Introduction
The balance between the formation of osteoblasts and
osteoclast resorption is key in bone remodeling. An imbalance
between the osteoclasts and osteoblasts leads to various
bone-related disorders, such as osteoporosis, osteomalacia and
osteopetrosis (1,2). Osteoclasts are derived from
hematopoietic cells of the monocyte/macrophage lineage that are
responsible for bone resorption. Receptor activator of nuclear
factor-κB (RANK) and its cognate ligand (RANKL) are a tumor
necrosis factor (TNF) superfamily receptor-ligand pair that are
essential in bone remodeling, immune function and mammary gland
development, in conjunction with various cytokines and hormones
(3–6). RANKL is a type II homotrimeric
transmembrane protein and it is the central regulator of the
differentiation, activation, and survival of osteoclasts which are
the primary cells for physiological and pathological bone
resorption (7). RANKL-induced
activation of RANK on the surface of osteoclast progenitor cells
leads to the stimulation of TNF receptor-associated factors (TRAFs)
and the subsequent activation of several downstream signaling
molecules, including nuclear factor-κB (NF-κB), mitogen-activated
protein kinases (MAPKs), activating protein 1 (AP-1), nuclear
factor of activated T cells cytoplasmic, calcineurin-dependent 1
(NFATc1), and phosphatidylinositol 3-kinase, resulting in the
differentiation of osteoclast progenitor cells into cells that
finally fuse to form multinucleated bone-resorbing osteoclasts
(8,9). These signaling pathways ultimately
lead to the induction and activation of the transcription factors
involved in the expression of genes that characterize
osteoclasts.
Recent studies have revealed that NFATc1 is a
critical transcription factor in osteoclastogenesis. NFAT is a
calcineurin and calcium-regulated transcription factor. The release
of Ca2+ activates the calmodulin-regulated phosphatase
calcineurin, which binds to the N-terminal domain of NFATc1 and
dephosphorylates it (10,11). NFATc1-deficient embryonic stem
cells can not differentiate into osteoclasts. The over-expression
of constitutively active NFATc1 in bone marrow
monocytes/macrophages causes precursor cells to undergo efficient
differentiation even in the absence of RANKL. Suggesting that
NFATc1 is important for osteoclastogenesis (10–13).
NFATc1 undergoes nuclear translocation and regulates the expression
of a number of osteoclast specific genes, such as cathepsin K,
TRAP, calcitonin receptor as well as NFATc1 itself (12). The expression of NFATc1 appeared to
be induced by RANKL stimulation, and the suppression or knockdown
of NFATc1 inhibits RANKL-induced osteoclastogenesis in vitro
(13). c-Src is critical to the
function of osteoclasts, but not for osteoclast formation (14). Osteoclasts deficient in c-Src do
not develop normal podosomes, cannot give rise to mature
superstructures and fail to move properly (15). In addition, osteoclasts deficient
in c-Src exhibit reduced motility and abnormal organization of the
ruffle border and lack the cytoskeletal elements which are required
for bone resorption.
Our previous study showed that
RANK/RANKL/osteoprotegerin (OPG) may modulate the process of
osteoclast activation and subsequent bone resorption (16). However, the different
concentrations of RANKL impact NFATc1 and c-Src expression in
osteoclast-like cells remains unknown. In this study, RANKL
regulation of the gene expression of NFATc1 and c-Src in
osteoclast-like cell cultures derived from the bone marrow of rats
and the underlying molecular mechanism were investigated.
Materials and methods
Animals
Five-week-old female Sprague Dawley rats, weighing
120–130 g, were purchased from the Medical College of Xiamen
University Laboratory Animal Center (Xiamen, China; certificate no.
2007–0005). The rats were maintained at 22±2°C and 60% humidity
with good ventilation, an alternate 12 h natural light-dark cycle,
and unlimited access to standard rat food and tap water. This study
was approved by the medical ethics committee of the Zhongshan
Hospital of Xiamen University and conformed to National Institutes
of Health guidelines regarding animal experimentation.
Reagents
The 387A-1 kit used for cytological staining,
obtained from Sigma-Aldrich (St. Louis, MO, USA). α-minimum
essential medium (α-MEM) was obtained from Hyclone (Logan, UT,
USA). Fetal bovine serum (FBS) was obtained from (Biological
Industries, Beit-Haemek, Israel). Soluble recombinant rat RANKL and
M-CSF were obtained from PeproTech (Rocky Hill, NJ, USA). TRIzol
was obtained from Invitrogen, Thermo Fisher Scientific Inc.
(Waltham, MA, USA). DNA markers were obtained from TIANGEN Biotech
(Beijing) Co., Ltd. (Beijing, China). The RevertAid First Strand
cDNA Synthesis kit was obtained from Thermo Fisher Scientific Inc.
Primer sequences were obtained from Invitrogen, Thermo Fisher
Scientific Inc. All other chemicals were obtained from
Sigma-Aldrich.
Osteoclast-like cell formation in rat
marrow culture
Rat bone marrow cells were harvested as previously
described (16–18) with minor modifications. Briefly,
rat bone marrow cells were harvested 12 weeks after ovariectomy by
a previous method (17) with minor
modifications. For superficial antisepsis, the sacrificed rats by
cervical dislocation and decapitation without anesthesia and were
soaked twice for 5 min in 1% iodophors liquid (Shanghai Likang
Disinfection Technology Co., Ltd., Shanghai, China). Deiodination
was completed with 75% alcohol. After rapid removal of soft tissues
under aseptic conditions, the femors and tibias were cleaned twice
with phosphate-buffered saline (PBS, pH 7.4). They were then rinsed
twice with α-minimum essential medium (Invitrogen, Thermo Fisher
Scientific, Inc.). Bone marrow cells were obtained from the femors
and tibias of the Sprague Dawley rats by flushing the marrow space
with α-MEM containing 15% FBS. The cells were then seeded into
6-well chamber slides (Nalge Nunc International, Naperville, IL,
USA) at 2 ml cell suspension/well in α-MEM containing FBS. The bone
slices were used for observe bone resorptive lacunae. The glass
slides were used for TRAP staining. After 9 days of culture with
osteoclast-like cells, the bone slices and glass slides were
removed for observation. After 5 h, non-adherent cells were removed
with α-MEM. Half the medium was replaced every day with fresh
medium. Bone marrow cells were cultured in α-MEM with 10% FBS
containing 25 ng/ml M-CSF. After 24 h, non-adherent cells were
collected and plated on a 6-well microplate at a density of
1×106 cells/cm2 and cultured for up to 9 days
in α-MEM containing 10% FBS with 25 ng/ml M-CSF and 0, 50, 75 or
100 ng/ml RANKL.
Tartrate-resistant acid phosphatase
(TRAP) assays
Osteoclast formation was evaluated using TRAP
staining with the formation of multiple nuclei. After 72 h of
culture, growing cells were removed and dried at room temperature.
The cells were fixed in 2.5% (v/v) glutaraldehyde (Shanghai
Ponsurebio Co., Ltd., Shanghai, China) at 4°C for 10 min. TRAP
staining fluid (Sigma-Aldrich) was added, and the plates were
incubated at 37°C for an additional 50 min. After the removal of
the TRAP solution using a transferpettor, the plates were washed
three times with distilled water and air-dried. The slices were
sealed with glycerogelatin. Osteoclast-like cell morphology,
including cellular configuration, size, and nuclei, was observed
under a microscope (BX40, Olympus, Tokyo, Japan).
Scanning electron microscopy of
osteoclasts
Areas of the resorption pits in the bone slices
formed by the osteoclasts may reflect the resorption activity. The
bone slices were removed and cleaned ultrasonically three times for
10 min each in a 0.25 M NH4OH solution to remove
adherent cells. They were then rinsed with PBS, fixed in 2.5% (v/v)
glutaraldehyde (Sigma-Aldrich), postfixed in 1% osmium tetroxide
(Sigma-Aldrich), dehydrated with a graded alcohol series, and
placed in alcohol with isopentyl acetate. Finally, the bone slice
samples were subjected to CO2 critical-point drying and
gold staining, and the resorptive lacunae were observed by scanning
electronic microscopy (JSM-6380LV, Jeol, Tokyo, Japan)
Experimental model
Marrow cells were harvested from the femors and
tibias of the rat and cultured in 6-well chamber slides. The marrow
cells were cultured in α-MEM containing 10% FBS, 1%
penicillin-streptomycin (North China Pharmaceutical Co., Ltd.,
Shijiazhuang, China), and M-CSF (25 ng/ml). After 24 h,
non-adherent cells were collected and seeded into 6-well culture
plates (1×106cells/well). Osteoclast-like cells were
randomly and equally divided into the M-CSF (25 ng/ml) group, M-CSF
(25 ng/ml)+RANKL (50 ng/ml) group, M-CSF (25 ng/ml)+RANKL (75
ng/ml) group and M-CSF (25 ng/ml)+RANKL (100 ng/ml) group and
cultured in 6-well plates for 9 days. NFATc1, c-Src and RANK mRNA
assays were subsequently performed on the 9th day.
Reverse transcription-quantitative
polymerase chain reaction analysis
Total RNA was isolated using TRIzol reagent
(Invitrogen, Thermo Fisher Scientific Inc.). First-strand cDNAs
were synthesized from 2 μg of total RNA in 20 μl
reactions using the RevertAid first strand cDNA synthesis kit
(Thermo Fisher Scientific Inc.). Each sample was analyzed in
triplicate. The light cycler-fast start DNA master SYBR green
(Roche, Thermo Fisher Scientific Inc.) was used according to the
manufacturer's instructions. qPCR was conducted in a real-time PCR
cycler (ABI7500, Applied Biosystems, Thermo Fisher Scientific
Inc.). Primers were synthesized by Sangon Biotech Shanghai, Co.,
Ltd. (Shanghai, China). The primer sequences, product size, and
cycling conditions are shown in Table
I. The relative mRNA levels of NFATc1, RANK and c-Src were
normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH)
mRNA. Four samples from each group were included in each
experiment, and experiments were conducted at least three times.
All reactions were run in triplicate, and data were analyzed using
the 2−ΔΔCq method (19).
 | Table IPrimer sequences, product size, and
condition for reverse transcription-quantitative polymerase chain
reaction analysis. |
Table I
Primer sequences, product size, and
condition for reverse transcription-quantitative polymerase chain
reaction analysis.
| Gene | Primer sequences
(5′–3′) | Conditions | Product size
(bp) |
|---|
| NFATc1 | F:
TTGGATTCTGACGAGCTGTG | (30) 95°C, 15 sec;
60°C, 60 sec; 60°C, 60 sec | 209 |
| R:
GTGCAGCTGGATCAAGAACA | | |
| c-Src | F:
ACTGTCACCT-CCCCACAAAG | (30) 95°C, 15 sec;
60°C, 60 sec; 60°C, 60 sec | 153 |
| R:
GACATTCCACCTTCCTCGTGT | | |
| RANK | F:
GCCAGCAAGAAGTGTGTGAA | (30) 95°C, 15 sec;
60°C, 60 sec; 60°C, 60 sec | 212 |
| R:
CCGGTCCGTGTACTCATCT | | |
| GAPDH | F:
AGACAGCCGCATCTTCTTGT | (30) 95°C, 15 sec;
60°C, 60 sec; 60°C, 60 sec | 207 |
| R:
CTTGCCGTGGGTAGAGTCAT | | |
Statistical analysis
All values were expressed as the mean ± standard
deviation. One-way analysis of variance followed by the Least
Significant Difference multiple comparison procedure was performed
using SigmaStat 17.0 (SPSS Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
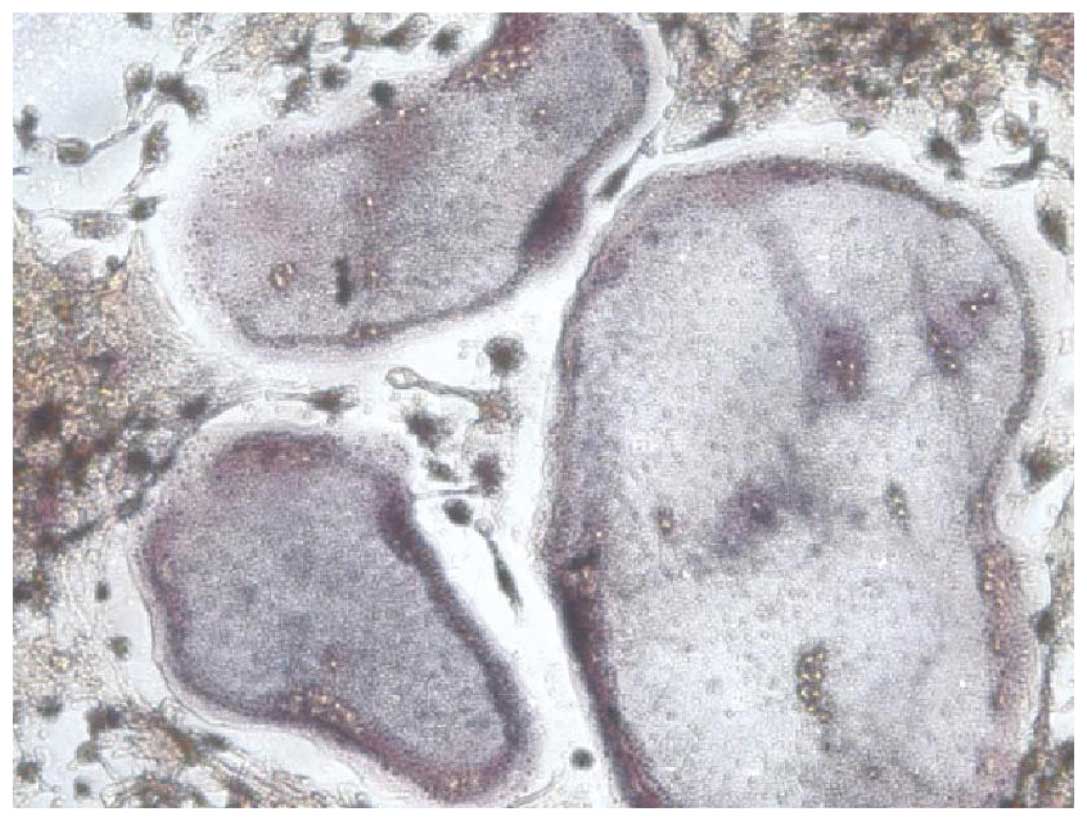
Cell observation
Cells were observed using an inverted phase contrast
microscope. The TRAP-positive cells with three nuclei were counted
as osteoclasts (Fig. 1). The
osteoclasts varied in shape when observed under the scanning
electron microscope. At the edge of cells, there were a large
number of pseudopodia-like protrusions. It formed a typical bone
resorption on the bone slices. The bone resorption was round, oval
or irregular. The bottom of the bone lacunae was rough and had
fibrous substrate (Fig. 2).
Regulation of NFATc1, c-Src and RANK mRNA
expression
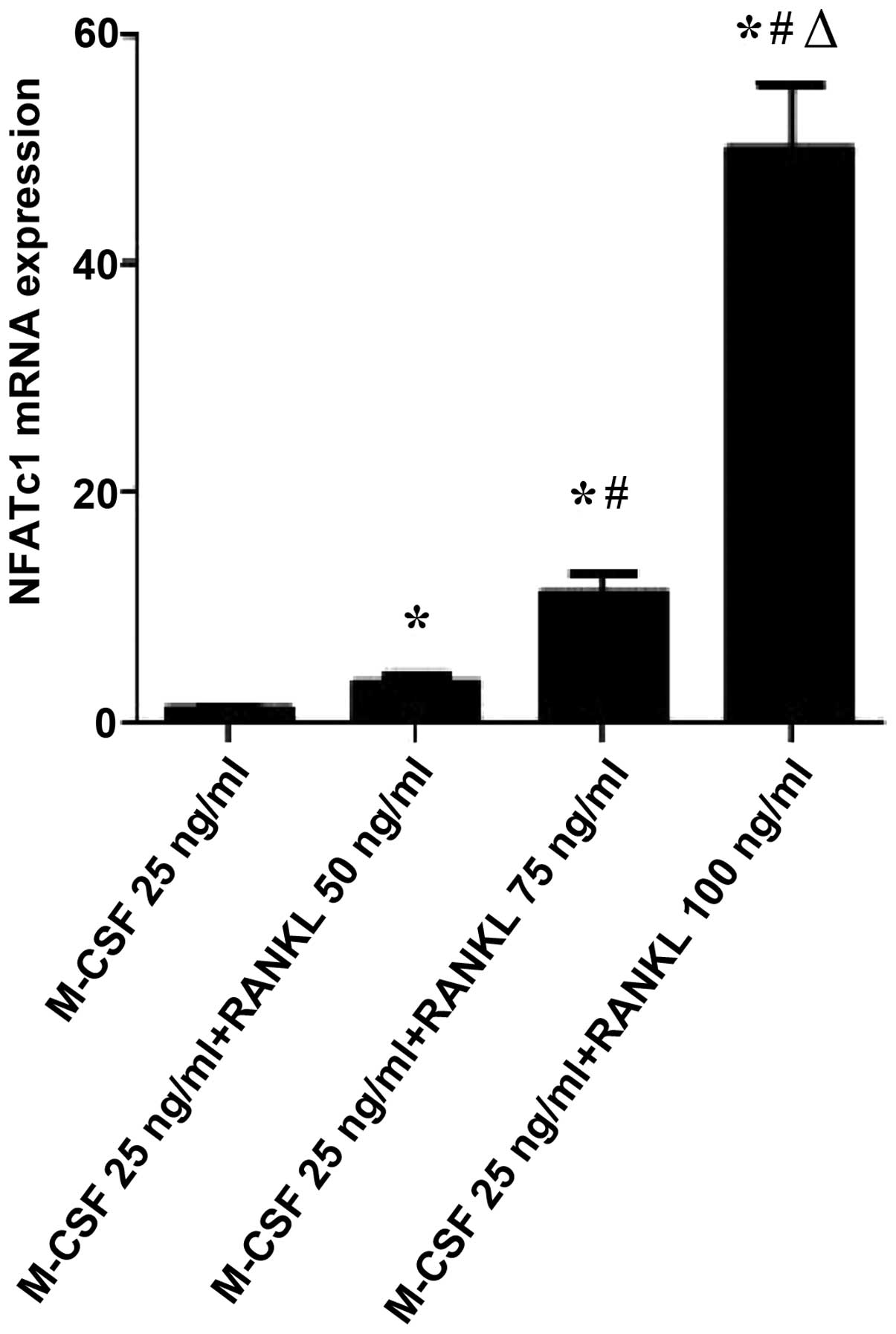
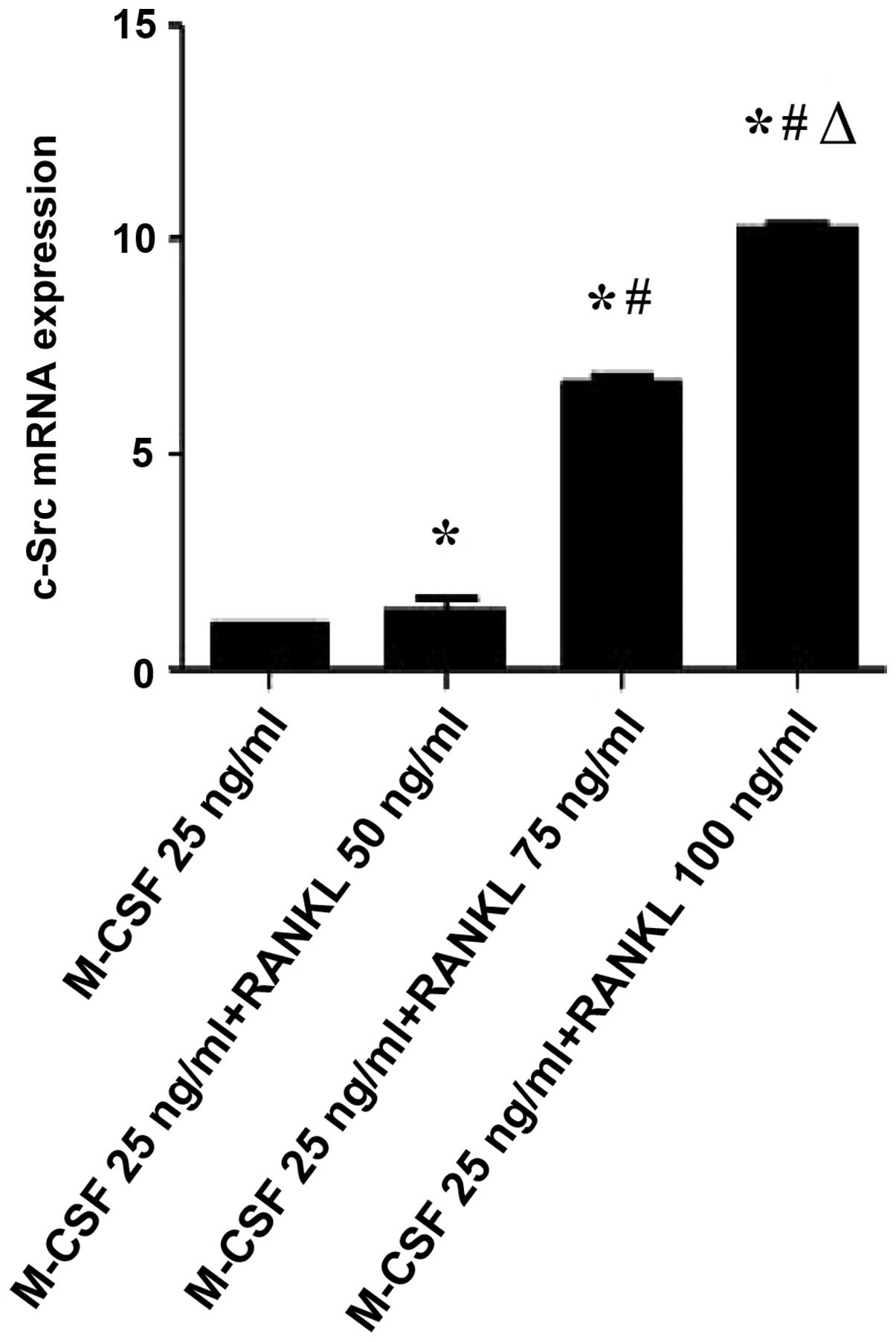
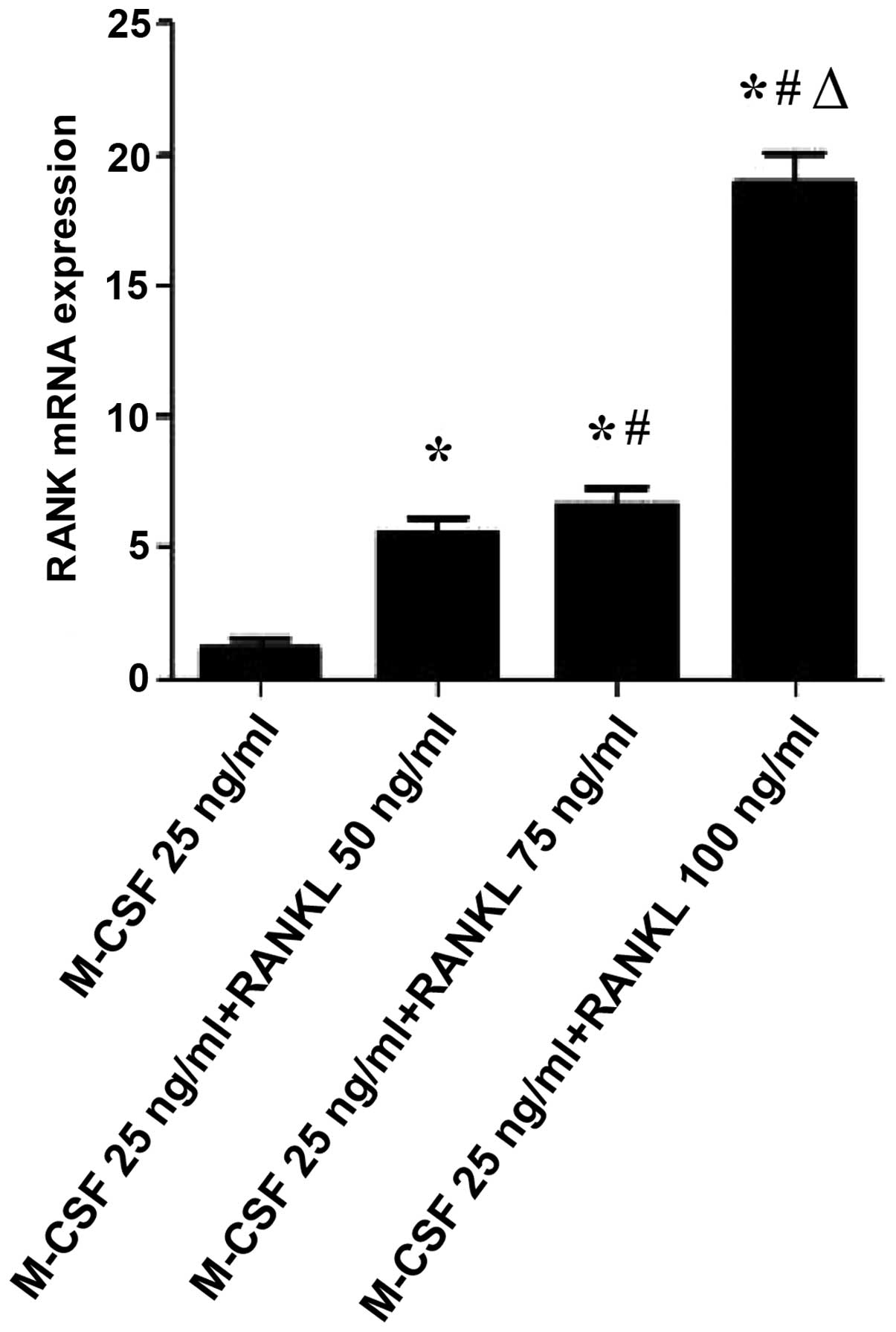
Given that RANKL induces osteoclast activation, it
was examined whether RANKL could regulate the gene expression of
NFATc1, c-Src and RANK, which is known to be associated with
osteoclast activation and function. As shown in Figs. 3Figure 4–5, RANKL induced the expression of various
osteoclast-activating genes, different concentrations of RANKL
induced the expression of NFATc1, c-Src and RANK mRNA. NFATc1,
c-Src and RANK mRNA expression in osteoclast-like cells following
treatment with different concentrations of RANKL. Compared with the
M-CSF (25 ng/ml)+RANKL (0 ng/ml) control group, the levels of
NFATc1 and c-Src mRNA expression were significantly increased in
the M-CSF (25 ng/ml)+RANKL (75 and 100 ng/ml, respectively) groups
(P<0.01). In addition, compared with the M-CSF (25 ng/ml)+RANKL
(0 ng/ml) group, the levels of RANK mRNA expression were
significantly increased in the M-CSF (25 ng/ml)+RANKL (50, 75 and
100 ng/ml, respectively) groups (P<0.01). Compared with the
M-CSF (25 ng/ml)+RANKL (50 ng/ml) group, the levels of NFATc1 mRNA
expression were significantly increased in the M-CSF (25
ng/ml)+RANKL (75 and 100 ng/ml) groups (P=0.013 and P<0.01,
respectively). Compared with the M-CSF (25 ng/ml)+RANKL (50 ng/ml)
group, the levels of c-Src mRNA expression were significantly
increased in M-CSF (25 ng/ml)+RANKL (75 and 100 ng/ml) groups
(P<0.01). Compared with the M-CSF (25 ng/ml)+RANKL (75 ng/ml)
group, the levels of NFATc1, c-Src and RANK mRNA expression were
significantly increased in tge M-CSF (25 ng/ml)+RANKL (100 ng/ml)
group (P<0.01).
Discussion
Osteoclast precursors that express RANK, a TNF
receptor family member, recognize RANKL and differentiate into
osteoclasts in the presence of M-CSF (3). M-CSF induces the proliferation of
pre-osteoclasts cells, sustains their survival and stimulates the
expression of RANK, the receptor of RANKL (20). RANKL expression by bone stromal
cells, such as osteoblasts, synovial fibroblasts and T cells is
increased adjacent to sites of pathological bone loss (17,21–23).
Upon binding to its receptor RANK, RANKL activates multiple
intracellular signaling pathways, including the MAPK and calcium
dependent pathways, and ultimately stimulates transcription factors
such as NF-κB, microphthalmia-associated transcription factor and
NFATc1 (9,24). NFATc1 is a critical transcription
factor in osteoclastogenesis.
The targeted disruption of c-Src impairs osteoclast
bone resorbing activity, causing osteopetrosis. Previous studies
have demonstrated that osteoclast activation is mediated by various
genes, including c-Src (14).
c-Src protein, a member of the non-receptor tyrosine kinase family,
was found to be highly expressed in osteoclasts (25). In addition, the expression of c-Src
appeared to be under the control of RANKL at the transcriptional
level in RAW264 mouse macrophage cells (26). Osteoclasts form normally in
Src− mice but are unable to resorb bone due to lack of a
ruffled border (14,27). Namely, osteoclasts isolated from
Src−/− mice presented with an abnormal cytoskeletal
structure, retarded cell migration, and consequently impaired bone
resorbing activity (15,28). While c-Src organizes the
cytoskeleton of the cell in response to cytokines, it does not
participate in RANKL-induced osteoclast formation.
NFATc1 and c-Src are master regulators of
RANKL-induced osteoclast differentiation and the present study
investigated the regulatory mechanism of NFATc1 and c-Src in
osteoclast activation. The principal aim of this study was to
assess whether an increase in the concentration of RANKL was
associated with changes in NFATc1, c-Src and RANK mRNA gene
expression in osteoclast-like cells. It was demonstrated that the
greater concentration of RANKL, the higher the expression of
NFATc1, c-Src and RANK in rat osteoclast-like cells. The expression
of NFATc1 appeared to be induced by RANKL stimulation. This further
demonstrated that RANKL regulates the expression of NFATc1 and
c-Src. These data suggest that NFATc1 is a key regulator of
osteoclasts through induction of c-Src.
In conclusion, these data demonstrated that RANKL
applied for 9 days could regulate the osteoclastic gene expression
of NFATc1, c-Src and RANK. These results indicate that RANKL may
modulate osteoclast activation and subsequent bone resorption
through RANK and NFATc1. It also demonstrated that c-Src
participates in the NFATc1 signaling pathway. This study suggests
that decreasing the expression of NFATc1, c-Src and RANK mRNA or
reducing the synthesis of RANKL may be a therapeutic strategy for
the treatment of In order to treat various metabolic bone
diseases.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81272168), the
Technological Project of Health Bureau, Xiamen, Fujian Province,
China (grant no. 3502Z20104031) and the Innovation Subject of
Medicine in Fujian Province (grant no. 2012-CXB-32).
References
|
1
|
Alliston T and Derynck R: Medicine:
Interfering with bone remodelling. Nature. 416:686–687. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karsenty G and Wagner EF: Reaching a
genetic and molecular understanding of skeletal development. Dev
Cell. 2:389–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Theill LE, Boyle WJ and Penninger JM:
RANK-L and RANK: T cells, bone loss, and mammalian evolution. Annu
Rev Immunol. 20:795–823. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Walsh MC, Kim N, Kadono Y, Rho J, Lee SY,
Lorenzo J and Choi Y: Osteoimmunology: Interplay between the immune
system and bone metabolism. Annu Rev Immunol. 24:33–63. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Binder NB, Niederreiter B, Hoffmann O,
Stange R, Pap T, Stulnig TM, Mack M, Erben RG, Smolen JS and
Redlich K: Estrogen-dependent and C-C chemokine
receptor-2-dependent pathways determine osteoclast behavior in
osteoporosis. Nat Med. 15:417–424. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fata JE, Kong YY, Li J, Sasaki T,
Irie-Sasaki J, Moorehead RA, Elliott R, Scully S, Voura EB, Lacey
DL, et al: The osteoclast differentiation factor
osteoprotegerin-ligand is essential for mammary gland development.
Cell. 103:41–50. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fuller K, Wong B, Fox S, Choi Y and
Chambers TJ: TRANCE is necessary and sufficient for
osteoblast-mediated activation of bone resorption in osteoclasts. J
Exp Med. 188:997–1001. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Teitelbaum SL and Ross FP: Genetic
regulation of osteoclast development and function. Nat Rev Genet.
4:638–649. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takayanagi H, Kim S, Koga T, Nishina H,
Isshiki M, Yoshida H, Saiura A, Isobe M, Yokochi T, Inoue J, et al:
Induction and activation of the transcription factor NFATc1 (NFAT2)
integrate RANKL signaling in terminal differentiation of
osteoclasts. Dev Cell. 3:889–901. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou P, Sun LJ, Dötsch V, Wagner G and
Verdine GL: Solution structure of the core NFATC1/DNA complex.
Cell. 92:687–696. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Na KL: Molecular understanding of
osteoclast differentiation and physiology. J Clin Endocr Metab.
25:264–269. 2010.
|
|
13
|
Ishida N, Hayashi K, Hattori A, Yogo K,
Kimura T and Takeya T: CCR1 acts downstream of NFAT2 in
osteoclastogenesis and enhances cell migration. J Bone Miner Res.
21:48–57. 2006. View Article : Google Scholar
|
|
14
|
Soriano P, Montgomery C, Geske R and
Bradley A: Targeted disruption of the c-src proto-oncogene leads to
osteopetrosis in mice. Cell. 64:693–702. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lakkakorpi PT, Nakamura I, Young M,
Lipfert L, Rodan GA and Duong LT: Abnormal localisation and
hyperclustering of alpha (v)beta (3) integrins and associated
proteins in Src-deficient or tyrphostin A9-treated osteoclasts. J
Cell Sci. 114:149–160. 2001.
|
|
16
|
Chen J, He JQ, Zheng SY and Huang LQ: OPG
inhibits gene expression of RANK and CAII in mouse osteoclast-like
cell. Rheumatol Int. 32:3393–3398. 2012. View Article : Google Scholar
|
|
17
|
Chen J and He CQ, Xia QJ, Huang LQ, Hu YJ
and He CQ: Effects of pulsed electromagnetic fields on the mRNA
expression of RANK and CAII in ovariectomized rat osteoclast-like
cell. Connect Tissue Res. 51:1–7. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pavlos NJ, Xu J, Riedel D, Yeoh JS,
Teitelbaum SL, Papadimitriou JM, Jahn R, Ross FP and Zheng MH:
Rab3D regulates a novel vesicular trafficking pathway that is
required for osteoclastic bone resorption. Mol Cell Biol.
25:5253–5269. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Arai F, Miyamoto T, Ohneda O, Inada T,
Sudo T, Brasel K, Miyata T, Anderson DM and Suda T: Commitment and
differentiation of osteoclast precursor cells by the sequential
expression of c-Fms and receptor activator of nuclear factor kappaB
(RANK) receptors. J Exp Med. 190:1741–1754. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Crotti T, Smith MD, Hirsch R, Soukoulis S,
Weedon H, Capone M, Ahern MJ and Haynes D: Receptor activator NF
kappaB ligand (RANKL) and osteoprotegerin (OPG) protein expression
in periodontitis. J Periodontal Res. 38:380–387. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Crotti TN, Smith MD, Findlay DM, Zreiqat
H, Ahern MJ, Weedon H, Hatzinikolous G, Capone M, Holding C and
Haynes DR: Factors regulating osteoclast formation in human tissues
adjacent to peri-implant bone loss: Expression of receptor
activator NFkappaB, RANK ligand and osteoprotegerin. Biomaterials.
25:565–573. 2004. View Article : Google Scholar
|
|
23
|
Crotti TN, Smith MD, Weedon H, Ahern MJ,
Findlay DM, Kraan M, Tak PP and Haynes DR: Receptor activator
NF-kappa B ligand (RANKL) expression in synovial tissue from
patients with rheumatoid arthritis, spondyloarthropathy,
osteoarthritis, and from normal patients: Semiquantitative and
quantitative analysis. Ann Rheum Dis. 61:1047–1054. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hirotani H, Tuohy NA, Woo JT, Stern PH and
Clipstone NA: The calcineurin/nuclear factor of activated T cells
signaling pathway regulates osteoclastogenesis in RAW264.7 cells. J
Biol Chem. 279:13984–13992. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thomas SM and Brugge JS: Cellular
functions regulated by Src family kinases. Annu Rev Cell Dev Biol.
13:513–609. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kumagai N, Ohno K, Tameshige R, Hoshijima
M, Yogo K, Ishida N and Takeya T: Induction of mouse c-src in
RAW264 cells is dependent on AP-1 and NF-kappaB and important for
progression to multinucleated cell formation. Biochem Biophys Res
Commun. 325:758–768. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lowe C, Yoneda T, Boyce BF, Chen H, Mundy
GR and Soriano P: Osteopetrosis in Src-deficient mice is due to an
autonomous defect of osteoclasts. Proc Natl Acad Sci USA.
90:4485–4489. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Boyce BF, Yoneda T, Lowe C, Soriano P and
Mundy GR: Requirement of pp60c-src expression for osteoclasts to
form ruffled borders and resorb bone in mice. J Clin Invest.
90:1622–1627. 1992. View Article : Google Scholar : PubMed/NCBI
|