Introduction
Numerous large-scale clinical studies have suggested
that hypertension is a risk factor for cardiovascular diseases
(1–3). High blood pressure promotes the
development of cardiovascular diseases via various pathological
processes, including endothelial dysfunction (4), inflammation (5) and myocardial fibrosis (5,6).
Though the pathogenesis of myocardial fibrosis in hypertension has
previously been heavily investigated, the mechanism remains to be
fully elucidated.
Periostin is a 90-kDa secretory protein that was
previously demonstrated to be secreted from a mouse osteoblastic
cell line (7). As an extracellular
matrix (ECM) protein, periostin is prominently expressed during
tissue fibrosis in the kidney (8–10),
lung (11,12) and heart (13,14).
The heart ventricles are rich in ECM secreted by fibroblasts, thus,
it has been hypothesized that periostin is involved in
hypertension-induced myocardial fibrosis.
Previous studies indicate that oxidative stress is
involved in the development of hypertension-induced myocardial
fibrosis (15–17), however, the exact mechanisms by
which oxidative stress induces interstitial fibrosis remain to be
elucidated. Multiple lines of evidence have indicated that
oxidative stress accelerates fibrosis via regulation of profibrotic
factors and ECM, thus, the present study hypothesizes that
oxidative stress may contribute to periostin expression. The
present study aimed to determine the effect of periostin in
myocardial fibrosis in an experimental hypertensive rat model and
to investigate the underlying molecular mechanisms.
Materials and methods
Experimental animals and treatment
The experimental and feeding protocols were approved
by the Ethics Committee of Drum Tower Hospital (Nanjing, China).
Rats (4 weeks old, male, n=16; weight, 90–115 g) were housed
together in an animal room at 22°C, normal atmosphere and 12 h
light/dark cycles, with free access to normal drinking water and
fed chow with 0.4% NaCl (normal group; n=8) or 8% NaCl
(hypertension group; n=8) for 36 weeks. At the end of the study,
the rats were anesthetized with 3% isoflurane and 8 hearts from
each group were rapidly excised and divided into three parts.
Cardiac fibroblasts used in present study were isolated from
another 10 normal rats. Physiological parameters, including blood
pressure and heart rate, were monitored every 4 weeks (BP-2006A;
Softron, Tokyo, Japan) until the animals were sacrificed.
Dihydroethidium (DHE) staining and
Masson's staining
At the endpoint of the experiment, the hearts were
rapidly excised from the rats and were divided into three parts.
One part was treated routinely and the others were flash frozen.
The tissue was fixed and embedded with optimal cutting temperature
compound (Department of Pathology, Drum Tower Hospital, Nanjing.
China), and was subsequently cut into 5 μm sections. The
frozen heart sections were then incubated with 4 μm/ml DHE
dye (Beyotime Institute of Biotechnology, Haimen, China) for 30 min
at 37°C. Oxidative stress was indicated by red staining and
visualized using a fluorescence microscope. Image Pro Plus 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA) was used to
quantify the red staining in three randomly selected fields in each
section (n=16). The second part of the tissue samples were fixed
with 4% paraformaldehyde, embedded in wax and cut into 5-μm
sections. Masson's staining was used to assess myocardial fibrosis.
Collagen volume fraction was analyzed using Image Pro Plus 6.0
software to quantify the collagen deposition in the heart tissue.
The remaining tissue was used for western blot analysis.
Isolation of fibroblasts
Cardiac fibroblasts were isolated from adult rats as
previously described (18).
Briefly, following anesthesia with 3% isoflurane of the rats, the
hearts were rapidly excised, rinsed and homogenized in cold
phosphate-buffered saline (PBS). Then, the tissues were digested in
Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan, TX, USA)
containing 0.1% collagenase type II (Nanjing SunShine Biotechnology
Co., Ltd., Nanjing, China) at 37°C. The resulting supernatant was
centrifuged at 36°C for 30 min at 500 × g and resuspended, prior to
seeding in DMEM containing 10% fetal bovine serum (Hyclone). After
1 h, the supernatant was removed and the adherent cells were
maintained as cardiac fibroblasts. Unless otherwise stated, cardiac
fibroblasts at passage 1 to 3, cultured at 37°C and 5%
CO2, were used in the present study and were
serum-starved overnight prior to treatments with Ang II (1, 2 or 4
μM; Sigma-Aldrich, St. Louis, MO, USA) or NAC (5 or 10 nM;
Beyotime Institute of Biotechnology, Haimen, China) for 24 h.
Immunofluorescence for α-smooth muscle
actin (α-SMA)
When adherent cells reached 80–90% confluency, they
were seeded in 24-well plates. Following treatment with Ang II (4
μM) or NAC (10 nM) for 24 h, the fibroblasts were washed
with PBS 3 times and fixed with 4% paraformaldehyde for 60 min at
room temperature. After blocking with 2% BSA at room temperature
for 1 h, the cells were incubated with rabbit anti-α-SMA primary
antibody (cat. no. ab5694; 1:100; Abcam, Cambridge, UK). Following
two washes with PBS, the cells were incubated with Alexa
Fluor® 594-conjugated goat anti-rabbit secondary
antibody (cat. no. A-11037; 1:400; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) for 1 h under dark conditions
at room temperature. Finally, the cells were counterstained with
4′,6-diamidino-2-phenylindole (Sigma-Aldrich, St. Louis, MO, USA)
for 10 min, and then visualized using a confocal laser scanning
microscope.
Measurement of intracellular reactive
oxygen species (ROS) in fibroblasts
Intracellular ROS generation was detected using the
fluorescent probe 2′,7′-dichlorofluorescin diacetate (DCFH).
Briefly, fibroblasts were plated in 24-well plates. Following
treatment with Ang II or NAC, the medium was removed and the cells
were washed with PBS. A solution of 5 μM DCFH probe in
serum-free DMEM was then added to the cells for 30 min at 37°C. The
plates were washed twice and the intracellular ROS levels were
detected by microscope.
Western blotting
Tissue or treated cells were homogenized in lysis
buffer containing a 1:100 dilution of protease inhibitor
(Sigma-Aldrich). The lysates were centrifuged at 13,800 × g for 10
min and proteins were harvested from the supernatants. Protein
concentration was detected using a bicinchoninic acid protein assay
kit (Pierce, Rockford, IL, USA). Equal quantities of proteins (30
μg) were separated by 10% SDS-PAGE and electroblotted to
polyvinylidene fluoride membranes. Following blocking with 5% milk,
the membranes were incubated with primary antibodies overnight at
4°C. The membranes were washed with Tris-buffered saline containing
0.1% Tween 20 then incubated with secondary antibody (cat. no.
sc-2004; 1:10,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
for 2 h at room temperature. Proteins were detected using enhanced
chemiluminescence reagents (Pierce) and images were obtained by
exposure to films. The densities of each band were analyzed using
Bio-Rad Quantity One v4.62 imaging software (Bio-Rad Laboratories,
Inc., Hercules, CA USA). The results are presented as a percentage
change compared with that in controls following normalization to
the β-actin bands for each sample. The following antibodies were
used: Anti-α-SMA (cat. no. ab5654; 1:1,000; Abcam); anti-β-actin
(cat. no. sc-130657; 1:2,000; Santa Cruz Biotechnology, Inc.),
anti-periostin (cat. no. sc-67233; 1:500; Santa Cruz Biotechnology,
Inc.), anti-p47phox (cat. no. A1148; 1:1,000; ABclonal Biotech Co.,
Ltd., Cambridge, MA, USA), anti-gp91phox (cat. no. sc-20782;
1:1,000; Santa Cruz Biotechnology, Inc.), and anti-p66shc (cat. no.
sc-1695; 1:1,000; Santa Cruz). All blots were performed on three
independent occasions.
Statistical analysis
Analyses were performed using SPSS software (version
21.0; IBM SPSS, Armonk, NY, USA). Differences between 2 groups were
analyzed by Student's t-test and one-way analysis of variance was
used to test significance for >2 groups. Student-Neuman-Keuls or
Dunnett's T3 tests were used for post-hoc multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Blood pressure increased in high-salt
induced hypertensive rats
As a high salt diet is an important risk factor in
hypertension, an in vivo hypertension model was produced
with a sustained high salt diet for 36 weeks. At the end of the
experimental period, blood pressure levels and heart rate of the
animals were measured. Compared with the normal rats (0.4% NaCl
diet), systolic blood pressure (P<0.01; Fig. 1A), diastolic blood pressure
(P<0.05; Fig. 1B) and mean
blood pressure (P<0.05; Fig.
1C) were significantly increased in the high salt (8% NaCl
diet)-induced hypertensive rats. At week 36, no significant
differences in heart rate were observed between the 2 groups
(Fig. 1D).
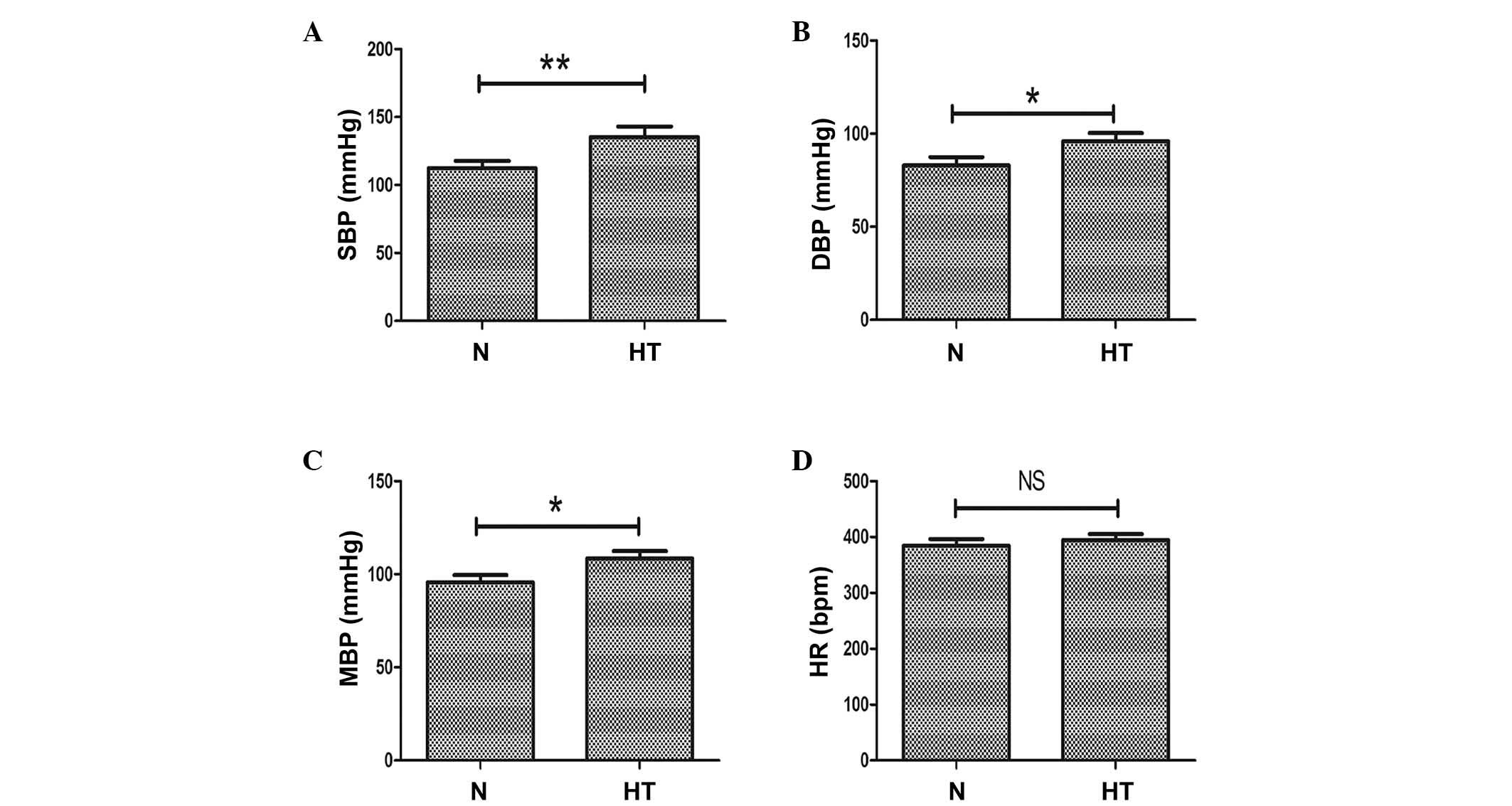 | Figure 1Evaluation of SBP, DBP, MBP and HR in
normal and high-salt induced hypertensive rats. (A) SBP, (B) DBP
and (C) MBP measurements in high salt-induced hypertensive and
normal rats. (D) HR in normal and hypertensive rats. n=8 per group,
*P<0.05, **P<0.01, comparisons
indicated by brackets. N, normal rats group; HT, hypertensive rats
group; SBP, systolic blood pressure; DBP, diastolic blood pressure;
MBP, mean blood pressure; HR, heart rate; NS, not significant. |
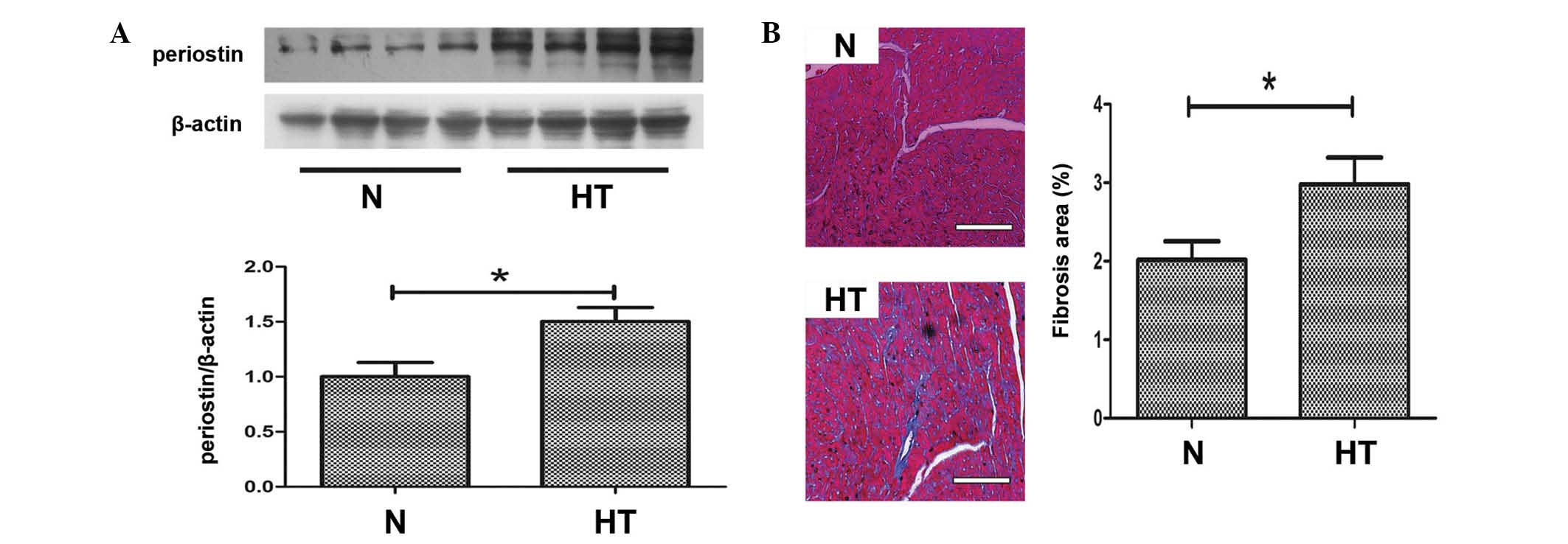
Periostin is upregulated and myocardial
fibrosis observed in hypertensive rats
Periostin has previously been considered to be an
important mediator of fibrosis, thus, western blotting was
performed to detect the expression levels of periostin in the
hearts of the 2 groups. As demonstrated in Fig. 2A, the protein expression levels of
periostin in hypertensive hearts was significantly upregulated
compared with normal hearts (P<0.05). Histological observations
following Masson's trichrome staining indicated that myocardial
fibrosis was induced in hypertensive rats compared with normal rats
(P<0.05; Fig. 2B).
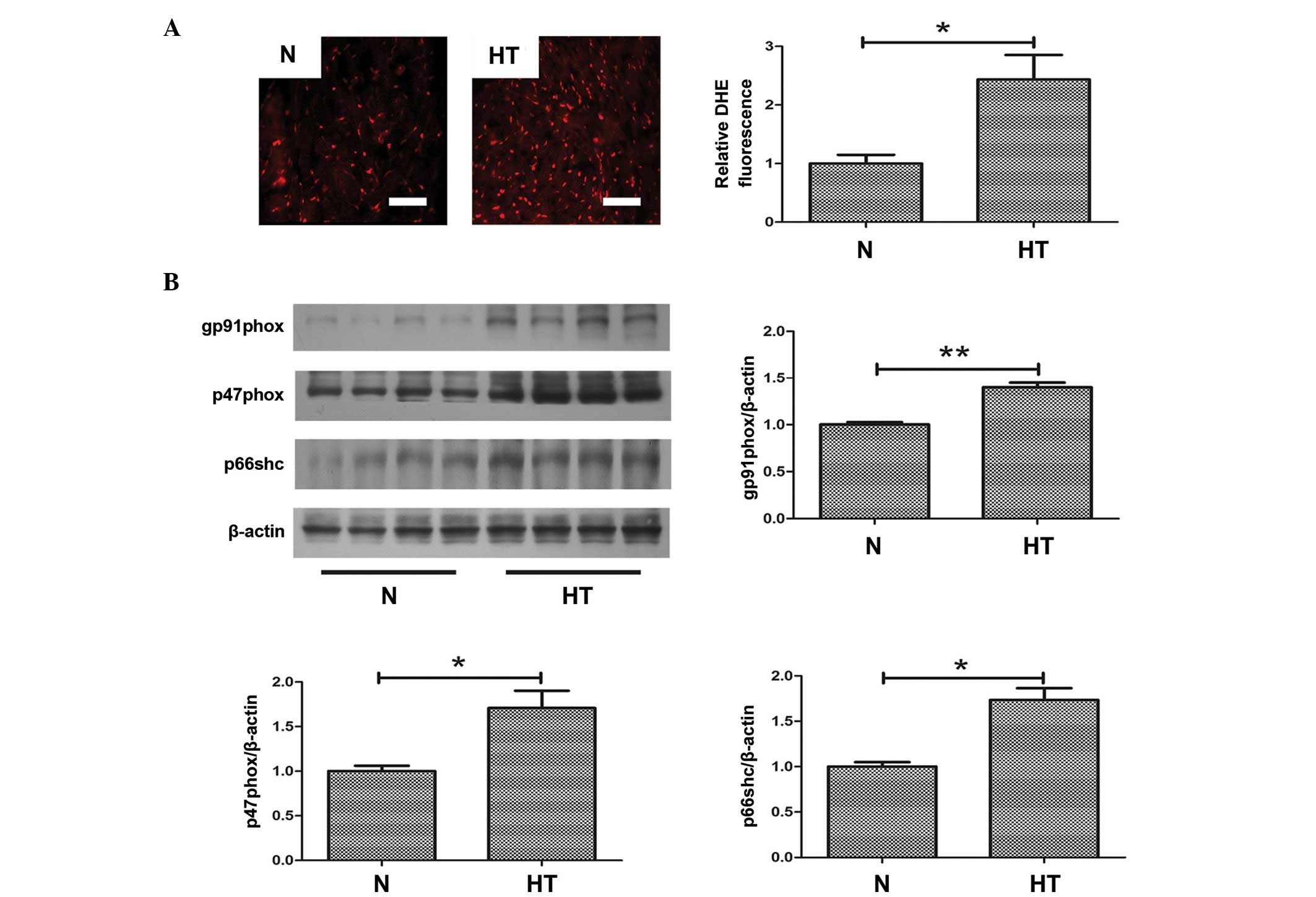
Oxidative stress is increased in
hypertensive hearts
To investigate the role of oxidative stress in
hypertensive fibrosis, ROS production was measured in samples from
normal and hypertensive rats. As demonstrated in Fig. 3A, oxidative stress, indicated by
DHE staining, was significantly increased in hypertensive hearts
compared with normal hearts (P<0.05; Fig. 3A). Furthermore, to evaluate the
increased oxidative stress in hypertensive hearts, the expression
levels of oxidative stress-associated proteins were determined.
Western blotting demonstrated that the protein expression level of
gp91phox (P<0.01), p47phox (P<0.05) and p66shc (P<0.05)
were significantly increased in the hearts of hypertensive rats
compared with normal rats (Fig.
3B). These proteins all contribute to oxidative stress.
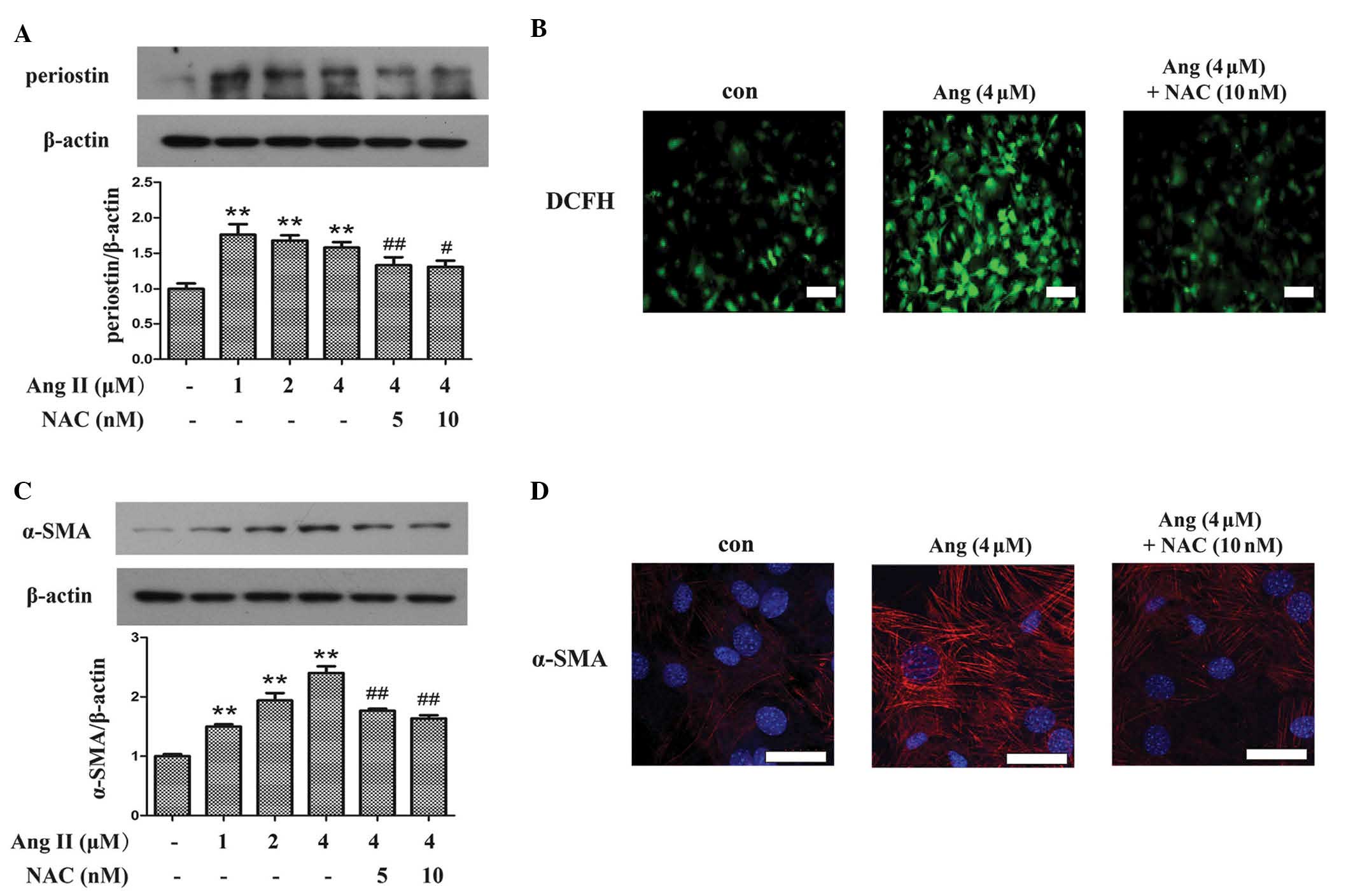
Angiotensin II (Ang II) increases
expression of periostin and α-SMA in cardiac fibroblasts via
oxidative stress
Ang II is important for the initiation of myocardial
fibrosis, thus, in vitro studies using Ang II to treat
cardiac fibroblasts were performed to investigate oxidative stress
and periostin expression levels in hypertensive fibrosis. As
demonstrated in Fig. 4A, compared
with the control group, incubation of fibroblasts with Ang II
significantly upregulated the expression of periostin (P<0.01),
whereas the expression was suppressed by pretreating with
N-acetyl-L-cysteine (NAC), an inhibitor of ROS (P<0.01 and
P<0.05 for 5 and 10 nM NAC, respectively). The anti-oxidative
effect of NAC was demonstrated using a DCFH probe, which indicated
that NAC attenuated Ang II-induced ROS production in fibroblasts
(Fig. 4B). To investigate the
association between periostin and the differentiation of
fibroblasts, the expression of α-SMA was detected. Western blot
analysis (Fig. 4C) and
immunofluorescence (Fig. 4D)
demonstrated that the increased expression levels of α-SMA induced
by Ang II compared with control, was abrogated by NAC treatment
(P<0.01).
Discussion
The current study demonstrated the contribution of
oxidative stress-induced periostin to the development of myocardial
fibrosis in hypertension.
Hypertensive myocardial fibrosis contributing to
myocardial remodeling is the pathophysiological basis of
hypertension and eventually results in heart failure. Myocardial
fibrosis is considered to be an early event of heart failure by
reducing contractile efficiency and demanding greater cardiac
contractile force. Fibrosis is a complex condition mediated by
profibrotic factors, an imbalance of collagen synthesis and
degradation, upregulation of ECM and increased oxidative stress.
The present study focused on ECM and oxidative stress in
hypertensive hearts.
Increased periostin and oxidative stress
in hypertensive rats
Periostin, a member of the fasciclin family, is an
ECM protein that promotes progression of fibrosis via regulating
ECM homeostasis (19,20). Periostin was previously
demonstrated to be upregulated in numerous heart diseases. A
previous study by Zhao et al (14) demonstrated that periostin
expression was upregulated and associated with myocardial fibrosis
in human failing hearts. Another investigation in mice following
transverse aortic constriction confirmed the effect of periostin in
heart failure (13). Meyer et
al (21) demonstrated that
periostin expression levels as a biomarker associated with ECM
remodeling contributing to heart failure were significantly higher
in men compared with women. The results of the present study
demonstrated that periostin was also increased in hypertensive
hearts. This finding is consistent with another investigation that
demonstrated that eplerenone enhanced cardioprotective effects in
hypertensive heart failure by regulating ECM proteins, including
periostin (22). However, to the
best of our knowledge, the mechanisms that regulate periostin
expression have not been previously investigated.
Oxidative stress
A major source of ROS in cardiovascular diseases is
the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase
family (17). NADPH oxidase
consists of multiple subunits: Membrane-bound proteins (including
gp91phox); cytosolic regulation subunits (such as p47phox); and a
small GTP-binding protein Rac 1 (23). The current study demonstrated that
p47phox and gp91phox were upregulated in hypertensive hearts, which
is consistent with the findings of others studies suggesting that
NADPH oxidase contributes to kidney and aortic media damage in
hypertension (24,25).
In addition, p66shc, an isoform of SRC transforming
protein 1, has also been recognized as an important mediator in
oxidative stress in the cardiovascular system (26–28).
Notably, consistent with previous findings (29), the present study demonstrated that
p66shc modulates ROS production in hypertension.
Furthermore, DHE staining was also used to evaluate
oxidative stress in hypertensive rats. DHE staining further
supported the results of ph47phox, gp91phox and p66shc, suggesting
that hypertensive hearts exhibited increased oxidative stress
compared with normal hearts. Consistently with the findings of the
present study, an accumulating body of evidence suggests that
oxidative stress is an essential characteristic of hypertension
(30–33). Thus, considering the present study
and previous findings in hypertension, ROS generation may be
important for the development of hypertension.
Ang II increases expression of periostin
via oxidative stress
As periostin and oxidative stress were upregulated
in hypertension, an in vitro experiment to investigate their
association was conducted.
A number of studies have demonstrated that Ang II is
the most important hormone that regulates myocardial fibrosis
(18,34), which is supported by previous
studies that have demonstrated that Ang II antagonists can prevent
fibrosis in various diseases (33,35).
Ang II promotes systemic arteriole contraction that results in
hypertension via binding Ang II type I receptor, and contributes to
myofibrosis in hypertension-associated heart diseases. Following
stimulation by certain factors, including Ang II, cardiac
fibroblasts can differentiate to myofibroblasts, exhibiting
increased expression of a number of cytoskeletal protein, including
α-SMA and collagen. Differentiation of cardiac fibroblasts into
myofibroblasts contributes to cardiac fibrosis and eventually
affects the dynamic contractile properties of the heart (36). Thus, the present study performed
in vitro experiments using cardiac fibroblasts treated with
Ang II to investigate the effect of oxidative stress and periostin
in hypertensive fibrosis.
The current study demonstrated that Ang II increased
the expression of periostin in fibroblasts, which is also supported
by the previous finding that periostin was induced by Ang II in
cardiac fibroblasts (13).
Notably, in the present study, upregu-lation of periostin was
accompanied by an increase in ROS generation, while inhibition of
ROS blocked the induction of periostin, suggesting that Ang II
upregulates the expression of periostin via an oxidative
stress-mediated signaling pathway. To the best of our knowledge,
this is the first report demonstrating that oxidative stress also
regulates periostin expression induced by Ang II.
Consistent with previous findings, the present study
demonstrated that Ang II upregulates the expression of α-SMA in
fibroblasts by western blot analysis and immunofluores-cence
(18), suggesting that Ang II
induces differentiation of cardiac fibroblasts into myofibroblasts.
Notably, the results indicated that ROS-induced periostin
expression may be important for Ang II-induced differentiation of
cardiac fibroblasts.
In conclusion, the results of the present study
demonstrated that oxidative stress-induced upregulation of
periostin may be important for in myocardial fibrosis in
hypertensive rats. Therefore, periostin may serve as a potential
target for the prevention of hypertension-induced myocardial
fibrosis.
Acknowledgments
The current study was supported by grants from the
Natural Science Foundation of China (grant nos. 81070195, 81200148
and 81270281), the Jiangsu Key Laboratory for Molecular Medicine of
Nanjing University, Jiangsu Provincial Special Program of Medical
Science (grant no. BL2012014), the State Key Laboratory of
Pharmaceutical Biotechnology (grant no. KF-GN-200901), the Peak of
Six Personnel in Jiangsu Province (grant no. 2013-WSN-008), Funds
for Distinguished Young Scientists in Nanjing (grant no. JQX13006),
and Natural Science Foundation of Jiangsu Province (grant no.
BK2010107).
References
|
1
|
Barasch E, Gottdiener JS, Aurigemma G,
Kitzman DW, Han J, Kop WJ and Tracy RP: Association between
elevated fibrosis markers and heart failure in the elderly: The
cardiovascular health study. Circ Heart Fail. 2:303–310. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kannan A and Janardhanan R: Hypertension
as a risk factor for heart failure. Curr Hypertens Rep. 16:4472014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Volpe M, McKelvie R and Drexler H:
Hypertension as an underlying factor in heart failure with
preserved ejection fraction. J Clin Hypertens (Greenwich).
12:277–283. 2010. View Article : Google Scholar
|
|
4
|
Lapu-Bula R and Ofili E: From hypertension
to heart failure: Role of nitric oxide-mediated endothelial
dysfunction and emerging insights from myocardial contrast
echocardiography. Am J Cardiol. 99:7D–14D. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Palaniyandi SS, Inagaki K and Mochly-Rosen
D: Mast cells and epsilonPKC: A role in cardiac remodeling in
hypertension-induced heart failure. J Mol Cell Cardiol. 45:779–786.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mori T, Kai H, Kajimoto H, Koga M, Kudo H,
Takayama N, Yasuoka S, Anegawa T, Kai M and Imaizumi T: Enhanced
cardiac inflammation and fibrosis in ovariectomized hypertensive
rats: A possible mechanism of diastolic dysfunction in
postmenopausal women. Hypertens Res. 34:496–502. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takeshita S, Kikuno R, Tezuka K and Amann
E: Osteoblast-specific factor 2: Cloning of a putative bone
adhesion protein with homology with the insect protein fasciclin I.
Biochem J. 294:271–278. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wallace DP, White C, Savinkova L, Nivens
E, Reif GA, Pinto CS, Raman A, Parnell SC, Conway SJ and Fields TA:
Periostin promotes renal cyst growth and interstitial fibrosis in
polycystic kidney disease. Kidney Int. 85:845–854. 2014. View Article : Google Scholar :
|
|
9
|
Bible E: Polycystic kidney disease:
Periostin is involved in cell proliferation and interstitial
fibrosis in polycystic kidney disease. Nat Rev Nephrol. 10:662014.
View Article : Google Scholar
|
|
10
|
Mael-Ainin M, Abed A, Conway SJ, Dussaule
JC and Chatziantoniou C: Inhibition of periostin expression
protects against the development of renal inflammation and
fibrosis. J Am Soc Nephrol. 25:1724–1736. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Naik PK, Bozyk PD, Bentley JK, Popova AP,
Birch CM, Wilke CA, Fry CD, White ES, Sisson TH, Tayob N, et al:
Periostin promotes fibrosis and predicts progression in patients
with idiopathic pulmonary fibrosis. Am J Physiol Lung Cell Mol
Physiol. 303:L1046–L1056. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Okamoto M, Hoshino T, Kitasato Y, Sakazaki
Y, Kawayama T, Fujimoto K, Ohshima K, Shiraishi H, Uchida M, Ono J,
et al: Periostin, a matrix protein, is a novel biomarker for
idiopathic interstitial pneumonias. Eur Respir J. 37:1119–1127.
2011. View Article : Google Scholar
|
|
13
|
Liu W, Zi M, Tsui H, Chowdhury SK, Zeef L,
Meng QJ, Travis M, Prehar S, Berry A, Hanley NA, et al: A novel
immunomodulator, FTY-720 reverses existing cardiac hypertrophy and
fibrosis from pressure overload by targeting NFAT (nuclear factor
of activated T-cells) signaling and periostin. Circ Heart Fail.
6:833–844. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao S, Wu H, Xia W, Chen X, Zhu S, Zhang
S, Shao Y, Ma W, Yang D and Zhang J: Periostin expression is
upregulated and associated with myocardial fibrosis in human
failing hearts. J Cardiol. 63:373–378. 2014. View Article : Google Scholar
|
|
15
|
Redout EM, van der Toorn A, Zuidwijk MJ,
van de Kolk CW, van Echteld CJ, Musters RJ, van Hardeveld C, Paulus
WJ and Simonides WS: Antioxidant treatment attenuates pulmonary
arterial hypertension-induced heart failure. Am J Physiol Heart
Circ Physiol. 298:H1038–H1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Worou ME, Belmokhtar K, Bonnet P, Vourc'h
P, Machet MC, Khamis G and Eder V: Hemin decreases cardiac
oxidative stress and fibrosis in a rat model of systemic
hypertension via PI3K/Akt signalling. Cardiovasc Res. 91:320–329.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paravicini TM and Touyz RM: NADPH
oxidases, reactive oxygen species, and hypertension: Clinical
implications and therapeutic possibilities. Diabetes Care. 31(Suppl
2): S170–S180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bai J, Zhang N, Hua Y, Wang B, Ling L,
Ferro A and Xu B: Metformin inhibits angiotensin II-induced
differentiation of cardiac fibroblasts into myofibroblasts. PLoS
One. 8:e721202013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamaguchi Y: Periostin in skin tissue and
skin-related diseases. Allergol Int. 63:161–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu AY, Zheng H and Ouyang G: Periostin, a
multifunctional matricellular protein in inflammatory and tumor
microenvironments. Matrix Biol. 37:150–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meyer S, van der Meer P, van Deursen VM,
Jaarsma T, van Veldhuisen DJ, van der Wal MH, Hillege HL and Voors
AA: Neurohormonal and clinical sex differences in heart failure.
Eur Heart J. 34:2538–2547. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Muñoz-Pacheco P, Ortega-Hernández A,
Caro-Vadillo A, Casanueva-Eliceiry S, Aragoncillo P, Egido J,
Fernández-Cruz A and Gómez-Garre D: Eplerenone enhances
cardioprotective effects of standard heart failure therapy through
matricellular proteins in hypertensive heart failure. J Hypertens.
31:2309–2318; discussion 2319. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Elnakish MT, Hassanain HH, Janssen PM,
Angelos MG and Khan M: Emerging role of oxidative stress in
metabolic syndrome and cardiovascular diseases: Important role of
Rac/NADPH oxidase. J Pathol. 231:290–300. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian N, Moore RS, Phillips WE, Lin L,
Braddy S, Pryor JS, Stockstill RL, Hughson MD and Manning RD Jr:
NADPH oxidase contributes to renal damage and dysfunction in Dahl
salt-sensitive hypertension. Am J Physiol Regul Integr Comp
Physiol. 295:R1858–R1865. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Akasaki T, Ohya Y, Kuroda J, Eto K, Abe I,
Sumimoto H and Iida M: Increased expression of gp91phox homologues
of NAD (P)H oxidase in the aortic media during chronic
hypertension: Involvement of the renin-angiotensin system.
Hypertens Res. 29:813–820. 2006. View Article : Google Scholar
|
|
26
|
Lebiedzińska M, Suski J, Duszyński J and
Wieckowski MR: Role of the p66Shc protein in physiological state
and in pathologies. Postepy Biochem. 56:165–173. 2010.In
Polish.
|
|
27
|
Lee SK, Chung JI, Park MS, Joo HK, Lee EJ,
Cho EJ, Park JB, Ryoo S, Irani K and Jeon BH: Apurinic/apyrimidinic
endonuclease 1 inhibits protein kinase C-mediated p66shc
phos-phorylation and vasoconstriction. Cardiovasc Res. 91:502–509.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Di Stefano V, Cencioni C, Zaccagnini G,
Magenta A, Capogrossi MC and Martelli F: P66ShcA modulates
oxidative stress and survival of endothelial progenitor cells in
response to high glucose. Cardiovasc Res. 82:421–429. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Camici GG, Sudano I, Noll G, Tanner FC and
Lüscher TF: Molecular pathways of aging and hypertension. Curr Opin
Nephrol Hypertens. 18:134–137. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin CX, Rhaleb NE, Yang XP, Liao TD,
D'Ambrosio MA and Carretero OA: Prevention of aortic fibrosis by
N-acetyl-seryl-asp artyl-lysyl-proline in angiotensin II-induced
hypertension. Am J Physiol Heart Circ Physiol. 295:H1253–H1261.
2008. View Article : Google Scholar
|
|
31
|
Rizzi E, Guimaraes DA, Ceron CS, Prado CM,
Pinheiro LC, Martins-Oliveira A, Gerlach RF and Tanus-Santos JE:
β1-Adrenergic blockers exert antioxidant effects, reduce matrix
metalloproteinase activity, and improve renovascular
hypertension-induced cardiac hypertrophy. Free Radic Biol Med.
73:308–317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen X, Mori T, Guo Q, Hu C, Ohsaki Y,
Yoneki Y, Zhu W, Jiang Y, Endo S, Nakayama K, et al: Carbonyl
stress induces hypertension and cardio-renal vascular injury in
Dahl salt-sensitive rats. Hypertens Res. 36:361–367. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chaykovska L, Alter ML, von Websky K,
Hohmann M, Tsuprykov O, Reichetzeder C, Kutil B, Kraft R, Klein T
and Hocher B: Effects of telmisartan and linagliptin when used in
combination on blood pressure and oxidative stress in rats with
2-kidney-1-clip hypertension. J Hypertens. 31:2290–2298; discussion
2299. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu J, Carretero OA, Liao TD, Peng H,
Shesely EG, Xu J, Liu TS, Yang JJ, Reudelhuber TL and Yang XP:
Local angiotensin II aggravates cardiac remodeling in hypertension.
Am J Physiol Heart Circ Physiol. 299:H1328–H1338. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yamada Y, Tsuboi K, Hattori T, Murase T,
Ohtake M, Furukawa M, Ueyama J, Nishiyama A, Murohara T and Nagata
K: Mechanism underlying the efficacy of combination therapy with
losartan and hydrochlorothiazide in rats with salt-sensitive
hypertension. Hypertens Res. 34:809–816. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Leask A: Potential therapeutic targets for
cardiac fibrosis: TGFbeta, angiotensin, endothelin, CCN2 and PDGF,
partners in fibroblast activation. Circ Res. 106:1675–1680. 2010.
View Article : Google Scholar : PubMed/NCBI
|