Introduction
Status convulsion (SC) is a common neurological
emergency in childhood. The incidence of SC in Kenya has been
reported to be 268 and 227 per 100,000 per year in children aged
1–11 and 12–59 months, respectively (1). Anticonvulsant drugs are frequently
used to treat SC (2). It has been
reported that SC is associated with the differentiation of
oligodendrocyte precursor cells, which secrete neurite growth
inhibitory factors, including neurite outgrowth inhibitor protein A
(NogoA), oligodendrocyte myelin glycoprotein (OMgp) and
myelin-associated glycoprotein (MAG) (3). In the Ras homolog gene family, member
A (RhoA)/Rho-associated protein kinase (ROCK) signaling pathway,
the myelin-derived inhibitors OMgp, NogoA and MAG may combine with
the leucine rich repeat and immunoglobulin-like domain-containing
protein 1 (Lingo-1)-Nogo-p75/tumor necrosis factor receptor
superfamily, member 19 (TROY) complex and modulate axon propagation
(3). Myelin-derived inhibitors may
combine with the Lingo-1-Nogo-p75/TROY complex to activate Rho and
its downstream effectors, ultimately resulting in functional
alterations in microtubules and actin, and impairment of neuronal
function (4–6). As a component of the Nogo-66
receptor/p75 signaling complex, Lingo-1 primarily regulates the
RhoA/ROCK signaling pathway via activation of RhoA (4,7).
RhoA/ROCK signaling is involved in various diseases of the central
nervous system (CNS), including optic nerve and spinal cord
injuries, stroke and neurodegenerative diseases. The expression
levels of ROCK are upregulated in spinal cord injury, spinal cord
nerve inflammation, demyelinating disease, stroke and other CNS
insults; ROCK inhibitors promote neurite outgrowth and the recovery
of neurological function (8).
Currently, fasudil is the only approved ROCK
inhibitor and it has been demonstrated to be neuroprotective
against glutamate-associated excitotoxicity (9). In neurology, fasudil is used for the
treatment of conditions including vertebral artery insufficiency,
acute cerebral infarction and cerebral vasospasm following
subarachnoid hemorrhage (10–13).
Fasudil specifically blocks the activity of ROCK through
competition with adenosine triphosphate (ATP) for the ROCK
catalytic domain ATP binding site (14). As a potent inhibitor of ROCK,
fasudil may inhibit various other protein kinases, which may
counteract each other in the regulation of neuronal excitability
(15). Recent studies have
demonstrated that fasudil may effectively stimulate neurite
proliferation (16) and serves a
vital role in curing spinal muscular atrophy in rats (17). ROCK inhibitors, including Y-27632
and fasudil, have additionally been revealed to possess
antiepileptic effects (14).
However, few studies have reported the application of fasudil in
the treatment of SC.
The present study investigated the effects of SC on
the RhoA/ROCK signaling pathway using western blot analysis.
Subsequently, the beneficial effects of fasudil on cognitive
function and brain injury in SC rats were determined using an
object-in-place memory task, Morris Water Maze (MWM) test and
histopathology.
Materials and methods
Animals
All animal procedures conformed to the National
Institutes of Health guidelines (18) and were approved by the Ethics
Committee of Chongqing Medical University Animal Center (Chongqing,
China). A total of 56 healthy male Sprague Dawley rats [age, 21
days; weight, 42–52 g; animal certificate no. SCXK (Yu) 2012–0001;
specific pathogen free (SPF)] were provided by the Chongqing
Medical University Animal Center. All rats were housed in an
SPF-class animal room at a temperature of 25±2°C, humidity of
50±10%, 12 h light/dark cycle and freely available food and
water.
Preparation of SC model rats
The SC model was established by the lithium
chloride-pilocarpine method, as described previously (19). Initially, an intraperitoneal
injection of 127 mg/kg lithium chloride (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was administered to all test rats. After 18–20
h, 30 mg/kg pilocarpine (Sigma-Aldrich; Merck KGaA) was
administered by intraperitoneal injection, which induced SC in the
rats. Seizure was divided into five classes according to the Racine
classification method (20), and
SC was defined as Class 5 or 4 seizures 60 min in duration. All
treated rats received 10 mg/kg atropine sulfate (Southwest
Pharmaceutical Co., Ltd., Chongqing, China) and 10 mg/kg diazepam
(Tianjin Jinyao Amino Acid Co., Ltd., Tianjin, China) by
intraperitoneal injection 15 and 60 min following the onset of SC,
respectively. A total of 30 min after the intraperitoneal injection
of pilocarpine, Sprague Dawley rats that had not experienced Class
4 seizures were treated with a second dose of 10 mg/kg pilocarpine;
if class 4 seizures still did not occur, the rats were removed from
the study (n=4). In addition, control rats were injected
intraperitoneally with lithium, atropine and chloral hydrate,
without pilocarpine.
Study design
In the present study, 16 rats were divided into
control and SC groups (n=8/group), to investigate the effect of SC
on the RhoA/ROCK signaling pathway. On day 1 following
establishment of the SC model, rats were sacrificed by decapitation
1–2 min after intraperitoneal injection of 10% chloral hydrate
(Dalian Meilun Biotech Co., Ltd., Dalian, China), and subsequently,
brain tissues were obtained from rats to analyze protein expression
levels of MAG, myelin oligodendrocyte glycoprotein (MOG), NogoA,
Lingo-1 and RhoA in the RhoA/ROCK signaling pathway, by western
blotting. A further 40 rats were used to investigate the effects of
fasudil on cognitive function following SC. These were divided into
four groups (n=10/group): Group I (healthy untreated rats), group
II (healthy rats treated with fasudil), group III (SC rats), group
IV (SC rats treated with fasudil). On day 1 following SC induction,
rats in groups II and IV received intraperitoneal injections of 10
mg/kg fasudil (catalog no. HA-1077, Tianjin Chase Sun
Pharmaceutical Co. Ltd., Tianjin, China) dissolved in saline. Rats
in group I and III received intraperitoneal injections of the same
volume of saline. The cognitive function of rats was assessed 30
days later.
Western blot analysis
The hippocampal region was separated from brain
tissues of the 16 rats in the first SC model and used for western
blot analysis, to determine the protein expression levels of MAG,
MOG, NogoA, Lingo-1, RhoA and phosphorylated (p)-RhoA. Brain
tissues were homogenized with a lysis buffer solution containing:
50 mM Tris-HCl (pH 7.4), 400 mM NaCl, 2 mM EGTA, 1 mM EDTA, 1 mM
dithiothreitol, 10 mM phenylmethylsulphonyl fluoride, 10 µg/ml
leupeptin, 1 µg/ml pepstatin and 1 mM benzamidine. The protein
concentration was determined using a Bicinchoninic Acid assay.
Total cell lysates were prepared in 1X SDS buffer and equal amounts
(50 µg/lane) were loaded on 8% gels, subjected to gel
electrophoresis and subsequently transferred onto polyvinylidene
difluoride membranes (0.22 µm, EMD Millipore, Billerica, MA, USA).
Membranes were blocked with 5% bovine serum albumin (BSA; Huayi
Bioengineering Co. Ltd., Hubei, China) and incubated overnight at
4°C with the following specific antibodies: Mouse anti MAG
(1:5,000; EMD Millipore, catalog no. MAB1567), mouse anti MOG
(1:5,000; EMD Millipore; catalog no. MAB5680), rabbit anti NogoA
(1:5,000; Abcam, Cambridge, UK; catalog no. ab62024), rabbit anti
Lingo-1 (1:5,000; Abcam; catalog no. ab23631), mouse anti RhoA
(1:5,000; Abcam; catalog no. ab54835), rabbit anti p-RhoA (1:5,000;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA); catalog no.
sc-32954) or mouse anti β-actin (1:5,000; Sigma-Aldrich; catalog
no. A3854). Following this, the membranes were washed with TBS
containing Tween-20 and incubated with horse radish peroxidase
(HRP)-conjugated goat anti-rabbit IgG (1:10,000; Beijing
Biosynthesis Biotechnology Co., Ltd., Beijing, China; catalog no.
bs-0296G-HRP) or goat anti-mouse IgG (1:5,000; Beijing Biosynthesis
Biotechnology Co., Ltd., catalog no. bs-0296G-HRP) secondary
antibodies for 2 h at room temperature. Target proteins were
detected using an Enhanced Chemiluminescence kit (Beyotime
Institute of Biotechnology, Haimen, China). The ratios of target
proteins to internal control were calculated using ImageJ software
version 1.5.0 (National Institutes of Health, Bethesda, MD,
USA).
Object-in-place memory task
All 40 rats in the second SC model were assessed on
object-in-place recognition memory 30 days following fasudil
injection. Exploration occurred in a square and open-topped arena
(100 cm3) of black plexiglass. The floor of the arena
was covered with sawdust. An overhead camera and video recorder
were used to monitor and record the behavior of the rats for
subsequent analysis. This procedure lasted for 4 days and comprised
an adaptive phase and an acquisition phase separated by a delay
from a recognition test. The first two days was the adaptive phase,
during which rats were placed into the arena for 10 min at the same
time each day.
During the familiarization phase, two identical
objects (A1 and A2) were placed near the two corners at either end
of one side of the arena (10 cm from each adjacent wall). The rats
were placed into the arena facing the center of the opposite wall
and allowed 3 min in the arena to find A1 and A2. Exploratory
behavior was defined as the animal directing its nose toward the
object at a distance of <2 cm. Other behaviors, including
looking around while sitting on or resting against the object, were
not considered as exploration. The delay between the phases was 5
min or 1 h depending on the experiment. During the delay period,
the objects were cleaned with 75% alcohol to remove olfactory cues
and any sawdust. For the object-in-place memory test, the two
objects were removed and one replaced with a familiar object (A3)
and one with a novel object (B) and the subjects were allowed to
explore the objects for 3 min. To assess the spatial recognition
ability of the rats, the position of object B was altered. The time
spent exploring the two objects that had altered position was
compared with the time spent exploring the two objects that had
remained in the same position. If object-in-place memory is intact,
the subject will spend more time exploring the two objects that had
altered position, compared with the two objects that are in the
same locations. The positions of the objects in the test and the
objects used as novel or familiar were counterbalanced among rats.
The discrimination ratio was calculated using the following
equation: Discrimination ratio (%)=[novel (s)/familiar (s)]
×100.
MWM test
On day 2 following the object-in-place memory task,
spatial learning and memory was assessed using the SLY-WMS MWM
system version 2.0 (Beijing Sunny Instruments, Co., Ltd., Beijing,
China) in the 40 rats of the second SC model, as described
previously (21,22). Briefly, the procedure took place in
a black circular pool (diameter, 200 cm; depth, 50 cm; inside and
outside painted black) filled with 24°C water (dyed with black ink
to be opaque) to a depth of 30 cm. The entire procedure was
recorded by an overhead camera for subsequent analysis. The MWM
tests were performed on six consecutive days and comprised initial
spatial training (day 1), spatial reversal training (days 2–5) and
probe test (day 6).
Histopathology
On the day following the final MWM test, one rat in
each group was randomly selected for hematoxylin and eosin (HE) and
Nissl staining. Hippocampal organotypic tissues were obtained from
rats, as aforementioned. For cross-sectional observation, tissues
were fixed in 4% paraformaldehyde for 24 h and routinely processed
into 4-µm thick paraffin-embedded sections. All sections were
placed on slides pretreated with 3-aminopropyl-triethoxysilane, and
dried overnight in an oven at 60°C. HE staining and Nissl staining
were performed using conventional methods.
For HE staining, deparaffinized sections were
incubated with hematoxylin solution for 8 sec, differentiated with
0.5% HCl for 10 sec, stained with lithium carbonate solution for 20
sec and counterstained with eosin staining solution for 10 sec.
Sections were observed under a light microscope (Eclipse 55i; Nikon
Corporation, Tokyo, Japan). For Nissl staining, deparaffinized
sections were dehydrated in alcohol, immersed in 1% cresyl fast
violet solution for 23–30 min, differentiated in 95% alcohol,
hyalinized in xylene and mounted with neutral gum. Sections were
observed under a light microscope.
SC susceptibility analysis
SC was subsequently induced for a second time by
intraperitoneal injection of pilocarpine, as aforementioned, in
second SC model rats two days after the MWM test. The time from the
injection of pilocarpine to Class 4 seizures (SC latency) of each
rat was recorded for subsequent analysis.
Statistical analysis
Statistical analyses were conducted using SPSS
software version 17.0 (SPSS, Inc., Chicago, IL, USA). Data for
western-blotting analysis and animal behavior analysis are
expressed as the mean ± standard deviation of test rats in each
group. The number of times the platform was passed was analyzed by
Wilcoxon signed-rank-sum test. The protein expression levels of
neurite growth inhibitory factors between-groups were compared
using a one-way analysis of variance, followed by the least
significant difference post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of neurite growth
inhibitory factors in the RhoA/ROCK signaling pathway
To investigate the effect of SC on the protein
expression levels of neurite growth inhibitory factors in the
RhoA/ROCK signaling pathway, western blot analysis was performed.
On day 1 following SC model establishment, the hippocampal CA1
region was separated from rat brain tissues. Lingo-1 is a
transmembrane protein. As presented in Fig. 1, Lingo-1 expression levels were
significantly increased in the SC group compared with the control
group (P<0.05). In addition, the expression levels of MAG and
MOG demonstrated a similar pattern (P<0.05). However, NogoA
expression levels were greater in the control group compared with
the SC group (P<0.05). In addition, the protein expression
levels of RhoA and p-RhoA were measured. The expression levels of
RhoA were not significantly altered following SC (P>0.05).
However, the protein expression levels of p-RhoA were significantly
increased in the SC group compared with the control group
(P<0.01).
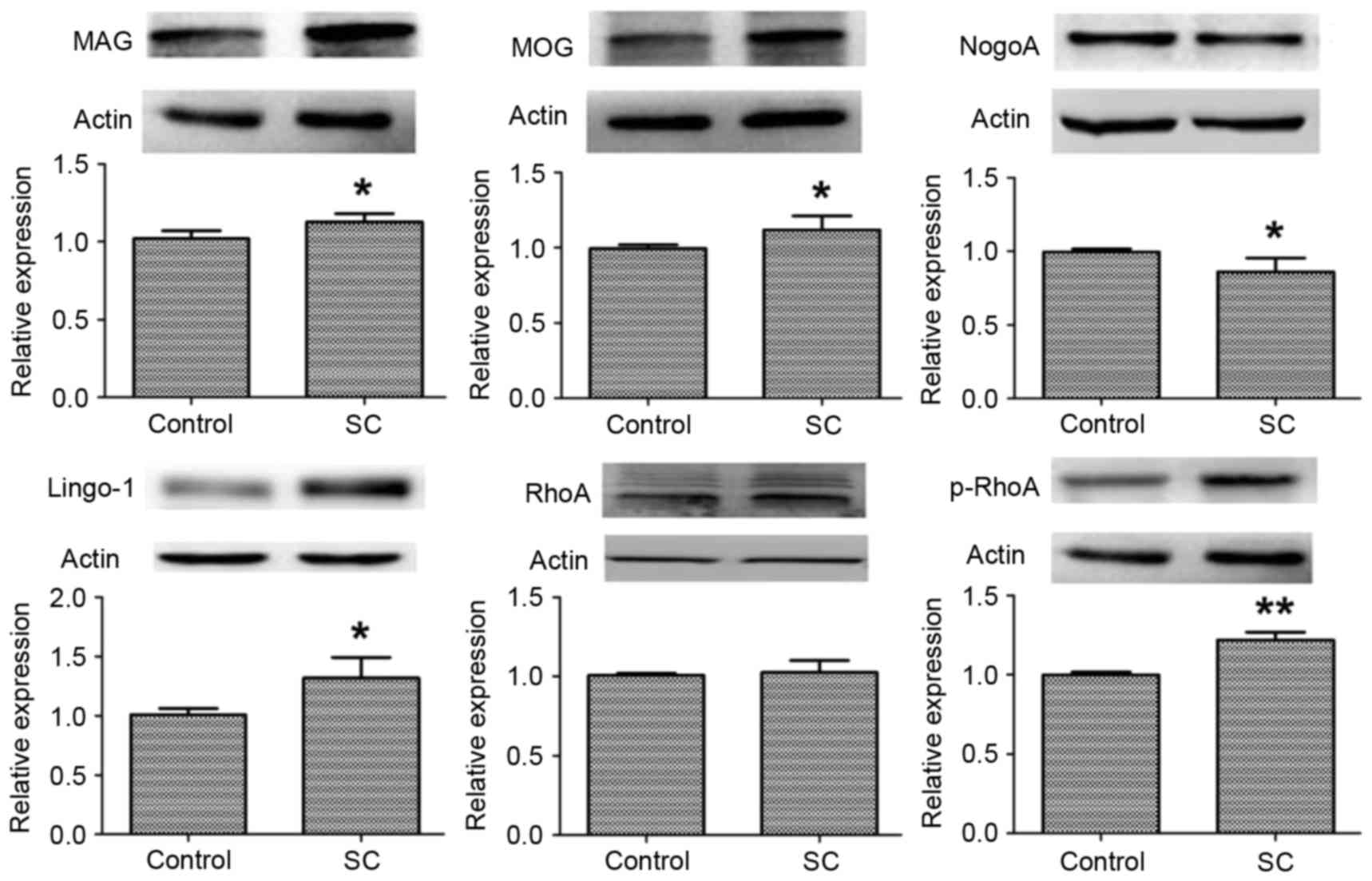 | Figure 1.Protein expression levels of neurite
growth inhibitory factors in the hippocampal CA1 region. The
protein expression levels of MAG, MOG, NogoA, Lingo-1, RhoA and
p-RhoA were measured by western blotting; β-actin served as an
internal control. Data are presented as the mean ± standard
deviation of the rats in each group. *P<0.05 and **P<0.01 vs.
control. MAG, myelin-associated glycoprotein; MOG, myelin
oligodendrocyte glycoprotein; NogoA, neurite outgrowth inhibitor
protein A; Lingo-1, leucine rich repeat and immunoglobulin-like
domain-containing protein 1; RhoA, Ras homolog gene family, member
A; p-, phosphorylated. |
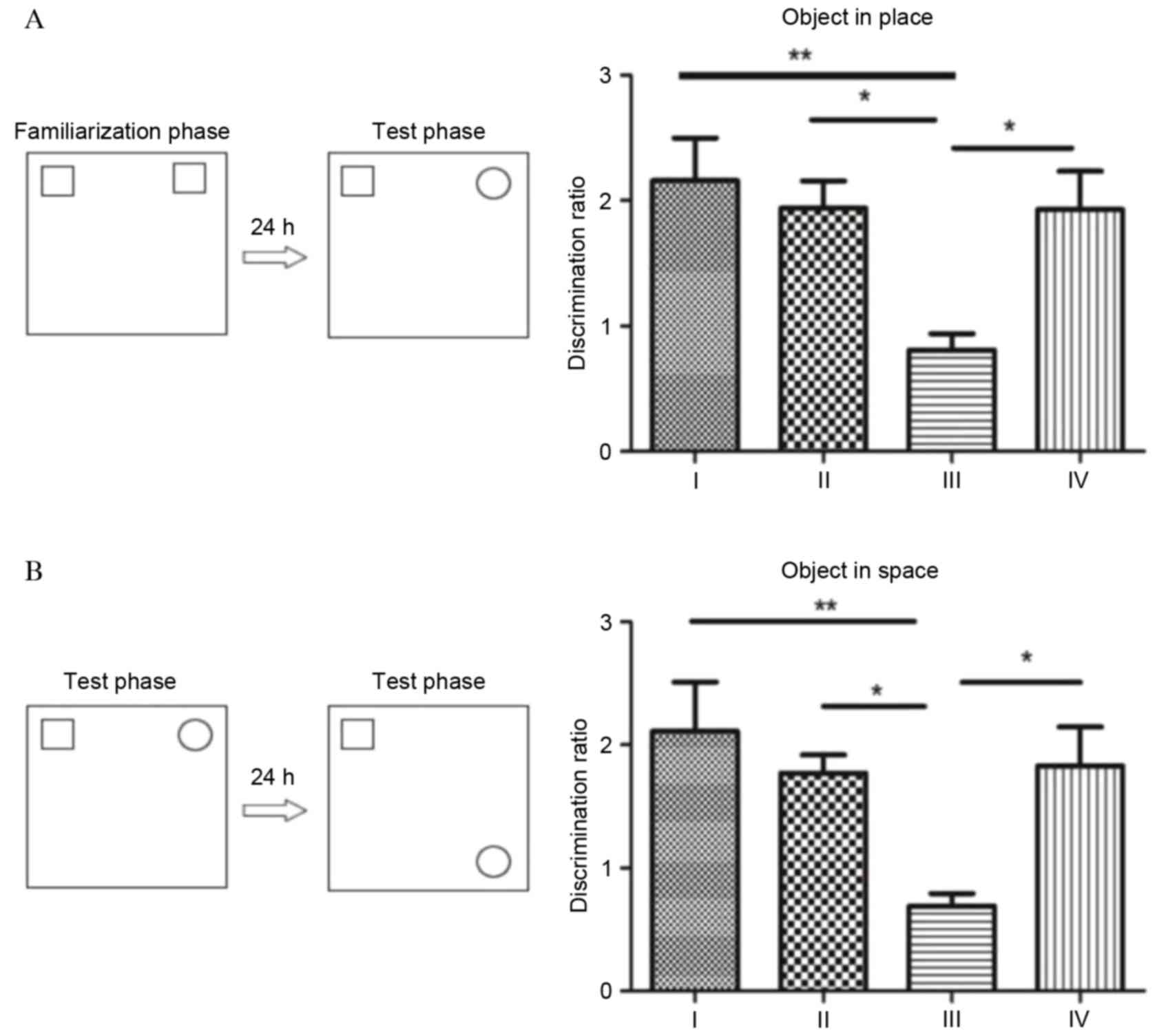
Object-in-place memory task
To measure the cognitive function of rats, the
object-in-place memory task was performed (Fig. 2). In the object location
recognition test, analysis of acquisition phase exploration
revealed significant differences between the four groups. As
presented in Fig. 2A, no
significant differences were observed in object-in-place memory
between group I, II and IV (P>0.05). By contrast, rats in group
III demonstrated significantly impaired object-in-place memory. The
discrimination ratio of rats in group III was significantly
decreased compared with groups I (P<0.01) and II (P<0.05).
Fasudil significantly improved the object-in-place memory in rats,
with the discrimination ratio significantly increased in group IV
compared with group III (P<0.05).
In the object-in-place memory test, significant
memory is inferred when rats spend more time exploring the objects
that switched locations compared with the objects that remained in
the same location as the sample phase. Analysis of the
discrimination ratio during the acquisition phase revealed
significant differences among the four groups, which demonstrated a
similar profile to the object location recognition test (Fig. 2). Although the discrimination ratio
was greater in group I rats compared with groups II and IV, no
significant differences were observed between the three groups
(P>0.05). Following SC, rats in group III demonstrated impaired
object-in-place memory, with a discrimination ratio significantly
reduced compared with groups I (P<0.01), II and IV
(P<0.05).
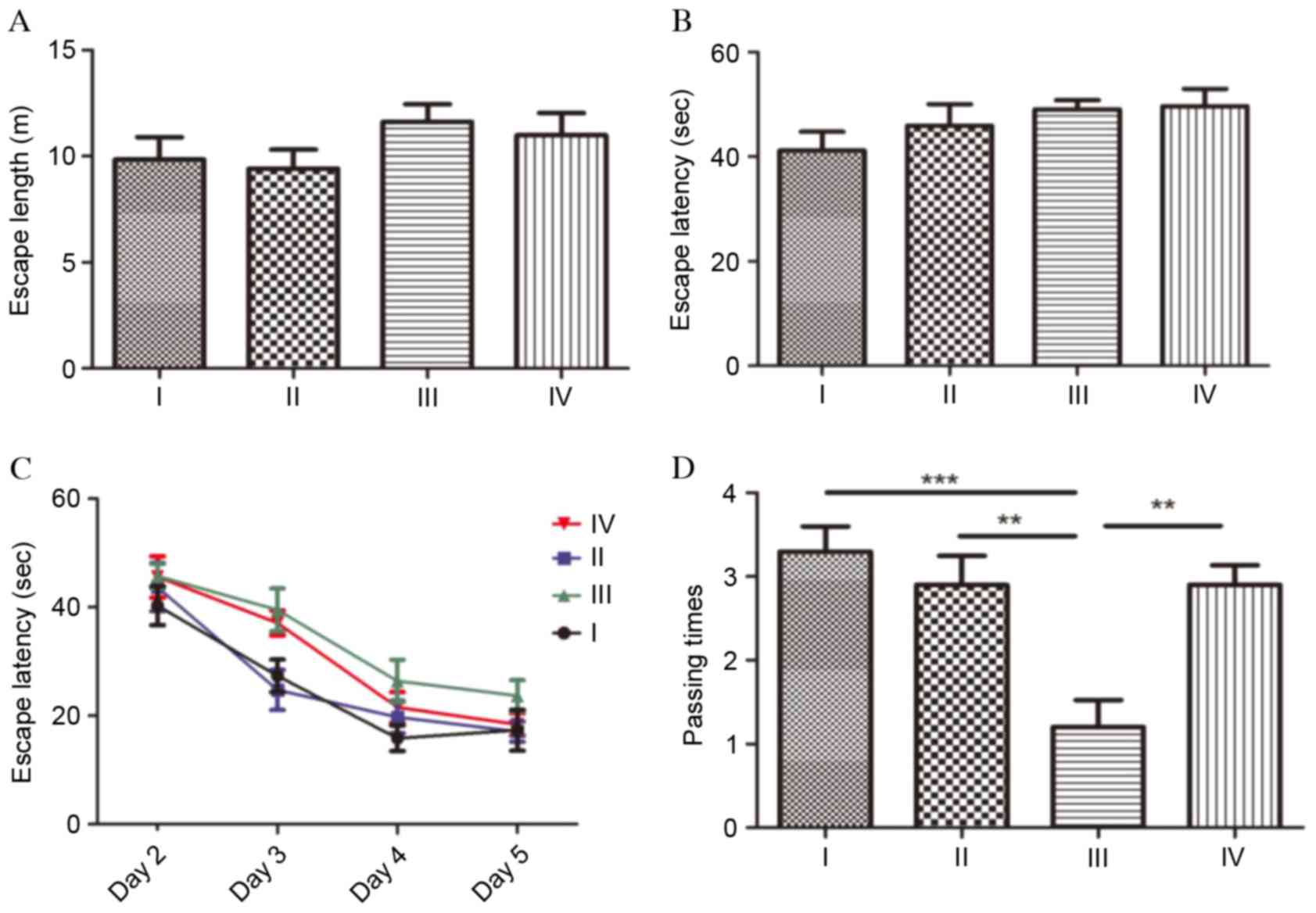
MWM test
To evaluate spatial learning and the memory of the
rats, the MWM test was performed (Fig.
3). The test was performed on 6 consecutive days and comprised
initial spatial training (day 1), spatial reversal training (days
2–5) and probe test (day 6). At the initial spatial training phase,
no significant differences were observed in escape length (Fig. 3A) or escape latency (Fig. 3B) between the four groups
(P>0.05). During the spatial reversal training phase, the escape
latencies of group III were longer than the other three groups;
however, compared with group III, there were no significant
differences between the groups (P>0.05; Fig. 3C). Escape latencies in each group
demonstrated a gradually decreasing profile. At the probe test
phase, the number of times the platform quadrant area was passed
was analyzed during a 60 sec period. Compared with the rats in
group I, the number of times the platform quadrant area was passed
was significantly decreased in group III (P<0.001), which
indicated that cognitive function was severely impaired following
SC. The passing times in group III rats were additionally
significantly reduced compared with group II rats (P<0.01).
Fasudil markedly promoted spatial learning and memory in rats
following SC, as the passing times in group IV were significantly
increased compared with group III (P<0.01).
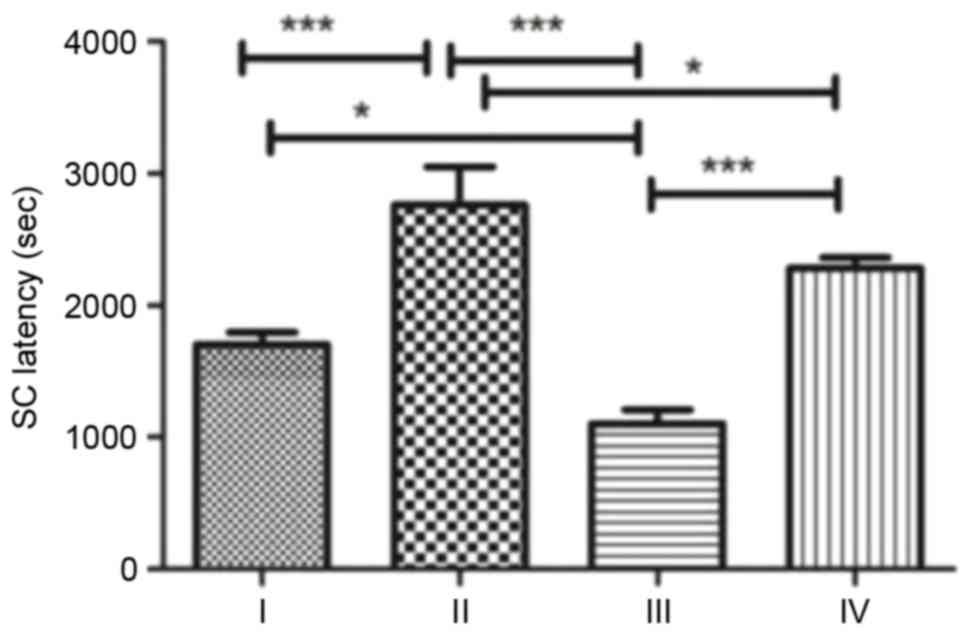
SC susceptibility analysis
SC was induced for a second time by intraperitoneal
injection of pilocarpine and the time from the injection to Class 4
seizures in each group was recorded as SC latency (Fig. 4). Rats in group II demonstrated the
longest SC latency, significantly longer compared with groups I,
III (P<0.001) and IV (P<0.05). Rats in group III demonstrated
the shortest SC latency. Following treatment with fasudil, SC
latency in group IV increased to approximately twice that of group
III (P<0.001).
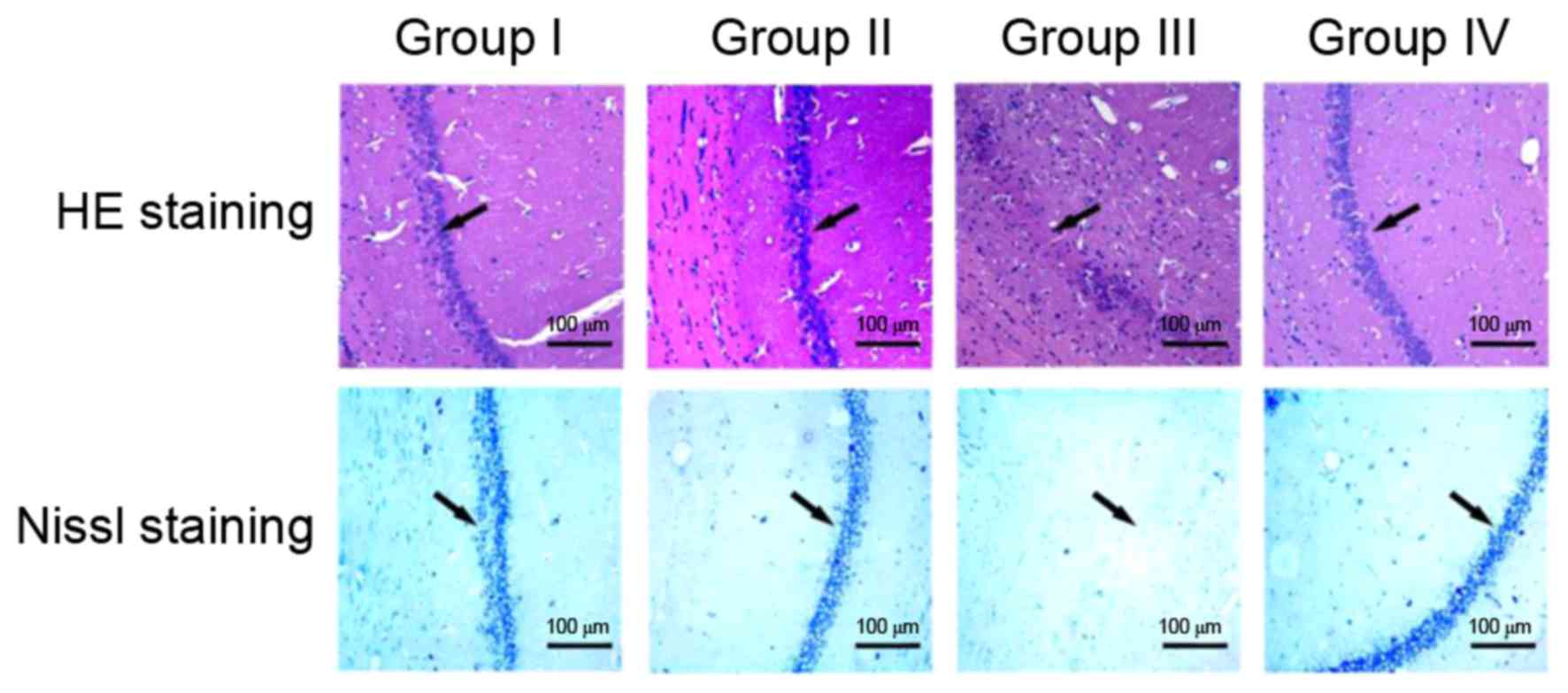
Histopathology
Morphological analysis of the hippocampal
organotypic tissue of rats was performed using HE and Nissl
staining (Fig. 5). HE and Nissl
staining revealed similar results in the four groups. In groups I
and II, no clear differences were observed in the hippocampal CA1
region of rats in HE and Nissl staining; the neuronal cells of the
hippocampal CA1 region were intact, regularly arranged and evenly
distributed. In group III, the number of hippocampal CA1 neurons
and Nissl bodies was decreased and HE staining revealed poorly
arranged neurons. Nissl staining of the hippocampal CA1 in group
III revealed an unclear structure with almost no neurons. In group
IV, fasudil treatment markedly promoted the recovery of the CA1
regional structure, including clearly visible Nissl bodies and a
relatively regular arrangement of neurons, although a small
quantity of irregular dark cellular matter was present.
Discussion
A number of studies have demonstrated that SC is
closely associated with CNS (23,24).
The majority of neurite growth inhibitory factors, including
Lingo-1, MAG, NogoA and OMgp, may activate the RhoA/ROCK signaling
pathway and further modulate axon propagation (6,7).
Generalized seizures exhibit distinct neuroanatomical substrates
that initiate and sustain seizure activity (25). The anticonvulsant effects of the
MCAT cells on acute SC may be associated with inhibitory factors
and immune-modulatory mechanisms assigned to mesenchymal cells in
the hippocampus (26). The
hippocampus is a structure susceptible to SC and serves an
important role in memory function (27). In the present study, rat
hippocampal CA1 tissues were selected for the study of fasudil on
cognitive function. The protein expression levels of Lingo-1, MAG,
MOG and p-RhoA were significantly increased one day after SC. SC
had no effect on RhoA expression levels, but significantly reduced
NogoA expression levels. These results indicated that the
alterations in neurite growth inhibitory factors were not fully
synchronized and ultimately resulted in CNS damage, which may be
induced by RhoA phosphorylation following SC and the activation of
the RhoA/ROCK signaling pathway. Lingo-1 is a leucine-rich repeat
transmembrane protein, which may transmit a signal of axon growth
inhibition to cells and convert non-activated RhoA-guanosine
diphosphate to an activated form of RhoA-guanosine triphosphate,
and further activate downstream ROCK (4). The alterations in Lingo-1 expression
levels were consistent with those of p-RhoA. The activated ROCK may
alter the cytoskeleton dynamics and inhibit axon propagation,
leading to the collapse of the neuronal cytoskeleton and CNS damage
(5).
Fasudil is a 5-isoquinoline sulfonamide derivative
and primarily inhibits ROCK, which is involved in a variety of
cellular functions, including apoptosis and smooth muscle
contraction. A previous study indicated that fasudil improved CNS
injury, promoted axonal regeneration and reduced cell death
(28). In a study of other
neurological diseases, fasudil was revealed to possess a protective
effect against CNS injury (16).
Lingor et al (29) applied
three inhibitors (Y-27632, fasudil and dimethylfasudil) to suppress
the activity of ROCK, which effectively induced nerve cell
regeneration following CNS injury. Whether fasudil has a protective
effect on seizure-induced brain injury remains to be determined.
The results of the present study demonstrated that fasudil may
improve the structure of the hippocampus and cognitive function in
rats. The hippocampal CA1 region almost disappeared in the SC rats
of group III, and recovered to almost normal in the rats of group
IV.
The object-in-place memory task and the MWM test
were used to analyze cognitive function. The specificity of the
recognition memory deficit in the object-in-place test suggested
that cognitive functions associated with the prefrontal cortex may
be particularly susceptible to alteration following prenatal
infection (30). Novel object
recognition was used as a facile behavior test for evaluating drug
effects in an Alzheimer's disease AbetaPP/PSl mouse model (31). The MWM test is recognized as a
method which effectively evaluates learning and memory in animal
models (32). Hippocampal
long-term depression mediates spatial reversal learning in the MWM
test (21). The hippocampus and
dorsal striatum serve critical roles in navigation based on spatial
or cue-based strategies and the spatial and cue versions of the MWM
are impaired by lesions in the dorsal hippocampus and dorsal
striatum, respectively (22). The
object-in-place memory task and MWM test demonstrated similar
results in the present study, which suggested that fasudil
significantly improved the cognitive ability of SC rats. Rats in
group III exhibited the lowest discrimination ratio, the least
number of times passing the platform quadrant area and the longest
escape length and latency. Following intraperitoneal injection with
fasudil, learning and memory in rats of group IV was significantly
improved.
Furthermore, SC susceptibility was analyzed and the
time from injection with pilocarpine to Class 4 seizures was
recorded in each group. SC latency was the shortest in group III;
therefore, rats in group III had the greatest susceptibility to SC,
which was consistent with previous studies (33,34).
Notably, fasudil lengthened SC latency compared with group III. In
addition, the SC latency in rats of group II was greater compared
with group I. There are certain limitations to the present study.
Lingo-1 is a transmembrane protein, which is located upstream of
the signaling pathway of interest. Further studies are required to
elucidate whether an improved therapeutic effect may be achieved in
rats following SC when the expression of Lingo-1 is inhibited.
In conclusion, the results of the present study
demonstrated that fasudil improves cognitive function and CNS
injury in rats, and decreases SC susceptibility. Fasudil therefore
has a protective effect on convulsive brain injury. Fasudil and SC
may regulate the CNS by affecting the expression levels of neurite
growth inhibitory factors in the RhoA/ROCK signaling pathway.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81371452), the
Natural Science Foundation of Chongqing City (grant no.
cstc2013jjB0031), the Foundation of Chongqing Municipal Commission
of science and technology (grant no. 2015XMSB000712) and the PhD
Programs Foundation of Ministry of Education of China (grant no.
20125503110011).
Glossary
Abbreviations
Abbreviations:
|
MAG
|
myelin-associated glycoprotein
|
|
MOG
|
myelin oligodendrocyte
glycoprotein
|
|
NogoA
|
neurite outgrowth inhibitor protein
A
|
|
Lingo-1
|
leucine rich repeat and
immunoglobulin-like domain-containing protein 1
|
|
RhoA
|
Ras homolog gene family, member A
|
|
SC
|
status convulsion
|
|
ROCK
|
Rho-associated protein kinase
|
|
OMgp
|
oligodendrocyte myelin
glycoprotein
|
|
CNS
|
central nervous system
|
References
|
1
|
Sadarangani M, Seaton C, Scott JA, Ogutu
B, Edwards T, Prins A, Gatakaa H, Idro R, Berkley JA, Peshu N, et
al: Incidence and outcome of convulsive status epilepticus in
Kenyan children: A cohort study. Lancet Neurol. 7:145–150. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Appleton R, Martl T and Phillips B: Drug
management for acute tonic-clonic convulsions including convulsive
status epilepticus in children. Cochrane Database Syst Rev:
CD001905. 2002. View Article : Google Scholar
|
|
3
|
Lee JK, Geoffroy CG, Chan AF, Tolentino
KE, Crawford MJ, Leal MA, Kang B and Zheng B: Assessing spinal axon
regeneration and sprouting in Nogo-, MAG-, and OMgp-deficient mice.
Neuron. 66:663–670. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mi S, Lee X, Shao Z, Thill G, Ji B, Relton
J, Levesque M, Allaire N, Perrin S, Sands B, et al: LINGO-1 is a
component of the Nogo-66 receptor/p75 signaling complex. Nat
Neurosci. 7:221–228. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luo L: Actin cytoskeleton regulation in
neuronal morphogenesis and structural plasticity. Annu Rev Cell Dev
Biol. 18:601–635. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stankiewicz TR and Linseman DA: Rho family
GTPases: Key players in neuronal development, neuronal survival,
and neurodegeneration. Front Cell Neurosci. 8:3142014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fujita Y and Yamashita T: Axon growth
inhibition by RhoA/ROCK in the central nervous system. Front
Neurosci. 8:3382014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roloff F, Scheiblich H, Dewitz C,
Dempewolf S, Stern M and Bicker G: Enhanced neurite outgrowth of
human model (NT2) neurons by small-molecule inhibitors of Rho/ROCK
signaling. PLoS One. 10:e01185362015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kitaoka Y, Kitaoka Y, Kumai T, Lam TT,
Kuribayashi K, Isenoumi K, Munemasa Y, Motoki M, Kobayashi S and
Ueno S: Involvement of RhoA and possible neuroprotective effect of
fasudil, a Rho kinase inhibitor, in NMDA-induced neurotoxicity in
the rat retina. Brain Res. 1018:111–118. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ishiguro M, Kawasaki K, Suzuki Y, Ishizuka
F, Mishiro K, Egashira Y, Ikegaki I, Tsuruma K, Shimazawa M,
Yoshimura S, et al: A Rho kinase (ROCK) inhibitor, fasudil,
prevents matrix metalloproteinase-9-related hemorrhagic
transformation in mice treated with tissue plasminogen activator.
Neuroscience. 220:302–312. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saito A, Inoue M, Kon H, Imaruoka S,
Basaki K, Midorikawa H, Sasaki T and Nishijima M: Effectiveness of
intraarterial administration of fasudil hydrochloride for
preventing symptomatic vasospasm after subarachnoid hemorrhage.
Acta Neurochir Suppl. 120:297–301. 2015.PubMed/NCBI
|
|
12
|
Iwabuchi S, Hayashi M, Yokouchi T, Sato K,
Nakayama H, Harashina J, Iwama J, Ishii M, Hiramoto Y, Hirai N, et
al: Prophylactic intra-arterial administration of fasudil
hydrochloride for vasospasm following subarachnoid haemorrhage.
Acta Neurochir Suppl. 120:167–169. 2015.PubMed/NCBI
|
|
13
|
Liu YH, Zhao Y, Huang FZ, Chen YH, Wang
HX, Bonney E and Liu BQ: Combination of early constraint-induced
movement therapy and fasudil enhances motor recovery after ischemic
stroke in rats. Int J Neurosci. 126:168–173. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Inan S and Büyükafşar K: Antiepileptic
effects of two Rho-kinase inhibitors, Y-27632 and fasudil, in mice.
Br J Pharmacol. 155:44–51. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davies S, Reddy H, Caivano M and Cohen P:
Specificity and mechanism of action of some commonly used protein
kinase inhibitors. Biochem J. 351:95–105. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen S, Luo M, Zhao Y, Zhang Y, He M, Cai
W and Liu A: Fasudil stimulates neurite outgrowth and promotes
differentiation in C17.2 neural stem cells by modulating notch
signalling but not autophagy. Cell Physiol Biochem. 36:531–541.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Coque E, Raoul C and Bowerman M: ROCK
inhibition as a therapy for spinal muscular atrophy: Understanding
the repercussions on multiple cellular targets. Front Neurosci.
8:2712014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the care and use of laboratory animals. 8th.
Washington, DC: National Academies Press; 2011
|
|
19
|
Lively S and Brown IR: Analysis of the
extracellular matrix protein SC1 during reactive gliosis in the rat
lithium-pilocarpine seizure model. Brain Res. 1163:1–9. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Racine RJ: Modification of seizure
activity by electrical stimulation: II. Motor seizure.
Electroencephalogr Clin Neurophysiol. 32:281–294. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong Z, Bai Y, Wu X, Li H, Gong B, Howland
JG, Huang Y, He W, Li T and Wang YT: Hippocampal long-term
depression mediates spatial reversal learning in the Morris water
maze. Neuropharmacology. 64:65–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Miyoshi E, Wietzikoski EC, Bortolanza M,
Boschen SL, Canteras NS, Izquierdo I and Da Cunha C: Both the
dorsal hippocampus and the dorsolateral striatum are needed for rat
navigation in the Morris water maze. Behav Brain Res. 226:171–178.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
D'Agostino DP, Pilla R, Held HE, Landon
CS, Puchowicz M, Brunengraber H, Ari C, Arnold P and Dean JB:
Therapeutic ketosis with ketone ester delays central nervous system
oxygen toxicity seizures in rats. Am J Physiol Regul Integr Comp
Physiol. 304:R829–R836. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chapman CD, Frey WH II, Craft S, Danielyan
L, Hallschmid M, Schiöth HB and Benedict C: Intranasal treatment of
central nervous system dysfunction in humans. Pharm Res.
30:2475–2484. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Browning RA, Wang C, Nelson DK and Jobe
PC: Effect of precollicular transection on audiogenic seizures in
genetically epilepsy-prone rats. Exp Neurol. 155:295–301. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tamura B, Almeida D, Felizardo R, Olanda
G, Bocca L, Pinhal N, Alves-de-Moraes L, Covolan L, Cãmara N and
Longo BM: Convulsive seizure protection after hippocampal
transplantation of mesenchymal cells from adipose tissue in mice. J
Stem Cell Res Ther. 4:22014.
|
|
27
|
Scharfman HE: Epileptogenesis in the
parahippocampal region. Parallels with the dentate gyrus. Ann N Y
Acad Sci. 911:305–327. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nishio Y, Koda M, Kitajo K, Seto M, Hata
K, Taniguchi J, Moriya H, Fujitani M, Kubo T and Yamashita T:
Delayed treatment with Rho-kinase in-hibior dose not enhance axonal
regeneration or functional recovery after spinal cord injury in
rats. Exp Neurol. 200:392–397. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lingor P, Teusch N, Schwarz K, Mueller R,
Mack H, Bähr M and Mueller BK: Inhibition of Rho kinase (ROCK)
increases neurite outgrowth on chondroitin sulphate proteoglycan in
vitro and axonal regeneration in the adult optic nerve in vivo. J
Neurochem. 103:181–189. 2007.PubMed/NCBI
|
|
30
|
Howland JG, Cazakoff BN and Zhang Y:
Altered object-in-place recognition memory, prepulse inhibition,
and locomotor activity in the offspring of rats exposed to a viral
mimetic during pregnancy. Neuroscience. 201:184–198. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang R, Xue G, Wang S, Zhang L, Shi C and
Xie X: Novel object recognition as a facile behavior test for
evaluating drug effects in AβPP/PS1 Alzheimer's disease mouse
model. J Alzheimers Dis. 31:801–812. 2012.PubMed/NCBI
|
|
32
|
Vorhees CV and Williams MT: Morris water
maze: Procedures for assessing spatial and related forms of
learning and memory. Nat Protoc. 1:848–858. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Khazipov R, Khalilov I, Tyzio R, Morozova
E, Ben-Ari Y and Holmes GL: Developmental changes in GABAergic
actions and seizure susceptibility in the rat hippocampus. Eur J
Neurosci. 19:590–600. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moshé SL, Albala BJ, Ackermann RF and
Engel J Jr: Increased seizure susceptibility of the immature brain.
Brain Res. 283:81–85. 1983. View Article : Google Scholar : PubMed/NCBI
|