Introduction
Infertility affects ~15% of couples worldwide, with
half of cases reported to be due to the male partner (1). Treatments for male infertility have
not been developed as the associated molecular mechanisms are not
well understood (2). A more
detailed investigation of the physiological mechanisms of
spermatogenesis is required for an improved understanding of
infertility prior to the development of therapies for this
condition.
Spermatogenesis is a complicated developmental
process. Spermatogonia differentiate into spermatocytes and
spermatids during two meiotic divisions, which leads to the
production of mature sperm (3).
Defects at any stage of this process may lead to infertility. The
process involves various genes, which encode proteins that in turn
are important for specific stages of germ cell development. Theses
genes are primarily expressed in spermatogenic cells, and are
regulated at the transcriptional or post-transcriptional level. The
identification of germ cell-specific or testes-specific genes
involved in these unique events provides a means by which to
dissect the differentiation program and to study the mechanisms of
spermatogenesis. Previous studies have identified a number of
testes-specific genes in humans and mice, including fibronectin
type 3 and ankyrin repeat domains 1 (4), A kinase (PRKA) anchor protein 3
(5), protein lifeguard 5 (6), protease, serine 41 (7), spermatogenesis associated 33
(2) and testis-specific serine
kinase 4 (8). In addition to the
aforementioned genes it is possible that additional testis-specific
genes exist and provide unique functions.
Coiled-coil domain containing 38 (Ccdc38) was
selected from the expressed sequence tags (ESTs) obtained through
the comparison of testes gene libraries with the libraries of other
tissues and cell lines using the differential digital display
program (9). The EST profile of
Ccdc38 in Unigene (Mm.477086) indicates that the
Ccdc38 transcript is detected solely in murine testes, which
is consistent with the report at BioGPS.org
(10). Ccdc38 orthologs are
present in other species, including Rattus norvegicus (Gene
ID: 500823), Bos taurus (Gene ID: 517752), Pan
troglodytes (Gene ID: 738083) and humans (Gene ID: 120935).
The present study determined that the
testes-specific gene, Ccdc38, was only expressed in mouse
testes. This gene is primarily expressed in spermatogonia and
spermatocytes. Additionally, it was identified that CCDC38 may
interact with ubiquitinated histone H2A (uH2A) in mouse testes.
This suggests that Ccdc38 is a testes-specific gene, which
is important for spermatogenesis in mice.
Materials and methods
Samples
Twenty male and twenty female BALB/c adult mice
(25–30 g) were purchased from the Laboratory Animals Center of
South Medical University (Guangzhou, China). Mice were treated
under a pathogen-free condition at ~22°C under a 12 h light/dark
cycle. All the animals had free access to standard water and chow.
Male and female mice were allowed to mate spontaneously, and the
day of birth was assigned as day 1. Testes tissues were
individually collected from the 1, 2, 3, 4, 6, 8 weeks, and 6
months-old mice following sacrifice by cervical dislocation. The
remaining organs, including the heart, brain, lungs, spleen,
kidneys, liver, epididymis and bladder from the adult offspring
were immediately frozen in RNAlater (Qiagen, Inc., Valencia, CA,
USA). The protocol was approved by The Ethics Committee of Peking
University Shenzhen Hospital (Shenzhen, China).
Antibodies
The rabbit polyclonal anti-CCDC38 antibody (cat. no.
ab170231), the mouse monoclonal anti-glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) antibody (cat. no. ab8245), the mouse
monoclonal anti-enhanced green fluorescent protein (EGFP) antibody
(cat. no. ab184601) and the rabbit polyclonal anti-hemagglutinin
(HA) tag antibody (cat. no. ab137838) were purchased from Abcam
(Cambridge, UK). The mouse anti-H2A antibody was purchased from EMD
Millipore (Billerica, MA, USA). Anti-rabbit-Alexa Fluor 488 (cat.
no. ab150073) and anti-mouse-Cy3 (cat. no. ab97035) were purchased
from Abcam.
Construction of plasmids and cell
culture
The full length of the Ccdc38 cDNA was
amplified by polymerase chain reaction (PCR) with the forward (F)
5′-ATGGCATCCCAGATGC-3′ and the reverse (R)
5′-ACTAAAAAAGTACTCTTCGTC-3′ primers, and then inserted into
pCDNA3.1/HA plasmids via BamH1 and Xhol. The full
length of the H2A cDNA was amplified by PCR with the F
5′-ATGTCTGGACGTGGCAAACAG-3′ and the R 5′-TTATTTCCCCTTGGCCTTGTGG-3′
primers and then inserted into pEGFP-C1 plasmids via BamH1
and EcoR1. Reaction condirions were as follows: 98°C for 2
min; 37 cycles of 98°C for 10 sec, 55°C for 30 sec and 72°C for 2
min; followed by 72°C for 5 min. The PCR products were cloned and
sequenced by Invitrogen; Thermo Fisher Scientific, Inc.. Two cell
lines, TM4 and HEK293T were obtained from the American Type Culture
Collection (Manassas, VA, USA). The cells were maintained in
Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Thermo Fisher Scientific,
Inc.) at 37°C in a humidified atmosphere with 5%
CO2.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from mice tissues using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) in accordance
with the manufacturer's protocol. Total RNA (1 µg) was used
as a template for the reverse transcription using the PrimeScript
RT Enzyme Mix I (Takara Bio, Inc., Shiga, Japan). Reverse and
forward oligonucleotide primers were designed using Primer Express
version 2.0 software (Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The primer sequences were as follows:
Ccdc38, F 5′-CTTGTCCTGTTAGTCCTGTATAG-3′, R
5′-CGTAGAGATGAAGTGTGATGAT-3′; and Gapdh (as an internal
control) F 5′-AGTGGCAAAGTGGAGATT-3′, R 5′-GTGGAGTCATACTGGAACA-3′.
The following PCR conditions were used: 98°C for 2 min; 32 cycles
of 98°C for 10 sec, 55°C for 30 sec and 72°C for 30 sec; and 72°C
for 7 min using a LightCycler 480II instrument (Roche Diagnostics
Deutschland GmbH, Mannheim, Germany). Testes samples obtained at
different time points and other tissues were amplified in
triplicate. The amplified products were resolved on a 2% agarose
gel, stained with ethidium bromide (Sigma-Aldrich, St. Louis, MO,
USA) and visualized with an ultraviolet imaging system (Universal
Hood II, Bio-Rad Laboratories, Hercules, CA, USA). Data were
normalized to the expression of Gapdh and were quantified
according to the 2−ΔΔCq method (11).
Western blotting
The protein extracts of various mouse tissues (20
µg) were subjected to 12% sodium dodecyl sulfate
polyacrylamide gel and transferred to polyvinylidene fluoride
membrane (EMD Millipore). Following blocking with 5% non-fat milk,
the membranes were incubated with rabbit anti-CCDC38 (1:500, cat.
no. ab170231, Abcam) or anti-GAPDH antibody (1:5,000, cat. no.
ab181602, Abcam) antibodies overnight at 4°C. Subsequently, the
membranes were washed three times with Tris-buffered saline with
Tween-20 buffer for 15 min. Then, the membranes were incubated with
horseradish peroxidase (HRP)-conjugated goat anti-rabbit secondary
antibody (1:2,000, cat. no. ab97051, Abcam, CA, USA) for 1 h at
room temperature. Finally, the positive bands were examined using
an enhanced chemiluminescence kit (Thermo Fisher Scientific, Inc.).
The densitometry of each band was analyzed using Image-Pro Plus 6.0
software (Image Pro-Plus 6.0; Media Cybernetics, Silver Spring, MD,
USA). The CCDC38 expression level in tested from mice aged 1–8
weeks and 6 months was normalized to GAPDH expression.
Immunohistochemistry and
immunofluorescence
Paraffin sections (3–5 µm) of testes samples
from 1–8 weeks and 6 months-old mice was used for staining. were
prepared as previously described (12). The sections were blocked in 10%
goat serum and then incubated with rabbit anti-CCDC38 antibody
(dilution, 1:300), rabbit anti-uH2A (dilution, 1:100) antibody
overnight at 4°C. The sections were washed with phosphate-buffered
saline (PBS) three times and incubated for 1 h at 37°C with the
anti-rabbit-Alexa Fluor 488 (1:500) and anti-mouse-Cy3 (1:500)
secondary antibodies. Subsequently, the slides were incubated with
50 µl of 1X DAPI solution for 5 min at room temperature in
the dark, followed by washing with 100 µl PBS three times.
The slides were then treated with DAPI for 5 min at room
temperature, washed in PBS, mounted, and observed under ×100 LSM
710 oil lens (Zeiss GmbH, Jena, Germany). DAB staining was
performed according to the manufacturer's recommended protocol (ABC
kit; Maixin Biotechnology, Fuzhou, China). The DAB slides were
observed under ×40 Leica DM4000B lens (Leica Micro-systems GmbH,
Wetzlar, Germany). The level of nonspecific staining was determined
by omission of the incubation step with the primary antibody.
Hoeschst staining
The Hoechst 33258 staining kit (Invitrogen; Thermo
Fisher Scientific, Inc.) was utilized to observe cell nuclear
staining in mouse testes. Paraffin sections were prepared as
described previously (12).
Following culture with primary and secondary antibodies, the slides
were incubated with 50 µl of 1X Hoechst 33258 solution for 5
min at room temperature in the dark, followed by washing with 100
µl PBS three times. The results were observed under a LSM
710 oil lens at ×100 magnification (Zeiss GmbH, Jena, Germany).
Co-immunoprecipitation (co-IP) assay
Whole testes were prepared with lysis buffer (10 mM
Tris pH 7.4, 1.0% Triton X-100, 0.5% NP-40, 150 mM NaCl, 20 mM NaF,
1 mM EDTA, 1 mM EGTA, and 0.2 mM PMSF) supplemented with protease
inhibitors. For the in vivo co-IP assay, the supernatant was
incubated with anti-CCDC38 (10 µg) and anti-uH2A (5
µg) antibodies overnight at 4°C. Protein A/G beads (60
µl) were then added to each sample, and the mixtures were
incubated at 4°C for 1 h. For the in vitro co-IP assay, the
pEGFP-H2A overexpression plasmid was synthesized by Invitrogen;
Thermo Fisher Scientific, Inc. The CCDC38-pCDNA3.1/HA (Guang-dong
and Shenzhen Key Laboratory of Male Reproductive Medicine and
Genetics; Shenzhen, China) was transfected into HEK293T cells, with
pEGFP-H2A (Guangdong and Shenzhen Key Laboratory of Male
Reproductive Medicine and Genetics) or the vector control
(pEGFP-C1; Invitrogen, Thermo Fisher Scientific, Inc.) using
Lipofectamine 2000 transfection reagent (Invitrogen, Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
cells were harvested 48 h after transfection and extracted with the
aforementioned lysis buffer. Subsequently, the supernatant was
incubated with anti-EGFP (5 µg) and anti-HA (2 µg)
antibodies overnight at 4°C. Protein A/G beads (60 µl) were
also added to every sample, and the mixtures were incubated at 4°C
for 1 h. The beads were then washed three times with lysis buffer,
boiled in sample buffer containing 0.2 M dithiothreitol, and
analyzed by western blotting as previously described.
Bioinformatic analysis
Bioinformatics related to the genome, chromosome,
mRNA and protein products of Ccdc38 were obtained using the
National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/), Mouse Genome
Informatics (http://www.informatics.jax.org/) and the UCSC Genome
bioinformatics (http://genome.ucsc.edu/). The SIB Bioinformatics
Resource Portal (http://www.expasy.org/) was applied to analyze the
domains and motifs of CCDC38 protein.
Statistical analysis
Each of the experiments was repeated at least three
times. Data are presented as the mean ± standard deviation.
Student's t-test was used to compare the difference between two
groups. SPSS version 17.0 statistical software (SPSS, Inc.,
Chicago, IL, USA) was used for statistical analyses. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of Ccdc38 by in silico
screening
The testes-specific gene, Ccdc38 was
identified using the UniGene libraries. To further characterize the
expression of Ccdc38, its structure and function was
analyzed using systematic bioinformatic methods. The Ccdc38
gene encodes a protein with a predicted molecular weight of ~65
kDa. The homology with other vertebrates in GenBank was also
determined, indicating that several predicted homologues only exist
in mammals (Fig. 1A), with mouse
CCDC38 sharing a high sequence homology with other mammalian
homologues. The CCDC38 protein also had three coiled-coil domains
in the Pfam database. Sequence analysis indicated that CCDC38 is a
serine-rich protein. Two potential sites (12 serine phosphorylation
sites and 1 threonine phosphorylation site) were revealed by
post-translational modification analysis (Fig. 1B), which suggests that this protein
may be important for kinase signaling during spermatogenesis.
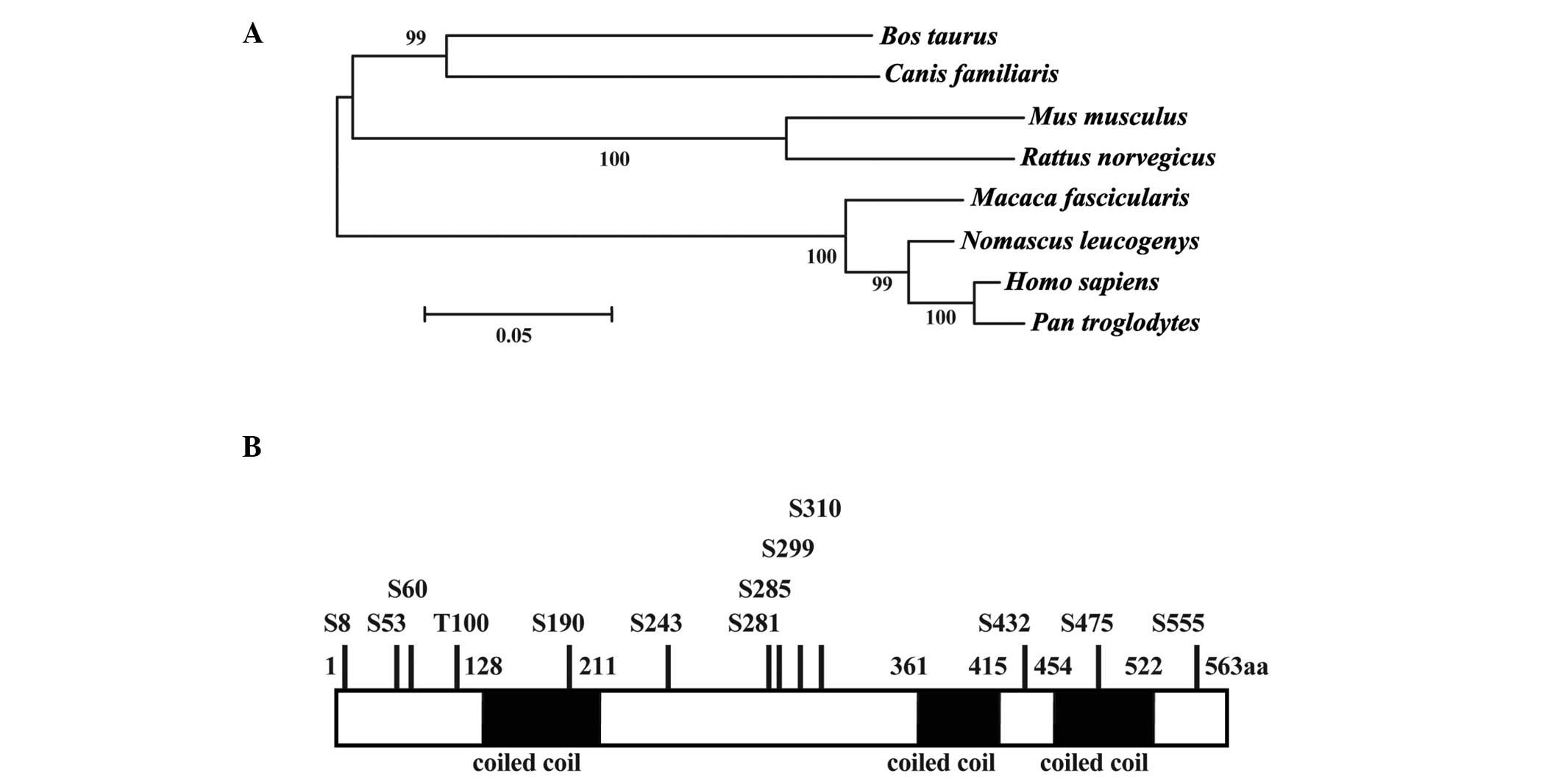 | Figure 1Phylogenetic tree, domains, and
modification sites of CCDC38. (A) Phylogenetic tree of CCDC38 in
mammals. Phylogenetic analysis was performed with MEGA5. Numbers on
the branches represent the bootstrap values from 1,000 replicates
obtained using the Neighbor-Joining method. The scale bar
corresponds to the estimated evolutionary distance units. GenBank
accession numbers are as follows: Bos taurus,
XP_003586123.2; Canis familiaris, XP_854813.2; Mus
musculus, XP_006513691.1; Rattus norvegicus,
XP_006241333.1; Macaca fascicularis, XP_005571963.1;
Nomascus leucogenys, XP_003259733.1; Homo sapiens,
XP_006719292.1; Pan troglodytes, XP_001144908.1. (B)
Schematic mapping of potential protein domains and
post-translational modification sites. The predicted sites for
phosphorylation in CCDC38 are indicated. CCDC38, coiled-coil domain
containing 38; S/T, Serine/Threonine phosphorylation-sites. |
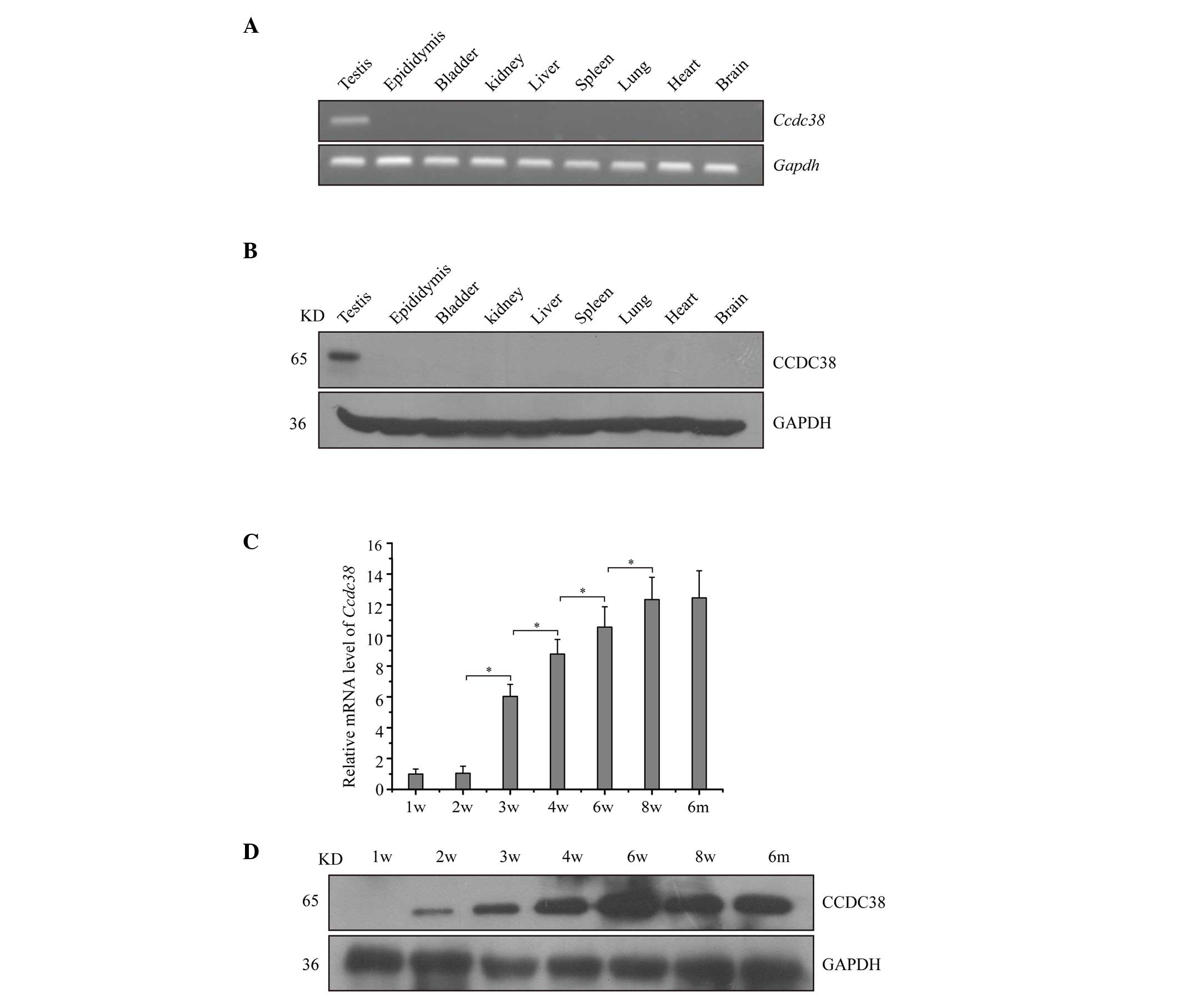
Expression of CCDC38 in mice
The mRNA expression of Ccdc38 in various
adult mouse tissues was determined using RT-PCR and RT-qPCR.
Ccdc38 was exclusively expressed in testes (Fig. 2A). To determine its protein level,
a polyclonal anti-CCDC38 antibody was used against the coding
region of CCDC38 in western blotting. The analysis indicated a
unique band at 65 kDa in mouse testes, consistent with the
predicted molecular weight by Computing pI/Mw (13). The protein level of CCDC38 was
consistent with the mRNA analysis displaying a testis-specific
expression pattern (Fig. 2B).
Additionally, the timing of mouse CCDC38 expression for mRNA and
protein levels during postnatal testis development was
investigated. As presented in Fig.
2C, The expression of Ccdc38 mRNA from 2 to 8 weeks was
gradually increased, as compared with that at 1 week. Western blot
analysis indicated that the CCDC38 protein was initially expressed
at 2 weeks and the expression pattern was consistent with its mRNA
expression (Fig. 2D). This
indicates that CCDC38 is testes-specific in mice and is
developmentally regulated during spermatogenesis.
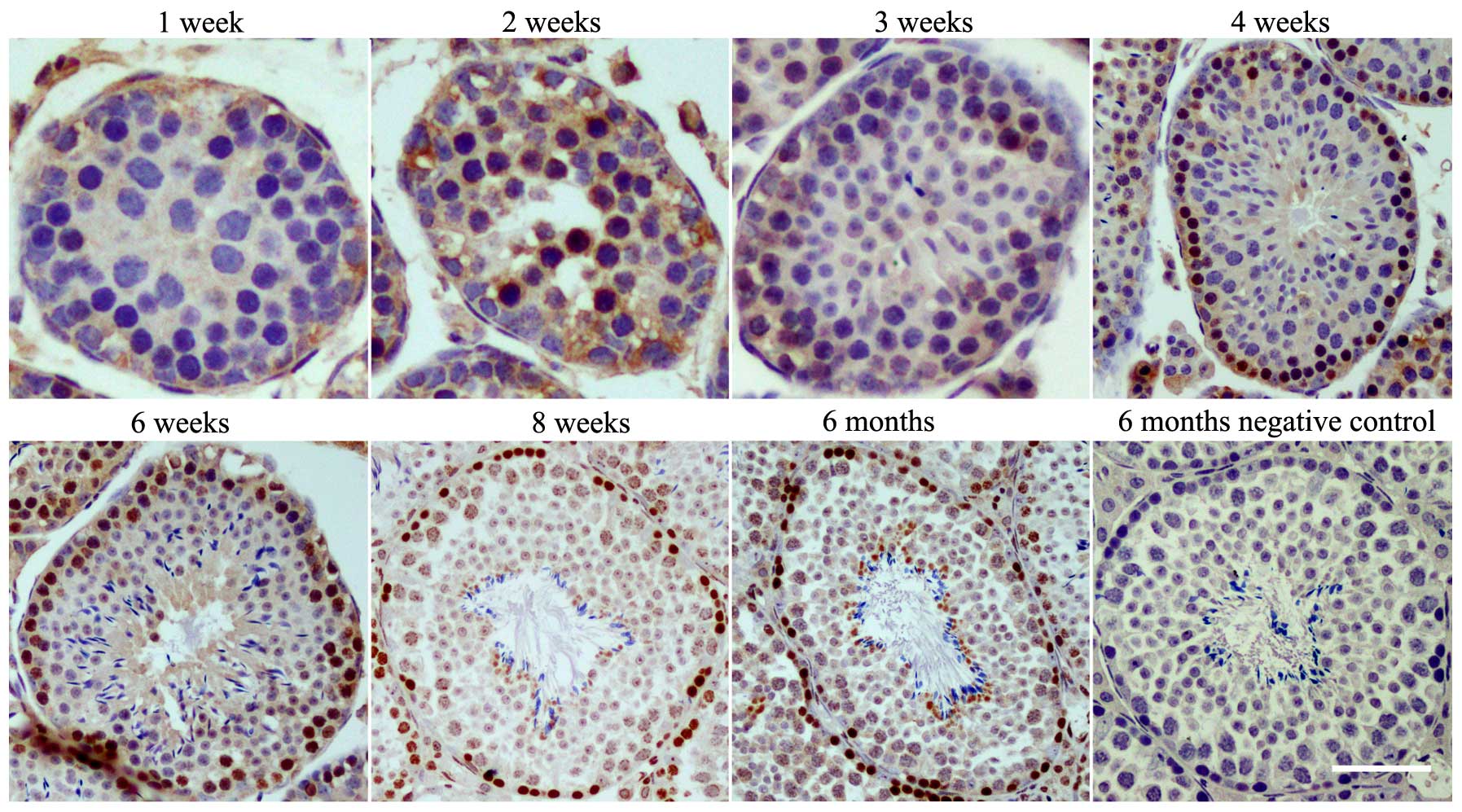
CCDC38 protein is predominantly expressed
in spermatogonia and spermatocytes
Immunocytochemical staining on sections of the
immature and adult mouse testes was employed to investigate the
expression levels CCDC38 protein in testes. CCDC38 was
predominantly expressed in the nucleus of the spermatogonia and
spermatocytes of mice aged from 2 weeks to adults (8 weeks)
(Fig. 3).
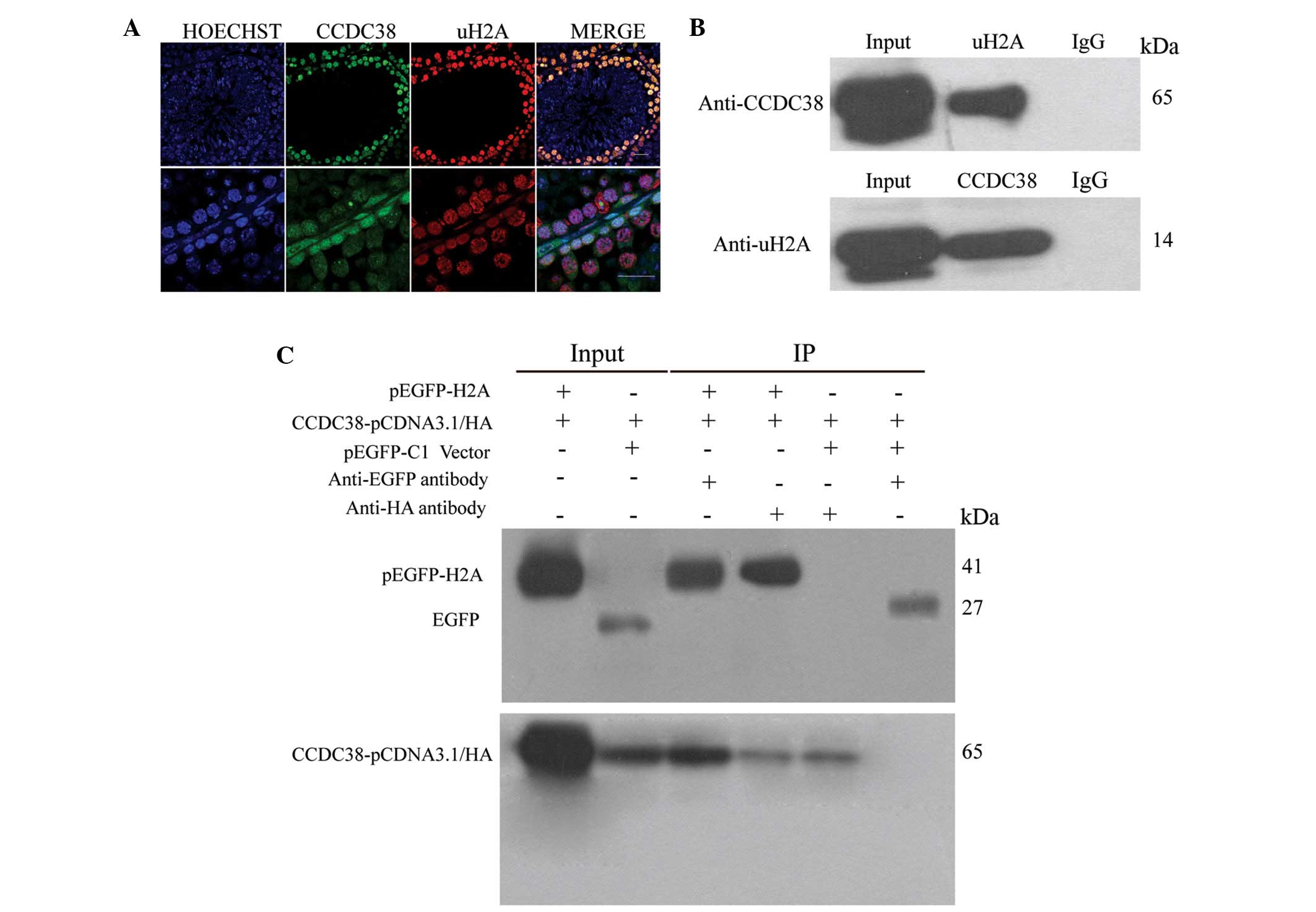
CCDC38 protein interacts with uH2A
CCDC38 and uH2A were examined by immunofluorescence
in the testes of adult mice. The results indicated that CCDC38 and
uH2A protein were partially co-localized in the nucleus of
spermatogonia and spermatocytes (Fig.
4A). The co-IP results indicated that the CCDC38 protein may be
co-immunoprecipitated by anti-uH2A and anti-EGFP antibodies in
vivo and in vitro, which demonstrated that CCDC38
interacted with uH2A in mouse testes (Fig. 4B and C).
Discussion
There are ~2,000 genes regulating the process of
spermatogenesis (14), and further
studies of key genes that regulate this process will aid in the
understanding of spermatogenesis. In the present study, the
testes-specific gene, Ccdc38, was identified, which was
associated with spermatogenesis. The association was supported by
the multiple tissue analysis of Ccdc38, which indicated that
it was exclusively expressed in the testes. Further
immunohistochemistry analysis indicated that the CCDC38 protein was
mainly localized in the nuclei of spermatogonia and spermatocytes
of the seminiferous tubules. In addition, the mRNA and protein
expression levels of CCDC38 were increased postnatally, from
2–8 weeks in the mice testes. Previous studies have
indicated that murine germ cells enter the meiotic prophase ~10
days after birth and proceed with the first wave of meiosis during
the remaining 10 days in the mice (15,16).
Therefore, it is possible that the developmentally-regulated
expression pattern of CCDC38 may contribute to spermatogenic
events.
Spermatogenesis involves various histone
modifications including ubiquitination, phosphorylation,
methylation and acetylation (17).
A previous study indicated that a high quantity of uH2A was
detected in the pachytene stage of spermatocytes, by immunoblot and
immunohistochemical analysis of wild-type mice testes (18). The current study indicated that
CCDC38 protein is localized in nuclei of spermatogonia and
spermatocytes. CCDC38 may interact with uH2A in murine testes, as
indicated by the use of a vector machine, previous experimental
results, combined with auto-covariance for prediction of
protein-protein interactions from protein sequences (19). In order to test this hypothesis
immunocolocalization and co-IP assays were performed. It was
determined that CCDC38 and uH2A partially co-localized in the
nuclei of spermatogonia and spermatocytes. Additionally, co-IP
demonstrated that CCDC38 may interact with uH2A in vivo and
in vitro. H2A ubiquitination contributes to various cellular
processes, including DNA damage repair and transcriptional
regulation by gene silencing or repression (20–22).
Therefore, it is possible that CCDC38 may be important for
transcriptional silencing via interactions with uH2A during
spermatogenesis.
In conclusion, the present study identified the
testes-specific gene Ccdc38 is conserved in mammalian
species. The expression and localization of the CCDC38 protein
indicates that it may be important for spermatogenesis. In
addition, CCDC38 may also regulate spermatogenesis by interacting
with uH2A. The molecular mechanisms of CCDC38 in spermatogenesis
require further investigation.
Acknowledgments
The present study was funded by grants from the
National Natural Science Foundation of China (grant nos. 31271244
and 31471344) and Shenzhen Project of Science and Technology (grant
nos. XB201104220045A and JCYJ20140415162543017).
References
|
1
|
De Kretser DM and Baker HW: Infertility in
men: Recent advances and continuing controversies. J Clin
Endocrinol Metab. 84:3443–3450. 1999.PubMed/NCBI
|
|
2
|
Chen H, Yi M, Sheng Y, Cheng H and Zhou R:
A novel testis-enriched gene Spata33 is expressed during
spermatogenesis. PLoS One. 8:e678822013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Luk AC, Chan WY, Rennert OM and Lee TL:
Long noncoding RNAs in spermatogenesis: Insights from recent
high-throughput transcriptome studies. Reproduction. 147:R131–R141.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dong WW, Huang HL, Yang W, Liu J, Yu Y,
Zhou SL, Wang W, Lv XC, Li ZY, Zhang MY, et al: Testis-specific
Fank1 gene in knockdown mice produces oligospermia via apoptosis.
Asian J Androl. 16:124–130. 2014. View Article : Google Scholar :
|
|
5
|
Xu K, Yang L, Zhao D, Wu Y and Qi H: AKAP3
synthesis is mediated by RNA binding proteins and PKA signaling
during mouse spermiogenesis. Biol Reprod. 90:1192014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mariotti M, Smith TF, Sudmant PH and
Goldberger G: Pseudogenization of testis-specific Lfg5 predates
human/Neanderthal divergence. J Hum Genet. 59:288–291. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoneda R and Kimura AP: A testis-specific
serine protease, Prss41/Tessp-1, is necessary for the progression
of meiosis during murine in vitro spermatogenesis. Biochem Biophys
Res Commun. 441:120–125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wei Y, Wang X, Fu G and Yu L: Testis
specific serine/threonine kinase 4 (Tssk4) maintains its kinase
activity by phosphorylating itself at Thr-197. Mol Biol Rep.
40:439–447. 2013. View Article : Google Scholar
|
|
9
|
Li D and Lu GX: Identification and
expression of a novel human testis-specific gene by digital
differential display. Chin Med J (Engl). 117:1791–1796. 2004.
|
|
10
|
Wu C, Orozco C, Boyer J, Leglise M,
Goodale J, Batalov S, Hodge CL, Haase J, Janes J, Huss JW III and
Su AI: BioGPS: An extensible and customizable portal for querying
and organizing gene annotation resources. Genome Biol. 10:R1302009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Y, Qin D, Tang A, Zhou D, Qin J, Yan
B, Diao R, Jiang Z, Cai Z and Gui Y: Developmental expression
pattern of a novel gene, TSG23/Tsg23, suggests a role in
spermatogenesis. Mol Hum Reprod. 15:223–230. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wilkins MR, Gasteiger E, Bairoch A,
Sanchez JC, Williams KL, Appel RD and Hochstrasser DF: Protein
identification and analysis tools in the ExPASy server. Methods Mol
Biol. 112:531–552. 1999.PubMed/NCBI
|
|
14
|
Krausz C and Giachini C: Genetic risk
factors in male infertility. Arch Androl. 53:125–133. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goetz P, Chandley AC and Speed RM:
Morphological and temporal sequence of meiotic prophase development
at puberty in the male mouse. J Cell Sci. 65:249–263.
1984.PubMed/NCBI
|
|
16
|
Bellvé AR, Cavicchia JC, Millette CF,
O'Brien DA, Bhatnagar YM and Dym M: Spermatogenic cells of the
prepuberal mouse. Isolation and morphological characterization. J
Cell Biol. 74:68–85. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
An JY, Kim EA, Jiang Y, Zakrzewska A, Kim
DE, Lee MJ, Mook-Jung I, Zhang Y and Kwon YT: UBR2 mediates
transcriptional silencing during spermatogenesis via histone
ubiquitination. Proc Natl Acad Sci USA. 107:1912–1917. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baarends WM, Hoogerbrugge JW, Roest HP,
Ooms M, Vreeburg J, Hoeijmakers JH and Grootegoed JA: Histone
ubiquitination and chromatin remodeling in mouse spermatogenesis.
Dev Biol. 207:322–333. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo Y, Yu L, Wen Z and Li M: Using support
vector machine combined with auto covariance to predict
protein-protein interactions from protein sequences. Nucleic Acids
Res. 36:3025–3030. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Leeb M and Wutz A: Ring1B is crucial for
the regulation of developmental control genes and PRC1 proteins but
not X inactivation in embryonic cells. J Cell Biol. 178:219–229.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Richly H and Di Croce L: The flip side of
the coin: Role of ZRF1 and histone H2A ubiquitination in
transcriptional activation. Cell Cycle. 10:745–750. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Richly H, Rocha-Viegas L, Ribeiro JD,
Demajo S, Gundem G, Lopez-Bigas N, Nakagawa T, Rospert S, Ito T and
Di Croce L: Transcriptional activation of polycomb-repressed genes
by ZRF1. Nature. 468:1124–1128. 2010. View Article : Google Scholar : PubMed/NCBI
|