Introduction
Colorectal cancer is a leading cause of morbidity
and mortality in patients worldwide. A significant number of men
and women are at risk of developing invasive colon cancer in their
lifetime (1). Inflammatory bowel
disease is a group of inflammatory conditions in the human large
and small intestines. It is recognized that chronic inflammation is
a risk factor for tumor development, including colorectal cancer
(2). Thus, targeting inflammatory
pathways has been shown to be effective in preventing the formation
of colon tumors and their malignant progression in animal and human
studies (3–7).
There is growing evidence to suggest that certain
natural products possess anti-inflammatory and anticancer
potential. Mahonia bealei is an evergreen shrub of the
Berberidaceae family, native to East Asia, and North and Central
America. In Chinese folk medicine, the root, stem and leaf of M.
bealei have been used for thousands of years as herbal
medicines to treat various ailments, in particular gastrointestinal
inflammation-related illnesses, such as diarrhea and dysentery
(8). Modern phytochemistry studies
have identified a number of alkaloids from M. bealei, such
as berberine, palmatine and jatrorrhizine, the latter two being
protoberberine alkaloids. These compounds have been shown to
exhibit antioxidant, anti-inflammatory, and anticancer effects
(9).
Compared with the extensive research on berberine
(9), the investigation of
palmatine is limited. Notably, several recently published studies
have demonstrated that palmatine not only has anticancer potential,
but is also an anti-inflammatory agent (10,11).
Thus the aim of this study explore the effect of palmitine on colon
cancer.
Our previous study observed the antitumor effects of
different botanicals. For example, using a colon tumor-xenograft
nude mouse model, significant antitumor activities of American
ginseng and Oplopanax horridus were identified (12,13).
However, the nude mouse is not a gut disease-specific animal model.
Thus, in this study, to ascertain the anticancer effects of
palmatine, in vitro evaluation was used to demonstrate that
palmatine significantly inhibited colorectal cancer cell
proliferation and the expression of inflammatory cytokine
interleukin (IL)-8. Since it is desirable to use specific gut
inflammatory and tumorigenesis animal models for in vivo
studies ApcMin/+
mice, a model with a mutant Apc gene, were then used for
investigation. This Min (multiple intestinal neoplasia)
mouse is characterized by early lethality, colon tumors, and
development of a number of polyps in the small intestine (14,15).
To increase the possibility of gut diseases, the mice were fed
high-fat rodent chow to mimic a Western diet (16), as previous research has shown that
high-fat diet intake is able to promote colon cancer by increasing
fatty acids and secondary bile acids in the colonic lumen (17). Data from the present study showed
that palmatine significantly reduced the progression of high-fat
diet-enhanced gut inflammation and tumorigenesis in
ApcMin/+ mice,
supported by the inflammatory cytokine level reductions. In
addition, mice displayed increased survival rates, which was
consistent with the pharmacological observations. These results
suggest that palmatine has a potential clinical utility as a colon
cancer therapeutic.
Materials and methods
Chemicals
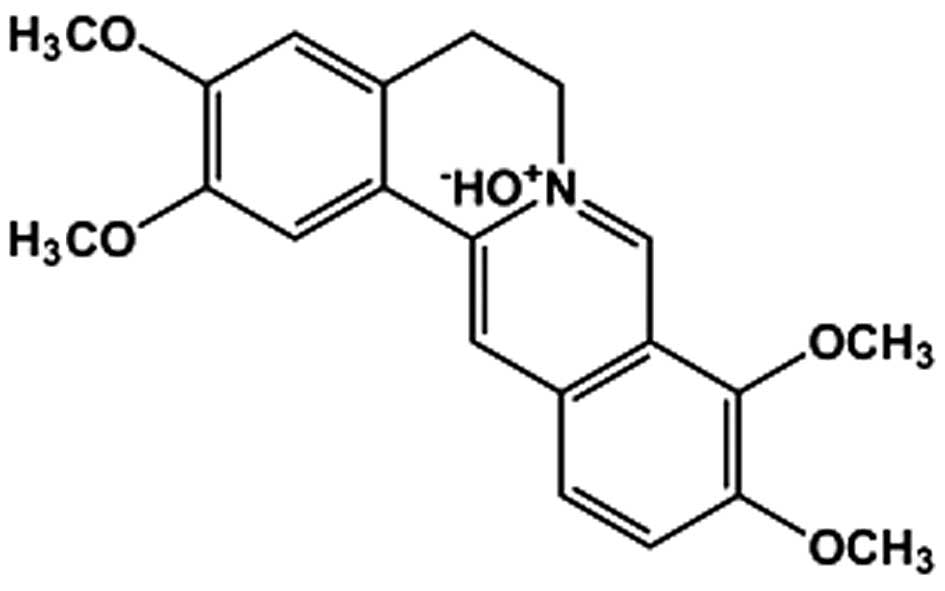
Palmatine (98% purity; Fig. 1) was isolated from Mahonia
bealei as previously described (18). McCoy's 5A and Dulbecco's modified
Eagle's medium (DMEM), fetal bovine serum (FBS), penicillin and
streptomycin were obtained from Mediatech, Inc. (Herndon, VA, USA).
Dimethylsulfoxide and lipopolysaccharide were obtained from
Sigma-Aldrich (St. Louis, MO, USA).
Cell culture and proliferation assay
HT-29 and SW-480 human colorectal cancer cell lines
were obtained from the American Type Tissue Collection (Rockville,
MD, USA), with HT-29 maintained in McCoy's 5A and SW-480 in DMEM.
All medium were supplemented with 10% FBS, penicillin (100 IU/ml)
and streptomycin (100 µg/ml). The colon cancer cells were
subcultured twice a week and incubated in a humidified atmosphere
with 5% CO2 at 37°C.
Young adult mouse colon (YAMC) cells were obtained
from the Digestive Disease Research Core Center at the University
of Chicago (Chicago, IL, USA), and were grown in RPMI-1640
(Mediatech, Inc., Herndon, VA, USA) medium supplemented with 5%
neonatal calf serum (Hyclone; GE Healthcare Life Sciences, Logan,
UT, USA) ITS+ (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA; 6.25 mg/ml insulin, 6.25 mg/ml transferrin,
6.25 ng/ml selenous acid, 5.35 mg/ml linoleic acid, and 1.25 mg/ml
bovine serum albumin), 5 IU/ml murine interferon-γ (PeproTech,
Inc., Rocky Hill, NJ, USA), penicillin and streptomycin. The YAMC
cells were cultured in a humidified atmosphere with 5%
CO2 at 33°C.
Palmatine was dissolved in DMSO and stored in small
aliquots at −20°C prior to use. The cancer cells were seeded in
96-well plates at a density of 5,000 cells/well, allowed to attach
overnight and then treated with different concentrations of
palmatine. Cell proliferation was measured at 48 h using the Cell
Titer 96 Aqueous MTS Reagent (Promega Corporation, Madison, WI,
USA) according to the manufacturer's instructions. The absorbance
was read by an automated microplate reader (Epoch; Bio-Tek
Instruments, Winooski, VT, USA) set to a wavelength of 490 nm. Data
are expressed as the percentage of treated cells vs. control
(vehicle set at 100%) (19).
IL-8 secretion analysis
The HT-29 and SW-480 cells were seeded in 24-well
plates and cultured for 72 h. Cell monolayers were washed with
phosphate-buffered saline and fresh medium was added in the
presence of 100 ng/ml lipopolysaccharide (LPS) or different
concentrations of palmatine plus 100 ng/ml LPS. After incubation
for 6 h, the culture medium was collected, the secreted IL-8 was
quantified by enzyme-linked immunosorbent assay (ELISA; Thermo
Fisher Scientific, Inc.), and anti-inflammatory activities were
calculated.
Animals and experimental protocol
The experimental protocols were approved by the
Institutional Animal Care and Use Committee of the University of
Chicago. Male C57BL/6J-ApcMin/J (n=5) and female
C57BL/6J (n=5) mice were purchased from Jackson Laboratory (Bar
Harbor, ME, USA) for breeding. Mice were caged under controlled
room temperature, humidity and light (12-h light:dark cycle) and
allowed free access to rodent chow and tap water. After weaning,
genotyping was conducted via tail biopsy using polymerase chain
reaction (PCR)-based assays to identify
ApcMin/+ mice
(14).
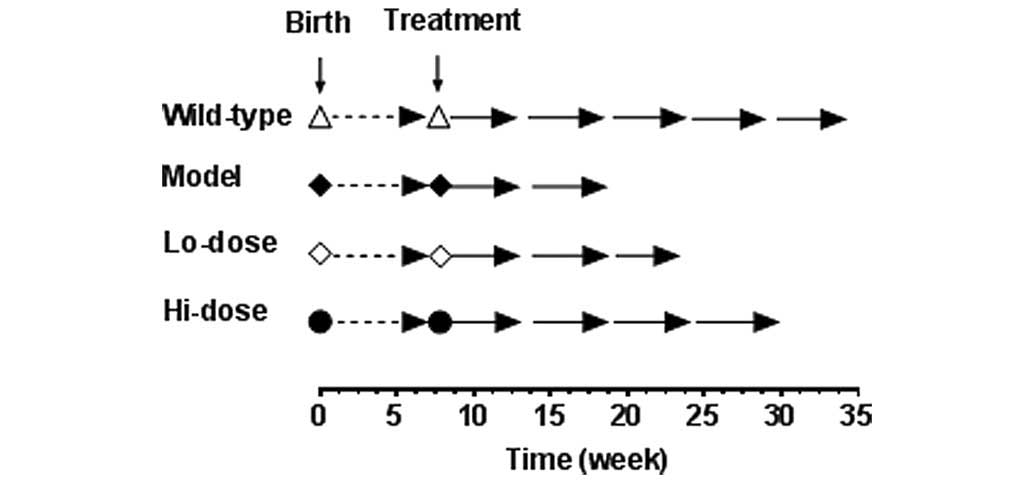
The study protocol is shown in Fig. 2. Before they were 8-weeks old, all
mice consumed standard rodent chow. Starting at week 8, the animals
received a Western high-fat diet, and the four experimental groups
(n=5 per group) are as follows: i) Wild-type mice as negative
control; ii)
ApcMin/+ mice in
untreated model group as a positive control; iii)
ApcMin/+ mice
receiving a low-dose of palmatine in animal chow, equivalent to 10
mg/kg/day; and iv)
ApcMin/+ mice
receiving a high-dose of palmatine in animal chow, equivalent to 20
mg/kg/day. The two doses of palmatine were selected based on data
obtained from a pilot study in which the effective oral dose of
palmatine started at 6–8 mg/kg/day. No significant adverse events
were observed in mice that were administered palmatine.
The high-fat Western diet (Harlan Laboratories,
Madison, WI, USA) contains 20% fat and includes beef tallow (35
g/kg), lard (30 g/kg) and corn oil (80 g/kg) (16). For the drug groups, palmatine was
evenly mixed with the high-fat chow. The daily oral palmatine dose
was calculated based on the amount of chow consumption. Animal body
weight was measured at least once a week.
Animal survival data was obtained when a mouse was
critically ill (as determined by Animal Care Facility staff, who
assisted in the regular monitoring of the animals) and expected to
die within the next 36 h. At that point, the animal was sacrificed
by cervical dislocation after collecting blood from the heart,
followed by the removal of the organs and subsequent measurements
and analyses. The small intestine and colon were harvested, flushed
immediately with ice-cold PBS and cut open longitudinally. Two
independent investigators who were blinded with respect to the
treatment groups counted tumor numbers and sizes under a dissection
microscope (SZX7 Stereo Microscope; Olympus Corporation, Tokyo,
Japan). Small intestine and colon tissue samples were fixed in 10%
neutral-buffered formalin, embedded in paraffin blocks, and
processed by routine histological staining. Gut tissue was
collected for reverse transcription-quantitative PCR analysis of
inflammatory cytokines. Plasma samples were collected to determine
the levels of cytokines using the ELISA kits.
In addition, at the end of the experiment, the
spleen, thymus and liver were removed and weighed. Organ index was
calculated based on the following equation: Organ index (%) = organ
weight (g) / body weight (g) x 100 (20,21).
Histological assessment
Colon and small intestine samples (2 mm2)
were harvested from the
APCmin/+ mice. Two
wedge-shaped incisions were made with a scalpel (blade no. 15c;
Swann Morton, Ltd., Sheffield, UK). Subsequent to harvesting, the
material was placed in a sterile test tube filled with a 5%
formalin solution.
Following fixation, the material was put through a
Thermo Scientific Citadel Tissue Processor (Thermo Fisher
Scientific, Inc.) using a standard processor program (rinsing in
50, 90 and 98% alcohol series in turns for 1 h each, rinsing three
times with xylene and embedding in paraffin). Once embedded in
paraffin blocks and cut with a semi-automatic Shandon Finesse 325
microtome (Thermo Fisher Scientific, Inc.) to a thickness of 7
µm, the material was stained with hematoxylin and eosin
(H&E; Leica Microsystems, Inc., Buffalo Grove, IL, USA)
solution using a Thermo Scientific Gemini AS Automated Slide
Stainer (Thermo Fisher Scientific, Inc.). Histological examination
was performed by descriptive analysis under light microscopy with
an Olympus BX41 microscope (Olympus Corporation) and the stained
sections were examined for histopathological changes by a
gastrointestinal pathologist.
RNA extraction and RT-qPCR
Total RNA was isolated from mouse small intestine
and colon tissues using the miRNeasy kit (Qiagen, Valencia, CA,
USA) according to the manufacturer's instructions, and was used as
a template to synthesize cDNA for RT-qPCR. First strand cDNA was
synthesized the using Thermo Scientific Maxima First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.). RT-qPCR was
performed on a 7900HT real-time PCR system (Applied Biosystems,
Foster City, CA, USA) with the following cycling conditions: 1 PCR
cycle, 95°C, 10 min; 40 PCR cycles, 60°C, 1 min, 95°C, 15 sec.
RT-qPCR with SYBR Green dye (Qiagen) was used to determine the
expression of genes. Signals were analyzed by the ABI Prism
Sequence Detection System (version 2.2; Thermo Fisher Scientific,
Inc.). The 2−ΔΔCq method for relative quantification was
used (22). Primers for RT-qPCR
were purchased from Sigma-Aldrich and are listed in Table I. β-actin was used as an endogenous
control. Each sample was run in triplicate.
 | Table IPrimers used for RT-qPCR analysis of
inflammatory cytokines. |
Table I
Primers used for RT-qPCR analysis of
inflammatory cytokines.
| Gene | Primer
sequence |
|---|
| IL1α | F:
5′-CGAAGACTACAGTTCTGCCATT-3′ |
| R:
5′-GACGTTTCAGAGGTTCTCAGAG-3′ |
| IL1β | F:
5′-GCAACTGTTCCTGAACTCAACT-3′ |
| R:
5′-ATCTTTTGGGGTCCGTCAACT-3′ |
| IL8 | F:
5′-ACTGAGAGTGATTGAGAGTGGAC-3′ |
| R:
5′-AACCCTCTGCACCCAGTTTTC-3′ |
| G-CSF | F:
5′-ATGGCTCAACTTTCTGCCCAG-3′ |
| R:
5′-CTGACAGTGACCAGGGGAAC-3′ |
| GM-CSF | F:
5′-GGCCTTGGAAGCATGTAGAGG-3′ |
| R:
5′-GGAGAACTCGTTAGAGACGACTT-3′ |
| β-actin | F:
5′-GGCTGTATTCCCCTCCATCG-3′ |
| R:
5′-CCAGTTGGTAACAATGCCATGT-3′ |
Statistical analysis
Data are presented as the mean ± standard
derivation. Polyp data was analyzed using a one-way analysis of
variance (ANOVA) with Newman-Keuls post-hoc analysis. All
inflammatory mediator data were analyzed using a two-way ANOVA with
Student Newman-Keuls post hoc analysis. GraphPad Prism software
(version 6.0; GraphPad Software, Inc., La Jolla, CA, USA) was used
for the analyses, including the Kaplan-Meier survival curve.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects on colorectal cancer cell
proliferation and secretion of inflammatory cytokine IL-8
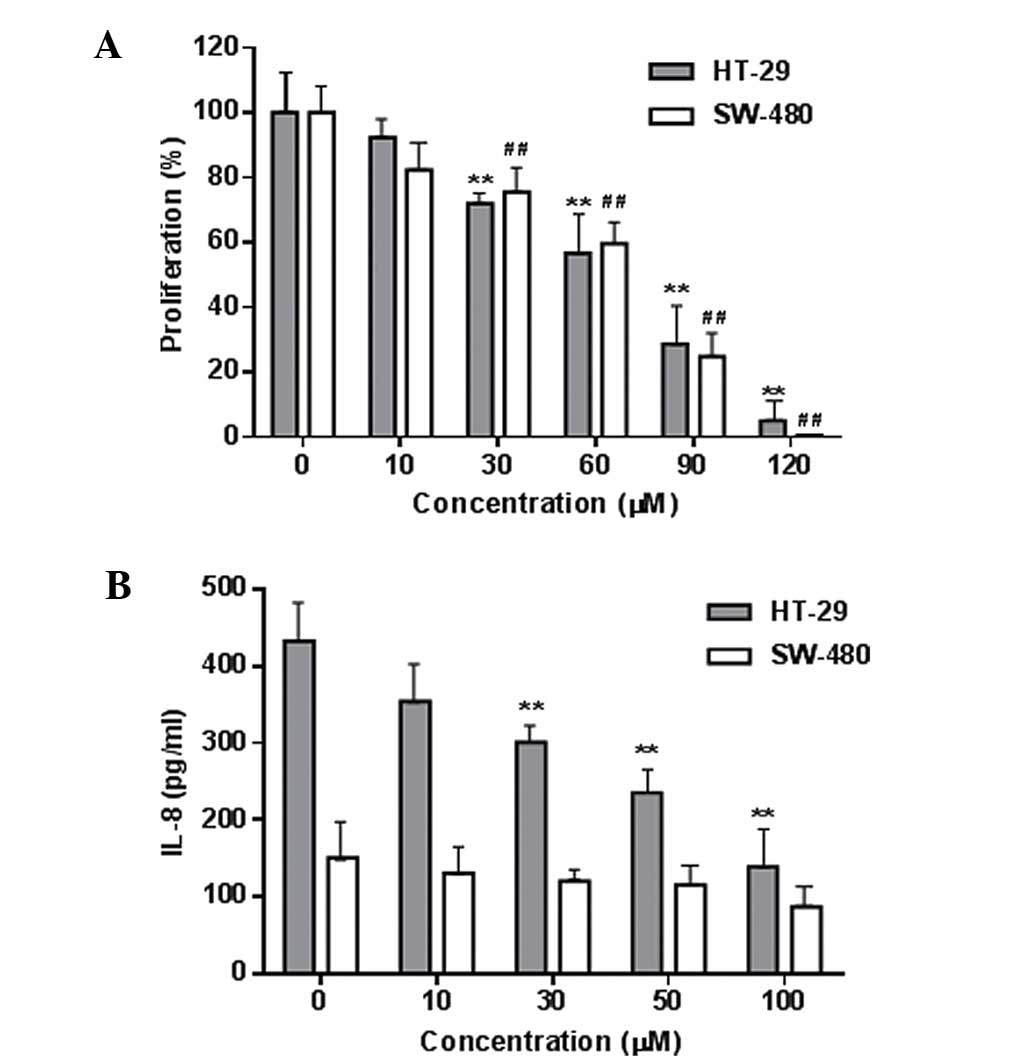
Fig. 3A shows the
concentration-related inhibitory effects of palmatine on HT-29 and
SW-480 colorectal cancer cell growth (P<0.01 compared with the
control). The IC50 for HT-29 and SW-480 cells was 68.3
and 72.6 µM, respectively. Using the YAMC cells, it was also
observed that palmatine did not significantly affect the growth of
these non-cancer cells even at high concentrations (100–120
µM). These results suggested that palmatine possesses in
vitro antiproliferative potential on human colorectal cancer
cells.
The LPS-induced production of the inflammatory
cytokine IL-8 in these two cancer cells was determined, and the
IL-8 level was higher in the HT-29 cells than that in the SW-480
cells (Fig. 3B). Palmatine
significantly reduced the IL-8 production in HT-29 cells (P<0.01
compared with the control). This result was consistent with a
previous study that demonstrated that the HT-29 cell line is a
suitable in vitro model for the evaluation of
anti-inflammation and anti-inflammatory responses (23).
Effects on body weight and organ index
using ApcMin/+ mice
To further evaluate the effects of palmatine on
colorectal cancer and inflammatory responses, the in vivo
ApcMin/+ mouse model
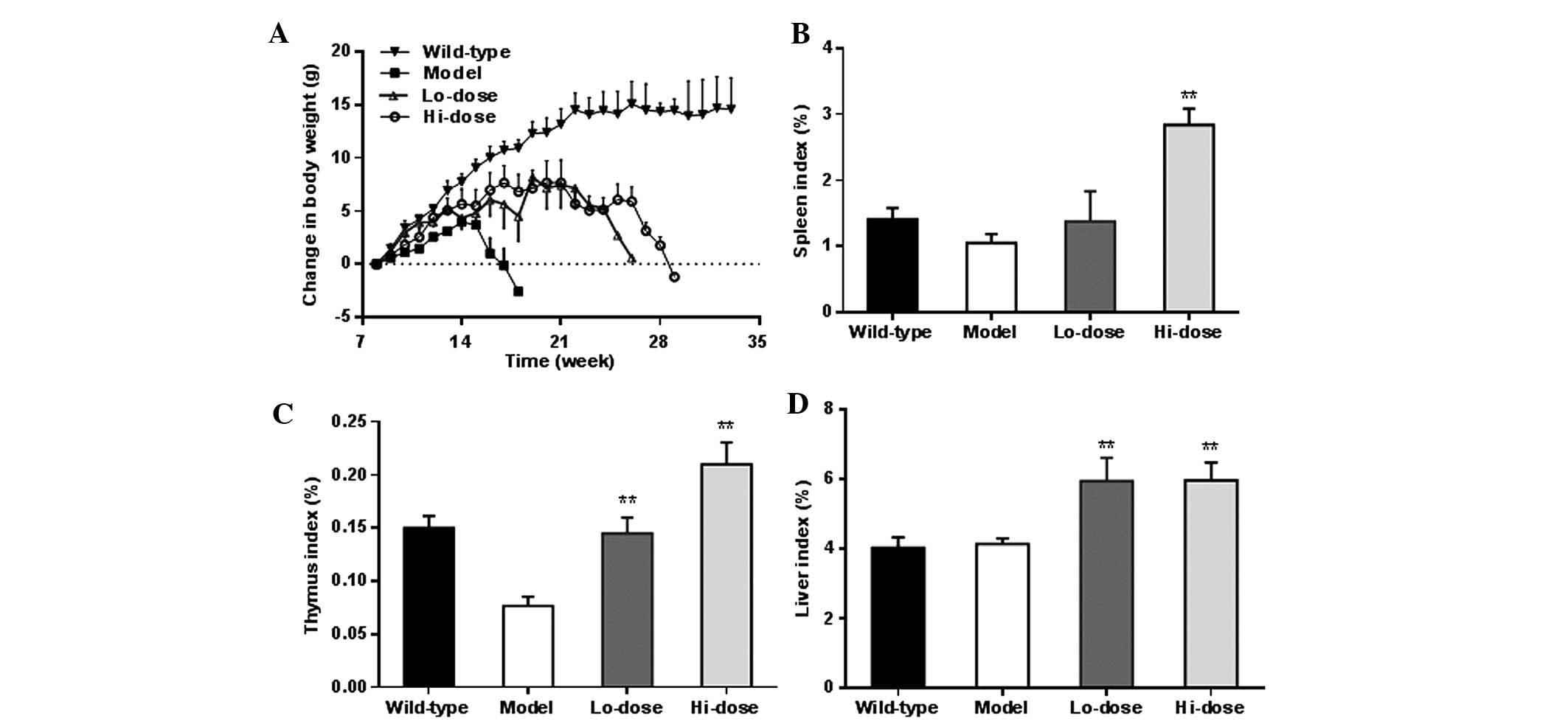
was employed. The weight changes of animals in the different
experimental groups (Fig. 3) after
week 8 are shown in Fig. 4A. As
expected, wild-type mice fed with a Western high-fat diet had
notable weight gain, approximately 13 g increase over the 14-week
observation period. However, the
ApcMin/+ mice in the
model group did not respond to the high-fat diet well. The body
weight increase was slower compared with that of the wild-type mice
up to week 7. From week 8, the mice in the model group lost weight
gradually until sacrifice. The
ApcMin/+ mice
treated with palmatine showed noticeably less body weight reduction
compared with the model group, and their weight only gradually
decreased from week 14.
Fig. 4B–D shows the
changes in the organ weight index of the spleen, thymus and liver,
respectively. In a dose-dependent manner, palmatine increased the
spleen index and thymus index compared with the model group
(P<0.01). The liver index was also significantly increased
following palmatine treatment (P<0.01).
Effects on ApcMin/+ mice life
spans
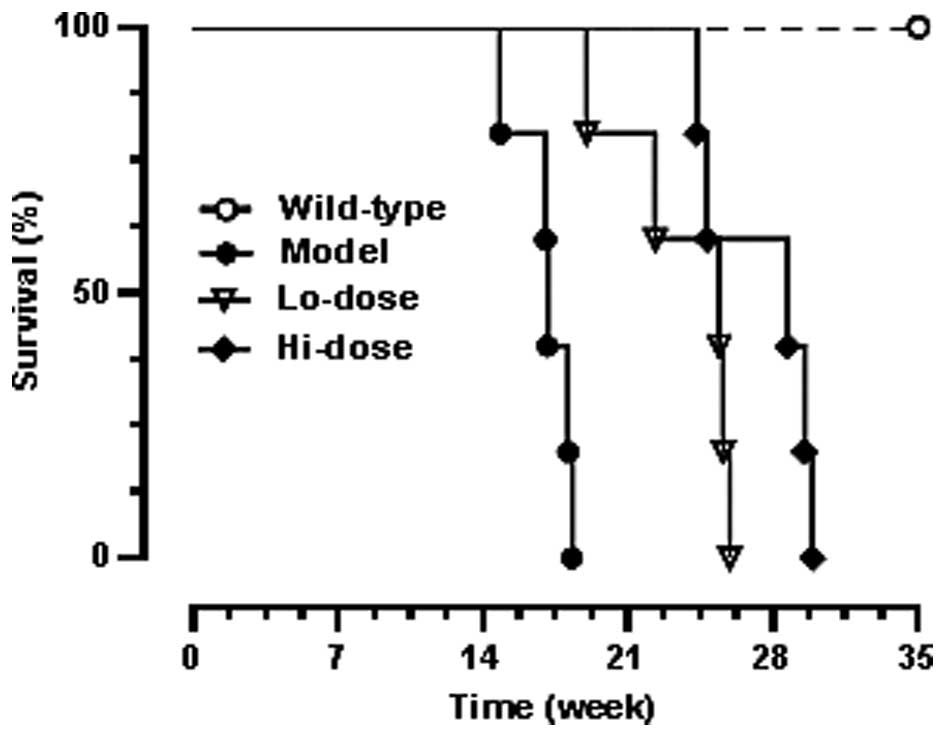
Fig. 5 shows the
Kaplan-Meier survival curve which demonstrated the dose-related
effects of palmatine on life span. Before week 19, all
ApcMin/+ mice in the
model group had died. The curve shows that both low-dose and
high-dose palmatine significantly increased mouse survival and
dose-related effects were observed (P<0.01 compared with the
model group).
Effects on gut histopathological
changes
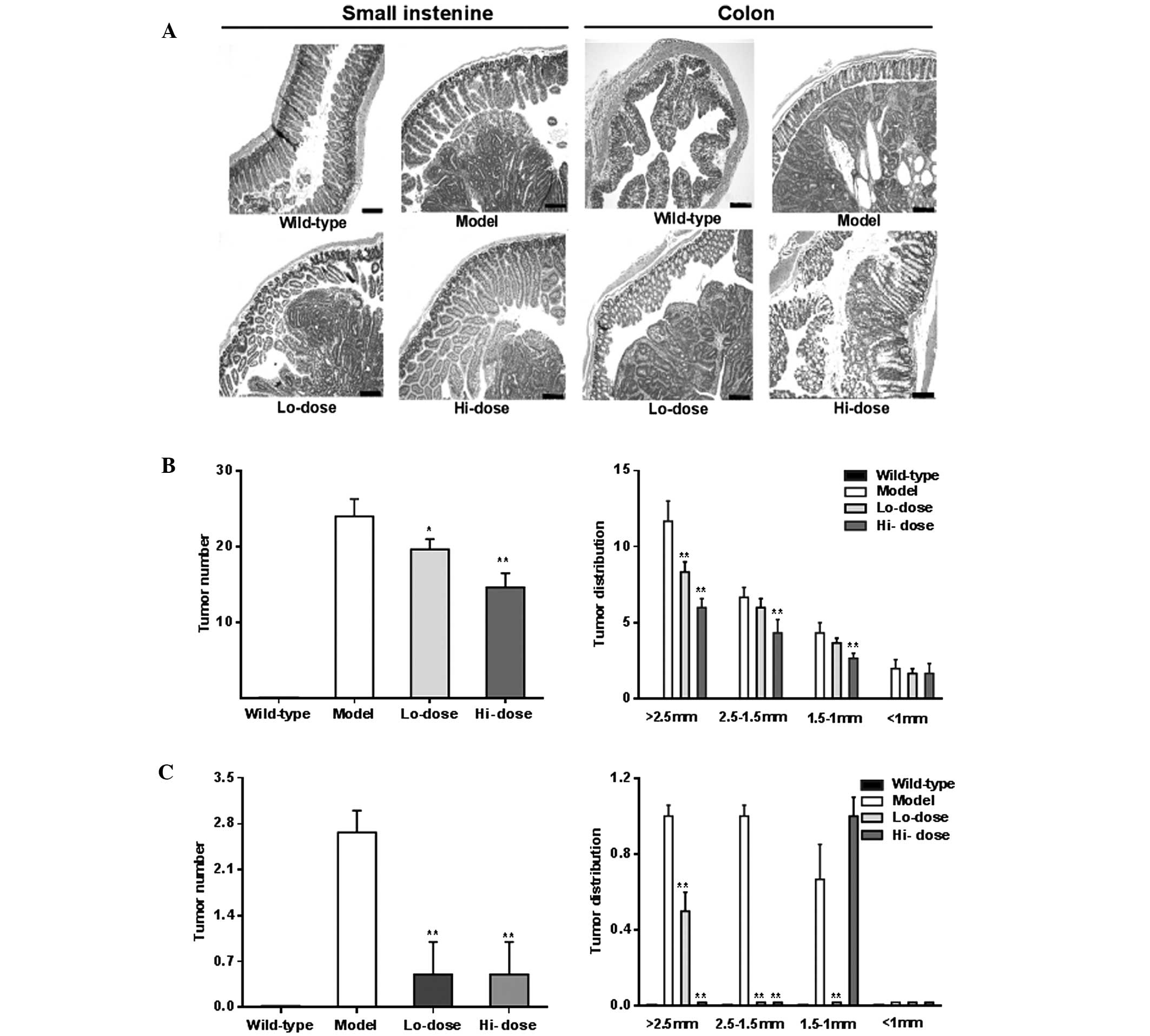
Fig. 6A shows
representative H&E staining histological sections of
experimental animals with different groups. The histology of the
model group showed prominent adenomatous changes along with
inflammatory lesions, such as neutrophil infiltration, and focal
adenomatous changes were also observed in the
ApcMin/+ mice. In
the palmatine treatment groups, however, the dysplastic changes
were reduced in the small intestine and colon sections.
Effects on tumor multiplicity
changes
Compared with the model group, following palmatine
treatment, tumor numbers were significantly reduced in the small
intestine and colon as shown in Fig.
6B and C (P<0.01). Tumor distribution data of the gut,
particularly from the colon, showed a significant decrease in the
number of large tumors and an increase in the number of small
tumors, supporting the effects of palmatine in the attenuation of
the tumorigenesis.
Effects on inflammatory cytokine
expression in gut tissue and serum
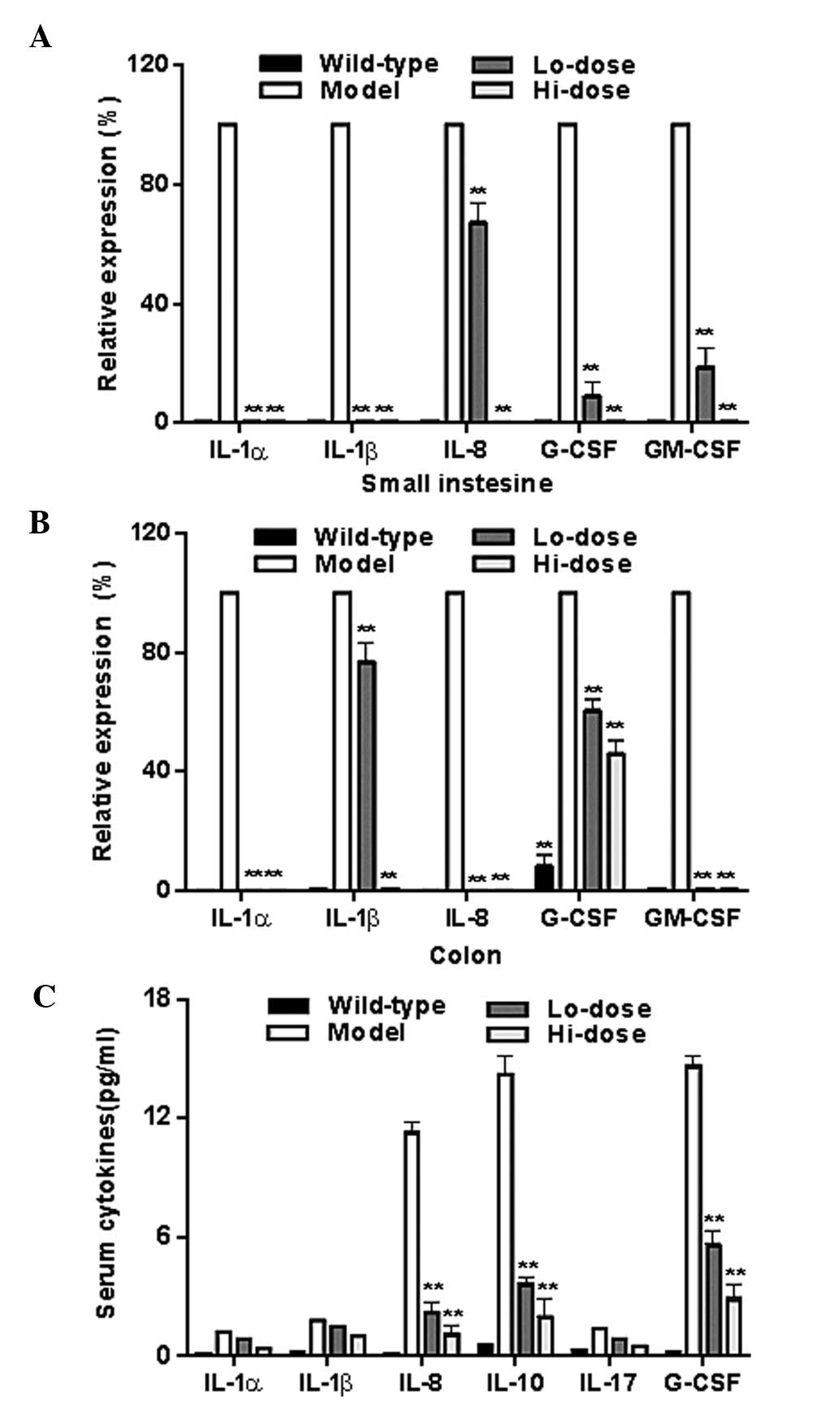
Fig. 7A and B show
the mRNA levels of data of IL-1α, IL1-β, IL-8, granulocyte-colony
stimulating factor (G-CSF), and granulocyte-colony stimulating
factor (GM-CSF) in the gut tissue from the model group and
palmatine treated groups as determined by RT-qPCR. The levels of
all of these inflammatory cytokines were reduced significantly
following treatment (all P<0.01 compared with the model group).
Fig. 7C illustrates the
significant reductions of IL-8, IL-10 and G-CSF in serum after
palmatine treatment (P<0.01 compared with the model group).
Discussion
Gut inflammation can initiate and promote stimuli
and mediators, generating a tumor-prone microenvironment. Chronic
inflammation is a risk factor for tumor development, including
colorectal cancer (3), and the
inhibition of the inflammatory pathway is proven to be effective in
preventing the initiation and progression of colon cancer (2). The use of non-steroidal
anti-inflammatory drugs (NSAIDS) can reduce colon tumorigenesis,
but concerns and long-term risks of NSAIDs render this form of
prevention unsuitable as a general recommendation in clinical
practice (24). Thus, there is a
strong motivation for exploring alternative strategies, including
the use of herbal medicines, in the management of malignancies in
the gastrointestinal system (25–27).
It was hypothesized that the natural compound
palmatine may have anti-colorectal cancer potential, and actions of
the compound were at least in part mediated by anti-inflammatory
activity. To test this hypothesis, the anti-inflammation and
anti-colorectal cancer activities were evaluated using different
experimental paradigms. The in vitro effects of palmatine on
HT-29 and SW-480 colorectal cancer cell proliferation were
observed. As expected, the compound showed concentration-dependent
inhibitory effects on cancer cell growth. In addition, the levels
of IL-8 in these two types of cancer cell were detected, and
palmatine was observed to significantly reduce the IL-8 production
of HT-29 cells. In vitro cell models can be used to screen
natural compounds with anti-inflammatory activity. In this study,
the IL-8 level was tested first using two cancer cell lines
following treatment with LPS. Since HT-29 cells were sensitive to
LPS stimulation compared with SW-480 cells, data suggested that
HT-29 is a suitable cell model to quickly screen for
anti-inflammatory effects of natural compounds.
The role of IL-8 in cancer cells and within the
tumor microenvironment has been presented, and the IL-8 activity
appeared to be associated with tumor progression through its
potential function in the regulation of angiogenesis, cancer cell
growth and migration, leukocyte infiltration and modification of
immune responses (28). The
aforementioned functions of IL-8 may be crucial for the in
vitro palmatine effects observed in the present study, which
were subsequently evaluated in in vivo studies.
Two gut-specific animal models, AOM/DSS mice and
ApcMin/+ mice, have
been employed in a number of research studies. AOM/DSS is a
chemical compound-induced gut disease model, characterized by early
inflammation followed by tumor development (29,30).
This chemically induced murine model has been used frequently to
investigate inflammation-related colon tumorigenesis in the last
decade. Ginseng is a commonly used herbal medicine worldwide and
has been highly investigated for its chemopreventive effects
(31–33). Our group previously used the
AOM/DSS mouse model in our ginseng research (30). The
ApcMin/+ mouse model
is another commonly used gut-specific carcinogenesis model. This
mouse model has a mutant in the Apc gene resulting in the
growth of small intestine polyps and colon tumors, and the
association of inflammation with tumorigenesis has been reported
(34). Using the anti-inflammatory
agent celecoxib, a cyclooxygenase-2 inhibitor belonging to the
NSAID family, tumor preventive and therapeutic effects in
ApcMin/+ mice have
been demonstrated (34). To verify
the in vitro antiproliferation and anti-inflammation effects
of palmatine, an in vivo evaluation was performed using the
ApcMin/+ mouse
model.
Like a number of other herbal medicines, M.
bealei is orally administered. In the present study, oral
administration of palmatine, an active constituent in M.
bealei, was observed to significantly attenuate the progression
of high-fat diet enhanced inflammation and tumorigenesis in
ApcMin/+ mice. Body
weight changes and mice survival data suggested that palmatine
administration resulted in intestinal homeostasis and improved
general condition. Based on a previous study, the median lethal
dose (LD50) of palmatine was 1,533.68 mg/kg (35). In our study, the high dose of
palmatine was 20 mg/kg, only ~1.5% of its reported LD50.
Thus, the palmatine doses in this study should be safe and without
toxicity to the animals. Further safety and pharmacokinetic
observations of palmatine will be conducted in our future
studies.
Our gut histological observations showed that,
compared with the model group, the inflammatory lesions in the
small intestine and colon in the palmatine treatment groups were
markedly reduced, with less neutrophil infiltration and focal
adenomatous change. In addition, the dysplastic changes were
greatly reduced, supporting the effects of palmatine on gut
inflammation and tumorigenesis. Furthermore, RT-qPCR analysis data
showed that, compared with the model group, palmatine significantly
reduced the gene expression of several inflammatory cytokines,
IL-1α, IL1-β, IL-8, G-CSF and GM-CSF, in the small intestine and
colon. In addition, levels of IL-8, IL-10 and G-CSF in the serum
were also significantly reduced following palmatine administration.
As an anti-inflammatory compound, palmatine likely suppresses the
overexpression of inflammatory cytokines in cancerous and
non-cancerous tissues.
In conclusion, using a gut-specific inflammation and
tumorigenesis
ApcMin/+ mouse
model, it was reported that palmatine significantly reduced gut
inflammation, tumor initiation and tumor progression. The observed
effects were supported by the body weight changes, organ index
results, mice survival results, gut tissue histology, and gut
inflammation cytokine data. Results obtained from the current study
suggest that palmatine may have a therapeutic role in colorectal
cancer chemoprevention.
Acknowledgments
This study was supported in part by a grant (grant
no. BK20131309) from The Natural Science Foundation of Jiangsu
Province (China), and NIH/NCCAM grants (grant nos. AT 004418 and
AT005362).
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics. CA Cancer J Clin. 64:104–117. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McCarthy N: Tumorigenesis: All together
now. Nat Rev Cancer. 13:148–149. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Madka V and Rao CV: Anti-inflammatory
phytochemicals for chemoprevention of colon cancer. Curr Cancer
Drug Targets. 13:542–557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moreno-Jimenez MR, Trujillo-Esquivel F,
Gallegos-Corona MA, Reynoso-Camacho R, González-Laredo RF,
Gallegos-Infante JA, Rocha-Guzmán NE and Ramos-Gomez M:
Antioxidant, anti-inflammatory and anticarcinogenic activities of
edible red oak (Quercus spp.) infusions in rat colon carcinogenesis
induced by 1,2-dimethylhydrazine. Food Chem Toxicol. 80:144–153.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu C, Wen XD, Zhang Z, Zhang CF, Wu XH,
Martin A, Du W, He TC, Wang CZ and Yuan CS: American ginseng
attenuates azoxymethane/dextran sodium sulfate-induced colon
carcinogenesis in mice. J Ginseng Res. 39:14–21. 2015. View Article : Google Scholar
|
|
6
|
Saleem M: Lupeol, a novel
anti-inflammatory and anti-cancer dietary triterpene. Cancer Lett.
285:109–115. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang X, Song ZJ, He X, Zhang RQ, Zhang CF,
Li F, Wang CZ and Yuan CS: Antitumor and immunomodulatory activity
of genkwanin on colorectal cancer in the APC(Min/+) mice. Int
Immunopharmacol. 29:701–707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chinese Pharmacopoeia Commission:
Pharmacopoeia of the People's Republic of China (Part 1) China.
Medical Science Press; Beijing: 2010
|
|
9
|
Bhadra K and Kumar GS: Therapeutic
potential of nucleic acid-binding isoquinoline alkaloids: Binding
aspects and implications for drug design. Med Res Rev. 31:821–862.
2011. View Article : Google Scholar
|
|
10
|
Zhang L, Li J, Ma F, Yao S, Li N, Wang J,
Wang Y, Wang X and Yao Q: Synthesis and cytotoxicity evaluation of
13-n-alkyl berberine and palmatine analogues as anticancer agents.
Molecules. 17:11294–11302. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jung J, Choi JS and Jeong CS: Inhibitory
activities of palmatine from coptis chinensis against helicobactor
pylori and gastric damage. Toxicol Res. 30:45–48. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang CZ, Du GJ, Zhang Z, Wen XD, Calway T,
Zhen Z, Musch MW, Bissonnette M, Chang EB and Yuan CS: Ginsenoside
compound K, not Rb1, possesses potential chemo-preventive
activities in human colorectal cancer. Int J Oncol. 40:1970–1976.
2012.PubMed/NCBI
|
|
13
|
Wang CZ, Zhang Z, Huang WH, Du GJ, Wen XD,
Calway T, Yu C, Nass R, Zhao J, Du W, et al: Identification of
potential anticancer compounds from Oplopanax horridus.
Phytomedicine. 20:999–1006. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fichera A, Guo Y, Romero L, Michelassi F
and Arenas RB: Quantitation of in vivo gene delivery by restriction
enzyme PCR generated polymorphism. J Surg Res. 69:188–192. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mohammed A, Janakiram NB, Li Q, Choi CI,
Zhang Y, Steele VE and Rao CV: Chemoprevention of colon and small
intestinal tumorigenesis in APC (Min/+) mice by licofelone, a novel
dual 5-LOX/COX inhibitor: Potential implications for human colon
cancer prevention. Cancer Prev Res (Phila). 4:2015–2026. 2011.
View Article : Google Scholar
|
|
16
|
Dougherty U, Mustafi R, Wang Y, Musch MW,
Wang CZ, Konda VJ, Kulkarni A, Hart J, Dawson G, Kim KE, et al:
American ginseng suppresses Western diet-promoted tumorigenesis in
model of inflammation-associated colon cancer: Role of EGFR. BMC
Complement Altern Med. 11:1112011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Van der Meer R, Lapré JA, Govers MJAP and
Kleibeuker JH: Mechanisms of the intestinal effects of dietary fats
and milk products on colon carcinogenesis. Cancer Lett. 114:75–83.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jung HA, Yoon NY, Bae HJ, Min BS and Choi
JS: Inhibitory activities of the alkaloids from Coptidis Rhizoma
against aldose reductase. Arch Pharm Res. 31:1405–1412. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Yu C, Zhang CF, Wu XH, Wen XD,
Anderson S, Du W, Huang WH, Li SP, Wang CZ and Yuan CS:
Chemopreventive effects of oplopantriol A, a novel compound
isolated from Oplopanax horridus, on colorectal cancer. Nutrients.
6:2668–2680. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bin-Hafeez B, Haque R, Parvez S, Pandey S,
Sayeed I and Raisuddin S: Immunomodulatory effects of fenugreek
(Trigonella foenum graecum L.) extract in mice. Int
Immunopharmacol. 3:257–265. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zheng YQ, Wei W, Zhu L and Liu JX: Effects
and mechanisms of Paeoniflorin, a bioactive glucoside from paeony
root, on adjuvant arthritis in rats. Inflamm Res. 56:182–188. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang J, Xie Y, Feng Y, Zhang L, Huang X,
Shen X and Luo X: (−)-Epigallocatechingallate induces apoptosis in
B lymphoma cells via caspase-dependent pathway and Bcl-2 family
protein modulation. Int J Oncol. 46:1507–1515. 2015.PubMed/NCBI
|
|
23
|
Grimoud J, Durand H, de Souza S, Monsan P,
Ouarné F, Theodorou V and Roques C: In vitro screening of
probiotics and synbiotics according to anti-inflammatory and
anti-proliferative effects. Int J Food Microbiol. 144:42–50. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Garcia Rodriguez LA, Cea-Soriano L,
Tacconelli S and Patrignani P: Coxibs: Pharmacology, toxicity and
efficacy in cancer clinical trials. Recent Results Cancer Res.
191:67–93. 2013. View Article : Google Scholar
|
|
25
|
Chen S, Qu X, Wan P, Li QW, Wang Z, Guo F,
Bai L, Hu Z, Tan W and Li J: Norcantharidin inhibits
pre-replicative complexes assembly of HepG2 cells. Am J Chin Med.
41:665–682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu S, Wang XJ, Liu Y and Cui YF:
PI3K/AKT/mTOR signaling is involved in
(−)-epigallocatechin-3-gallate-induced apoptosis of human
pancreatic carcinoma cells. Am J Chin Med. 41:629–642. 2013.
View Article : Google Scholar
|
|
27
|
Wang CY, Bai XY and Wang CH: Traditional
Chinese medicine: A treasured natural resource of anticancer drug
research and development. Am J Chin Med. 42:543–559. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yuan A, Chen JJ, Yao PL and Yang PC: The
role of interleukin-8 in cancer cells and microenvironment
interaction. Front Biosci. 10:853–865. 2005. View Article : Google Scholar
|
|
29
|
De Robertis M, Massi E, Poeta ML, Carotti
S, Morini S, Cecchetelli L, Signori E and Fazio VM: The AOM/DSS
murine model for the study of colon carcinogenesis: From pathways
to diagnosis and therapy studies. J Carcinog. 10:92011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wen XD, Wang CZ, Yu C, Zhang Z, Calway T,
Wang Y, Li P and Yuan CS: Salvia miltiorrhiza (dan shen)
significantly ameliorates colon inflammation in dextran sulfate
sodium induced colitis. Am J Chin Med. 41:1097–1108. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gu C, Qiao J, Zhu M, Du J, Shang W, Yin W,
Wang W, Han M and Lu W: Preliminary evaluation of the interactions
of Panax ginseng and Salvia miltiorrhiza Bunge with 5-fluorouracil
on pharmacokinetics in rats and pharmacodynamics in human cells. Am
J Chin Med. 41:443–458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park EY, Kim MH, Kim EH, Lee EK, Park IS,
Yang DC and Jun HS: Efficacy comparison of Korean ginseng and
American ginseng on body temperature and metabolic parameters. Am J
Chin Med. 42:173–187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Qi LW, Wang CZ and Yuan CS: Ginsenosides
from American ginseng: Chemical and pharmacological diversity.
Phytochemistry. 72:689–699. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jacoby RF, Seibert K, Cole CE, Kelloff G
and Lubet RA: The cyclooxygenase-2 inhibitor celecoxib is a potent
preventive and therapeutic agent in the min mouse model of
adenomatous polyposis. Cancer Res. 60:5040–5044. 2000.PubMed/NCBI
|
|
35
|
Yi J, Ye X, Wang D, He K, Yang Y, Liu X
and Li X: Safety evaluation of main alkaloids from Rhizoma
Coptidis. J Ethnopharmacol. 145:303–310. 2013. View Article : Google Scholar
|