Introduction
It is estimated that more than 6 million children
receive anesthesia every year (1).
In modern anesthesiology, general anesthesia is the most common
practice for surgery or relief from procedural pain, particularly
in children (2). A previous study
suggested that anesthetic exposure can lead to neurotoxicity in the
developing brain (3). Similarly,
children exposed to anesthetics during early life have been
reported to exhibit a higher incidence of learning deficits at
adolescence (4). These results
prompt concerns regarding the possible detrimental effects of
commonly used anesthetics in the pediatric population.
An alkyl phenol derivative, 2,6-di-isopropylphenol
(propofol), was introduced as a sedative and anesthetic agent in
1977 due to its rapid onset and recovery (5). It demonstrated neuroprotective
effects in the adult brain in vivo (6) and in vitro (7). However, in the developing brain,
propofol is able to cause neurotoxicity by inducing apoptosis
(8), disturbing neuronal
proliferation (9) and disrupting
synapse formation (10).
Additionally, long-term behavioral deficits in pups exposed to
propofol have been previously observed (8). The cellular mechanisms and signaling
pathways that underlie propofol-induced neurotoxicity are poorly
understood. The phosphatidylinositol-4,5-bisphosphate 3-kinase
(PI3K)/v-akt murine thymoma viral oncogene homolog 1 (Akt)
signaling pathway is pivotal for cell growth, proliferation and
survival (11). Activation of
PI3K/Akt signaling is widely recognized to provide protection
against cerebral ischemia/reperfusion injury (12). Phosphorylation of the downstream
protein, glycogen synthase kinase-3β (GSK-3β), also improves
long-term memory in hippocampal-associated tasks (13) and inhibits propofol-induced
lysosome/mitochondrial apoptosis in macrophages (14). Whether this signaling pathway is
associated with propofol-induced neurotoxicity remains to be
determined.
Several drugs have been demonstrated to exhibit
neuroprotective effects in different cerebral injury models. Among
them, dexmedetomidine (Dex), a potent and highly selective
α2-adrenoceptor agonist and a widely used auxiliary anesthetic, has
previously been reported to reduce isoflurane- and ketamine-induced
neuroapoptosis in vivo and in vitro (15,16).
The neuroprotective effect of Dex may be associated with its
anti-oxidant (17) and
anti-apoptotic (15) activity, its
positive impact on astrocyte brain-derived neurotrophic factor
expression (18), or inhibition of
the PI3K/Akt/GSK-3β pathway (19).
However, the effect of Dex on propofol-induced neurotoxicity in the
developing brain has not been determined. Thus, the current study
aimed to examine whether Dex has an effect on apoptosis and
neurobehavioral changes elicited by repeated propofol exposure in
neonatal rats, and whether the PI3K/Akt/GSK-3β pathway is
involved.
Materials and methods
Animals and experimental groups
All experiments were conducted in accordance with
protocols approved by the Peking University Biomedical Ethics
Committee Experimental Animal Ethics Branch (Beijing, China) and
the National Institutes of Health guide for the Care and Use of
Laboratory Animals. In total, 75 male postnatal day 7 (P7) Sprague
Dawley rats (obtained from the Department of Laboratory Animal
Science, Peking University Health Science Centre, Beijing, China)
weighing 12–16 g were used in the current study. Pups were bred and
maintained under standard housing conditions (24±2°C; 12 h:12 h,
light/dark), and had access to food and water ad libitum.
Rats were randomly divided into three groups (25 animals per group)
using the random table method. Groups were defined as control
(CON), propofol administration (PRO), and Dex preconditioning prior
to propofol administration (Dex + PRO).
Dex preconditioning and propofol
administration
Dex + PRO group rats were intraperiotoneally (i.p.)
administered with Dex (Jiangsu Hengrui Medicine Co., Ltd., Jiangsu,
China) at 75 µg/kg (1 µg/ml) in saline every day for
seven consecutive days from P7 to P13. Following 20 min of Dex
preconditioning, 30 mg/kg of propofol (Diprivan; Astra-Zeneca,
London, UK) was administered (i.p.) to pups from PRO and Dex + PRO
groups every 90 min up to a cumulative dose of 90 mg/kg each day
(n=25). Pups in the CON group were injected with an equal volume of
intralipid (Huarui Pharmaceutical Co., Wuxi, China) under the same
conditions. For the anesthesia procedure, pups were placed in a
temperature-controlled incubator (28°C) until they could
successfully perform the righting reflex. To avoid the rebreathing
of CO2, the inspired CO2 concentration was
continuously monitored and maintained at <1% by adjusting the
fresh gas flow. During anesthesia, respiratory frequency and skin
color were observed to detect apnea and hypoxia. If apnea occurred,
rats received a pain stimulus; if breathing did not restart or
resuscitation efforts were necessary, rats were excluded from
further processing and analysis. The animals were observed for a
further 90 min until they were awake and were returned to their
mothers following the last injection.
Blood gas analysis and body weight
To determine whether propofol anesthesia would cause
physiological side effects, including hypoxia, hypercapnia, or
hypoglycemia, five rats in each group were selected as
cardiorespiratory control animals (n=5) following the last
injection of propofol at P7, as described in a previous study
(20). Cardiorespiratory control
rats were not used for any other part of the study. Every pup from
each group was weighed at P7, P10, P13, P17, P21, P24 and P27 to
monitor body development.
Tissue preparation
For western blot studies, at P29, five rats per
group were sacrificed by decapitation. Hippocampi were isolated
immediately on ice and stored at −80°C until used. For terminal
deoxynucleotidyl transferase-mediated dUTP nick-end labeling
(TUNEL) assays, five rat pups per group were deeply anesthetized
with sodium pentobarbital (45 mg/kg, i.p.; Sigma-Aldrich, St.
Louis, MO, USA), perfused transcardially with 0.1 M
phosphate-buffered saline (PBS; pH=7.4) and then perfused with 4%
(w/v) paraformaldehyde in 0.1 M PBS. The brains were removed and
post-fixed with the identical fixative overnight at 4°C.
Formalin-fixed hippocampal tissue was dehydrated, embedded in
paraffin, and sliced into 5 µm-thick sections.
TUNEL fluorescent assay
According to the protocol of our previous study
(21), hippocampal apoptosis was
detected by TUNEL using an in situ cell death detection kit
(Roche Applied Science, Mannheim, Germany). Apoptosis was
quantified by calculating the percentage of TUNEL-positive nuclei
out of total nuclei in an average of 20 high-power fields for each
section in a blinded manner.
Western blotting
Following experimental exposure, rat pups (n=5 per
group) were sacrificed and western blot analysis was performed as
previously described (20). In
brief, hippocampus tissues were homogenized in cold
radioimmunoprecipitation assay buffer (Applygen Technologies Inc.,
Beijing, China), and the quantity of protein in the supernatants
was determined using a bicinchoninic acid protein assay kit
(Applygen Technologies Inc.). Protein samples (60 mg protein/lane)
were separated by 8 or 10% SDS-PAGE. Following transfer to
nitrocellulose membranes, the blots were probed using the following
primary antibodies: Rabbit anti-Akt (Ser473; cat. no.
9272), anti-phosphorylated (p)-Akt (Ser473; cat. no.
9271), anti-GSK-3β (Ser9; cat. no. 9315), and
anti-p-GSK-3β (Ser9; cat. no. 9336) antibodies (1:1,000;
Cell Signaling Technology, Inc., Danvers, MA, USA); and rabbit
anti-B-cell CLL/lymphoma 2 (Bcl-2; cat. no. sc-783),
anti-Bcl-2-associated X protein (Bax; cat. no. sc-526) and
anti-caspase-3 (cat. no. 7148) antibodies (1:200; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). Fluorescent secondary
antibodies (1:10,000; cat. nos. 926–32211 and 926–32210; LI-COR,
Inc., Lincoln, NE, USA) were used to detect the binding of primary
antibodies. Proteins were visualized by scanning the membrane on an
Odyssey Infrared Imaging System (version 3.0; LI-COR, Inc.).
Behavioral studies
As described in a previous study (20), the Morris water maze (MWM) test,
including place and probe trials, was used to evaluate spatial
learning and memory of rats (P29–P33) in the three groups (n=10) by
investigators blinded to the group conditions. At P28, a day before
the formal MWM test, rats were checked for their ability to swim to
a visible platform to determine whether non-cognitive impairment
had occurred (e.g. visual or swimming deficits). In the MWM test,
rats received four training trials daily for four consecutive
days.
Statistical analysis
Data are expressed as the mean ± standard error and
were analyzed using SPSS software version 20 for Windows (IBM SPSS,
Armonk, NY, USA). Body weight, average swimming speed and escape
latency of pups were analyzed by two-way repeated-measures analysis
of variance (ANOVA), with Bonferroni post-hoc analysis. The
remaining data were analyzed with one-way ANOVA, followed by
Bonferroni post-hoc analysis. P<0.05 was considered to indicate
a statistically significant difference.
Results
Physiological parameters following
propofol exposure
There were no significant differences in arterial
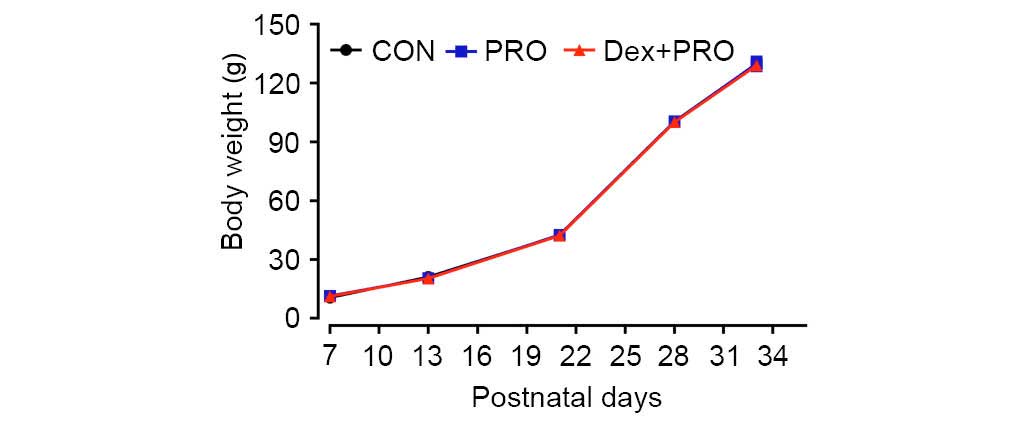
blood gas values, blood glucose concentrations (Table I) or body weights (Fig. 1) among the three treatment groups.
These results reduce the possibility that the anesthesia protocol
of the current study caused hypoxia and hypercapnia, which may
result in neuronal apoptosis, hypoglycemia and delayed puberty.
 | Table IDex preconditioning and PRO
administration have no effect on arterial blood gas values of
neonatal rats. |
Table I
Dex preconditioning and PRO
administration have no effect on arterial blood gas values of
neonatal rats.
| Group | pH | PO2
(mmHg) | PCO2
(mmHg) | Lactate
(mmol/l) | Glucose
(mmol/l) |
|---|
| CON | 7.408±0.177 | 88.6±1.75 | 40.2±1.43 | 2.96±0.27 | 6.0±0.71 |
| PRO | 7.398±0.171 | 89.8±1.80 | 40.8±1.46 | 3.02±0.11 | 6.6±0.75 |
| Dex+PRO | 7.396±0.221 | 90.0±1.95 | 39.6±1.08 | 3.00±0.15 | 5.8±0.97 |
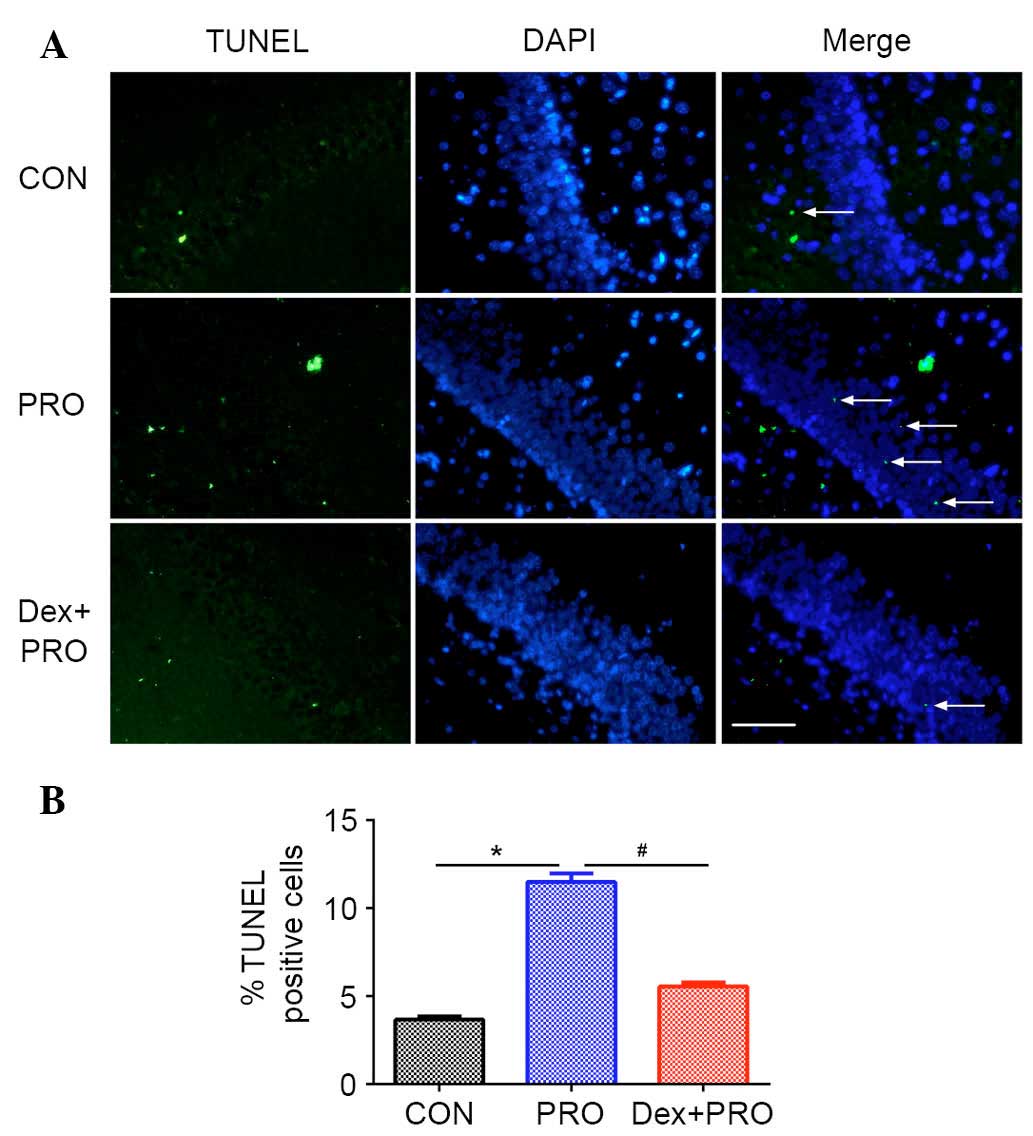
Dex preconditioning attenuates
propofol-induced hippocampal apoptosis
The number of TUNEL-positive cells per
mm2 in the hippocampal CA1 region was higher in the
propofol-treated animals compared with control animals (P=0.0007;
Fig. 2). Pretreatment with 75
µg/kg Dex 20 min prior to propofol exposure significantly
reduced the number of TUNEL-positive cells in the CA1 region
compared with the PRO group (P=0.0005; Fig. 2).
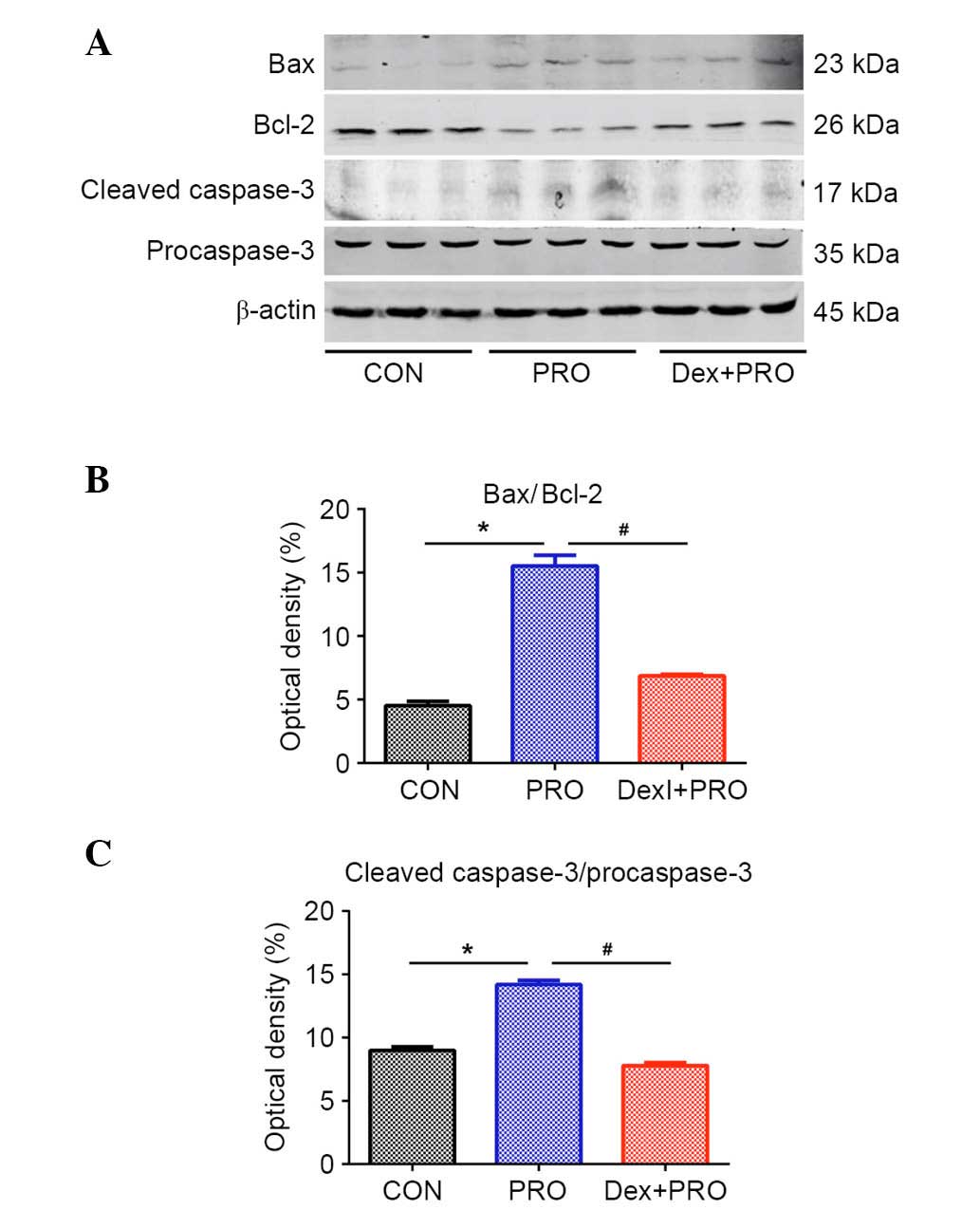
Dex preconditioning regulates the ratio
of Bax/Bcl-2 and caspase-3 activation
Western blot analysis (Fig. 3A) demonstrated that, compared with
controls, rats subjected to repeated propofol exposure exhibited a
significantly higher ratio of Bax/Bcl-2 (P=0.0006; Fig. 3B) and caspase-3 activation
(P=0.0008; Fig. 3C). However,
compared with the PRO group, Dex pretreatment alleviated the change
in Bax/Bcl-2 ratio and caspase-3 activation induced by propofol
(P=0.0009).
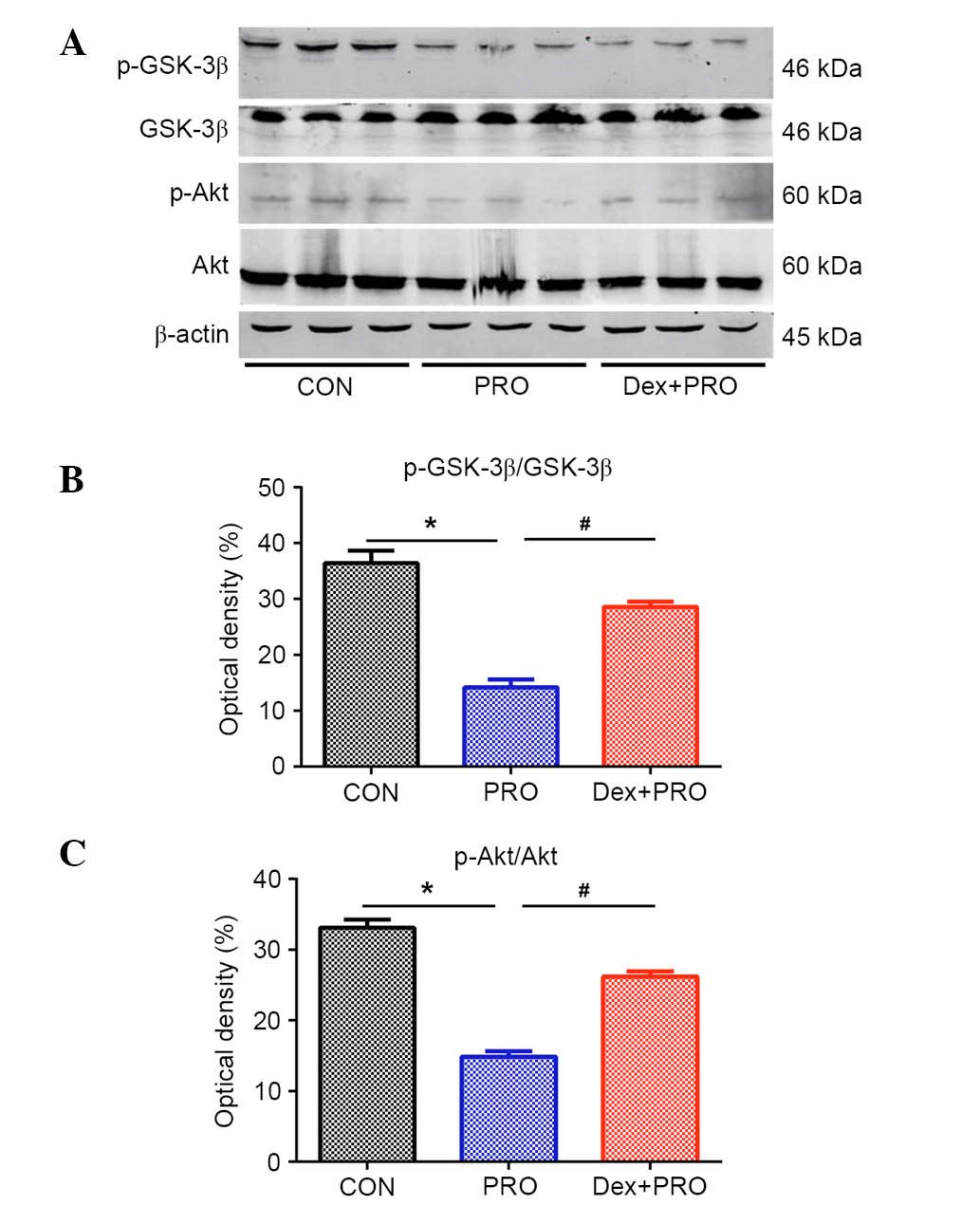
Dex preconditioning activates the
PI3K/Akt/GSK-3β signaling pathway
PRO group rats exhibited a significantly lower ratio
of p-Akt/Akt (P= 0.03) and p-GSK-3β/GSK-3β (P=0.04) compared with
controls. However, compared with the PRO group, the ratios were
significantly reduced by Dex pretreatment (P=0.0008; Fig. 4).
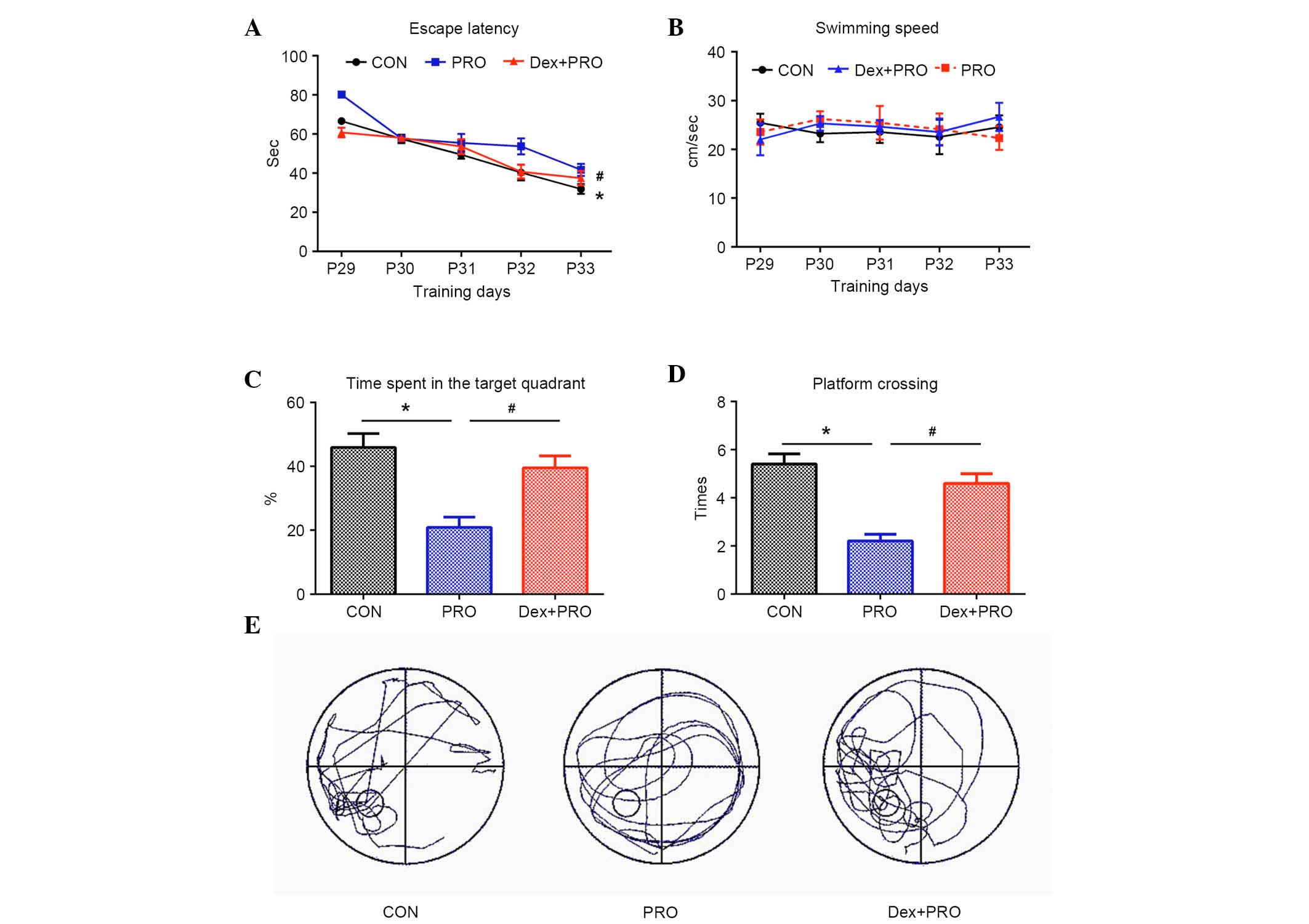
Dex preconditioning attenuates
propofol-induced cognitive deficits
During the MWM test, the escape latency to locate
the submerged platform was significantly higher in the PRO group
rats compared with controls (P=0.0005) and the Dex + PRO group
(P=0.0007) at P33 (Fig. 5A), and
there were no significant differences in swimming speeds among the
three groups (Fig. 5B). In the
probe test, compared with controls, the time spent on the platform
area (P=0.0009; Fig. 5C) and the
number of platform crossings (P=0.0006; Fig. 5D) were significantly reduced in the
PRO group rats. Furthermore, Dex significantly increased the time
spent on the platform area and platform crossing compared with the
PRO group rats (P=0.02).
Discussion
The present study demonstrated that repeated
propofol-induced juvenile spatial learning/memory impairments in
neonatal rats were ameliorated by pretreatment with 75 µg/kg
Dex 20 min prior to challenge. Dex pretreatment alleviated
hippocampal apoptosis, regulated the imbalance of Bax/Bcl-2
expression and caspase-3 activation, and normalized the
phosphorylation of Akt and GSK-3β. These results strongly suggested
that Dex preconditioning protects the developing brain against
propofol-induced neurotoxicity and hippocampal apoptosis, a process
that may be mediated by the PI3K/Akt/GSK-3β signaling pathway.
Almost all general anesthetics have dual effects of
neuroprotection and neurotoxicity, depending on the concentration,
duration and the age of exposure (22,23).
During early postnatal life, which is termed the brain growth-spurt
period, neuronal proliferation, migration, differentiation and
synapse formation occur. Increasing experimental data in
pre-clinical settings suggest that, during this period, brains are
more vulnerable to anesthetics (6,8). The
current study used P7 rats in the experimental model, as during
this time neurogenesis is still occurring, thus neurons have a high
degree of plasticity and are vulnerable to internal and external
environment.
As one of the most widely used general anesthetics,
propofol has previously been demonstrated to induce neuronal
apoptosis, which may contribute to further memory impairment, in
several studies (8). However,
there is some controversy regarding the appropriate dose of
propofol to use in P7 rats. Gao et al (24) demonstrated that 75 mg/kg propofol
i.p. once a day at regular 24-h intervals for 7 days in rats from
P7 to P13 impairs learning and memory of pups without visible signs
of cyanosis. However, Pesić et al (25) observed that 50 mg/kg propofol
immediately induces loss of the righting reflex and promotes
cyanotic changes in skin color. In a preliminary experiment, it was
observed in our laboratory that 75 mg/kg propofol could lead to
respiratory depression and cyanosis over a long time (data not
shown). Thus, the anesthesia scheme was adjusted to the protocol
used by Bercker et al (26)
(30 mg/kg propofol every 90 min up to a cumulative dose of 90 mg/kg
every day for seven consecutive days), and no clearly discernible
anoxic events occurred.
Several protein families contribute to apoptotic
cell death regulation. Among these proteins, Bcl-2 is
anti-apoptotic and Bax is pro-apoptotic, thus, their relative
expression determines cell viability. Activation of the downstream
protein caspase-3 was previously demonstrated to be induced by
propofol in postnatal brains during maturation (25). In the present study, cognitive
impairments associated with propofol exposure were accompanied by
significant increases in the Bax/Bcl-2 ratio, caspase-3 expression
and the number of TUNEL-positive cells. These results further
suggested that apoptosis in the hippocampus is important for
propofol neurotoxicity.
Dex, a potent and highly selective α2-adrenoceptor
agonist, is commonly administered by the continuous infusion method
in pediatric patients for anesthesia and sedation, and occasionally
it is also received as a bolus. In addition to its hypnotic and
analgesic effect in several brain injury models, it has previously
been demonstrated to exhibit anti-apoptotic properties in both
pre-clinical and clinical studies (15,18).
A previous report demonstrating that the expression of Bcl-2 and
Mdm-2 proto-oncogene was greater following Dex treatment compared
with controls suggests that the neuroprotective properties of Dex
involve ultra-early modulation of the balance between pro- and
anti-apoptotic proteins (27). In
the current study, 75 µg/kg Dex was chosen as the
experimental dose based on a previous study, which demonstrated
that pretreatment with 75 µg/kg prior to anesthesia exposure
significantly reduced isoflurane-induced apoptosis, whereas Dex
alone did not induce a change (19). The addition of Dex potentially
increases the depth of anesthesia, however, the gas blood results
measured in the present study demonstrated no evident physiological
parameter changes. The results of the current study are similar to
the report by Sanders et al (16), which demonstrated that, following
Dex preconditioning, the number of TUNEL-positive cells were
reduced, the expression of apoptotic markers (Bax/Bcl-2, cleaved
caspase-3/caspase-3) was reduced and spatial learning/memory was
improved. These results demonstrate that Dex exerts an important
anti-apoptotic effect against propofol-induced injury.
However, the underlying mechanism of the effects of
Dex in the central nervous system remains to be clearly delineated.
It is suggested that Dex provides its neuroprotective effect via
the α2-adrenergic receptor subtype (28). However, as the α2-adrenoceptor
antagonist, atipamezole, only partially reverses the
neuroprotective effects of Dex following isoflurane-induced
neurotoxicity in rats (16), other
mechanisms may be involved.
It is recognized that the PI3K/Akt/GSK-3β signaling
pathway also participates in the protective effect of Dex against
ischemia-reperfusion injury in the brain (29). Akt is a serine/threonine kinase and
is activated by phosphorylation under normal physiological
conditions. Activated Akt (p-Akt, Ser473) then
phosphorylates GSK-3β at Ser9 to decrease the activity
of GSK-3β. Activation of Akt is involved in anti-apoptotic
signaling via phosphorylation of Bcl-2 associated agonist of cell
death, an important member of the Bcl-2 family that can inhibit the
inactivation of Bcl-2 (30).
GSK-3β, a homologous mammalian isoform of GSK-3, is highly
expressed in the brain, particularly in the hippocampus, striatum
and thalamus (31). It
phosphorylates a wide variety of cellular substrates and thereby
regulates various cellular processes, ranging from glycogen
metabolism to cell survival and neuronal polarity (32). Phosphorylation of GSK-3β at
Ser9 negatively regulates pro-apoptotic activity
(14), and has been previously
demonstrated to improve long-term memory in hippocampal-associated
tasks (13). In the current study,
the expression of p-GSK-3β at Ser9 and p-Akt at
Ser473 were downregulated by propofol, which was
accompanied by an increase in apoptosis markers (Bax) and spatial
deficits, and these changes were ameliorated by Dex, which is
similar to the observations of Linseman et al (33). These results suggest that Dex can
ameliorate propofol-induced disturbances to the PI3K/Akt/GSK-3β
signaling pathway in the developing brain, alleviate hippocampal
apoptosis and inhibit learning/memory deficits. However, the exact
effect of Dex on the PI3K/Akt/GSK-3β signaling pathway requires
further investigation.
Disruption of intracellular calcium homeostasis,
particularly excessive calcium release from intracellular stores,
mediates general anesthetic-induced neuronal apoptosis (34,35).
Furthermore, calcium is also a regulator of PI3K/Akt/GSK-3β
signaling (36), and Dex has been
previously demonstrated to affect calcium release from
intracellular stores in astrocytes (37). Thus, calcium release may act as an
inhibitor of the PI3K/Akt/GSK-3β pathway and an inducer of
apoptosis, which may be associated with the neuroprotective effect
of Dex on propofol-induced neurotoxicity. However, this issue
requires further investigation.
There are several limitations of the current study.
Notably, a ʻDex aloneʼ group was not included in the study.
Although a previous study has already shown that one dose of 75
µg/kg Dex alone does not increase neuronal apoptosis
compared with controls (38), it
would be beneficial to include a Dex alone group in future studies
to further confirm these findings. Additionally, the effect of Dex
on long-term hippocampal-dependent memory was not determined.
Furthermore, although there are reports of propofol infusions being
used for five days or more in pediatric intensive care units
(ICUs), repeated administration of propofol for seven consecutive
days in the present study is highly unlikely to occur in pediatric
anesthesia. This dosing protocol was designed to simulate repeated
propofol infusion in a pediatric ICU, and whether this dose of
propofol (30 mg/kg, i.p.) in rodents is relevant to clinical
practice is difficult to predict. However, the results of the
current study provide a novel basis for exploring the activity of
Dex in neurogenesis following propofol anesthesia in neonate
brains. Further studies investigating this issue are required.
In conclusion, the in vivo data of the
present study suggests that repeated exposure to propofol
anesthesia impairs juvenile spatial learning and memory in neonatal
rats, and this impairment may be associated with hippocampal
apoptosis. Dex administration prior to propofol anesthesia improves
cognitive dysfunction, partially by attenuating hippocampal
apoptosis and regulating the PI3K/Akt/GSK-3β signaling pathway.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant no. 28342016).
References
|
1
|
Sun LS, Li G, DiMaggio CJ, Byrne MW, Ing
C, Miller TL, Bellinger DC, Han S and McGowan FX: Feasibility and
pilot study of the Pediatric Anesthesia NeuroDevelopment Assessment
(PANDA) project. J Neurosurg Anesthesiol. 24:382–388. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu C, Gao J, Karlsson N, Li Q, Zhang Y,
Huang Z, Li H, Kuhn HG and Blomgren K: Isoflurane anesthesia
induced persistent, progressive memory impairment, caused a loss of
neural stem cells, and reduced neurogenesis in young, but not
adult, rodents. J Cereb Blood Flow Metab. 30:1017–1030. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sun L: Early childhood general anaesthesia
exposure and neurocognitive development. Br J Anaesth. 105(Suppl
1): pp. i61–i68. 2010, View Article : Google Scholar
|
|
4
|
Wilder RT, Flick RP, Sprung J, Katusic SK,
Barbaresi WJ, Mickelson C, Gleich SJ, Schroeder DR, Weaver AL and
Warner DO: Early exposure to anesthesia and learning disabilities
in a population-based birth cohort. Anesthesiology. 110:796–804.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kay B and Rolly G: I.C.I. 35868, a new
intravenous induction agent. Acta Anaesthesiol Belg. 28:303–316.
1977.PubMed/NCBI
|
|
6
|
Harman F, Hasturk AE, Yaman M, Arca T,
Kilinc K, Sargon MF and Kaptanoglu E: Neuroprotective effects of
propofol, thiopental, etomidate, and midazolam in fetal rat brain
in ischemia-reperfusion model. Childs Nerv Syst. 28:1055–1062.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rossaint J, Rossaint R, Weis J, Fries M,
Rex S and Coburn M: Propofol: Neuroprotection in an in vitro model
of traumatic brain injury. Crit Care. 13:R612009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Creeley C, Dikranian K, Dissen G, Martin
L, Olney J and Brambrink A: Propofol-induced apoptosis of neurones
and oligodendrocytes in fetal and neonatal rhesus macaque brain. Br
J Anaesth. 110(Suppl 1): i29–i38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Krzisch M, Sultan S, Sandell J, Demeter K,
Vutskits L and Toni N: Propofol anesthesia impairs the maturation
and survival of adult-born hippocampal neurons. Anesthesiology.
118:602–610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sanchez V, Feinstein SD, Lunardi N,
Joksovic PM, Boscolo A, Todorovic SM and Jevtovic-Todorovic V:
General anesthesia causes long-term impairment of mitochondrial
morphogenesis and synaptic transmission in developing rat brain.
Anesthesiology. 115:992–1002. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gray JJ, Bickler PE, Fahlman CS, Zhan X
and Schuyler JA: Isoflurane neuroprotection in hypoxic hippocampal
slice cultures involves increases in intracellular Ca2+ and
mitogen-activated protein kinases. Anesthesiology. 102:606–615.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dewachter I, Ris L, Jaworski T, Seymour
CM, Kremer A, Borghgraef P, De Vijver H, Godaux E and Van Leuven F:
GSK3beta, a centre-staged kinase in neuropsychiatric disorders,
modulates long term memory by inhibitory phosphorylation at
serine-9. Neurobiol Dis. 35:193–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsing CH, Chen YH, Chen CL, Huang WC, Lin
MC, Tseng PC, Wang CY, Tsai CC, Choi PC and Lin CF: Anesthetic
propofol causes glycogen synthase kinase-3β-regulated
lysosomal/mitochondrial apoptosis in macrophages. Anesthesiology.
116:868–881. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Duan X, Li Y, Zhou C, Huang L and Dong Z:
Dexmedetomidine provides neuroprotection: Impact on
ketamine-induced neuroapoptosis in the developing rat brain. Acta
Anaesthesiol Scand. 58:1121–1126. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sanders RD, Xu J, Shu Y, Januszewski A,
Halder S, Fidalgo A, Sun P, Hossain M, Ma D and Maze M:
Dexmedetomidine attenuates isoflurane-induced neurocognitive
impairment in neonatal rats. Anesthesiology. 110:1077–1085. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z, Kou D, Li Z, He Y, Yu W and Du H:
Effects of propofol-dexmedetomidine combination on ischemia
reperfusion-induced cerebral injury. Neuro Rehabilitation.
35:825–834. 2014.PubMed/NCBI
|
|
18
|
Degos V, Charpentier TL, Chhor V, Brissaud
O, Lebon S, Schwendimann L, Bednareck N, Passemard S, Mantz J and
Gressens P: Neuroprotective effects of dexmedetomidine against
glutamate agonist-induced neuronal cell death are related to
increased astrocyte brain-derived neurotrophic factor expression.
Anesthesiology. 118:1123–1132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Y, Zeng M, Chen W, Liu C, Wang F, Han
X, Zuo Z and Peng S: Dexmedetomidine reduces isoflurane-induced
neuroapoptosis partly by preserving PI3K/Akt pathway in the
hippocampus of neonatal rats. PLoS One. 9:e936392014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li ZQ, Rong XY, Liu YJ, Ni C, Tian XS, Mo
N, Chui DH and Guo XY: Activation of the canonical nuclear
factor-κB pathway is involved in isoflurane-induced hippocampal
interleukin-1β elevation and the resultant cognitive deficits in
aged rats. Biochem Biophys Res Commun. 438:628–634. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liang Y, Li Z, Mo N, Li M, Zhuang Z, Wang
J, Wang Y and Guo X: Isoflurane preconditioning ameliorates renal
ischemia-reperfusion injury through antiinflammatory and
anti-apoptotic actions in rats. Biol Pharm Bull. 37:1599–1605.
2014. View Article : Google Scholar
|
|
22
|
Kawaguchi M, Furuya H and Patel PM:
Neuroprotective effects of anesthetic agents. J Anesth. 19:150–156.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou Z and Ma D: Anaesthetics-induced
neurotoxicity in developing brain: An update on preclinical
evidence. Brain Sci. 4:136–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao J, Peng S, Xiang S, Huang J and Chen
P: Repeated exposure to propofol impairs spatial learning, inhibits
LTP and reduces CaMKIIα in young rats. Neurosci Lett. 560:62–66.
2014. View Article : Google Scholar
|
|
25
|
Pesić V, Milanović D, Tanić N, Popić J,
Kanazir S, Jevtović-Todorović V and Ruzdijić S: Potential mechanism
of cell death in the developing rat brain induced by propofol
anesthesia. Int J Dev Neurosci. 27:279–287. 2009. View Article : Google Scholar
|
|
26
|
Bercker S, Bert B, Bittigau P,
Felderhoff-Müser U, Bührer C, Ikonomidou C, Weise M, Kaisers UX and
Kerner T: Neurode-generation in newborn rats following propofol and
sevoflurane anesthesia. Neurotox Res. 16:140–147. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Engelhard K, Werner C, Eberspächer E,
Bachl M, Blobner M, Hildt E, Hutzler P and Kochs E: The effect of
the alpha 2-agonist dexmedetomidine and the N-methyl-D-aspartate
antagonist S (+)-ketamine on the expression of apoptosis-regulating
proteins after incomplete cerebral ischemia and reperfusion in
rats. Anesth Analg. 96:524–531. 2003.
|
|
28
|
Ma D, Hossain M, Rajakumaraswamy N, Arshad
M, Sanders RD, Franks NP and Maze M: Dexmedetomidine produces its
neuroprotective effect via the alpha 2A-adrenoceptor subtype. Eur J
Pharmacol. 502:87–97. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu YM, Wang CC, Chen L, Qian LB, Ma LL,
Yu J, Zhu MH, Wen CY, Yu LN and Yan M: Both PI3K/Akt and ERK1/2
pathways participate in the protection by dexmedetomidine against
transient focal cerebral ischemia/reperfusion injury in rats. Brain
Res. 1494:1–8. 2013. View Article : Google Scholar
|
|
30
|
Wang H, Luo QF, Peng AF, Long XH, Wang TF,
Liu ZL, Zhang GM, Zhou RP, Gao S, Zhou Y and Chen WZ: Positive
feedback regulation between Akt phosphorylation and fatty acid
synthase expression in osteosarcoma. Int J Mol Med. 33:633–639.
2014.
|
|
31
|
Leroy K and Brion JP: Developmental
expression and localization of glycogen synthase kinase-3beta in
rat brain. J Chem Neuroanat. 16:279–293. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yuskaitis CJ and Jope RS: Glycogen
synthase kinase-3 regulates microglial migration, inflammation and
inflammation-induced neurotoxicity. Cell Signal. 21:264–273. 2009.
View Article : Google Scholar
|
|
33
|
Linseman DA, Butts BD, Precht TA, Phelps
RA, Le SS, Laessig TA, Bouchard RJ, Florez-McClure ML and
Heidenreich KA: Glycogen synthase kinase-3beta phosphorylates Bax
and promotes its mitochondrial localization during neuronal
apoptosis. J Neurosci. 24:9993–10002. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao YL, Xiang Q, Shi QY, Li SY, Tan L,
Wang JT, Jin XG and Luo AL: GABAergic excitotoxicity injury of the
immature hippocampal pyramidal neurons' exposure to isoflurane.
Anesth Analg. 113:1152–1160. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sinner B, Friedrich O, Zink W, Zausig Y
and Graf BM: The toxic effects of s (+)-ketamine on differentiating
neurons in vitro as a consequence of suppressed neuronal Ca2+
oscillations. Anesth Analg. 113:1161–1169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei H and Inan S: Dual effects of
neuroprotection and neurotoxicity by general anesthetics: Role of
intracellular calcium homeostasis. Prog Neuropsychopharmacol Biol
Psychiatry. 47:156–161. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen Y, Zhao Z, Code WE and Hertz L: A
correlation between dexmedetomidine-induced biphasic increases in
free cytosolic calcium concentration and energy metabolism in
astrocytes. Anesth Analg. 91:353–357. 2000.PubMed/NCBI
|
|
38
|
Liao Z, Cao D, Han X, Liu C, Peng J, Zuo
Z, Wang F and Li Y: Both JNK and P38 MAPK pathways participate in
the protection by dexmedetomidine against isoflurane-induced
neuroapoptosis in the hippocampus of neonatal rats. Brain Res Bull.
107:69–78. 2014. View Article : Google Scholar : PubMed/NCBI
|