Introduction
Bladder cancer (BC) is one the most common
urological malignancies worldwide. In China, BC is the most common
genitourinary malignancy and its incidence has increased over the
last decades (1). Following
surgery and adjuvant chemotherapy, bladder cancer frequently recurs
in a more aggressive form (2,3).
Therefore, it is necessary to develop novel treatment strategies to
improve the poor prognosis of bladder cancer patients.
MicroRNAs (miRs) are small non-coding RNAs which
regulate gene expression at transcriptional and
post-transcriptional levels (4).
Increasing evidence has indicated that miRs have important roles in
various biological processes (5–7).
Aberrant expression of miRs is known to be correlated with cancer
progression (8,9). Upregulation of certain oncogenic
miRs, including miR-145, miR-150, miR-19a and miR-155, in bladder
cancer cells has been documented (10–12).
However, certain tumor-suppressive miRs, including miR-320c and
miR-24–1, inhibit tumor invasion and metastasis (13,14).
miR-335 expression has been observed to be
deregulated in several types of cancer, and its implication in
tumorigenesis and metastasis of has also been reported (15,16).
In pediatric acute lymphoblastic leukemia, deregulated miR-335 that
targets mitogen-activated protein kinase (MAPK)1 has been
implicated in poor outcome (17).
However, the clinical significance and biological roles of miR-335
in bladder cancer have yet to be fully elucidated. Therefore, the
present study assessed the expression of miR-335 in bladder cancer
tissues and adjacent non-tumor tissues, as well as bladder cancer
cell lines. Furthermore, the effects of miR-335 on the
proliferation and migration of bladder cancer cells, in addition to
the underlying mechanisms, were investigated.
Materials and methods
Clinical specimens
Tissue samples were collected from surgical
specimens of 50 bladder cancer patients who underwent surgery at
Nantong Tumor Hospital (Nantong, China) between May 2010 and July
2014. The corresponding adjacent non-neoplastic tissues were
obtained at the same time and used as controls. All samples were
immediately snap-frozen in liquid nitrogen and stored at −80°C
prior to RNA extraction. Patient demographics are displayed in
Table I. The Clinical Research
Ethics Committee of Nantong Tumor Hospital (Nantong, China)
approved the study protocol and written informed consent was
obtained from all patients.
 | Table IClinical features and tumor
characteristics of patients in the present study. |
Table I
Clinical features and tumor
characteristics of patients in the present study.
| Clinical
characteristic | ACCs |
|---|
| Age, years |
| ≥65 | 27 |
| <65 | 23 |
| Gender |
| Male | 42 |
| Female | 8 |
| pT stage |
| Ta, Tis, T1 | 21 |
| T2–T4 | 29 |
| Tumor grade |
| Low grade | 19 |
| High grade | 31 |
| Lymph node
status |
| N0 | 17 |
| N1, N2 | 33 |
Cell culture
The T24 and 5637 human bladder cancer cell lines
were obtained from the Cell Bank of Type Culture Collection of
Chinese Academy of Sciences (Shanghai, China) as was the TCHu169
immortalized human bladder epithelial cell line, which was also
used. The bladder cancer cells were cultured in RPMI-1640 (Gibco;
Thermo Fisher Scientific, Waltham, MA, USA) medium supplemented
with 10% fetal bovine serum (FBS; Biological Industries, Cromwell,
CT, USA) and 100 µg/ml penicillin-streptomycin (Gibco;
Thermo Fisher Scientific), while the TCHu169 cells were propagated
in F-12K medium (Gibco; Thermo Fisher Scientific) containing 10%
FBS. Cells were incubated in a humidified atmosphere of 95% air and
5% CO2 at 37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Prior to RNA extraction, tissues were immersed in
RNAlater (Ambion; Thermo Fisher Scientific) and stored at 20°C.
Subsequently, total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific)
and quantified using a Synergy HT Multi-Mode Microplate Reader
(Biotek Instruments, Inc., Winooski, VT, USA). Total RNA, including
miR, was extracted using the mirVana miRNA isolation kit (Ambion).
RNA (1 µg) was reverse transcribed to cDNA using random
primers and a Reverse Transcription kit (Takara Biotechnology Co.,
Ltd., Dalian, China), in a final volume of 20 µl. RT was
performed at 37°C for 15 min, then at 85°C for 5 sec. qPCR was
performed using the Power SYBR Green PCR (Takara), according to the
manufacturer's protocol. RT-qPCR was performed using a SYBR Green
Premix Ex Taq (Takara Bio Inc., Otsu, Japan) in a Light Cycler 480
(Roche, Basel, Switzerland). U6 small nuclear RNA and β-actin mRNA
were used as internal controls. Primers were synthesized by
Shanghai Sangon Biological Engineering Technology Services
(Shanghai, China). All reactions were run in triplicate. The
primers had the following sequences: miR-335 forward,
5′-TCAAGAGCAATAACGAAAAATGT-3′ and reverse,
5′-GCTGTCAACGATACGCTACGT-3′; U6 forward, 5′-CGCTTCGGCAGCACATATAC-3′
and reverse, 5′-TTCACGAATTTGCGTGTCAT-3′; β-actin forward,
5′-AGTGTGACGTGGACATCCGCAAAG-3′ and reverse,
5′-ATCCACATCTGCTGGAAGGTGGAC-3′; miR-335 mimics forward,
5′-GCUAGAAGAACUAUUUGCUUU-3′ and reverse,
5′-AAAGCAAATAGTTCTTCTAGC-3′; miR-NC forward,
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-AAAGCTGACACGTTCGGAGAA-3′. The relative mRNA expression levels
were calculated using the 2−ΔΔCq method (18), in which values were normalized to
those of β-actin.
miRNA mimics, small interfering (si)RNAs
and transfection
miR-335 mimic and siRNAs specific for MAPK1 were
synthesized by RiboBio (Guangzhou, China). miR-335 mimics and
negative control miR (miR-NC) were used for gain-of-function
experiments, whereas MAPK1 siRNA and control siRNA were used in the
loss-of-function experiments. Transfection with miRNAs or siRNAs
was performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific) according to the manufacturer's instructions and the
cycling conditions were as follows: 94°C for 2 min for 2 cycles,
followed by 40 cycles of 94°C for 15 sec, 58°C for 25 sec, and 72°C
for 30 sec. Cells in the logarithmic growth phase were seeded in a
10 cm dish for RNA and protein extraction, in a six-well plate for
protein extraction and apoptosis assay, and in a 96-well plate for
MTT assay and luciferase reporter assay. Small interfering RNAs
(siRNAs) against MAPK1 were also obtained from RiboBio. The two
sets of siRNA sequences were as follows: siRNA-1 (negative control)
forward, 5′-CUCUACGUAAGAUCCAGCUUU-3′ and reverse,
5′-AGCUGGAUCUUACGUAGAGUU-3′; siRNA-2 forward,
5′-AGCAAAUAGUUCCUAGCUUUU-3′ and reverse,
5′-AAGCUAGGAACUAUUUGCUUU-3′.
Cell proliferation assay
Cells were transfected with 10 nM miRNA or siRNA by
reverse transfection and seeded in 96-well plates at
3×103 cells per well. After 72 h of incubation, cell
proliferation was determined using an MTT assay (Invitrogen)
according to the manufacturer's protocol. The number of cells per
well was determined by measuring the absorbance at 540 nm. All
experiments were performed in triplicate.
Apoptosis analysis
Following transfection for 48 h, cells were
harvested and re-suspended in 1X binding buffer (BioVision, Inc.,
Milpitas, CA, USA) at a density of 1×106 cells/ml.
Double staining with fluorescein isothiocyanate-conjugated Annexin
V and propidium iodide (PI; Annexin V-Phycoerythrin Apoptosis
Detection kit; BD Biosciences, Franklin Lakes, NJ, USA) was used to
evaluate the percentage of apoptotic cells. In the results of the
apoptosis analysis, the left upper, right upper, left lower and
right lower quadrants represent necrotic cells, late apoptotic
cells, normal cells and early apoptotic cells, respectively. The
right lower quadrant was selected for evaluation of the levels of
apoptosis in cells. The cells were immediately analyzed using a
FACScan flow cytometer (BD Biosciences), where Annexin V-positive
and PI-negative cells were designated as apoptotic cells. Finally,
optical density was determined at 540 nm by a POLARstar+Optima (BMG
Labtech GmbH, Ortenberg, Germany). The aforementioned experiment
was repeated three times, and the results were analyzed using Cell
Quest software (version 2.7; BD Biosciences) where the left upper,
right upper, left lower and right lower quadrants represent
necrotic cells, late apoptotic cells, normal cells and early
apoptotic cells, respectively. The right lower quadrant was
selected for evaluation of the levels of apoptosis in cells.
Cell migration assay
Transwell inserts with a pore size of 8 µm
(Corning, Inc., Corning, NY, USA) were used to determine the
migratory capacity of the tumor cells. After transfection for 24 h,
1×106 cells were starved in medium without FBS for 24 h,
then re-suspended in the FBS-free medium and placed in the top
chambers of the Transwell inserts with each experimental condition
performed in triplicate. The cells remaining on the upper membrane
were removed with cotton wool, while the cells that had migrated to
the bottom of the membrane were then fixed with 95% ethanol and
stained with 0.1% crystal violet (Beyotime Institute of
Biotechnology, Haimen, China) Five fields of view for each insert
were randomly selected and images were captured under a light
microscope (CKX41SF; Olympus Corporation, Tokyo, Japan) at ×200
magnification. All experiments were performed in triplicate.
Protein extraction and western blot
analysis
Bladder cancer cells were seeded onto six-well
plates (1×107 cells/well) and were transfected for 48 h.
Cells were lysed with radioimmunoprecipitation assay buffer
containing protease inhibitors (Thermo Fisher Scientific). The
protein concentration was determined using a bicinchoninic protein
assay kit (Pierce Biotechnology, Inc., Rockford, IL, USA). The
respective tissue proteins (30 µg) were loaded onto 10%
sodium dodecyl sulfate-polyacrylamide gels and separated by
electrophoresis, followed by transfer onto a polyvinylidene
difluoride membranes (Millipore, Billerica, MA, USA). The blots
were probed with primary rabbit anti-MAPK11 monoclonal antibody
(cat no. ab117949; Abcam, Cambridge, MA, USA) diluted 1:500 or
mouse monoclonal anti-β-actin antibody diluted 1:2,000 (cat no.
0869100; MP Biomedicals, Santa Ana, CA, USA) in Tris-buffered
saline with 0.05% Tween 20 containing 5% non-fat milk at 4°C
overnight. The membranes were washed 3 times for 10 min each using
Tris-buffered saline comprising 50 mM Tris (pH 7.4), 0.9% NaCl and
0.05% Tween-20, and were then incubated with horseradish
peroxidase-conjugated secondary antibodies: Anti-MAPK11 [1C2]
monclonal mouse antibody (cat no. ab117949; Abcam) and anti-actin
monoclonal antibody (1:10,000; cat no. 0869100; MP Biomedicals,
LLC, Santa Ana, CA, USA). Blots were visualized using enhanced
chemiluminescence substrates (Pierce Biotechnology, Inc.). β-Actin
was used as an endogenous control.
Statistical analysis
Data analyses were performed using the SPSS
statistical package 15.0 (SPSS Inc., Chicago, IL, USA). Values are
expressed as the mean ± standard deviation. Data were analyzed
using Pearson's χ2 test and Fisher's exact test.
P<0.05 was considered to indicate a statistically significant
difference between values.
Results
miR-335 is downregulated in bladder
cancer tissues
To the best of our knowledge, the present study was
the first to assess the expression of miR-335 in human bladder
cancer. RT-qPCR analysis was employed to quantify the expression
levels of miR-335 in 50 pairs of bladder cancer tissues and the
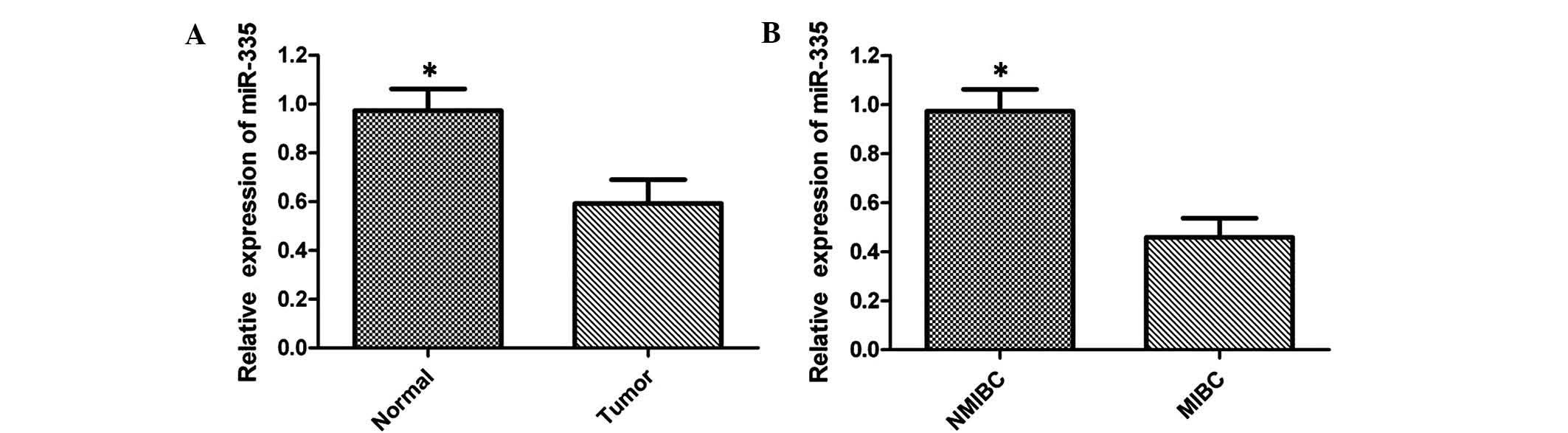
adjacent non-neoplastic tissues. The results revealed that miR-335
expression levels in bladder cancer tissues were significantly
lower compared with those in healthy tissues, and 29/50 samples
displayed a reduction of ≤50% (Fig.
1A). To further investigate the association between the
expression of miR-335 and the clinicopathological characteristics,
the levels of miR-335 between muscle invasive bladder cancer (MIBC)
and non-muscle invasive bladder cancer (NMIBC) were compared. The
results indicated that expression of miR-335 was lower in MIBC
compared with that in NMIBC, suggesting that miR-335 expression was
significantly associated with an aggressive tumor phenotype
(Fig. 1B). These results indicated
that miR-335 may represent a potential tumor suppressor in bladder
cancer.
miR-335 is downregulated in bladder
cancer cell lines
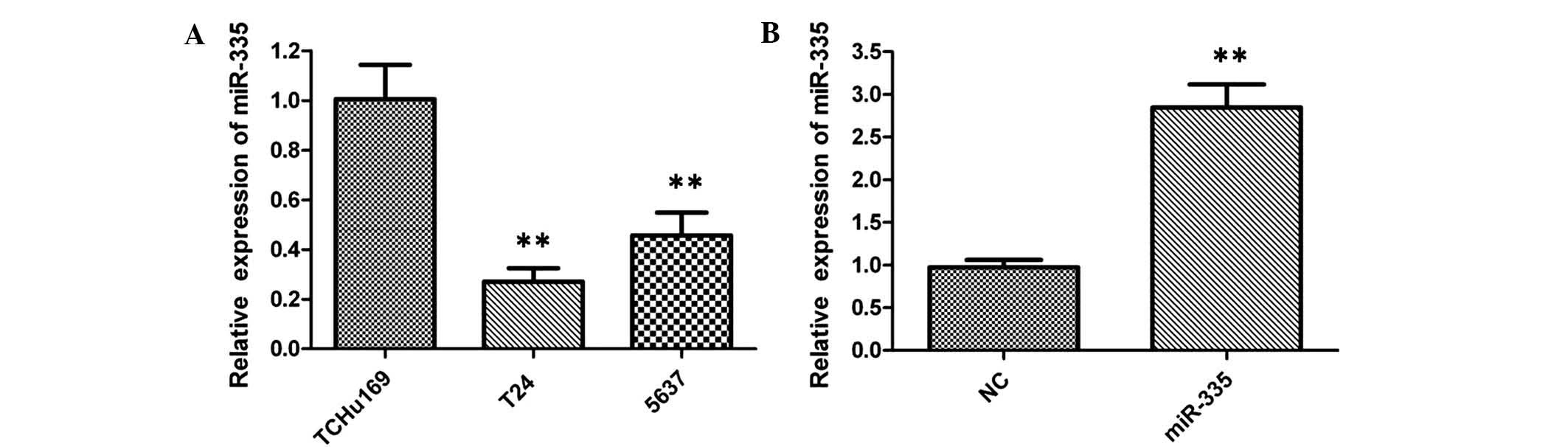
When compared with the TCHu169 normal bladder cell
line, the expression of miR-335 was significantly downregulated in
T24 and 5637 bladder cancer cells, indicating that low levels of
miR-335 may be relevant to the development of bladder cancer
(Fig. 2A). After transfection of
T24 cells with miR-335 mimics for 48 h, upregulation of miR-335
expression levels compared to those in miR-NC-transfected cells
were verified by RT-qPCR (Fig.
2B).
miR-335 suppresses bladder cancer cell
proliferation and migration, and induces apoptosis
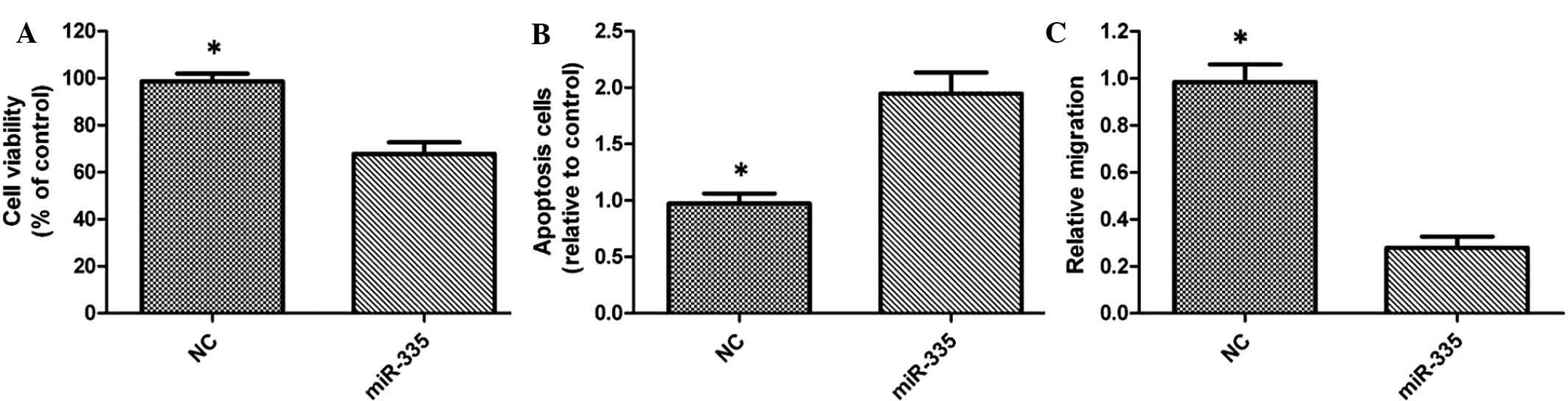
In order to assess whether miR-335 is able to
suppress bladder cancer progression in vitro, the present
study performed a gain-of-function study via transfection of
miRNA-335 into T24 cells. The results of the MTT assay revealed
that overexpression of miR-335 significantly inhibited the
proliferation of miR-335-transfected T24-cells (Fig. 3A). Flow cytometric analysis
revealed a significant increase in the number of apoptotic cells in
miR-335-transfected cells, when compared with that in the control
group (Fig. 3B). Furthermore, the
Transwell assay indicated a significant inhibition of T24 cell
migration following transfection with miR-335 (Fig. 3C).
miR-335 exerts is tumor-suppressor
effects in bladder cancer through targeting MAPK1
The present study aimed to elucidate the underlying
mechanism of the tumor-suppressive role of miR-335 in bladder
cancer. MAPK1 has been verified as a functional target of miR-335,
as miR-335 efficiently controls MAPK1 expression by directly
targeting a sequence motif in the coding region of MAPK1 in
pediatric acute lymphoblastic leukemia (17). MAPK signaling is associated with
proliferation and drug resistance in a variety of cancers (19). However, the possible involvement of
MAPK1 in the tumor suppressor role of miR-334 in bladder cancer has
remained elusive.
To further confirm that MAPK1 is a target gene of
miR-335, RT-qPCR and western blot analysis were used to detect
effects of miR-335 mimics on the expression of MAPK1 in T24 cells.
The expression of MAPK1 was evidently decreased after
overexpression of miR-335 at the mRNA (Fig. 4A) and protein level (Fig. 4B) compared with that in the
negative control-transfected cells. These results indicated that
miR-335 downregulated the expression of MAPK1 in bladder cancer
cells.
Inhibition of MAPK1 suppresses the
proliferation and migration of T24 cells
To investigate whether miR-335 exerts its
tumor-suppressive function through the MAPK1 pathway, MAPK1 was
silenced in T24 cells using siRNA. RT-qPCR and western blot
analyses confirmed that the siRNAs siMAPK1_1 and siMAPK1_2
significantly decreased the mRNA and protein expression of MAPK1
(Fig. 5A and B). Following MAPK1
silencing, the growth rate of T24 cells was significantly reduced
compared with that in the negative control-transfected group
(Fig. 5C). Furthermore, a
Transwell assay demonstrated that the migratory capacity of cells
transfected with siMAPK1_1 or siMAPK_2 was significantly inhibited
compared with that of control-transfected cells (Fig. 5D).
Discussion
Aberrant expression of miRs is implicated in the
pathogenesis of most cancer types (20), and miRs have important roles in
regulating tumor development. Deregulation of miRs in bladder
cancer has been comprehensively reviewed (21), however, the function and mechanisms
of miR-335 in bladder tumorigenesis have remained to be
elucidated.
miR-335 is transcribed from its genomic region on
chromosome 7q32.2 and acts as a tumor suppressor in types of
various malignancy (22). Studies
have reported that miR-335 is downregulated in various cancer
types, including breast, prostate (23) and gastric cancers (15,24);
however, the function of miR-335 and its target genes in bladder
cancer have remained elusive. To the best of our knowledge, the
present study was the first to investigate the expression of
miR-335 in bladder cancer tissues and to assess the underlying
mechanisms. The expression of miR-335 was detected to be
significantly decreased in human bladder cancer tissues compared
with that in adjacent normal tissues. Furthermore, low expression
of miR-335 in bladder cancer was associated with a more aggressive
phenotype. Similar results were obtained by analysis of miR-335
expression in two bladder cancer cell lines and a normal bladder
epithelial cell line, which further strengthened the conclusion
that miR-335 was downregulated in bladder cancer.
A gain-of-function study was further performed in
the T24 bladder cancer cell line. Overexpression of miR-335 through
transfection with miR-335 mimics was observed to significantly
inhibit the proliferation of T24 cells, which was consistent with
the results of previous studies, which reported the
tumor-suppressive role of miR-335 in other cancer types (15,24).
In addition, a Transwell assay revealed a significant inhibition of
T24-cell migration following transfection with miR-335 mimics,
demonstrating that miR-335 impaired the migratory ability of
bladder cancer cells. Thus, downregulation of miR-335 may have a
critical function in bladder cancer development. The role of
miR-335 in bladder cancer remains to be confirmed in
vivo.
To further assess the mechanism by which miR-335
functions as a tumor suppressor in bladder cancer, the present
study examined the association between miR-335 and MAPK1 in bladder
cancer cells, revealing that miR-335 exerted its regulatory role in
bladder cancer cells by targeting MAPK1. MAPK is an intracellular
serine/threonine protein kinase which regulates cell proliferation,
differentiation, development and apoptosis. The MAPK signal pathway
consists of ERK, c-Jun N-terminal kinase (JNK)/stress-activated
protein kinase (SAPK), p38 and ERK5/BMK1 subgroup. Several studies
have suggested that JNK/SAPK and p38MAPK pathways are related to
cellular stress and apoptosis, and ERK pathway serves an important
role in cell proliferation and differentiation (25,26).
Furthermore, it was demonstrated that miR-335 inhibited bladder
cancer cell proliferation and migration via the MAPK1 pathway. The
MTT assay revealed a significant cell-growth inhibition following
transfection of T24 cells with MAPK1-targeted siRNA compared with
si-control transfection. Furthermore, the Transwell assay
demonstrated a significant inhibition of T24-cell migration
following transfection with siMAPK1_1 and siMAPK_2. These results
suggested that reduced miR-335 levels lead to elevated MAPK1
levels, which drives the progression of bladder cancer.
In conclusion, the present study suggested that
miR-335 is a potential tumor suppressor in bladder cancer. By
targeting MAPK1, miR-335 inhibits bladder cancer cell proliferation
and migratory potential bladder cancer cells. Therefore,
restoration of miR-335 may represent a promising therapeutic
strategy for bladder cancer.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
von der Maase H, Sengelov L, Roberts JT,
Ricci S, Dogliotti L, Oliver T, Moore MJ, Zimmermann A and Arning
M: Long-term survival results of a randomized trial comparing
gemcitabine plus cisplatin, with methotrexate, vinblastine,
doxorubicin, plus cisplatin in patients with bladder cancer. J Clin
Oncol. 23:4602–4608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shirodkar SP and Lokeshwar VB: Potential
new urinary markers in the early detection of bladder cancer. Curr
Opin Urol. 19:488–493. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar
|
|
6
|
Zhao Y, Samal E and Srivastava D: Serum
response factor regulates a muscle specific microRNA that targets
Hand2 during cardiogenesis. Nature. 436:214–220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cimmino A, Calin GA, Fabbri M, Iorio MV,
Ferracin M, Shimizu M, Wojcik SE, Aqeilan RI, Zupo S, Dono M, et
al: MiR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl
Acad Sci USA. 102:13944–13949. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sethi S, Ali S, Philip PA and Sarkar FH:
Clinical advances in molecular biomarkers for cancer diagnosis and
therapy. Int J Mol Sci. 14:14771–14784. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ling H, Fabbri M and Calin GA: MicroRNAs
and other non-coding RNAs as targets for anticancer drug
development. Nat Rev Drug Discov. 12:847–865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu Z, Xu T, Wang L, Wang X, Zhong S, Xu C
and Shen Z: MicroRNA-145 directly targets the insulin-like growth
factor receptor I in human bladder cancer cells. FEBS Lett.
588:3180–3185. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feng Y, Liu J, Kang Y, He Y, Liang B, Yang
P and Yu Z: MiR-19a acts as an oncogenic microRNA and is
up-regulated in bladder cancer. J Exp Clin Cancer Res. 33:672014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lei Y, Hu X, Li B, Peng M, Tong S, Zu X,
Wang Z, Qi L and Chen M: MiR-150 modulates cisplatin
chemosensitivity and invasiveness of muscle-invasive bladder cancer
cells via targeting PDCD4 in vitro. Med Sci Monit. 20:1850–1857.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inoguchi S, Seki N, Chiyomaru T, Ishihara
T, Matsushita R, Mataki H, Itesako T, Tatarano S, Yoshino H, Goto
Y, et al: Tumour-suppressive microRNA-24–1 inhibits cancer cell
proliferation through targeting FOXM1 in bladder cancer. FEBS Lett.
588:3170–3179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang X, Wu J, Lin Y, Zhu Y, Xu X, Liang Z,
Li S, Hu Z, Zheng X and Xie L: MicroRNA-320c inhibits tumorous
behaviors of bladder cancer by targeting Cyclin-dependent kinase 6.
J Exp Clin Cancer Res. 33:692014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi L, Jiang D, Sun G, Wan Y, Zhang S,
Zeng Y, Pan T and Wang Z: miR-335 promotes cell proliferation by
directly targeting Rb1 in meningiomas. J Neurooncol. 110:155–162.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu Y, Zhao F, Wang Z, Song Y, Luo Y, Zhang
X, Jiang L, Sun Z, Miao Z and Xu H: MicroRNA-335 acts as a
metastasis suppressor in gastric cancer by targeting Bcl-w and
specificity protein 1. Oncogene. 31:1398–1407. 2012. View Article : Google Scholar :
|
|
17
|
Yan J, Jiang N, Huang G, Tay JL, Lin B, Bi
C, Koh GS, Li Z, Tan J, Chung TH, et al: Deregulated MIR335 that
targets MAPK1 is implicated in poor outcome of paediatric acute
lymphoblastic leukaemia. Br J Haematol. 163:93–103. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
19
|
Murali R, Chandramohan R, Möller I, Scholz
SL, Berger M, Huberman K, Viale A, Pirun M, Socci ND, Bouvier N, et
al: Targeted massively parallel sequencing of angiosarcomas reveals
frequent activation of the mitogen activated protein kinase
pathway. Oncotarget. 6:36041–36052. 2015.PubMed/NCBI
|
|
20
|
Esquela-Kerscher A and Slack FJ: Oncomirs-
microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Catto JW, Alcaraz A, Bjartell AS, De Vere
White R, Evans CP, Fussel S, Hamdy FC, Kallioniemi O, Mengual L,
Schlomm T and Visakorpi T: MicroRNA in prostate, bladder, and
kidney cancer: A systematic review. Eur Urol. 59:671–681. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tavazoie SF, Alarcón C, Oskarsson T, Padua
D, Wang Q, Bos PD, Gerald WL and Massagué J: Endogenous human
microRNAs that suppress breast cancer metastasis. Nature.
451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Q, Liu X, Liu Y, Yang J, Lv G and Dong
S: MicroRNA-335 and -543 suppress bone metastasis in prostate
cancer via targeting endothelial nitric oxide synthase. Int J Mol
Med. 36:1417–1425. 2015.PubMed/NCBI
|
|
24
|
Yan Z, Xiong Y, Xu W, Gao J, Cheng Y, Wang
Z, Chen F and Zheng G: Identification of hsa-miR-335 as a
prognostic signature in gastric cancer. PLoS One. 7:e400372012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kamiyama M, Naguro I and Ichijo H: In vivo
gene manipulation reveals the impact of stress-responsive MAPK
pathways on tumor progression. Cancer Sci. 106:785–796. 2015.
View Article : Google Scholar : PubMed/NCBI
|