Introduction
Malignant melanoma (MM) originates from melanocytes
and is a highly malignant skin tumor, which accounts for 1–2% of
all skin cancer cases. MM has a high malignant potential, with a
high risk of ulceration and metastasis. The global incidence rates
of MM have increased rapidly over the past few decades, with an
annual incidence rate of 3–8% (1–3).
The Y-box binding protein-1 (YB-1), which contains a
‘cold shock’ region, is a multifunctional protein that is located
in the nucleus and the cytoplasm (4). YB-1 regulates the expression of
target genes indirectly or by binding to target gene sequences
directly, and the altering the splicing of target genes (5–8).
YB-1 expression is upregulated in a number of human tumors,
including ovarian cancer, breast cancer, glioblastoma, esophageal
cancer, gastric cancer, colon cancer and lung cancer (9–11),
and is therefore considered to be a prognostic marker. YB-1
overexpression is an indicator of increased tumor cell
proliferation, migration and invasion (9–11).
Additional studies have indicated that the expression levels of
YB-1, the epidermal growth factor receptor (EGFR) and the human
EGFR-2 (HER-2) are collectively increased in breast cancer tissues.
In addition, HER-2 may regulate the nuclear translocation of YB-1
directly or indirectly (11). In
contrast to its putative oncogenic role, inhibition of YB-1
expression has been demonstrated to increase the sensitivity of
melanoma cells to chemotherapy; an effect that is closely
associated with the phosphatidylinositol-4,5-bisphosphate
3-kinase/AKT serine/threonine kinase 1 signaling pathway (11,12).
This demonstrates that the expression of YB-1 may serve a role in
regulating tumor development and/or progression.
Advances in tumor biology have led to an increased
focus on the potential antitumor activities of antimicrobial
peptides. The human cathelicidin antimicrobial protein-18, also
known as LL-37, is a key antimicrobial molecule that has been
demonstrated to exhibit antitumor activities (13). It is the only member of the
cathelin family in humans, and is composed of 37 amino acid
residues in the C-terminal domain. LL-37 has been demonstrated to
serve an important functional role in regulating innate immunity
via neutrophils (13–15). In addition to its antibacterial and
immune activities, LL-37 is reportedly involved in angiogenesis and
cell proliferation (14,16). Studies have demonstrated that LL-37
expression is upregulated in ovarian, breast, prostate and lung
cancer, which may promote tumor cell proliferation, migration and
invasion (17–22). The oncogenic mechanisms by which
LL-37 functions may be associated with the formyl peptide receptor
(FPR)-like 1, EGFR and insulin-like growth factor 1 receptor;
however, the specific mechanisms remain unclear (15,23–25).
In addition, the role of LL-37 in regulating YB-1 expression has
not been elucidated.
In the present study, the effects of LL-37 on YB-1
expression on the proliferation, migration and invasion of MM cells
were investigated. The results demonstrated that LL-37 may promote
the progression of MM cells by upregulating YB-1 expression via the
nuclear factor-κB (NF-κB) signaling pathway.
Materials and methods
Cell culture
MM cell lines (A375 and A875; American Type Culture
Collection, Manassas, VA, USA) were cultured in Dulbecco's modified
Eagle's medium (Gibco; Thermo Fisher Scientific, Inc., Karlsruhe,
Germany) containing 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin (Sigma-Aldrich; Merck Millipore, Darmstadt, Germany).
Cells were maintained in a humidified chamber at 37°C and 5%
CO2.
Knockdown of YB-1 by small interfering
RNA (siRNA)
The YB-1 siRNA and negative control (NC) siRNA
oligonucleotide sequences (Shanghai GenePharma Co., Ltd., Shanghai,
China) employed in the present study were as follows: YB-1 siRNA 1,
sense 5′-GCAGACCGUAACCAUUAUATT−3′, antisense
5′-UAUAAUGGUUACGGUCUGCTT-3′; YB-1 siRNA 2, sense
5′-CGGCAAUGAAGAAGAUAAATT-3′, antisense 5′-UUUAUCUUCUUCAUUGCCGTT-3′;
YB-1 siRNA 3, 5′-CUGCCAUAAAGAAGAAUAATT-3′, antisense
5′-UUAUUCUUCUUUAUGGCAGTT-3′; and NC siRNA, sense
5′-UUCUUCGAACGUGUCACGUTT-3′ and antisense
5′-ACGUGACACGUUCGGAGAATT-3′. According to a GenBank (https://www.ncbi.nlm.nih.gov/genbank/)
sequence database search, the NC siRNA used in the present study
was not demonstrated to target any known mammalian gene. Cells were
seeded at a density of 1×105 cells/well in 6-well
plates. When 70–80% confluence was reached (~24 h after seeding),
the cells were transfected with siRNAs using the Lipofectamine 2000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Transfected cells
were incubated for a further 48 h, before the inhibition rate of
YB-1 expression was determined by western blot analysis.
Western blot analysis
At 4°C, ~2×106 cells were lysed with
100–400 µl of a mixture comprising PBS, 5 mM EDTA, 0.5% Triton
X-100, 20 mM NaF, 1 mM orthovanadate, 1 mM pyrophosphate and
protease inhibitors (0.1 mM PMSF, 10 µM pepstatin A, 10 µM
leupeptin and 25 µg/ml aprotinin) for 30 min, and then the cell
lysates were centrifuged (13,523 × g, 4°C, 15 min). Protein
extracts (20 µg) were separated using 10% SDS-PAGE and then
transferred to a polyvinylidene difluoride membrane. The membrane
was blocked using 5% non-fat milk in a 0.1% Tween-20-phosphate
buffer solution and incubated at room temperature for 1 h.
Membranes were subsequently incubated with the rabbit anti-human
YB-1 monoclonal antibody (1:500 dilution; cat. no. ab12148; Abcam,
Cambridge, UK) and β-actin antibody (cat. no. sc-47778; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) at 4°C overnight. The
membranes were then incubated with a goat anti-rabbit horseradish
peroxidase-conjugated secondary antibody (1:5,000 dilution; cat.
no. sc-2004; Santa Cruz Biotechnology, Inc.) for 1 h at room
temperature, and visualized using a chemiluminescence substrate
(EMD Millipore, Billerica, MA, USA). Chemiluminescence was detected
using an X-ray film.
Analysis of the viability and
migration of cells transfected with YB-1 siRNA
Transfected cells were seeded at a density of
3×103 cells/well in 96-well plates. Following 12 h, the
medium was replaced with serum-free medium and the cells were
cultured for a further 24 h. A total of 10 µl (5 mg/ml) MTT reagent
(Sigma-Aldrich; Merck Millipore) was added to each well, and cells
were incubated under normal conditions for a further 4 h. Following
the addition of 150 µl dimethyl sulfoxide (Sigma-Aldrich; Merck
Millipore), the absorbance was read at 490 nm using a microplate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA). A cell
invasion assay was performed using Transwell chambers (Costar;
Corning Life Sciences, Inc., Corning, NY, USA) pre-coated with
Matrigel matrix (BD Biosciences, Heidelberg, Germany). Briefly,
transfected cells were incubated without serum for 12 h before they
were resuspended in serum-free medium at a concentration of
2.5×105 cells/ml. A total of 200 µl cell suspension was
added to the upper Transwell chamber, while 500 µl of culture
medium containing 10% FBS was added to the lower chamber. Following
incubation for 24 h, residual cells in the upper chamber were
removed, and cells that had invaded the lower chamber were stained
with a staining solution (0.1% crystal violet ethanol). A total of
three randomly selected representative fields of view were
visualized under a microscope (magnification, ×200) and the average
number of invaded cells was calculated.
Cell viability assay following LL-37
stimulation
Cells (3×103 cells/well) were seeded in
96-well plates and cultured in the absence of serum for 24 h. LL-37
(Sigma-Aldrich; Merck Millipore) was then added at the 0, 0.05,
0.5, 5 and 20 µg/ml at 24, 48 and 72 h after the cells were seeded.
An MTT assay was used to examine cell viability using the
aforementioned procedure.
Cell migration and invasion assays
following LL-37 stimulation
Transwell chambers were used to examine cell
migration and invasion following LL-37 stimulation according to the
aforementioned Transwell assay procedures. In the upper chamber, a
0, 0.05, 0.5 or 5 µg/ml LL-37 was added. Matrigel matrix-coated
chambers were used to determine cell invasion, while cell migration
was determined using uncoated chambers.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
TRIzol reagent (Sigma-Aldrich; Merck Millipore) was
used to extract total RNA from cells following treatment with
LL-37. Total RNA was then reverse transcribed into cDNA using an RT
reaction kit (Promega Corporation, Madison, WI, USA) according to
the manufacturer's instructions. RT-qPCR analysis of YB-1 and
β-actin mRNA expression levels was conducted using SYBR Premix Ex
Taq II (Takara Biotechnology Co., Ltd, Dalian, China) and an
Mx3000P qPCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Reaction mixtures contained 5 µl cDNA (diluted 1:5), 1 µl
forward primer, 1 µl reverse primer, 10 µl qPCR Master Mix 10 and 3
µl nuclease-free water. Thermal cycling parameters consisted of 50
cycles of 95°C for 10 sec and 60°C for 30 sec. The primer sequences
were as follows: Human YB-1 forward,
5′-CAGAATAGTGAGAGTGGGG-3′, and reverse, 5′-ATGTAGTAAGGTGGGAACC-3′;
Human β-actin forward, 5′-TTCCATATCGTCCCAGTTGGT−3′, and
reverse, 5′-CCAGGGCGTTATGGTAGGCA-3′. mRNA levels were calculated
using the 2−ΔΔCq method (26).
Immunofluorescence staining
Following the transfer of a monolayer of cells
(~3×105 cells) to a cell climbing film (Medical
Equipment Factory, Xi'an Jiaotong University, Xi'an, China), LL-37
was added at 0, 0.05, 0.5 or 5 µg/ml and incubated for 48 h. Cells
were subsequently fixed with 4% paraformaldehyde at room
temperature for 10 min, and then treated with 5% Triton X-100 for
15 min followed by 1% goat serum for 30 min. Cells were incubated
with the rabbit anti-human YB-1 monoclonal antibody (1:50 dilution)
overnight at 4°C. Cells were subsequently incubated with a
fluorescein isothiocyanate-labeled goat anti-rabbit antibody [from
KIT-9710; UltraSensitive™ SP (Mouse/Rabbit) IHC Kit; Fuzhou Maixin
Biotechnology Co., Ltd., Fuzhou, China] at 37°C for 1 h, and then
stained with DAPI (Sigma-Aldrich; Merck Millipore) for 1 min.
Staining intensities were visualized using an inverted fluorescence
microscope (LSM 700; Zeiss AG, Oberkochen, Germany).
Signal transduction pathway
analysis
Cells were seeded at 1×105 cells/well in
6-well plates. They were initially treated with 10 µM
mitogen-activated protein kinase kinase (MEK) inhibitor (PD98059),
10 µM p38/mitogen-activated protein kinase (p38/MAPK) inhibitor
(SB203580) or 1 µM NF-κB inhibitor (PDTC; all from Abcam), for 30
min. Cells were subsequently treated with 0.5 µM LL-37 for a
further 24 h. The protein expression levels of YB-1 and β-actin
were then determined by western blot analysis using the
aforementioned procedures.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical significance between two groups was determined using
the Student's t-test or one-way analysis of variance with least
significant difference post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
YB-1 siRNA transfection inhibits YB-1
protein expression and reduces the viability and invasion of MM
cells
Total protein was extracted from A375 and A875 MM
cells at 48 h following transfection with YB-1 siRNA, and YB-1
protein expression levels were determined by western blotting.
Compared with the control group (transfected with non-targeting
siRNA), the protein expression levels of YB-1 in YB-1
siRNA-transfected cells were significantly reduced (Fig. 1A). Initially, three siRNAs were
tested and the most effective siRNA (siRNA1 in A375 cells and
siRNA2 in A875 cells) were used in the following studies. In
addition, the viability of YB-1 siRNA-transfected A375 and A875
cells was significantly decreased at 24 h following transfection
when compared with the control group (P=0.016 in A375 cells;
P=0.018 in A875 cells; Fig. 1B),
indicating that the YB-1 may regulate MM tumor cell viability.
Results from the Transwell invasion assay demonstrated that the
number of YB-1 siRNA-transfected cells that traversed the membrane
following 24 h was significantly lower when compared with that in
the control transfection group (P=0.026 in A375 cells; P=0.021 in
A875 cells; Fig. 1C). This
suggests that YB-1 may control tumor cell invasion. Therefore, YB-1
depletion may reduce the viability and invasiveness of A375 and
A875 cells in vitro.
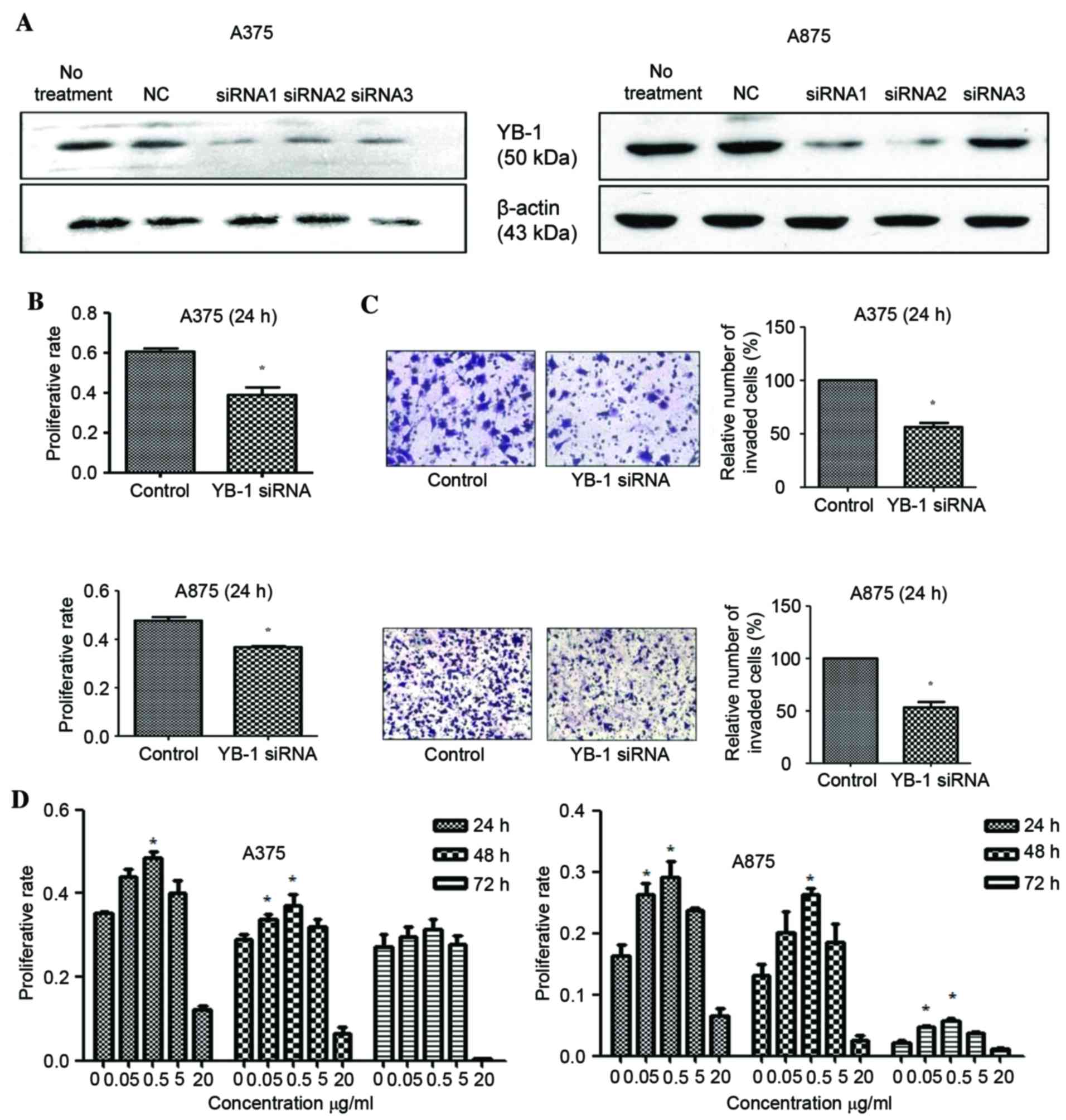 | Figure 1.Effect of YB-1 knockdown on MM cell
viability and invasion, and the effect of LL-37 exposure on MM cell
viability. (A) Western blot analysis showed that siRNA1 in A375
cells and siRNA2 in A875 cells successfully reduced YB-1 protein
expression. (B) Cell viability was reduced in A375 and A875 cells
following YB-1 siRNA transfection, as demonstrated by MTT assay
analysis. (C) The Transwell invasion assay demonstrated that the
number of invaded cells was reduced at 24 h following transfection
with YB-1 siRNA (magnification, ×200). (D) The viability of A375
and A875 cells following exposure to 0, 0.05, 0.5, 5 and 20 µg/ml
LL-37 for 24, 48 and 72 h as determined by MTT assay. Data are
presented as mean ± standard deviation of three independent
experiments, with five samples in each treatment group. *P<0.05
vs. control (non-targeting siRNA) or 0 µg/ml LL-37. YB-1, Y-box
binding protein 1; MM, malignant melanoma. |
LL-37 treatment increases the
viability of MM cells
In order to investigate the effect of LL-37
treatment on the viability of MM cells, A375 and A875 cells were
exposed to increasing concentrations of LL-37 for 24, 48 and 72 h.
Compared with the untreated group, exposure to 0.05, 0.5 and 5
µg/ml LL-37 was associated with an increase in cell viability at
all time points. A statistically significant increase was observed
following exposure of the two cell lines to 0.5 µg/ml at all time
points, with the exception of A375 cells treated for 72 h (P=0.003
in A375 cells; P=0.001 in A875 cells; Fig. 1D).
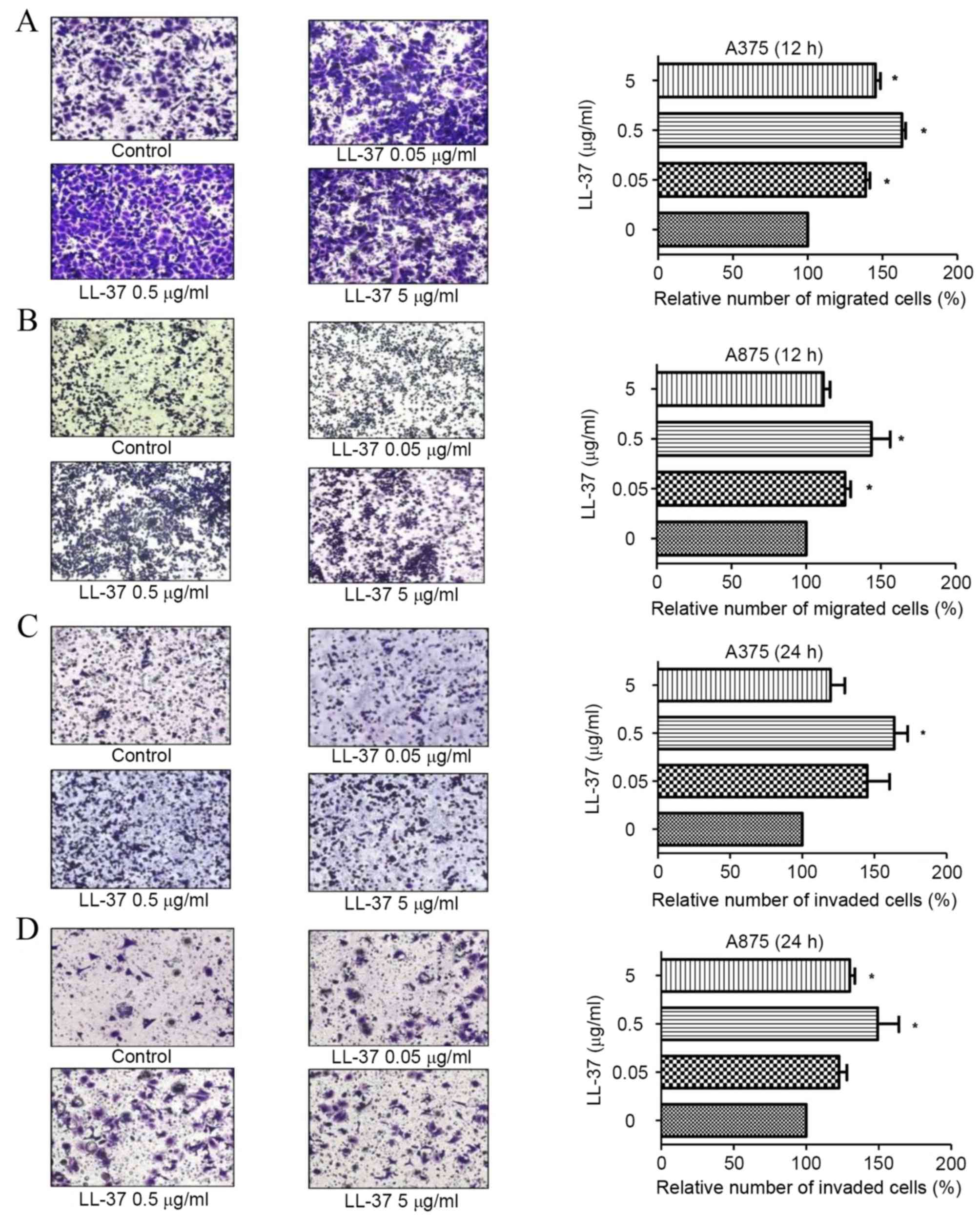
LL-37 promotes the migration and
invasion of MM cells
Compared with the untreated control group, exposure
of A375 and A875 cells to 0.05, 0.5 and 5 µg/ml LL-37 was
associated with an increase in cell migration after 12 h (P=0.018,
Fig. 2A; and P=0.001, Fig. 2B) and invasion after 24 h (P=0.011,
Fig. 2C; and P=0.015, Fig. 2D). A statistically significant
increase in A375 and A875 cell migration and invasion was observed
following exposure to 0.5 µg/ml LL-37 at both time points.
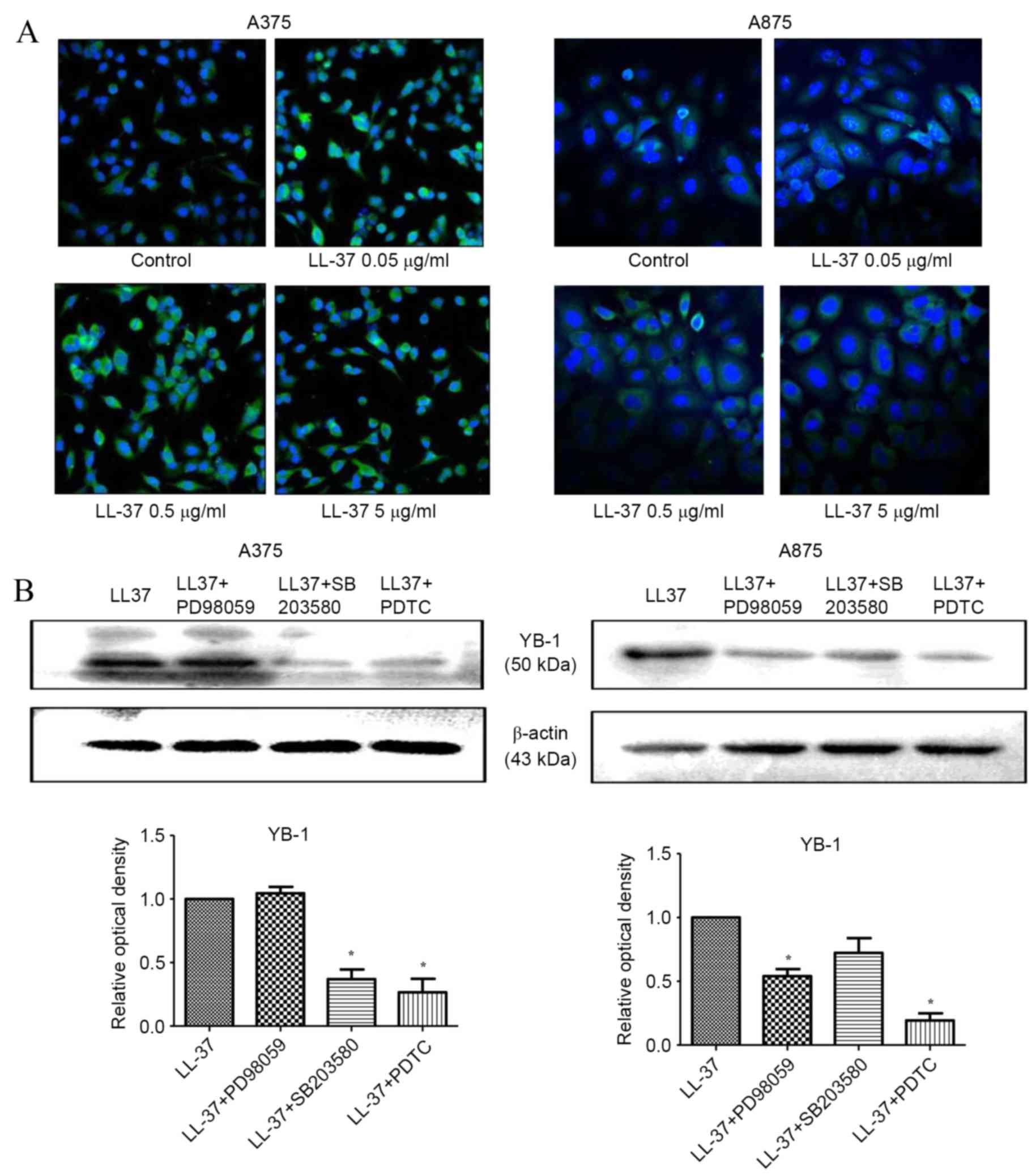
LL-37 promotes YB-1 expression in MM
cells
Compared with the untreated control, exposure of
A375 and A875 cells to 0.5 µg/ml LL-37 for 24 h, was associated
with a significant increase in YB-1 mRNA expression levels [P=0.048
(time) and P=0.031 (dose) in A375 cells; P=0.012 (time) and P=0.023
(dose) in A875 cells; Fig. 3A].
Western blot analysis of YB-1 protein expression demonstrated that
exposure to LL-37 for 48 h led to a marked increase in YB-1 protein
expression levels within A375 and A875 cells, with the most notable
increase observed at a concentration of 0.5 µg/ml (Fig. 3B and C). These results were
confirmed by immunofluorescence staining of YB-1 in cells following
exposure to increasing concentrations of LL-37, which demonstrated
increased fluorescence intensities (Fig. 4A). The most notable increase in
YB-1 expression was observed following exposure of A375 and A875
cells to 0.5 µg/ml LL-37 (Fig.
4A). These results suggest that LL-37 promoted the expression
of YB-1 in A375 and A875 cells.
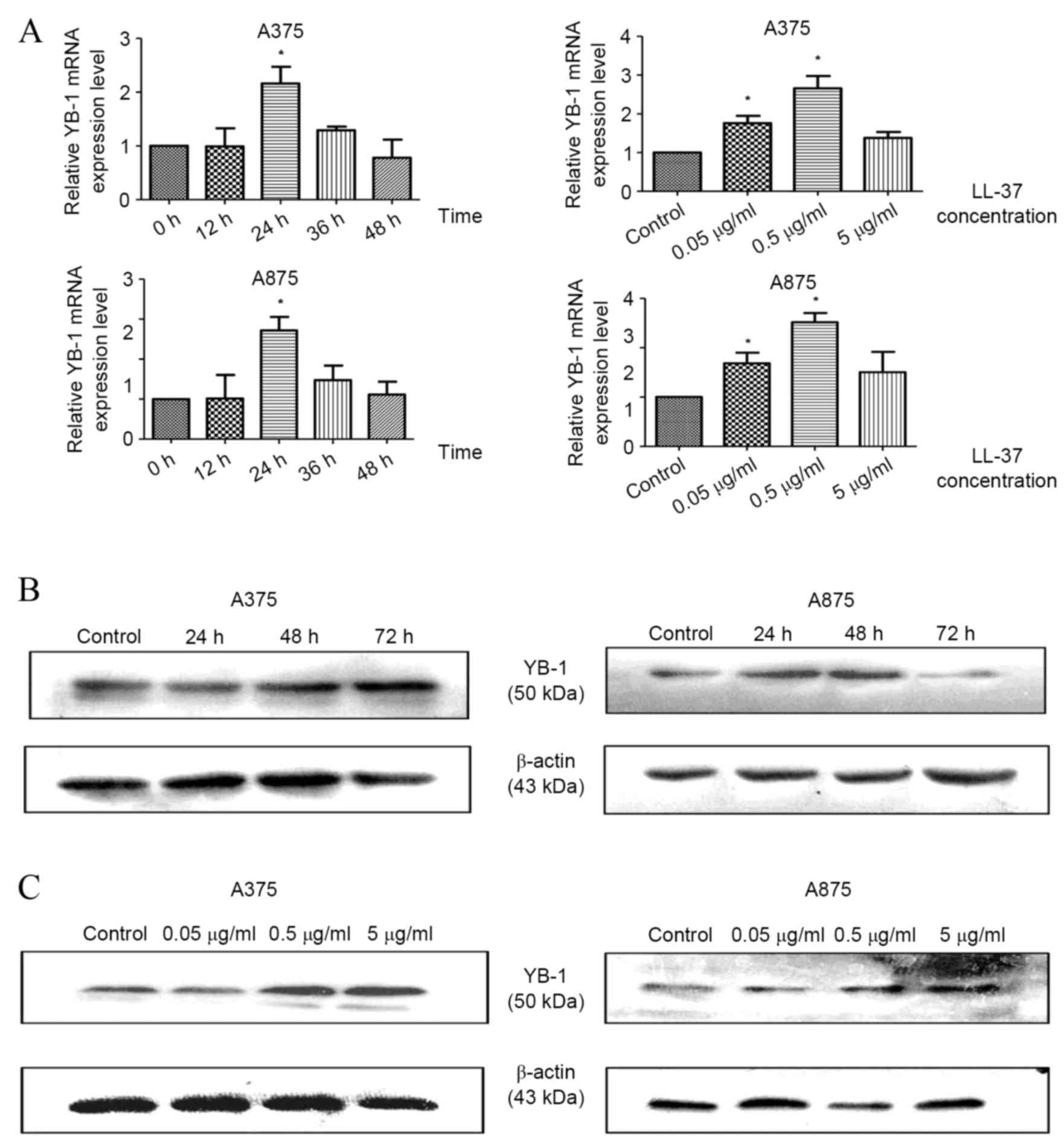 | Figure 3.LL-37 promotes YB-1 expression in
A375 and A875 cells. (A) Reverse transcription-quantitative
polymerase chain reaction analysis of YB-1 mRNA expression in cells
exposed to 0.5 µg/ml LL-37 for 0, 12, 24, 36 or 48 h or exposed to
0, 0.05, 0.5 and 5 µg/ml LL-37 for 24 h. The mRNA levels of YB-1
are shown relative to β-actin mRNA. Western blot analysis of YB-1
protein expression levels in A375 and A875 cells following exposure
to (B) 0.5 µg/ml LL-37 for 0, 24, 48 and 72 h, and (C) 0, 0.05, 0.5
and 5 µg/ml LL-37 for 48 h. Data are presented as the mean ±
standard deviation of three independent experiments. *P<0.05 vs.
control. LL-37, cathelicidin antimicrobial peptide; YB-1, Y box
binding protein 1; MM, malignant melanoma. |
Inhibition of the NF-κB signalling
pathway prevents LL-37-mediated induction of YB-1 expression
Based on earlier results demonstrating that YB-1
expression induced by LL-37 was mediated by the NF-κB signalling
pathway in ovarian cancer cells (27), the role of the NF-κB signalling
pathway in mediating this effect in MM cells was investigated. To
do this, A375 and A875 cells were treated with a MEK inhibitor
(PD98059), a p38/MAPK inhibitor (SB203580), or a NF-κB inhibitor
(PDTC) prior to treatment with LL-37. The p38/MAPK and NF-κB
inhibitors significantly reduced LL-37-induced YB-1 protein
expression levels in A375 cells, whereas the MEK and NF-κB
inhibitors significantly reduced the expression in A875 cells.
Thus, the NF-κB inhibitor PDTC significantly decreased the
LL-37-induced expression of YB-1 in both cell lines (P=0.001 in
A375 cells; P=0.002 in A875 cells; Fig. 4B). These results suggest that the
NF-κB signaling pathway may serve a role in mediating the induction
of YB-1 expression by LL-37 in MM cells.
Discussion
LL-37 is a member of the antimicrobial peptide
family, and its expression is correlated with the proliferation of
epidermal cells (13,14). Previous studies have demonstrated
that LL-37 may promote the progression of numerous malignant
tumors, including lung, ovarian and prostate cancer, as well as
oral squamous cell carcinoma. LL-37-mediated tumor progression is
suggested to be associated with the upregulation of EGFR and the
HER-2 receptor tyrosine kinase (15,23–25,28).
LL-37 transactivates the EGFR via matrix metallopeptidase
(MMP)-mediated cleavage of membrane-anchored EGFR-ligands (29), which may depend on
G-protein-coupled receptors (15,23–25).
In lung squamous cell carcinoma, LL-37 stimulates the proliferation
and invasion of tumor cells via its mitogenic effect on EGFR
phosphorylation, and the subsequent activation of the Ras/MAPK
signaling cascade (22). EGFR
signaling in lung cancer cells serves a direct role in promoting
cell proliferation, anti-apoptotic signaling pathways,
angiogenesis, invasion and metastasis (15,22).
The majority of EGFR ligands, such as transforming growth factor
and heparin-binding epidermal growth factor, are soluble
transmembrane precursors, which are released following proteolytic
cleavage (24,30,31).
These precursors diffuse freely, and bind to and activate the EGFR.
Thus, the oncogenic effects of LL-37 in specific tissues may be
mediated by the activation of EGFR (24,30,31).
In breast cancer, LL-37 promotes tumor progression via activation
of the HER-2-mediated signaling pathway, which may also be mediated
by FPR2 (20). LL-37 triggers the
activation of MAPK and the Janus kinase signal transducer in a
biochemical cascade that leads to the activation of multiple
transcription factors. This process may be dependent or independent
of FPR2 (23,25,32).
LL-37 enhances the invasiveness of ovarian cancer cells by
increasing the activity of tissue remodelling enzymes, such as the
MMP-2 (21,24,30,31).
FPR2 in ovarian cancer cells increases the expression of MMP-2,
thus inhibiting or blocking LL-37 receptors and promoting tumor
cell invasiveness (21,24,30,31).
Previous studies have demonstrated that YB-1
regulates epithelial cell proliferation and is overexpressed in
numerous tumors (33–36). In transgenic mice, an increase in
FPR2 expression promotes the transcription of YB-1 (12,37).
Previous studies have demonstrated that Y-box family proteins may
mediate the regulation of EGFR and HER-2 expression by binding to
the enhancer and promoter regions of these genes, respectively
(7,9). An additional study demonstrated that
upregulation of YB-1 is mediated by the upregulation of
extracellular signal-regulated kinase 2 (ERK2) and glycogen
synthase kinase-3 (GSK-3β) activity. ERK2 and GSK-3β activity is
critical for cell proliferation and apoptosis. Therefore, YB-1 may
be a downstream target of ERK2 and GSK-3β (11,38).
To the best of our knowledge, no previous report has
demonstrated the interaction between LL-37 and YB-1. However, the
current study demonstrated that LL-37, which shares certain common
features with YB-1, may promote the progression and malignant
biological behavior of numerous malignant tumors. The results of
the present study demonstrated that knockdown of YB-1 expression
inhibits the viability and invasion of A375 and A875 MM cells in
vitro. Following exposure to 0.05–5 µg/ml LL-37, A375 and A875
cells exhibited an increase in YB-1 expression. These results
suggest that LL-37 may function to upregulate YB-1 expression MM
cells. Previous studies have reported that LL-37 increases the
malignant potential of tumor cells, which may be associated with
activation of the Ras/MAPK and NF-κB signaling pathways (39,40).
Therefore, ERK, MAPK and NF-κB inhibitors were employed in the
present study to investigate the role of the NF-κB signaling
pathway in mediating LL-37-induced YB-1 expression. Inhibition of
NF-κB in A375 and A875 cell lines significantly reduced the
LL-37-induced YB-1 protein expression. These results suggest that
LL-37 may upregulate YB-1 expression via the NF-κB signaling
pathway. LL-37-mediated activation of EGFR and HER-2 is accompanied
by the phosphorylation of EGFR and activation of the subsequent
Ras/MAPK cascade, thereby promoting the expression of MMP-2 via
FPR2. In addition, EGFR, HER-2 and FPR2 may regulate YB-1
expression (22,23), and the overexpression of these
factors may enhance tumor cell proliferation and invasion. This
suggests that, LL-37 may promote the proliferation and invasion of
A375 and A875 cells through the upregulation of YB-1 expression.
NF-κB is a transcription factor that is known to regulate the
expression of multiple genes, and is involved in a wide range of
cellular responses (41,42). NF-κB activation serves a
significant role in promoting tumor metastasis, and inhibition of
NF-κB can prevent the induction of apoptosis (41,42).
In conclusion, the results of the present study are
consistent with the reported role of YB-1 as a marker of malignancy
in MM. In addition, the antimicrobial peptide LL-37 was
demonstrated to upregulate YB-1 expression and promote the
viability, migration and invasion of A375 and A875 cells in
vitro. Furthermore, the NF-κB signaling pathway may mediate
LL-37-induced YB-1 expression. This study provides a novel insight
into the association between LL-37 and YB-1 in MM, and provides
potential therapeutic targets for the treatment of MM.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81071299, 81371732 and
81573055) and was partially supported by the Fundamental Research
Funds for the Central Universities and for Changjiang Scholars and
Innovative Research Team in University (grant no. PCSIRT:1171).
References
|
1
|
Meier F, Satyamoorthy K, Nesbit M, Hsu MY,
Schittek B, Garbe C and Herlyn M: Molecular events in melanoma
development and progression. Front Biosci. 3:D1005–D1010. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eigentler TK, Caroli UM, Radny P and Garbe
C: Palliative therapy of disseminated malignant melanoma: A
systematic review of 41 randomised clinical trials. Lancet Oncol.
4:748–759. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bekele S, Bekele Y, Mulatu F, Lemma T,
Tilahun H, Gadisa E, Negussie S, Yamuah L, Wassie L, Abebe M, et
al: Recent trends of cutaneous leishmaniasis in Alert Hospital,
Addis Ababa. Ethiop Med J. (Suppl 1). S37–S41. 2014.
|
|
4
|
Kosnopfel C, Sinnberg T and Schittek B:
Y-box binding protein 1-a prognostic marker and target in tumour
therapy. Eur J Cell Biol. 93:61–70. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sakura H, Maekawa T, Imamoto F, Yasuda K
and Ishii S: Two human genes isolated by a novel method encode
DNA-binding proteins containing a common region of homology. Gene.
73:499–507. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Eliseeva IA, Kim ER, Guryanov SG,
Ovchinnikov LP and Lyabin DN: Y-box-binding protein 1 (YB-1) and
its functions. Biochemistry (Mosc). 76:1402–1433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shiota M, Izumi H, Onitsuka T, Miyamoto N,
Kashiwagi E, Kidani A, Yokomizo A, Naito S and Kohno K: Twist
promotes tumor cell growth through YB-1 expression. Cancer Res.
68:98–105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lasham A, Samuel W, Cao H, Patel R, Mehta
R, Stern JL, Reid G, Woolley AG, Miller LD, Black MA, et al: YB-1,
the E2F pathway, and regulation of tumor cell growth. J Natl Cancer
Inst. 104:133–146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lasham A, Print CG, Woolley AG, Dunn SE
and Braithwaite AW: YB-1: Oncoprotein, prognostic marker and
therapeutic target? Biochem J. 449:11–23. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takahashi M, Shimajiri S, Izumi H, Hirano
G, Kashiwagi E, Yasuniwa Y, Wu Y, Han B, Akiyama M, Nishizawa S, et
al: Y-box binding protein-1 is a novel molecular target for tumor
vessels. Cancer Sci. 101:1367–1373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stratford AL, Habibi G, Astanehe A, Jiang
H, Hu K, Park E, Shadeo A, Buys TP, Lam W, Pugh T, et al: Epidermal
growth factor receptor (EGFR) is transcriptionally induced by the
Y-box binding protein-1 (YB-1) and can be inhibited with Iressa in
basal-like breast cancer, providing a potential target for therapy.
Breast Cancer Res. 9:R612007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schittek B, Psenner K, Sauer B, Meier F,
Iftner T and Garbe C: The increased expression of Y box-binding
protein 1 in melanoma stimulates proliferation and tumor invasion,
antagonizes apoptosis and enhances chemoresistance. Int J Cancer.
120:2110–2118. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Durr UH, Sudheendra US and Ramamoorthy A:
LL-37, the only human member of the cathelicidin family of
antimicrobial peptides. Biochim Biophys Acta. 1758:1408–1425. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bucki R, Leszczyńska K, Namiot A and
Sokołowski W: Cathelicidin LL-37: A multitask antimicrobial
peptide. Arch Immunol Ther Exp (Warsz). 58:15–25. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu WK, Wang G, Coffelt SB, Betancourt AM,
Lee CW, Fan D, Wu K, Yu J, Sung JJ and Cho CH: Emerging roles of
the host defense peptide LL-37 in human cancer and its potential
therapeutic applications. Int J Cancer. 127:1741–1747. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Coffelt SB and Scandurro AB: Tumors sound
the alarmin(s). Cancer Res. 68:6482–6485. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hensel JA, Chanda D, Kumar S, Sawant A,
Grizzle WE, Siegal GP and Ponnazhagan S: LL-37 as a therapeutic
target for late stage prostate cancer. Prostate. 71:659–670. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gill K, Mohanti BK, Singh AK, Mishra B and
Dey S: The over expression of cathelicidin peptide LL37 in head and
neck squamous cell carcinoma: The peptide marker for the prognosis
of cancer. Cancer Biomark. 10:125–134. 2011.PubMed/NCBI
|
|
19
|
Kim JE, Kim HJ, Choi JM, Lee KH, Kim TY,
Cho BK, Jung JY, Chung KY, Cho D and Park HJ: The antimicrobial
peptide human cationic antimicrobial protein-18/cathelicidin LL-37
as a putative growth factor for malignant melanoma. Br J Dermatol.
163:959–967. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heilborn JD, Nilsson MF, Jimenez CI,
Sandstedt B, Borregaard N, Tham E, Sørensen OE, Weber G and Ståhle
M: Antimicrobial protein hCAP18/LL-37 is highly expressed in breast
cancer and is a putative growth factor for epithelial cells. Int J
Cancer. 114:713–719. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Coffelt SB, Waterman RS, Florez L, Höner
zu Bentrup K, Zwezdaryk KJ, Tomchuck SL, LaMarca HL, Danka ES,
Morris CA and Scandurro AB: Ovarian cancers overexpress the
antimicrobial protein hCAP-18 and its derivative LL-37 increases
ovarian cancer cell proliferation and invasion. Int J Cancer.
122:1030–1039. 2008.PubMed/NCBI
|
|
22
|
von Haussen J, Koczulla R, Shaykhiev R,
Herr C, Pinkenburg O, Reimer D, Wiewrodt R, Biesterfeld S, Aigner
A, Czubayko F and Bals R: The host defence peptide LL-37/hCAP-18 is
a growth factor for lung cancer cells. Lung Cancer. 59:12–23. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Coffelt SB, Tomchuck SL, Zwezdaryk KJ,
Danka ES and Scandurro AB: Leucine leucine-37 uses formyl peptide
receptor-like 1 to activate signal transduction pathways, stimulate
oncogenic gene expression, and enhance the invasiveness of ovarian
cancer cells. Mol Cancer Res. 7:907–915. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Coffelt SB, Marini FC, Watson K, Zwezdaryk
KJ, Dembinski JL, LaMarca HL, Tomchuck SL, zu Bentrup K Honer,
Danka ES, Henkle SL, et al: The pro-inflammatory peptide LL-37
promotes ovarian tumor progression through recruitment of
multipotent mesenchymal stromal cells. Proc Natl Acad Sci USA.
106:3806–3811. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Girnita A, Zheng H, Grönberg A, Girnita L
and Ståhle M: Identification of the cathelicidin peptide LL-37 as
agonist for the type I insulin-like growth factor receptor.
Oncogene. 31:352–365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao BX, Sun YB, Wang SQ, Duan L, Huo QL,
Ren F and Li GF: Grape seed procyanidin reversal of p-glycoprotein
associated multi-drug resistance via down-regulation of NF-κB and
MAPK/ERK mediated YB-1 activity in A2780/T cells. PLoS One.
8:e710712013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Weber G, Chamorro CI, Granath F, Liljegren
A, Zreika S, Saidak Z, Sandstedt B, Rotstein S, Mentaverri R,
Sánchez F, et al: Human antimicrobial protein hCAP18/LL-37 promotes
a metastatic phenotype in breast cancer. Breast Cancer Res.
11:R62009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tjabringa GS, Aarbiou J, Ninaber DK,
Drijfhout W, Sørensen OE, Borregaard N, Rabe KF and Hiemstra PS:
The antimicrobial peptide LL-37 activates innate immunity at the
airway epithelial surface by transactivation of the epidermal
growth factor receptor. J Immunol. 171:6690–6696. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chuang CM, Monie A, Wu A, Mao CP and Hung
CF: Treatment with LL-37 peptide enhances antitumor effects induced
by CpG oligodeoxynucleotides against ovarian cancer. Hum Gene Ther.
20:303–313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li D, Wang X, Wu JL, Quan WQ, Ma L, Yang
F, Wu KY and Wan HY: Tumor-produced versican V1 enhances
hCAP18/LL-37 expression in macrophages through activation of TLR2
and vitamin D3 signaling to promote ovarian cancer progression in
vitro. PLoS One. 8:e566162013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kittaka M, Shiba H, Kajiya M, Ouhara K,
Takeda K, Kanbara K, Fujita T, Kawaguchi H, Komatsuzawa H and
Kurihara H: Antimicrobial peptide LL37 promotes vascular
endothelial growth factor-A expression in human periodontal
ligament cells. J Periodontal Res. 48:228–234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yasen M, Kajino K, Kano S, Tobita H,
Yamamoto J, Uchiumi T, Kon S, Maeda M, Obulhasim G, Arii S and Hino
O: The up-regulation of Y-box binding proteins (DNA binding protein
A and Y-box binding protein-1) as prognostic markers of
hepatocellular carcinoma. Clin Cancer Res. 11:7354–7361. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang LL, He DL, Li X, Li L, Zhu GD, Zhang
D and Wang XY: Overexpression of coxsackie and adenovirus receptor
inhibit growth of human bladder cancer cell in vitro and in vivo.
Acta Pharmacol Sin. 28:895–900. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guay D, Garand C, Reddy S, Schmutte C and
Lebel M: The human endonuclease III enzyme is a relevant target to
potentiate cisplatin cytotoxicity in Y-box-binding protein-1
overexpressing tumor cells. Cancer Sci. 99:762–769. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shiota M, Yokomizo A, Tada Y, Uchiumi T,
Inokuchi J, Tatsugami K, Kuroiwa K, Yamamoto K, Seki N and Naito S:
P300/CBP-associated factor regulates Y-box binding protein-1
expression and promotes cancer cell growth, cancer invasion and
drug resistance. Cancer Sci. 101:1797–1806. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Koike K, Uchiumi T, Ohga T, Toh S, Wada M,
Kohno K and Kuwano M: Nuclear translocation of the Y-box binding
protein by ultraviolet irradiation. FEBS Lett. 417:390–394. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Oda Y, Ohishi Y, Basaki Y, Kobayashi H,
Hirakawa T, Wake N, Ono M, Nishio K, Kuwano M and Tsuneyoshi M:
Prognostic implications of the nuclear localization of
Y-box-binding protein-1 and CXCR4 expression in ovarian cancer:
Their correlation with activated Akt, LRP/MVP and P-glycoprotein
expression. Cancer Sci. 98:1020–1026. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang Y, Choi H, Seon M, Cho D and Bang SI:
LL-37 stimulates the functions of adipose-derived stromal/stem
cells via early growth response 1 and the MAPK pathway. Stem Cell
Res Ther. 7:582016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pistolic J, Cosseau C, Li Y, Yu JJ,
Filewod NC, Gellatly S, Rehaume LM, Bowdish DM and Hancock RE: Host
defence peptide LL-37 induces IL-6 expression in human bronchial
epithelial cells by activation of the NF-kappaB signaling pathway.
J Innate Immun. 1:254–267. 2009.PubMed/NCBI
|
|
41
|
Ji BC, Hsiao YP, Tsai CH, Chang SJ, Hsu
SC, Liu HC, Huang YP, Lien JC and Chung JG: Cantharidin impairs
cell migration and invasion of A375.S2 human melanoma cells by
suppressing MMP-2 and −9 through PI3K/NF-κB signaling pathways.
Anticancer Res. 35:729–738. 2015.PubMed/NCBI
|
|
42
|
Xia Y, Shen S and Verma IM: NF-κB, an
active player in human cancers. Cancer Immunol Res. 2:823–830.
2014. View Article : Google Scholar : PubMed/NCBI
|