Introduction
Idiopathic pulmonary fibrosis (IPF) is a common and
life-threatening disease, with a median survival rate of ~3–5 years
following diagnosis. However, its pathogenesis is complex and
remains to be fully elucidated (1). Microarray analysis has shown that the
instillation of bleomycin, a profibrotic drug, alters the gene
transcription pattern in the mouse lung by increasing the
expression of proinflammatory mediators, certain components of the
pulmonary extracellular matrix and genes that are induced by
transforming growth factor-β1 (TGF-β1) (2). TGF-β1 contributes to pulmonary
fibrosis in numerous animal models and in human pulmonary fibrotic
disease by inducing the epithelial-to-mesenchymal transition (EMT)
of alveolar epithelial cells (3,4).
Human cytomegalovirus (HCMV) is a member of the
herpes family of viruses. CMV infection is relatively common, with
~40–70% of adults being infected in the latent state. Latent CMV
infection, defined by being a carrier of the CMV genome without
active replication, is commonly asymptomatic in immunocompetent
individuals, whereas active CMV infection is associated with
clinical signs and symptoms, which include fever, sore throat and
leukopenia. HCMV infects the respiratory tract, and it has been
evaluated with regards to its association with IPF. Patients with
IPF show significantly higher DNA copy numbers in their blood,
compared with healthy controls (5). There is evidence that suggests an
association between the incidence of HCMV infection and the
incidence of IPF, and that CMV induces the secretion of TGF-β1 from
infected fibroblasts, astrocytes and osteosarcoma cells (6). However, the mechanism by which HCMV
may affect fibrosis remains to be elucidated.
In the present study, a mouse model of murine CMV
(MCMV) infection was used to investigate the hypotheses that CMV
may trigger IPF in a susceptible host, and/or that the presence of
CMV may alter IPF in response to a well-defined trigger of
pulmonary fibrosis, namely the chemical bleomycin. In addition, the
precedent exists for the possibility that CMV-infected alveolar
epithelial cells may induce the production or activation of TGF-β1
in pathological settings. TGF-β1 has been associated with EMT and
promotion of pulmonary fibrosis.
Materials and methods
Animals and materials
BALB/c mice (age, 4–6 weeks; weight, 15.9±1.5 g;
n=27) were purchased from The Jackson Laboratory (Bar Harbor, ME,
USA) and maintained at 18–22°C with a 10–14 h light/dark cycle and
access to food and water ad libitum. The mice were divided
into 3 groups of 9 mice. MCMV was cultured, as described previously
(7). In brief, the salivary
gland-passed MCMV (Smith strain) was prepared by homogenizing the
salivary glands of 10 BALB/c mice, sacrificed by cervical
dislocation, which had been infected with 104 plaque
forming units (PFU) 3 weeks previously. Aliquots (1 ml) of the
homogenized supernatants of the salivary glands were stored in
liquid nitrogen in RPMI 1640 (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) with 5% fetal bovine serum (Thermo Fisher
Scientific, Inc.), and titrated onto 2.5×105/ml NIH-3T3
cells (American Type Culture Collection, Manassas, VA, USA) and
cultured at 37°C in 5% CO2 for 7 days. Polymerase chain
reaction (PCR) reagents were purchased from Applied Biosystems;
Thermo Fisher Scientific, Inc. The PCR primers were purchased from
Invitrogen; Thermo Fisher Scientific, Inc. Bleomycin was purchased
from Sigma-Aldrich (St. Louis, MO, USA). The present study was
approved by the ethics committee of Anhui Medical University
(Hefei, China).
Mouse treatments
The salivary gland-passaged MCMV was suspended in
200 µl of Hanks' balanced salt solution (Thermo Fisher
Scientific, Inc.) with 3% fetal bovine serum. The mice were
infected via intraperitoneal injection with 105 PFU or,
as a control, mice were injected with Hanks' balanced salt
solution/3% fetal bovine serum. The mice (n=9; 3/group) were
sacrificed by cervical dislocation 3, 14 and 28 days following
infection and viral loads were analyzed using PCR. After 4 weeks,
subsets of the MCMV-infected and uninfected mice, respectively,
were intratracheally instilled with bleomycin or phosphate-buffered
saline (PBS) to induce fibrotic changes. Briefly, the mice were
anesthetized with sevoflurane inhalation (Abbott Laboratories,
Abbott Park, IL, USA) and placed in dorsal recumbency.
Transtracheal insertion of a 24-G animal feeding needle was used to
instillate bleomycin (0.75 U/ml) or vehicle (PBS), in a volume of
80 µl. The body weights of the mice were measured at least
twice each week thereafter. The mice were sacrificed 7 and 14 days
following instillation, and the lungs were removed for further
analysis, as described below.
PCR assessment of viral loads
Extraction of the MCMV DNA from the tissues was
performed using a QIAamp DNA Mini kit (Qiagen, Hilden, Germany). In
brief, 25 mg samples of tissues (10 mg of spleen; salivary gland;
lung; liver) were sonicated following the addition of 80 µl
PBS. Qiagen lysing buffer (100 µl; Qiagen) and 20 µl
proteinase K (Thermo Fisher Scientific, Inc.) were added, and the
sample was incubated at 56°C for ~3 h until it was completely
lysed. RNase A (200 µl) was then added, along with Qiagen
buffer AL (Qiagen). The sample was incubated at 70°C for 10 min and
200 µl ethanol (96–100%) was added, followed by
centrifugation at 6,000 × g for 1 min at 4°C in a spin column
several times with 500 µl Qiagen buffers AW1 and AW2
(Qiagen). The sample was then incubated for 5 min at room
temperature and centrifuged at 6,000 × g for 1 min at 4°C using 150
µl distilled water, following which the filtrate containing
the MCMV DNA was collected. The DNA quality was confirmed in an
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara,
CA, USA). The viral loads of MCMV were determined using standard
and real-time quantitative (q) PCR using an ABI 7500 (Applied
Biosystems; Thermo Fisher Scientific, Inc.). A quantity of 50 ng
total DNA (5 µl) was used in each reaction with 26.5
µl PCR Master mix (Thermo Fisher Scientific, Inc.), 15.75
µl distilled water, 1 µl forward and reverse primers
(10 µM), and the primer sequences were as follows: forward
5′-ATCTGGTGCTCCTCAGATCAGCTAA-3′ and reverse
5′-ATTGTTCATTGCCTGGGGAGTTT-3′. The thermocycling conditions were as
follows: 95°C for 10 min; and 40 cycles of 95°C for 15 sec and 60°C
for 1 min. The experiments were performed three times independently
and results were compared using the 2−∆∆Cq method
(8).
Histopathology
Lungs from each mouse were fixed overnight with
neutral-buffered formalin (Sigma-Aldrich) and embedded in paraffin
(Sigma-Aldrich). Sections of tissue (5 µM thick) were
mounted and stained with hematoxylin and eosin (Beyotime Institute
of Biotechnology, Haimen, China) to assess the degree of fibrosis.
The histological sections were reviewed by an experienced
pathologist using a fluorescence microscope (BX41; Olympus
Corporation, Tokyo, Japan).
Preparation of tissue extracts
The isolated lung tissues were rinsed in sterile
normal saline to remove blood and were briefly placed on a sterile
gauze to dry. To obtain tissue extracts, the collected tissues were
cut into small sections, and ~30–50 mg of the tissues were minced
and sonicated in 500 µl lysis buffer (Qiagen) with 50 mM
Tris-HCl (pH 7.5), containing 100 mM sodium fluoride, 30 mM sodium
pyrophosphate, 2 mM sodium molybdate, 1 mM sodium orthovanadate, 1
mM glycerophosphate and 1X protease inhibitor cocktail, on ice. The
samples were centrifuged at 12,000 × g for 20 min at 4°C. The clear
supernatant was collected and stored in aliquots at −80°C. Protein
quantification of the lysate was performed using the bicinchoninic
acid method.
Bioassay of TGF-β1 activity
TGF-β1 activity was measured by the inhibition of
(3H) thymidine incorporation into lung epithelial cells (CCL64), as
described by Jennings et al (9). Acid activation was performed, with
minor modification, to isolate free TGF-β molecules from the latent
complex. Briefly, 30 µl of tissue extracts (equivalent to
100–300 µg protein) were added to 200 µl of minimal
essential medium/bovine serum albumin, followed by the addition of
10 µl 4 N HCl, and agitated for 1 h at 4°C. Acid activation
was terminated by neutralization with 10 µl 4 N NaOH. The
tissue extracts were assessed prior to and following activation.
The activated tissue extracts were also examined following the
neutralization of TGF-β1 activity with rabbit anti-pan-TGF-β serum
(British Biotechnology, Oxon, UK; 1:100) at 37°C for 30 min. The
controls consisted of (3H) thymidine incorporation into cells
treated with a range of concentrations of purified TGF-β1 with and
without anti-TGF-β antibodies, cells incubated with 4 mM HCl,
TGF-β-neutralizing serum alone (1:100), medium alone, or non-immune
rabbit serum.
Reverse-transcription-qPCR
Total RNA was obtained from the lung tissues using
an RNeasy kit (Qiagen), according to the manufacturer's protocol.
The RNA quality was confirmed using an Agilent 2100 Bioanalyzer
(Agilent Technologies, Inc.). The RNA was reverse transcribed into
cDNA using an RT2 First Strand kit (Qiagen), and the resulting
cDNAs were used according to the manufacturer's protocol of the
commercial primer/probe sets (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The PCR was conducted as described above. The
results were normalized to 18S RNA and expressed as the fold change
from baseline (control group). The experiments were performed three
times independently and the results were compared using a standard
curve.
Immunoblotting assays
The protein in the lung tissue extracts was
quantified using the BCA Protein assay kit (Beyotime Institute of
Biotechnology) 50 µg samples were separated on 10% SDS-PAGE
gels (Thermo Fisher Scientific, Inc.), which were then transferred
onto polyvinylidene fluoride membranes (Merck Millipore, Darmstadt,
Germany).
Subsequently, the membranes were blocked with 50 ml
3% fat free milk for 1 h and incubated with primary antibodies as
follows: Mouse monoclonal anti-E-cadherin (1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA; cat. no. 14472), mouse
monoclonal anti-Vimentin (1:1,000; Cell Signaling Technology, Inc.;
cat. no. 3390), rabbit monoclonal phospho-SMAD2 (1:1,000; Cell
Signaling Technology, Inc.; cat. no. 3108) and β-actin (1:500;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA; sc-8432) at 4°C
overnight. Following washing with PBS three times, the membranes
were incubated with horseradish peroxidase-conjugated goat
anti-mouse (1:2,500; Santa Cruz Biotechnology, Inc.; cat. no. 2005)
and goat anti-rabbit (1:2,500; Santa Cruz Biotechnology, Inc.; cat.
no. sc-2004) secondary antibodies. at room temperature for 1 h. The
membranes were washed again, and the antigen-antibody reaction was
visualized and analyzed using an Amersham ECL detection system (GE
Healthcare Life Sciences, Amersham, UK).
Statistical analysis
Statistical analysis was performed with GraphPad
Prism 6.01 software (GraphPad Software, Inc., La Jolla, CA, USA)
using Student's two-tailed t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
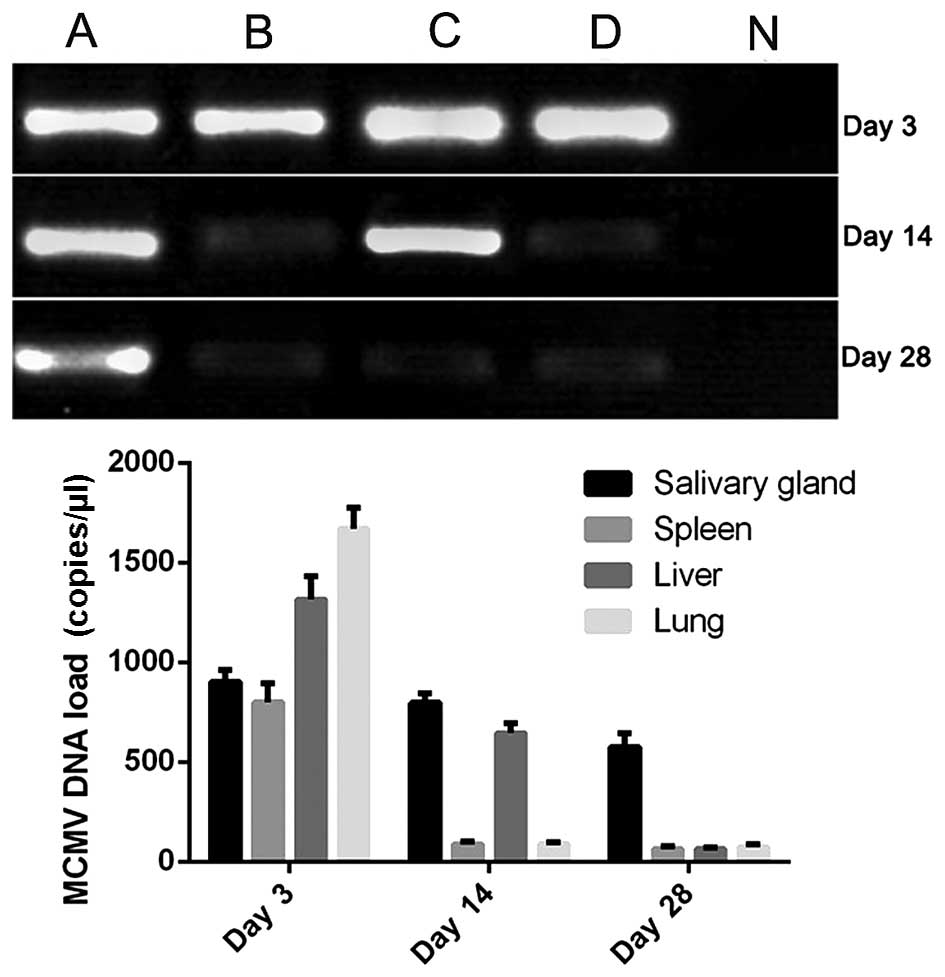
RT-qPCR results
The results of the RT-qPCR analyses are shown in
Fig. 1. The results revealed that
MCMV invaded the lung tissues, however it did not cause pulmonary
fibrosis. The viral DNA loads in the salivary glands, spleen, liver
and lungs were highest following inoculation for 3 days. Subsequent
to this, the numbers gradually reduced and were maintained at a low
level, which is similar the latent infection in humans.
Weight changes in the mice
Changes in the body weights of the mice are shown in
Fig. 2. The results suggested that
the weights of the mice decreased sharply following infection for 3
days, following which the weights recovered rapidly in the infected
group. No significant differences were observed between the control
group and infected group from 15 days post-infection.
Detection of TGF-β1 activity
The results of the analysis of TGF-β1 activity are
shown in Fig. 3. It was found that
the activity of TGF-β1 was higher in the MCMV+bleomycin group,
compared with the activity of TGF-β1 in the bleomycin group and
control group following treatment for 7 days. The activity reached
the highest level following treatment for 14 days. These results
suggested that the progression of fibrosis in the mice treated with
MCMV+bleomycin was more rapid, compared with that in the control
mice.
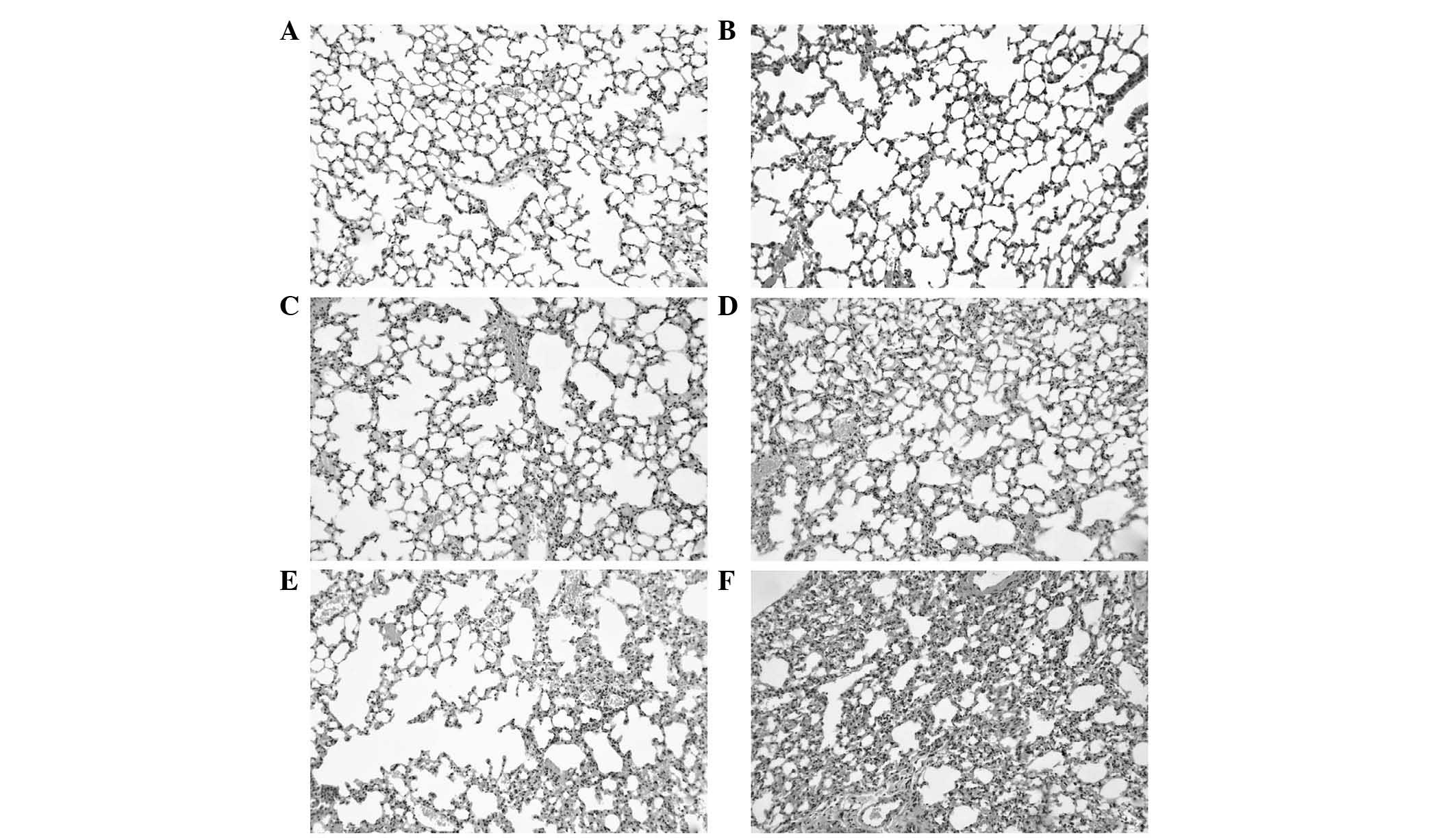
Detection of histopathology
The results of the pathological detection are shown
in Fig. 4. The results showed that
the degree of pulmonary fibrosis was more marked in the
MCMV+bleomycin group, compared with the bleomycin group following
treatment for 7 and 14 days.
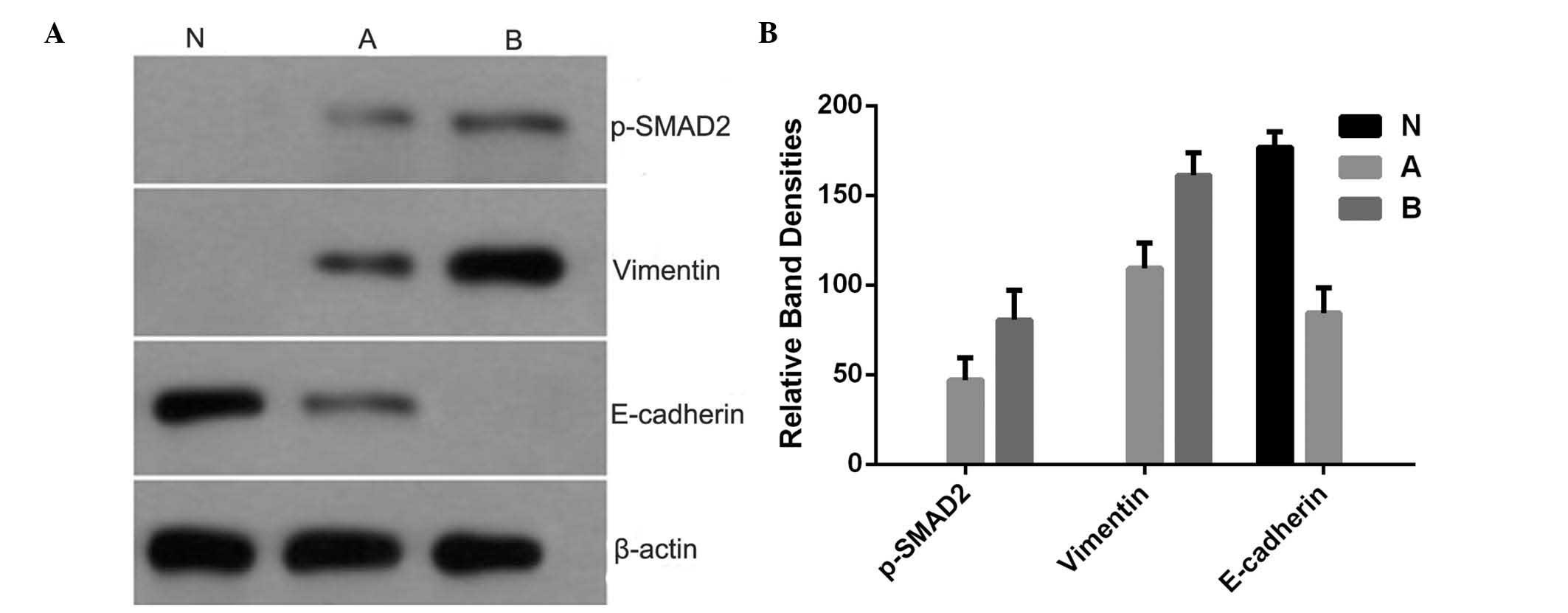
Western blot analysis of protein
expression levels
The results of the western blot analysis are shown
in Fig. 5. The results showed that
the protein levels of Vimentin and phospho-SMAD2 increased
following treatment with bleomycin and MCMV+bleomycin, whereas the
protein levels of E-cadherin decreased following treatment with
bleomycin and MCMV+bleomycin.
Discussion
The etiology of IPF remains to be elucidated.
However, previous studies based on animal models of pulmonary
fibrosis and lung tissues from patients with IPF have suggested a
dynamic pathobiological process, involving excessive wound healing
with chronic inflammation, apoptosis of epithelial and endothelial
cells, mesenchymal cell proliferation and activation with the
formation of fibroblasts/myofibroblasts foci, and excessive
deposition of extracellular matrix resulting in destruction of the
lung architecture and loss of lung functions (10). High throughput genomic profiling
studies have characterized certain gene changes, which occur during
the development of pulmonary fibrosis (9). During EMT, alveolar epithelial cells
demonstrate loss of epithelial characteristics and cellular
adhesions, develop changes in the actin cytoskeleton, induce the
expression of fibrogenic molecules and acquire a migratory
phenotype (11). These
fibroblastoid alveolar epithelial cells are key contributors to
pulmonary fibrosis, as the inhibition of TGF-β1-mediated EMT
prevents and reverses experimentally induced pulmonary fibrosis in
animal models (12,13).
Infection with HCMV is bimodal, occurring through
vertical and horizontal transmission in early childhood and through
sexual transmission in young adults. The viral loads in the
bronchoalveolar lavage cells of patients with IPF and healthy
volunteers are elevated, relative to the respective viral load in
the blood leukocytes, suggesting that the lungs are important in
the pathobiology of HCMV (14,15).
The production of TGF-β1 can be induced by transient transfection
with expression plasmids containing the HCMV immediate early 1 and
2 genes into fibroblasts and astrocytoma cells. In previous
studies, increases in TGF-β1 were associated with the induction of
TGF-β1 mRNA. However, the local effects of TGF-β1 are often
controlled in vivo by activation of the extracellular latent
form (16,17). In the present study, it was found
that MCMV invaded the lungs, however it did not cause pulmonary
fibrosis. The progression of fibrosis in the mice treated with
MCMV+bleomycin was more rapid, compared with that in the control
mice, and the degree of pulmonary fibrosis was more severe in the
MCMV+bleomycin group, compared with the bleomycin group following
treatment. The protein levels of EMT-associated genes, including
Vimentin and phospho-SMAD2 were upregulated following treatment
with MCMV+bleomycin. These results suggested that latent MCMV
infection aggravated pulmonary fibrosis in the mouse model,
possibly through the activation of TGF-β1.
In conclusion, the present study found that the
presence of CMV may alter IPF in response to a well-defined trigger
of pulmonary fibrosis, namely the chemical bleomycin. Latent MCMV
infection may have aggravated pulmonary fibrosis in the mouse model
through the activation of TGF-β1.
References
|
1
|
Hunninghake GW and Kalica AR: Approaches
to the treatment of pulmonary fibrosis. Am J Respir Crit Care Med.
151:915–918. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Munger JS, Huang X, Kawakatsu H, Griffiths
MJ, Dalton SL, Wu J, Pittet JF, Kaminski N, Garat C, Matthay MA, et
al: The integrin alpha v beta 6 binds and activates latent TGF beta
1: A mechanism for regulating pulmonary inflammation and fibrosis.
Cell. 96:319–328. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Haider Y, Malizia AP, Keating DT, Birch M,
Tomlinson A, Martin G, Ferguson MW, Doran PP and Egan JJ: Host
predisposition by endogenous transforming growth factor-beta1
overexpression promotes pulmonary fibrosis following bleomycin
injury. J Inflamm (Lond). 4:182007. View Article : Google Scholar
|
|
4
|
Kalluri R and Neilson EG:
Epithelial-mesenchymal transition and its implications for
fibrosis. J Clin Invest. 112:1776–1784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dworniczak S, Ziora D, Kapral M, Mazurek
U, Niepsuj G, Rauer R, Wilczok T and Kozielski J: Human
cytomegalovirus DNA level in patients with idiopathic pulmonary
fibrosis. J Physiol Pharmacol. 55(Suppl 3): S67–S75. 2004.
|
|
6
|
Michelson S, Alcami J, Kim SJ, Danielpour
D, Bachelerie F, Picard L, Bessia C, Paya C and Virelizier JL:
Human cytomegalovirus infection induces transcription and secretion
of transforming growth factor beta 1. J Virol. 68:5730–5737.
1994.PubMed/NCBI
|
|
7
|
Desmouliére A, Darby IA and Gabbiani G:
Normal and pathologic soft tissue remodeling: Role of the
myofibroblast, with special emphasis on liver and kidney fibrosis.
Lab Invest. 83:1689–1707. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Akbari A, Ghahremani MH, Mobini GR,
Abastabar M, Akhtari J, Bolhassani M and Heidari M: Down-regulation
of miR-135b in colon adenocarcinoma induced by a TGF-β receptor I
kinase inhibitor (SD-208). Iran J Basic Med Sci. 18:856–861.
2015.PubMed/NCBI
|
|
9
|
Jennings JC, Mohan S, Linkhart TA,
Widstrom R and Baylink DJ: Comparison of the biological actions of
TGF beta-1 and TGF beta-2: Differential activity in endothelial
cells. J Cell Physiol. 137:167–172. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sime PJ and O'Reilly KM: Fibrosis of the
lung and other tissues: New concepts in pathogenesis and treatment.
Clin Immunol. 99:308–319. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kasai H, Allen JT, Mason RM, Kamimura T
and Zhang Z: TGF-beta1 induces human alveolar epithelial to
mesenchymal cell transition (EMT). Respir Res. 6:562005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu YD, Hua J, Mui A, O'Connor R,
Grotendorst G and Khalil N: Release of biologically active
TGF-beta1 by alveolar epithelial cells results in pulmonary
fibrosis. Am J Physiol Lung Cell Mol Physiol. 285:L527–L539. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Y: Epithelial to mesenchymal
transition in renal fibrogenesis: Pathologic significance,
molecular mechanism, and therapeutic intervention. J Am Soc
Nephrol. 15:1–12. 2004. View Article : Google Scholar
|
|
14
|
Kossmann T, Morganti-Kossmann MC,
Orenstein JM, Britt WJ, Wahl SM and Smith PD: Cytomegalovirus
production by infected astrocytes correlates with transforming
growth factor-beta release. J Infect Dis. 187:534–541. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kwon YJ, Kim DJ, Kim JH, Park CG, Cha CY
and Hwang ES: Human cytomegalovirus (HCMV) infection in
osteosarcoma cell line suppresses GM-CSF production by induction of
TGF-beta. Microbiol Immunol. 48:195–199. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koli K, Saharinen J, Hyytiäinen M,
Penttinen C and Keski-Oja J: Latency, activation, and binding
proteins of TGF-beta. Microsc Res Tech. 52:354–362. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Annes JP, Munger JS and Rifkin DB: Making
sense of latent TGFbeta activation. J Cell Sci. 116:217–224. 2003.
View Article : Google Scholar
|