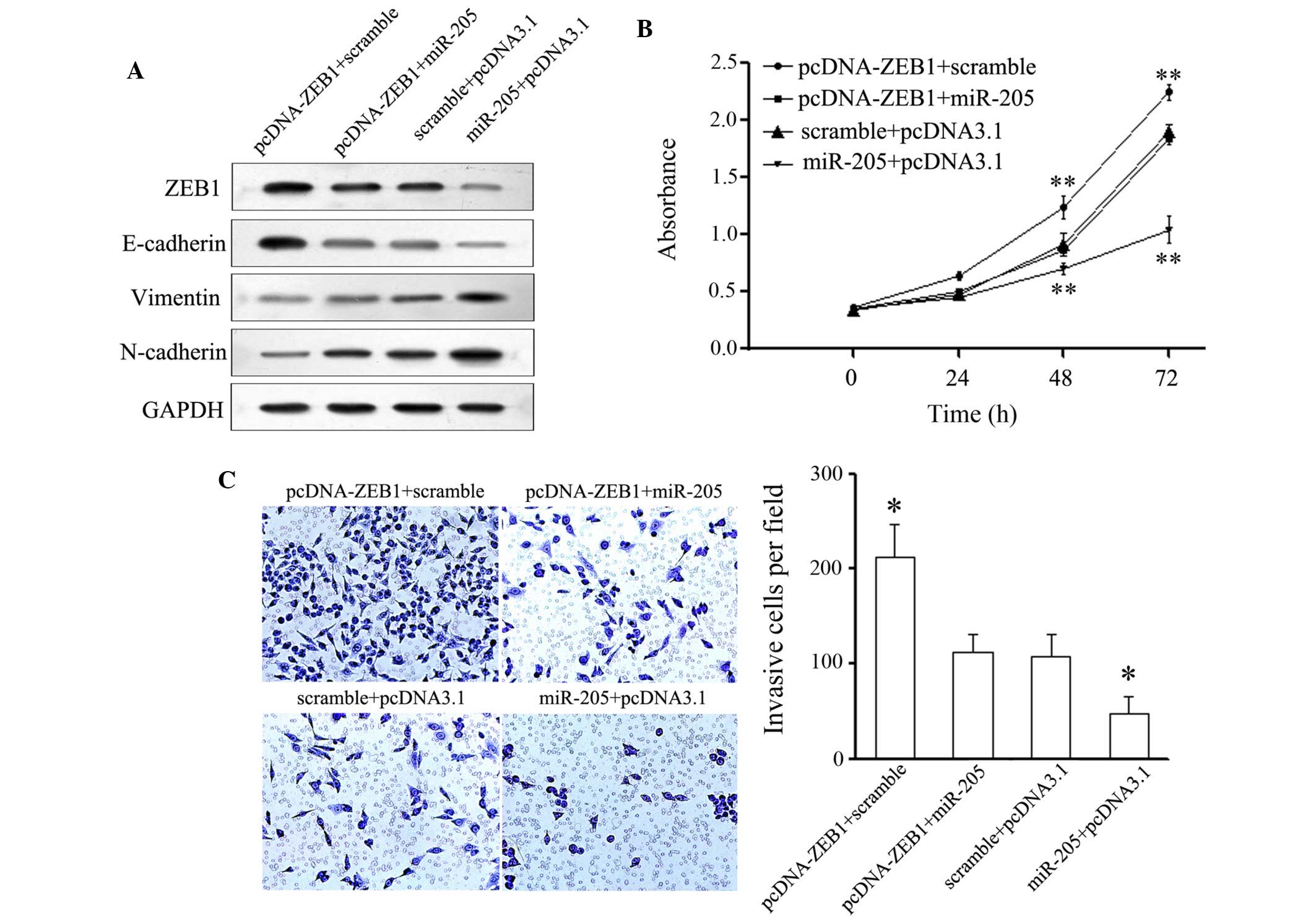
Therefore, a 'rescue' methodology was adopted to examine the
functional relevance of miR-205-ZEB1 interaction in NCI-H87
cells. A novel construct containing the full ORF of ZEB1 was
generated. Subsequently, NCI-H87 cells were co-transfected with
miR-205 or a scramble mimic alongside pcDNA-3.1-ZEB1 or
pcDNA-3.1 control constructs. Post-transfection, the expression
levels of ZEB1 were restored when the ZEB1 construct
was transfected into the NCI-H87 cells that had been transfected
with a miR-205 mimic for 24 h (Fig.
4A). In agreement with the restored expression of ZEB1,
increased cell proliferation was observed in the NCI-H87 cells
transfected with the ZEB1 construct following transfection
with the miR-205 mimic (Fig. 4B).
Furthermore, post-transfection with the ZEB1 construct, the
miR-205-mediated suppression of cell invasion (Fig. 4C) in NCI-H87 cells was also
partially attenuated. The number of invasive cells in the
pcDNA-ZEB1 + miR-205 and scramble + pcDNA3.1 groups were
significantly reduced, as compared with in the scramble +
pcDNA-ZEB1group, and were significantly increased as
compared with in the miR-205 + pcDNA3.1 group. Consistent with the
restored expression of ZEB1, suppression of N-cadherin was
restored, and the upregulation of E-cadherin was partially
attenuated (Fig. 4A). These
results indicate that ZEB1 may be a functional target of
miR-205, contributing to its role in the miR-205-mediated
suppression of cell invasion and EMT progression in GC cells.
Discussion
EMT is a fundamental process in embryonic
development, which is also considered an important step leading to
tumor invasion and metastasis (21). At present, previously unknown
markers, miRNAs, are considered to be important components of the
cancer signaling network and are emerging as novel biomarkers of
numerous diseases (22). miRNAs
are a group of endogenous, small, non-coding RNAs that modulate
protein expression by regulating the translational efficiency or
cleavage of targets (23). By
partially complementing the 3′-UTR of specific mRNAs, miRNAs induce
the genetic silencing of various target mRNAs, which are involved
in numerous biological processes, including EMT regulation
(24). Previous studies have
reported the important role of miRNAs in the initiation and
progression of GC (25–27), and numerous tumor-associated
circulating miRNAs have been reported to possess potential as
novel, non-invasive biomarkers for the early detection of GC
(28). Therefore, improved
knowledge regarding alterations in miRNA expression during GC
progression and metastasis may provide novel options for the
diagnosis and treatment of GC. The present study provided important
evidence in support of miR-205 functioning as a tumor suppressor in
GC.
miR-205 is a highly conserved gene among various
species, which has been closely associated with metastasis in
numerous types of cancer (11,12).
In melanoma specimens, E2F transcription factor 1 (E2F1) is
negatively regulated by miR-205. Overexpression of miR-205 leads to
a mediation of E2F1-regulated Akt phosphorylation and an
upregulation of p16-INK4A, resulting in the suppression of cell
proliferation (29). Furthermore,
by counteracting EMT, miR-205 expression is inversely associated
with the aggressive behavior of malignant mesothelioma and is able
to suppress its tumor proliferation and invasion (16). A previous study reported that the
expression levels of miR-205 were significantly downregulated in GC
tissue, as compared with in normal gastric tissue, and was
negatively associated with the clinical and pathological
characteristics of patients (17).
In addition, inhibition of miR-205 significantly promoted the
proliferation of GC cells, thus suggesting the suppressive role of
miR-205 in GC. However, the effects of miR-205 on metastasis and
EMT progression of GC cells, and the underlying molecular
mechanisms, remain largely unknown. The present study demonstrated
that restored expression of miR-205 in the NCI-H87 GC cell line
resulted in inhibition of cell proliferation, migration and
invasion. Alongside suppressed cell invasion, miR-205 suppressed
the expression of epithelial markers and upregulated the expression
of mesenchymal markers in GC cells. The present study further
confirmed the effects of miR-205 on the biological function of GC
cells, thus suggesting the tumor suppressor role of miR-205 in
GC.
To further identify the mechanisms underlying the
suppressive effects of miR-205, the putative targets of miR-205
were explored. Among these genes, ZEB1 was selected.
ZEB1, which is a member of the ZEB family, is a
transcriptional repressor that mediates its binding to paired
CAGGTA/G E-box-like promoter elements (30). Through suppressing the expression
of E-cadherin ZEB1 induces EMT and contributes to the
progression of malignant cancer (31). In addition, previous studies have
demonstrated that ZEB1 expression is positively correlated
with drug resistance in cancer cells (19), and ZEB1 knockdown was able
to chemosensitize pancreatic cancer cells to conventional
chemotherapy drugs, including gemcitabine, 5-fluorouracil and
cisplatin (32). The results of
the present study demonstrated that alterations in miR-205
expression in GC cells led to the opposite effects associated with
ZEB1 alterations, highlighting its negative regulation.
Furthermore, the ZEB1 mRNA 3′-UTR bears a binding site of
miR-205, and transfection with miR-205 resulted in the suppression
of the mRNA and protein expression levels of ZEB1.
To further analyze whether ZEB1 has an
important role in miR-205-mediated suppression of cell
proliferation and invasion, further rescue assays were performed.
It was suggested that miR-205 inhibited EMT progression via
targeting ZEB1 in GC cells, since restored expression of
ZEB1 could partially attenuate miR-205-mediated
downregulation of N-cadherin and up-regulation of E-cadherin.
Consistent with the suppression of EMT, the suppression of
proliferation and invasion were also partially restored. However,
overexpression of ZEB1 could not completely abolish
miR-205-mediated tumor suppression, thus suggesting that other
target genes may be involved in the suppression of GC. Therefore,
further research is required, in order to elucidate the exact
mechanisms underlying miR-205-mediated functions in GC cells. As
more information regarding the mechanisms is obtained, the
opportunity to manipulate them in cancer in order to suppress tumor
metastasis may arise.
In conclusion, the results of the present study
indicated that aberrant expression of miR-205 may have a role in
the tumor progression and prognosis of patients with GC.
Furthermore, these data suggested that miR-205 may function as a
tumor suppressor, and may modulate GC cell proliferation, invasion
and EMT progression by directly and negatively regulating
ZEB1. Therefore, the restored expression of miR-205 may be
considered a potential therapeutic strategy for the treatment of
GC.
Acknowledgments
The present study was supported by funds provided by
the Science and Technology Plan of Shandong University (grant no.
J11LF66).
References
|
1
|
Gill RS, Al-Adra DP, Nagendran J, Campbell
S, Shi X, Haase E and Schiller D: Treatment of gastric cancer with
peritoneal carcinomatosis by cytoreductive surgery and HIPEC: A
systematic review of survival, mortality, and morbidity. J Surg
Oncol. 104:692–698. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kanat O and O'Neil BH: Metastatic gastric
cancer treatment: A little slow but worthy progress. Med Oncol.
30:4642013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Klymkowsky MW and Savagner P:
Epithelial-mesenchymal transition: A cancer researcher's conceptual
friend and foe. Am J Pathol. 174:1588–1593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Montemayor-Garcia C, Hardin H, Guo Z,
Larrain C, Buehler D, Asioli S, Chen H and Lloyd RV: The role of
epithelial mesenchymal transition markers in thyroid carcinoma
progression. Endocr Pathol. 24:206–212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liang Q, Li L, Zhang J, Lei Y, Wang L, Liu
DX, Feng J, Hou P, Yao R, Zhang Y, et al: CDK5 is essential for
TGF-β1-induced epithelial-mesenchymal transition and breast cancer
progression. Sci Rep. 3:29322013. View Article : Google Scholar
|
|
7
|
Zhao L, Li W, Zang W, Liu Z, Xu X, Yu H,
Yang Q and Jia J: JMJD2B promotes epithelial-mesenchymal transition
by cooperating with β-catenin and enhances gastric cancer
metastasis. Clin Cancer Res. 19:6419–6429. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ryu HS, Park do J, Kim HH, Kim WH and Lee
HS: Combination of epithelial-mesenchymal transition and cancer
stem cell-like phenotypes has independent prognostic value in
gastric cancer. Hum Pathol. 43:520–528. 2012. View Article : Google Scholar
|
|
9
|
Hazan RB, Qiao R, Keren R, Badano I and
Suyama K: Cadherin switch in tumor progression. Ann NY Acad Sci.
1014:155–163. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Garg M: Targeting microRNAs in
epithelial-to-mesenchymal transition-induced cancer stem cells:
Therapeutic approaches in cancer. Expert Opin Ther Targets.
19:285–297. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kalogirou C, Spahn M, Krebs M, Joniau S,
Lerut E, Burger M, Scholz CJ, Kneitz S, Riedmiller H and Kneitz B:
MiR-205 is progressively down-regulated in lymph node metastasis
but fails as a prognostic biomarker in high-risk prostate cancer.
Int J Mol Sci. 14:21414–21434. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tucci P, Agostini M, Grespi F, Markert EK,
Terrinoni A, Vousden KH, Muller PA, Dötsch V, Kehrloesser S, Sayan
BS, et al: Loss of p63 and its microRNA-205 target results in
enhanced cell migration and metastasis in prostate cancer. Proc
Natl Acad Sci USA. 109:15312–15317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yi R, O'Carroll D, Pasolli HA, Zhang Z,
Dietrich FS, Tarakhovsky A and Fuchs E: Morphogenesis in skin is
governed by discrete sets of differentially expressed microRNAs.
Nat Genet. 38:356–362. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ason B, Darnell DK, Wittbrodt B, Berezikov
E, Kloosterman WP, Wittbrodt J, Antin PB and Plasterk RH:
Differences in vertebrate microRNA expression. Proc Natl Acad Sci
USA. 103:14385–14389. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fassina A, Cappellesso R, Guzzardo V,
Dalla Via L, Piccolo S, Ventura L and Fassan M:
Epithelial-mesenchymal transition in malignant mesothelioma. Mod
Pathol. 25:86–99. 2012. View Article : Google Scholar
|
|
17
|
Yin WZ, Li F, Zhang L, Ren XP, Zhang N and
Wen JF: Down-regulation of microRNA-205 promotes gastric cancer
cell proliferation. Eur Rev Med Pharmacol Sci. 18:1027–1032.
2014.PubMed/NCBI
|
|
18
|
Schmittgen TD, Zakrajsek BA, Mills AG,
Gorn V, Singer MJ and Reed MW: Quantitative reverse
transcription-polymerase chain reaction to study mRNA decay:
Comparison of endpoint and real-time methods. Anal Biochem.
285:194–204. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee JY, Park MK, Park JH, Lee HJ, Shin DH,
Kang Y, Lee CH and Kong G: Loss of the polycomb protein Mel-18
enhances the epithelial-mesenchymal transition by ZEB1 and ZEB2
expression through the downregulation of miR-205 in breast cancer.
Oncogene. 33:1325–1335. 2014. View Article : Google Scholar
|
|
20
|
Wellner U, Schubert J, Burk UC,
Schmalhofer O, Zhu F, Sonntag A, Waldvogel B, Vannier C, Darling D,
zur Hausen A, et al: The EMT-activator ZEB1 promotes tumorigenicity
by repressing stemness-inhibiting microRNAs. Nat Cell Biol.
11:1487–1495. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen K and Rajewsky N: The evolution of
gene regulation by transcription factors and microRNAs. Nat Rev
Genet. 8:93–103. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hui A, How C, Ito E and Liu FF: Micro-RNAs
as diagnostic or prognostic markers in human epithelial
malignancies. BMC Cancer. 11:5002011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yanaka Y, Muramatsu T, Uetake H, Kozaki K
and Inazawa J: miR-544a induces epithelial-mesenchymal transition
through the activation of WNT signaling pathway in gastric cancer.
Carcinogenesis. 36:1363–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Peng Y, Jin Z, Huang Q, Cheng Y,
Liu Y, Feng X, Yang M, Huang Y, Zhao Z, et al: Integrated miRNA
profiling and bioinformatics analyses reveal potential causative
miRNAs in gastric adenocarcinoma. Oncotarget. 6:32878–32889.
2015.PubMed/NCBI
|
|
28
|
Blanco-Calvo M, Calvo L, Figueroa A,
Haz-Conde M, Anton-Aparicio L and Valladares-Ayerbes M: Circulating
microRNAs: Molecular microsensors in gastrointestinal cancer.
Sensors (Basel). 12:9349–9362. 2012. View Article : Google Scholar
|
|
29
|
Dar AA, Majid S, de Semir D, Nosrati M,
Bezrookove V and Kashani-Sabet M: miRNA-205 suppresses melanoma
cell proliferation and induces senescence via regulation of E2F1
protein. J Biol Chem. 286:16606–16614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brabletz S and Brabletz T: The ZEB/miR-200
feedback loop - a motor of cellular plasticity in development and
cancer? EMBO Rep. 11:670–677. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gheldof A, Hulpiau P, van Roy F, De Craene
B and Berx G: Evolutionary functional analysis and molecular
regulation of the ZEB transcription factors. Cell Mol Life Sci.
69:2527–2541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arumugam T, Ramachandran V, Fournier KF,
Wang H, Marquis L, Abbruzzese JL, Gallick GE, Logsdon CD, McConkey
DJ and Choi W: Epithelial to mesenchymal transition contributes to
drug resistance in pancreatic cancer. Cancer Res. 69:5820–5828.
2009. View Article : Google Scholar : PubMed/NCBI
|