Introduction
Skeletal modeling is a physiological process
combining bone formation with bone resorption, where osteoclasts,
derived from hematopoietic mononuclear macrophages, are involved in
bone resorption. Under the stimulation of receptor activator for
nuclear factor-κb ligand (RANKL) and macrophage colony stimulating
factor, mononuclear macrophages fuse to form osteoclasts and
express osteoclast markers, including calcitonin receptor (CTR) and
RANK, with the activation of tartrate-resistant acid phosphatase
(TRAP) for bone resorption (1,2). As
previously reported, osteoclast differentiation is regulated by
several stimulators, including several types of growth factors
present in the bone microenvironment (3). It is of clinical significance to
investigate the regulatory mechanism of osteoclast differentiation,
which may contribute to the development of novel drugs for
inhibiting bone resorption.
Bone morphogenetic proteins (BMPs) are
multi-functional growth factors in the transforming growth factor-β
superfamily, which is associated with numerous molecular cascades
and signaling pathways (4). At
present, 18 different BMP-homologous molecules have been
identified, and have been shown to induce effects via the small
mothers against decapentaplegic (Smad) and mitogen-activated
protein kinase pathways through binding to different types of
receptors, including BMP receptor (BMPR)-I, BMPR-II and anaplastic
lymphoma kinases (ALKs) (4,5). A
number of previous studies have confirmed the effect of BMPs on
osteogenesis, including the differentiation of preosteoblasts and
bone matrix synthesis (5–8). The effects of BMPs on
osteoclastogenesis and bone resorption have also attracted
scientific interest, however, the effects remain to be fully
elucidated (1). It has been
suggested that BMP-9 may promote the differentiation of
osteoblasts, however, the effects of BMP-9 on the differentiation
of osteoclasts and bone resorption remain to be fully elucidated
(7,9). In the present study, mouse spleen
macrophages (RAW 264.7 cells) were cultured as osteoclast
precursors to investigate the roles of BMP-9 on osteoclast
proliferation and differentiation, and to examine the signaling
mechanism involved. The aim of the present study was to determine
the potential impact of BMP-9 on bone resorption in bone
homeostasis.
Materials and methods
Cell culture
Mouse spleen macrophages were purchased from the
Cell Bank of Chinese Academy of Sciences (Shanghai, China) for use
as osteoclast precursors. These cells were cultured in Dulbecco's
minimum essential medium (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Thermo
Fisher Scientific, Inc.) at 37°C in a humidified incubator
containing 5% CO2, and the medium was replaced every 3
days. The exponentially growing cells were seeded at a density of
2×104 cells/well in 48-well plates for the examinations of cell
proliferation, TRAP-positive cell staining and immunofluorescence
staining, or at a density of 2×105 cells/well in six-well plates
for enzyme-linked immunosorbent assay (ELISA) and western blot
analysis. Soluble recombinant human (rh)RANKL (100 ng/ml;
PeproTech, Inc., Rocky Hill, NJ, USA) was added to stimulate
osteoclastogenesis of the preosteoclasts. After 24 h of incubation,
different concentrations (0–150 ng/ml) of rhBMPs (R&D Systems,
Inc., Minneapolis, MN, USA) were added to the cultures. The medium,
BMPs and RANKL were replaced every 3 days. Triplicate cell cultures
were performed for the gene experiment, and quadruple cell cultures
were performed for the other experiments. Experiments were repeated
twice to confirm the results.
Cell proliferation assay
An MTT assay was performed to evaluate cell
viability, as previously described (2). Cells (8×103) were seeded
onto 96-well plates 24 h prior to treatment with ALK1-siRNA for 48
h using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h, 15 µl (5 mg/ml) MTT (Sigma-Aldrich;
Merck Millipore, Darmstadt, Germany) was added to each well, and
the cells were incubated for a further 4 h at 37°C. Subsequently,
150 µl dimethyl sulfoxide was added and the mixtures were agitated
for 10–15 min to fully dissolve the crystals. Absorbance (A) was
measured at 570 nm using a Tecan microplate reader (Männedorf,
Switzerland). Cell viability was calculated as the percentage
change in A570 between the control and treated cells.
TRAP staining of cells
After 72 h, the cells were fixed with 4%
paraformaldehyde for 10 min, washed with PBS, and then incubated in
TRAP staining (0.05 M acetate buffer, 0.03 M sodium tartrate, 100
g/ml naphthol AS-MX phosphate, 0.01% Triton X-100 and 0.3 mg/ml
Fast Red Violet LB stain) for 10 mins at 37°C. A phase contrast
microscope was used to observe and calculate the number of
osteoclasts.
ELISA
The protein expression of BMPR-IA, BMPR-IB, BMPR-II,
ALK1 receptors and CTR on the cell surface were detected using
ELISA (Biomedical Technologies, Inc., Stoughton, MA, USA) according
to the kit instructions. The results were standardized by the
quality of glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Immunofluorescence detection
The cells were fixed with 4% paraformaldehyde and
blocked with 5% BSA, and were then incubated overnight at 4°C with
the following primary antibody: Polyclonal goat anti-mouse ALK1
immunoglobulin (Ig) G (1:100; cat. no. 770-MA; R&D Systems,
Inc., Minneapolis, MN, USA), quenched with glycine (0.1 M) for 1 h,
incubated with secondary antibodies (polyclonal donkey anti-goat
IgG; 1:200; cat. no. NL003; R&D Systems, Inc.) for 2 h at room
temperature, and then washed three times with PBS. The cells were
then counterstained with DAPI (Vector Laboratories Canada, Inc.,
Burlington, ON, Canada), and the slides mounted in PBS-glycerol 50%
(v/v). The detection of fluorescence intensity was evaluated using
fluorescence microscopy.
Western blot analysis
At the end of the culture period, the cells were
washed with cold PBS, and lysed in the presence of protease
inhibitors. Protein concentrations were determined using the
Bio-Rad protein assay kit (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Total proteins (50 µg) were separated by 10% SDS-PAGE,
and transferred onto a PVDF membrane (EMD Millipore, Billerica, MA,
USA). The membrane was blocked with 5% BSA in Tris-buffered saline
Tween-20 for 1 h at room temperature. For visualization, the blots
were probed with antibodies against Smad2 (1:1,000 dilution; cat.
no. 3103), phosphorylated (p)-Smad2 (1:1,000 dilution; cat. no.
3104), Erk1/2 (1:500 dilution; cat. no. 4696), p-Erk1/2 (1:500
dilution; cat. no. 9101) and GAPDH (1:1,000 dilution; cat. no.
97166) (all Cell Signaling Technology, Inc., Danvers, MA, USA)
overnight at 4°C. Horseradish peroxidase-conjugated goat
anti-rabbit (cat. no. 7074) and horse anti-mouse (cat. no. 7076)
secondary antibodies (1:1,000; Cell Signaling Technology, Inc.)
were used to enable detection using a chemiluminescent system at
room temperature. The band densities were quantified using a
Bio-Rad Versa Doc imaging system (Bio-Rad Laboratories, Inc.), and
normalized to the GAPDH band as an internal control for each
group.
RNA inactivation
The HP custom siRNA kit (cat. no. 1027423) was
obtained from Qiagen (Mississauga, ON, Canada); the negative
control (NC) sequence was included. A total of 48 h prior to
stimulation, the differentiated cells were transfected either with
a mouse ALK1 siRNA (NM_001277255.1, 5′-GCAGGAAATCTCACCACAT-3′,
3′-AUGUGGUGAGAUUUCCUGCTT-5′; 1 ng/ml; Qiagen) or with an NC siRNA
(5′-GCAAATATCACCCAGGCAT-3′, 3′-AUGCCUGGGUGAUAUUUGCTT-5′; 1 ng/ml;
Qiagen) in a solution containing Lipofectamine® 2000
transfection reagent in serum-free Opti-MEM. The cells were
incubated at 37°C for 2 h with gentle stirring. The transfection
efficiency was evaluated using fluorescence microscopy in one well
of the culture containing a control green fluorescent protein
siRNA, and the subsequent downregulation of ALK1 was assessed using
western blotting.
Statistical analysis
Data are expressed as the mean ± standard deviation
of three or six independent experiments, and the statistical
significance was determined using a paired Student's t-test
or analysis of variance. Statistical analyses were conducted using
SPSS 19.0 software (SPSS, Inc., Chicagi, IL, USA) and graphs were
generated using GraphPad Prism 5.0 (GraphPad Software, Inc., La
Jolla, CA, USA). P<0.01 was considered to indicate a
statistically significant difference.
Results
Effects of BMP-9 on osteoclast
proliferation and differentiation
The MTT assay showed that RANKL (100 ng/ml) and
BMP-9 (50–150 ng/ml) alone promoted cell proliferation (Fig. 1A). The TRAP staining of the cells
showed that RANKL (100 ng/ml) stimulated the activation of TRAP in
the preosteoclasts (Fig. 1B and
Ca). However, BMP-9 (50–150 ng/ml) failed to induce the
activation of TRAP in the cells without the presence of RANKL
(Fig. 1Cb). In the presence of
RANKL (100 ng/ml), the percentage of TRAP-positive cells was
significantly elevated by BMP-9 (50–150 ng/ml) in a dose-dependent
manner (Fig. 1B and Cc and d). The
effect of BMP-9 on the activation of TRAP in the cells was not
associated with its effect on cell proliferation. In addition, the
expression of the CTR marker in the cell, detected using ELISA
showed a similar trend (Fig. 1D).
This suggested that BMP-9 promoted RANKL-induced osteoclast
differentiation in a dose-dependent manner at certain
concentrations (50–150 ng/ml).
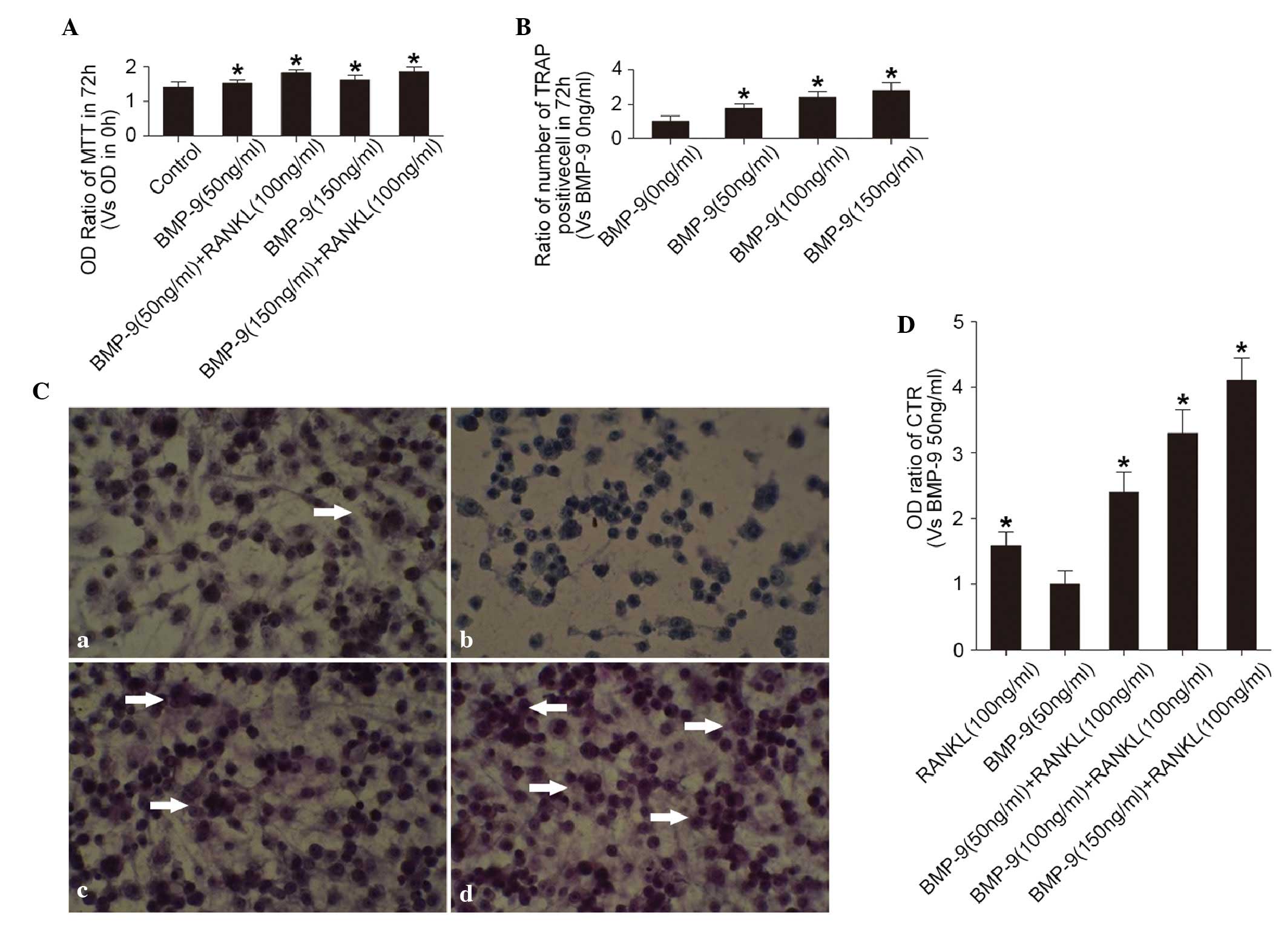 | Figure 1.Effects of BMP-9 on osteoclast
proliferation and differentiation. (A) Proliferation of cells in
different treatment groups cultured for 72 h, determined using an
MTT assay. OD ratio = OD (0 h) / OD (72 h). *P<0.01, vs. control
group. (B) Percentage of TRAP-positive cells in different treatment
groups cultured for 72 h. *P<0.01, vs. BMP-9 (0 ng/ml) group.
(C) TRAP staining of cells treated with (a) 100 ng/ml RANKL without
BMP-9, (b) 50 ng/ml BMP-9 without RANKL, (c) 50 ng/ml BMP-9 with
RANKL (100 ng/ml), (d) 150 ng/ml BMP-9 with RANKL (100 ng/ml). A
phase contrast microscope was used (magnification, ×20).
TRAP-positive cells are indicated by white arrows. (D) Expression
of CTR, assessed using an enzyme-linked immunosorbent assay,
following treatment with different concentrations of BMP-9 with or
without RANKL. *P<0.01, vs. BMP-9 (50 ng/ml) group. BMP-9, bone
morphogenetic protein-9; RANKL, receptor activator for nuclear
factor-κb ligand; CTR, calcitonin receptor; OD, optical density;
TRAP, tartrate-resistant acid phosphatase. |
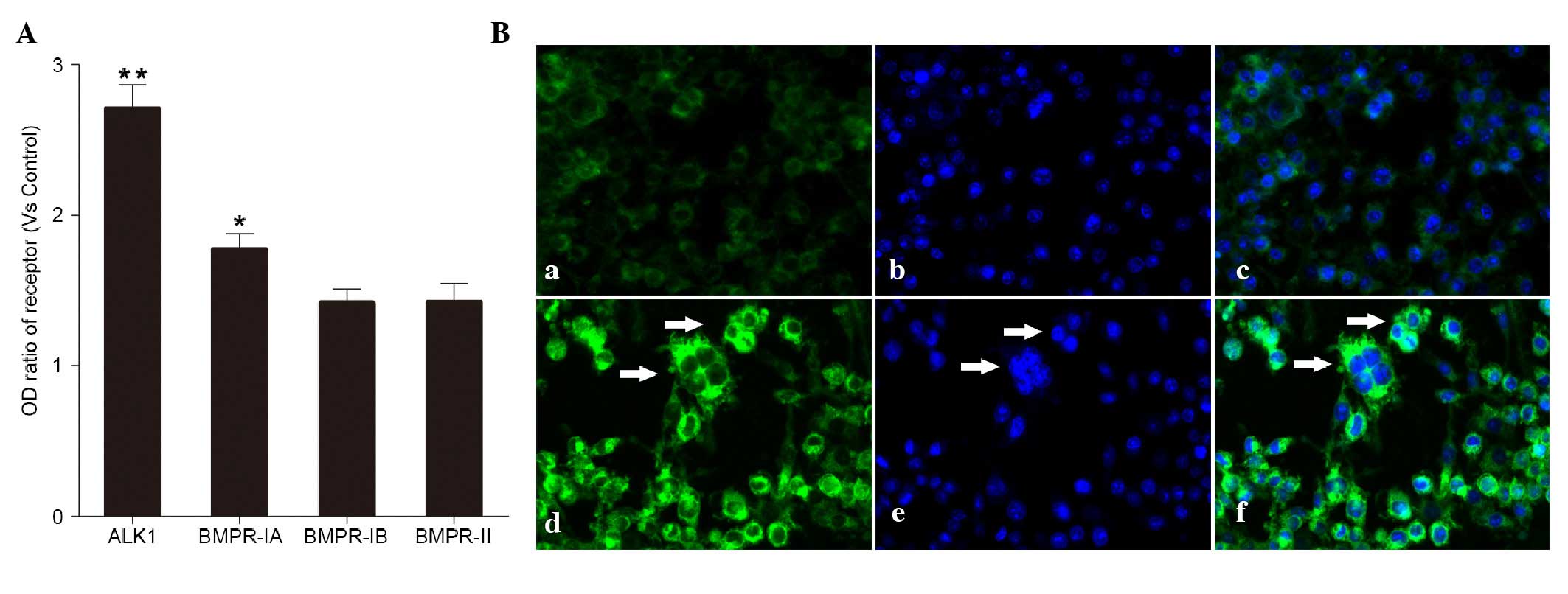
Effects of BMP-9 on the expression of
BMP receptors
The BMPR-IA, BMPR-IB, BMPR-II and ALK1 receptors
were detected in the cells induced by BMP-9 (100 ng/ml) using
ELISA. Among these receptors, the ALK1 receptor showed the highest
level of upregulation (Fig. 2A).
According to previous reports (10–12),
BMP-9 preferentially bind to its ALK1 receptor. Therefore, in the
present study, the ALK1 receptor was selected and its expression on
the cell surface was confirmed using immunofluorescence (Fig. 2Ba-f).
Effects of BMP-9 and the ALK1 receptor
on the induction of the cell signal transduction pathway
The Smad2 and non-Smad EPK1/2 signaling pathways are
the most common pathways in cells induced by BMP-9 (4,5). The
results of the western blot analysis showed that the
phosphorylation of ERK1/2 (p-ERK1/2) was decreased in the cells
following the induction of BMP-9, and was significantly reduced at
60 and 120 min, respectively (Fig. 3A
and B). However, no change was observed in the phosphorylation
of Smad2 during induction for 0–120 min (Fig. 3C and D). These results showed that
BMP-9 inhibited the ERK1/2 pathway in the osteoclast precursor.
Furthermore, the inhibited phosphorylation of ERK1/2 was reduced in
the cells pre-transfected with siRNA-ALK1 (Fig. 3E and F). This suggested that BMP-9
inhibited the phosphorylation of ERK1/2 through the ALK1 receptor
in the cells.
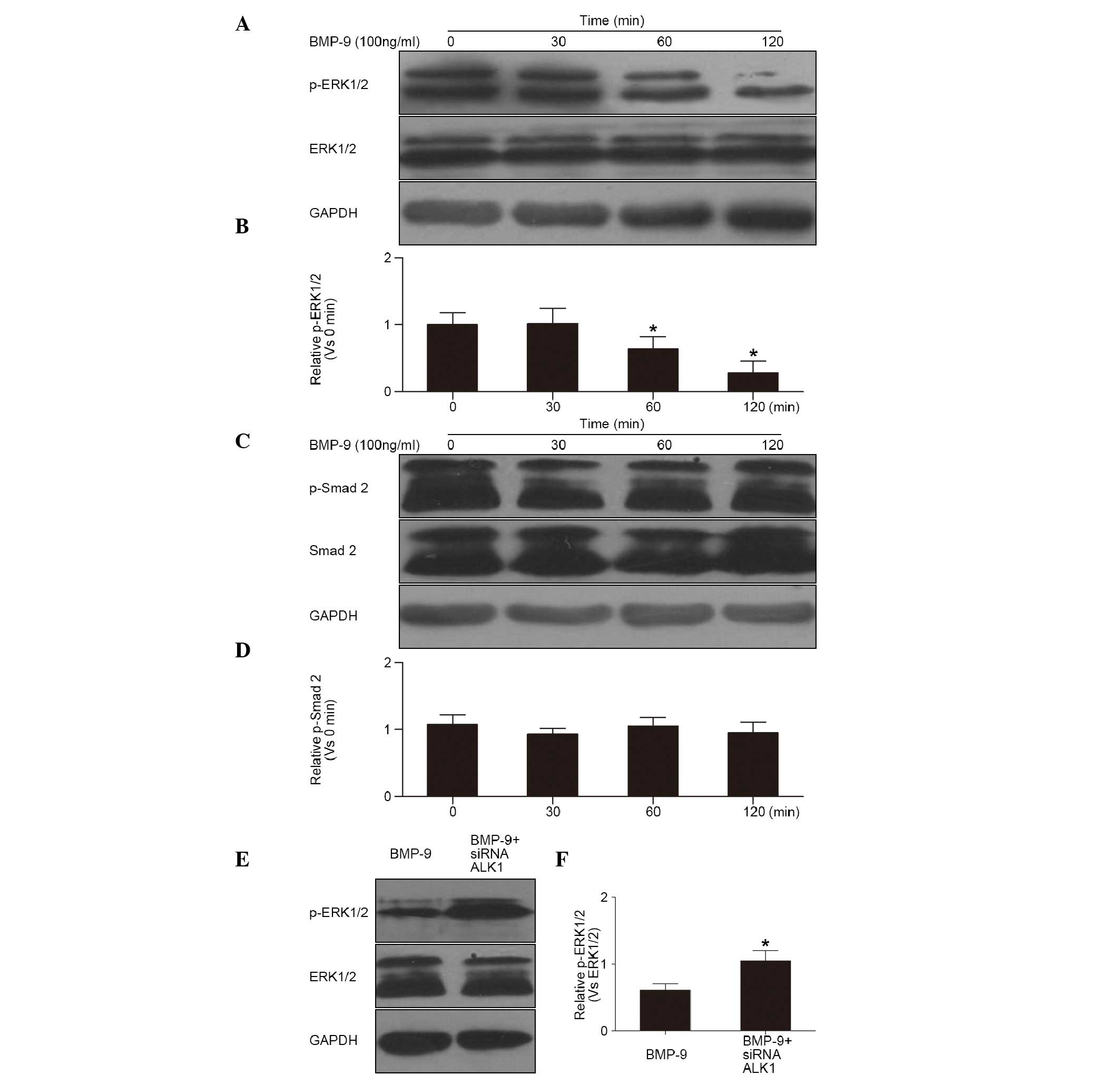 | Figure 3.Effects of BMP-9 and ALK1 receptor on
the cell signal transduction pathway. (A) Western blot of the
phosphorylation of ERK1/2 in cells treated with BMP-9 (100 ng/ml)
and (B) quantification (*P<0.01, vs. 0 min). (C) Western blot of
the phosphorylation of Smad2 in cells treated with BMP-9 (100
ng/ml) and (D) quantification (P>0.01, vs. 0 min). (E) Western
blot of the phosphorylation of ERK1/2 in cells pre-treated with
siRNA-ALK1 and (F) quantification (*P<0.01, vs. BMP-9). ERK1/2,
extracellular signal-regulated kinase 1/2; p-ERK1/2, phosphorylated
ERK1/2; Smad 2, small mothers against decapentaplegic 2; p-Smad2,
phosphorylated Smad 2; BMP-9, bone morphogenetic protein-9; ALK1,
anaplastic lymphoma kinase 1; siRNA, small interfering RNA. |
Effects of the ALK1 receptor and
ERK1/2 pathways on BMP-9-induced differentiation of
osteoclasts
In the presence of RANKL, BMP-9 (100 ng/ml) induced
the activation of TRAP in the cells Fig. 4Aa. The percentage of TRAP-positive
cells was reduced in the cells pre-transfected with siRNA-ALK1
Fig. 4Ab), but increased in the
cells exposed to U0126 (1,000 nmol/l), an inhibitor of the ERK1/2
pathway (Fig. 4Ac). These results
suggested that the promotion of BMP-9 on osteoclast differentiation
was reduced by inhibition of the ALK1 receptor, but enhanced by
inhibition of the ERK1/2 pathways. In addition, the protein
expression of CTR in the cells, determined using ELISA, showed
similar results (Fig. 4B).
Therefore, BMP-9 inhibited the intracellular ERK1/2 pathways
through mediation of the ALK1 receptor on the surface of
macrophagocytes, further promoting the differentiation into
osteoclasts.
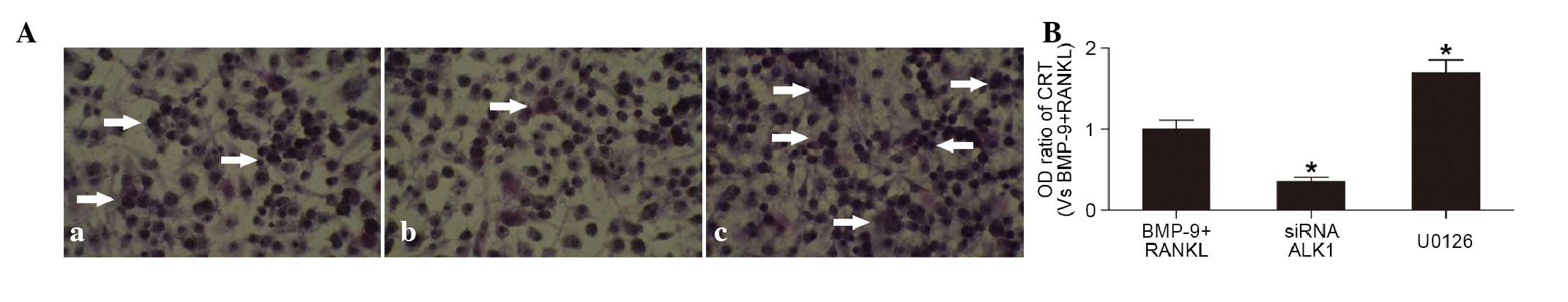 | Figure 4.Effects of the ALK1 receptor and
ERK1/2 pathways on BMP-9-induced osteoclast differentiation (A)
TRAP staining of cells (a) induced by BMP-9 (100 ng/ml) and RANKL
(100 ng/ml), and (b) pre-transfected with siRNA-ALK1 or (c)
cultured with U0126 (1,000 nmol/l) prior to BMP-9+RANKL. (Phase
contrast microscope; magnification, ×20). White arrows indicate
TRAP-positive cells. (B) Protein expression of CTR, determined
using an enzyme-linked immunosorbent assay, in cells treated with
BMP-9 (100 ng/ml) and RANKL (100 ng/ml), transfectecd with
siRNA-ALK1 or treated with U0126 (1,000 nmol/l). *P<0.01, vs.
BMP-9+RANKL group. BMP-9, bone morphogenetic protein-9; ALK1,
anaplastic lymphoma kinase 1; RANKL, receptor activator for nuclear
factor-κb ligand; CTR, calcitonin receptor; siRNA, small
interfering RNA; OD, optical density; TRAP, tartrate-resistant acid
phosphatase. |
Discussion
The present in vitro study showed that BMP
receptors, including BMPR-IA, BMPR-IB, BMPR-II and ALK1, were
detected in mouse spleen macrophages, and that BMP-9 promoted the
proliferation and osteoclast differentiation of the cells in the
presence of RANKL. The possible mechanism underlying the promotion
of differentiation by BMP-9 may be that BMP-9 inhibited the
intracellular EPK1/2 pathways through binding to ALK1 receptors on
the cell surface.
Previous studies have confirmed that BMPs, including
BMP-2, 4 and 7, not only promote osteogenisis differentiation, but
are also important in regulating osteoclast differentiation
(6,13). Due to its importance in
osteogenesis, BMP-9 is considered to be a growth factor offering
significant potential in clinical practice. However, there are few
reports on the roles of BMP-9 in osteoclast differentiation and
bone resorption. The effects of BMP-9 on osteoclast differentiation
were confirmed by the results of the present study, which also
offered novel clues to its possible mechanism. A previous study by
Fong et al (14) suggested
that BMP-9 did not promote osteoclast differentiation in human
mononuclear macrophages, however, it enhanced bone resorption by
significantly inhibiting the apoptosis of mononuclear macrophages
in the presence of RANKL. The findings of the present study showed
that BMP-9 promoted the proliferation of mouse spleen mononuclear
macrophages, and enhanced osteoclast differentiation only in the
presence of RANKL. These findings show the direct effects of BMP-9
on osteoclast precursors and bone resorption. However, the effect
of BMP-9 on cell differentiation was not associated with its effect
on cell proliferation, which suggested that BMP-9 may have
different effects on mononuclear macrophages from different
sources. In mononuclear macrophages in the mouse spleen, a study by
Zheng et al (15)
demonstrated that the BMP2/7 heterodimer promoted proliferation and
osteoclast differentiation in a dose-dependent manner in the
presence of RANKL in vitro. These findings suggest that BMP
factors have effects on osteoclast differentiation with diversity
according to the different osteoclast precursors.
Various types of BMP receptors, including BMPR-IA,
BMPR-IB, BMPR-II and ALK1,2, have been found in osteoclast
precursors from different sources. BMP-9 is the physiological
ligand of ALK1,2 in various cells (12,16),
and it has also been reported that BMP-9 binds to its BMPR-II
receptor in osteoclast precursors (12). In the present study, the expression
levels of BMPR-IA, BMPR-IB, BMPR-II and ALK1 receptors were
detected in the cells induced by BMP-9. Among these receptors, the
expression of ALK1 was upregulated significantly. Using an siRNA
inactivation assay, the ALK1 receptor was found to be important in
the effect of osteoclast differentiation by BMP-9. It is important
to acknowledge that the involvement of the other receptors on the
effects of BMP-9 cannot be excluded. The Smad2 and Smad-independent
ERK1/2 pathways are the primary signaling pathways in cells induced
by BMP-9 (16–18). The findings from the present study
showed that BMP-9 inhibited the phosphorylation of ERK1/2 pathways.
However, a study by Fong et al (14) indicated that BMP-9 promoted the
ERK1/2 pathways, suggesting that the binding of BMP-9 to different
receptors may have different effects.
There were two limitations to the present study: i)
As BMP-9 is similar to other BMPs, it may bind to different
receptors and activate, or inactivate, different signaling pathways
in different cell types. In the present study, only the ALK1
receptor and EPK1/2 signaling pathways were investigated, as
investigated in previous studies (4,5,7,16,19,20),
and their effects on the impact of BMP-9 were confirmed. However,
whether another receptor or signaling pathway is involved in this
signaling mechanism remains to be elucidated. Therefore,
considering the complexity of the signaling mechanism by BMPs,
further investigations are required on other receptors or signaling
pathways; ii) As in vitro results are not necessarily
consistent with the effects of BMP-9 in vivo, there is a
requirement to perform further in vivo investigations.
In conclusion, the presents study confirmed that
BMP-9 promoted the proliferation and differentiation of osteoclast
precursors in the presence of RANKL, which involved the ALK1
receptor and ERK1/2 pathways. These findings expand on current
understanding of the effects of BMPs on the regulation of
osteoclast differentiation and bone resorption, and provide
experimental evidence for further in vivo
investigations.
Acknowledgements
This study was supported by the Science Foundation
of Shanghai Science and Technology Commission (grant no.
11ZR1423900).
Glossary
Abbreviations
Abbreviations:
|
BMP-9
|
bone morphogenetic protein-9
|
|
RANKL
|
receptor activator for nuclear
factor-κb ligand
|
|
BMPR
|
bone morphogenetic protein
receptor
|
|
ALK1
|
anaplastic lymphoma kinase 1
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
CTR
|
calcitonin receptor
|
|
TRAP
|
tartrate-resistant acid
phosphatase
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
References
|
1
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parfitt AM, Mundy GR, Roodman GD, Hughes
DE and Boyce BF: A new model for the regulation of bone resorption,
with particular reference to the effects of bisphosphonates. J Bone
Miner Res. 11:150–159. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Graves L III and Jilka RL: Comparison of
bone and parathyroid hormone as stimulators of osteoclast
development and activity in calvarial cell cultures from normal and
osteopetrotic (mi/mi) mice. J Cell Physiol. 145:102–109. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bragdon B, Moseychuk O, Saldanha S, King
D, Julian J and Nohe A: Bone morphogenetic proteins: A critical
review. Cell Signal. 23:609–620. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schmierer B and Hill CS: TGFbeta-SMAD
signal transduction: Molecular specificity and functional
flexibility. Nat Rev Mol Cell Biol. 8:970–982. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lavery K, Swain P, Falb D and
Alaoui-Ismaili MH: BMP-2/4 and BMP-6/7 differentially utilize cell
surface receptors to induce osteoblastic differentiation of human
bone marrow-derived mesenchymal stem cells. J Biol Chem.
283:20948–20958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luu HH, Song WX, Luo X, Manning D, Luo J,
Deng ZL, Sharff KA, Montag AG, Haydon RC and He TC: Distinct roles
of bone morphogenetic proteins in osteogenic differentiation of
mesenchymal stem cells. J Orthop Res. 25:665–677. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Senta H, Park H, Bergeron E, Drevelle O,
Fong D, Leblanc E, Cabana F, Roux S, Grenier G and Faucheux N: Cell
responses to bone morphogenetic proteins and peptides derived from
them: Biomedical applications and limitations. Cytokine Growth
Factor Rev. 20:213–222. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bergeron E, Leblanc E, Drevelle O, Giguère
R, Beauvais S, Grenier G and Faucheux N: The evaluation of ectopic
bone formation induced by delivery systems for bone morphogenetic
protein-9 or its derived peptide. Tissue Eng Part A. 18:342–352.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bergeron E, Senta H, Mailloux A, Park H,
Lord E and Faucheux N: Murine preosteoblast differentiation induced
by a peptide derived from bone morphogenetic proteins-9. Tissue Eng
Part A. 15:3341–3349. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cunha SI and Pietras K: ALK1 as an
emerging target for antiangiogenic therapy of cancer. Blood.
117:6999–7006. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Scharpfenecker M, van Dinther M, Liu Z,
van Bezooijen RL, Zhao Q, Pukac L, Löwik CW and ten Dijke P: BMP-9
signals via ALK1 and inhibits bFGF-induced endothelial cell
proliferation and VEGF-stimulated angiogenesis. J Cell Sci.
120:964–972. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
David L, Feige JJ and Bailly S: Emerging
role of bone morphogenetic proteins in angiogenesis. Cytokine
Growth Factor Rev. 20:203–212. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fong D, Bisson M, Laberge G, McManus S,
Grenier G, Faucheux N and Roux S: Bone morphogenetic protein-9
activates Smad and ERK pathways and supports human osteoclast
function and survival in vitro. Cell Signal. 25:717–728. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng Y, Wang L, Zhang X, Zhang X, Gu Z
and Wu G: BMP2/7 heterodimer can modulate all cellular events of
the in vitro RANKL-mediated osteoclastogenesis, respectively, in
different dose patterns. Tissue Eng Part A. 18:621–630. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Broege A, Pham L, Jensen ED, Emery A,
Huang TH, Stemig M, Beppu H, Petryk A, O'Connor M, Mansky K and
Gopalakrishnan R: Bone morphogenetic proteins signal via SMAD and
mitogen-activated protein (MAP) kinase pathways at distinct times
during osteoclastogenesis. J Biol Chem. 288:37230–37240. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Itoh K, Udagawa N, Katagiri T, Iemura S,
Ueno N, Yasuda H, Higashio K, Quinn JM, Gillespie MT, Martin TJ, et
al: Bone morphogenetic protein 2 stimulates osteoclast
differentiation and survival supported by receptor activator of
nuclear factor-kappaB ligand. Endocrinology. 142:3656–3662. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jensen ED, Pham L, Billington CJ Jr, Espe
K, Carlson AE, Westendorf JJ, Petryk A, Gopalakrishnan R and Mansky
K: Bone morphogenic protein 2 directly enhances differentiation of
murine osteoclast precursors. J Cell Biochem. 109:672–682.
2010.PubMed/NCBI
|
|
19
|
Kaneko H, Arakawa T, Mano H, Kaneda T,
Ogasawara A, Nakagawa M, Toyama Y, Yabe Y, Kumegawa M and Hakeda Y:
Direct stimulation of osteoclastic bone resorption by bone
morphogenetic protein (BMP)-2 and expression of BMP receptors in
mature osteoclasts. Bone. 27:479–486. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie F, Liu W, Feng F, Li X, Yang L, Lv D,
Qin X, Li L and Chen L: A static pressure sensitive receptor APJ
promote H9c2 cardiomyocyte hypertrophy via PI3K-autophagy pathway.
Acta Biochim Biophys Sin (Shanghai). 46:699–708. 2014. View Article : Google Scholar : PubMed/NCBI
|