Introduction
Peritoneal dialysis (PD) is a primary treatment of
end-stage renal disease (ESRD). High glucose (HG) damages the
structure and function of the peritoneal membrane, which leads to
peritoneal fibrosis and ultrafiltration failure via inflammation,
apoptosis, oxidative stress and epithelial-mesenchymal transition
(EMT) (1–3). Recent studies have also revealed that
HG regulates autophagy in certain diseases (4–6).
Autophagy, which functions to maintain cell
homeostasis under various stress conditions, primarily degrades and
recycles misfolded proteins and damaged organelles (7). Previous studies have demonstrated
that HG modulates autophagy in multiple disease states (8–10).
However, the direct effects of autophagy on HG-induced peritoneal
mesothelial cells remain unknown. Macroautophagy, which is
generally referred to as autophagy, is a ubiquitous, genetically
programmed process. Central to this process is the formation of
autophagosomes, which are double- or multiple-membrane cytoplasmic
vesicles that are responsible for delivering cytoplasmic material
to lysosomes (11). Several
autophagy genes are involved in the process of autophagy, including
microtubule-associated protein 1A/1B-light chain 3 (LC3), Beclin-1,
ubiquitin-binding protein p62 (p62) and other autophagy-associated
proteins (12,13). Autophagy serves a protective role
in certain diseases such as renal ischemia/reperfusion, cancer, EMT
and inflammation (14–16). The effects of autophagy on EMT and
inflammation have also been proposed to be involved in cancer
(17,18).
Vitamin D has been demonstrated to serve an
important role in the regulation of cell differentiation, cell
proliferation, immunomodulation, inflammation and autophagy
(19–22). 1,25(OH)2D3,
the active form of vitamin D, regulates bone, calcium and phosphate
metabolism through the vitamin D receptor (VDR). The VDR forms a
heterodimer with the retinoid X receptor and regulates gene
expression in the nucleus. Research has demonstrated that
1,25(OH)2D3 has protective effects in a
number of diseases (23–25). It was previously revealed that
1,25(OH)2D3 attenuates apoptosis, oxidative
stress and inflammation in HG treated-peritoneal mesothelial cells
(1). Therefore, the present study
hypothesized that 1,25(OH)2D3 may regulate
autophagy and protect mesothelial cells during PD.
Earlier research performed by the authors of the
present study revealed that HG induced peritoneal injury,
apoptosis, oxidative stress, inflammation and EMT, and
1,25(OH)2D3 attenuated HG-induced peritoneal
injury (1,2). In the present study, the biological
function of autophagy in HG-stimulated human peritoneal mesothelial
cells (HPMCs) and peritoneal mesothelium was investigated. It was
also determined whether 1,25(OH)2D3
attenuates HG-induced peritoneal injury via autophagy and the
molecular mechanism of this effect. The present study found that
1,25(OH)2D3 treatment may provide an improved
solution to the peritoneal fibrosis and ultrafiltration
failure.
Materials and methods
Reagents
Fetal bovine serum (FBS), penicillin, streptomycin
and RPMI-1640 were all obtained from Gibco; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA).
1,25(OH)2D3 was purchased from Sigma-Aldrich;
Merck KGaA (Darmstadt, Germany). Anti-Beclin-1 (3738), anti-LC3
(4108), anti-p62 (5114), anti-mTOR (2972), anti-p-mTOR (2971),
anti-β-actin (4967) and HRP conjugate anti-rabbit (5127) IgG
antibodies were all purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Culture of HPMCs
HPMCs were kindly provided by Professor Na Di and
Professor Xu Huimian (The First Affiliated Hospital of China
Medical University, Shenyang, China) and were cultured in RPMI-1640
medium supplemented with 10% FBS, 100 UI/ml penicillin and 100
µg/ml streptomycin. HPMCs were cultured as previously described
(1). HPMCs were dissociated for 1
min with trypsin-EDTA and the subcultivation ratio was 1:3 or 1:4.
Cells at passage 5–10 were used in all experiments. A concentration
of 10−7 mol/l 1,25(OH)2D3 was used
in the present study, according to previous published research
(1). HPMCs were classified into
the following four groups: Control group; 10−7 mol/l
1,25(OH)2D3 treatment only group; 126 mM HG
treatment only group; 10−7 mol/l
1,25(OH)2D3 pretreatment followed by 126 mM
HG incubation.
Animals and experimental
treatments
All animal procedures were approved by the
Experimental Animals Ethics Committee of China Medical University
(Liaoning, China). Kunming male mice (28–30 g, aged 8–12 weeks)
were purchased from the Department of Laboratory Animals, China
Medical University (Liaoning, China), and housed in a room with
controlled temperature (22°C) on a 12-h light/dark cycle. Food and
water were given ad libitum throughout the experiment. Following
one week to adapt to their new environment, 35 Kunming male mice
were randomly assigned into seven groups (n=5/group): Control group
(no dialysate or saline was infused); saline group (mice received
50 ml/kg saline by intraperitoneal (IP) injection every day for 4
weeks); low dose vitamin D group [the mice were subjected to IP
injections of 1 µg/kg 1,25(OH)2D3 once weekly
(every Monday) for 4 weeks]; high dose vitamin D group [the mice
were subjected to IP injections of 5 µg/kg
1,25(OH)2D3 once weekly (every Monday) for 4
weeks]; the peritoneal dialysis (PD) group [injected IP with
conventional 4.25% peritoneal dialysis fluid (PDF; Baxter
Healthcare Ltd., Guangzhou, China) 50 ml/kg daily for 4 weeks]; the
PD + low dose vitamin D group [mice were injected IP with
conventional 4.25% PDF 50 ml/kg daily, then were subjected to IP
injections of 1 µg/kg 1,25(OH)2D3 once weekly
(every Monday) for 4 weeks]; the PD + high dose vitamin D group
[mice were injected IP with conventional 4.25% PDF 50 ml/kg daily,
then were subjected to IP injections of 5 µg/kg
1,25(OH)2D3 once weekly (every Monday) for 4
weeks]. At the end of the experimental period (4 weeks), the mice
were starved for 12 to 13 h and then sacrificed by cervical
dislocation; the visceral peritoneum was used for western
blotting.
Transmission electron microscopy
HPMCs (2×107) were fixed at room
temperature with 2.5% glutaraldehyde for 2 h, then washed with 0.1
M PBS. The samples were then postfixed in 1% osmium tetroxide for 2
h at room temperature, dehydrated in a series of graded ethanol,
and embedded in Epon 812 (Structure Probe, Inc., West Chester, PA,
USA). Ultra-thin sections were cut (70 nm), stained with 2% uranyl
acetate and lead citrate for 20 min at room temperature, and
examined using transmission electron microscopy.
Western blotting
Western blot analysis was carried out as previously
described (1). All experiments
were repeated ≥3 times. Representative protein bands and
densitometric data are displayed in the figures. Total proteins
were extracted using lysis buffer (Pierce; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), quantified with the BCA
Protein assay kit (Pierce; Thermo Fisher Scientific, Inc.) and
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). Protein (50 µg) was transferred onto
polyvinylidene fluoride (PVDF) membranes (EMD Millipore, Billerica,
MA, USA). BSA solution (10%; Sigma-Aldrich; Merck KGaA) was used to
reduce non-specific antibody binding at 37°C for 1 h. The blots
were incubated overnight at 4°C with antibodies against
anti-Beclin-1 (3738, 1:1,000), anti-LC3 (4108, 1:1,000), anti-p62
(5114, 1:1,000), anti-mTOR (2972, 1:1,000), anti-p-mTOR (2971,
1:1,000) and anti-β-actin (4967, 1:1,000) (all from Cell Signaling
Technology, Inc.). Subsequently, samples were incubated with HRP
conjugate anti-rabbit immunoglobulin G antibodies (5127, 1:1,000;
Cell Signaling Technology, Inc.) at 37°C for 2 h. Target proteins
on the PVDF membrane were visualized using ECL Western Blotting
Substrate (Pierce; Thermo Fisher Scientific, Inc.) and captured
using a DNR bioimaging system (DNR Bio-Imaging Systems, Ltd., Neve
Yamin, Israel).
Statistical analysis
Statistical analysis was performed using SPSS
software version 18 (SPSS, Inc., Chicago, IL, USA). Data are
presented as the mean ± standard error. Multiple group comparisons
were made using a one-way analysis of variance followed by Tukey's
multiple comparison test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Influence of
1,25(OH)2D3 on HG-induced autophagy
inhibition in HPMCs
To assess the potential underlying mechanism by
which 1,25(OH)2D3 regulates HG-induced
peritoneal injury in HPMCs, cell autophagy was determined by
transmission electron microscopy. Compared with the HG group, the
number of autophagosomes was enhanced in the HG +
1,25(OH)2D3 group (Fig. 1A). To further confirm the influence
of 1,25(OH)2D3 on autophagy, western blotting
was performed to detect Beclin-1, LC3-II and p62 expression.
Compared to the control group, HG downregulated the expression
levels of Beclin-1 and LC3-II, and upregulated the expression
levels of p62 (Fig. 1B-E).
However, it was revealed that 1,25(OH)2D3
itself induced autophagy, and HG-induced autophagy inhibition may
be attenuated by co-treatment with 10−7 mol/l
1,25(OH)2D3.
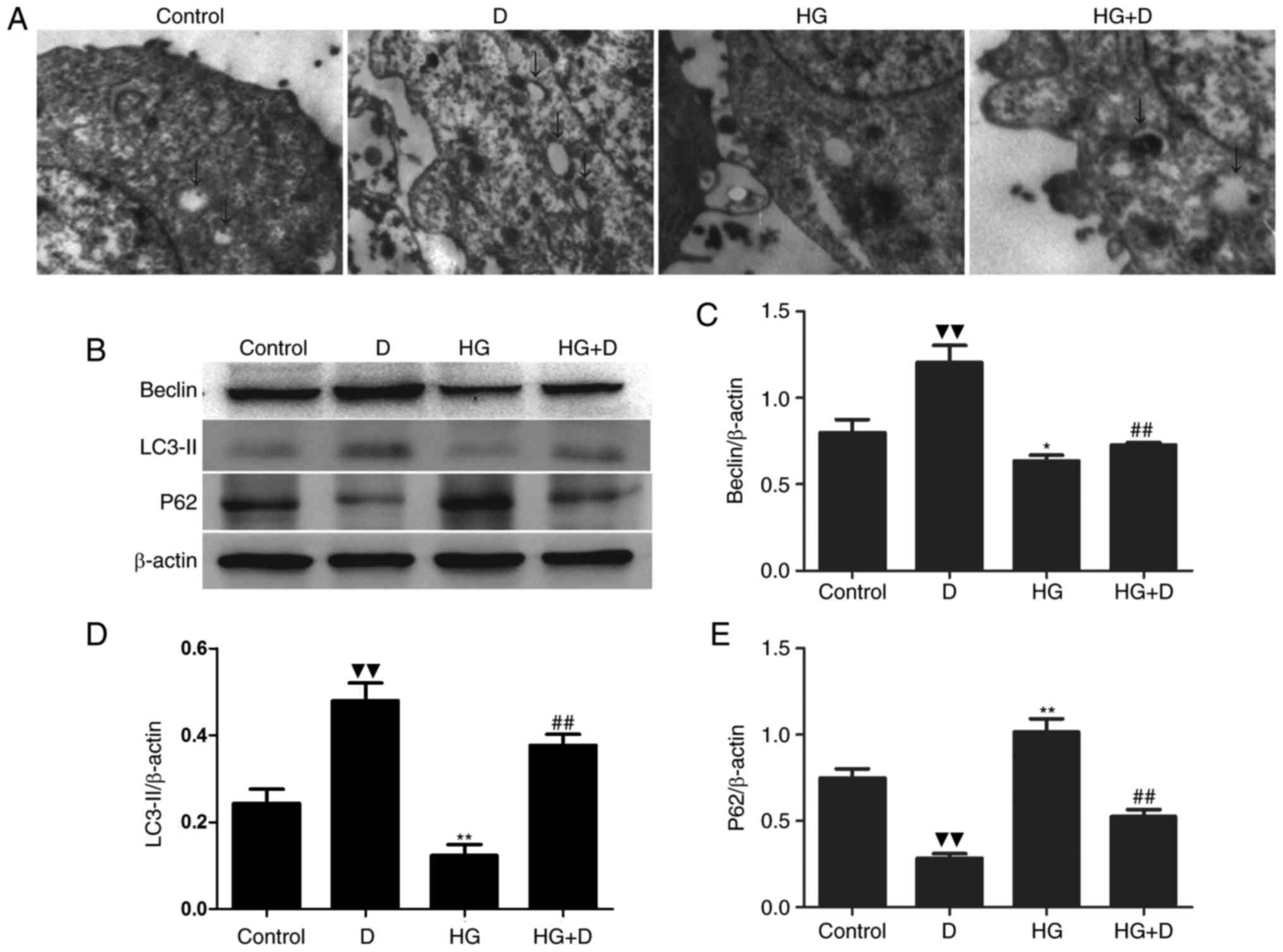 | Figure 1.1,25(OH)2D3
attenuated HG-induced autophagy inhibition. (A) HPMCs were exposed
to 126 mM HG in the presence or absence of 10−7 mol/l
1,25(OH)2D3 pretreatment. Representative
transmission electron microscopy (×4,000) images are displayed.
Arrows, autophagosomes. (B) Western blotting was performed with the
antibodies of Beclin-1, LC3-II and p62. (C-E) Protein expression
levels were assessed using densitometry and are expressed as
relative intensities. Each value represents the mean ± standard
error (n=3). ▼▼P<0.01 vs. control; *P<0.05 vs.
control; **P<0.01 vs. control; ##P<0.01 vs. HG
group. HG, high glucose; HPMCs, human peritoneal mesothelial cells;
LC3-II, microtubule-associated proteins 1A/1B light chain 3B; p62,
ubiquitin-binding protein; D, 1,25(OH)2D3; HG
+ D, high glucose and 1,25(OH)2D3. |
Effects of
1,25(OH)2D3 on the mTOR pathway in HG-treated
HPMCs
The mTOR signaling pathway has been reported to be
involved in autophagy and EMT (17); however, whether
1,25(OH)2D3 regulates autophagy via the mTOR
pathway in HPMCs remains unknown. The present study therefore
examined the effect of 1,25(OH)2D3 on the
mTOR signaling pathway in HPMCs.
mTOR activation was measured by western blotting
with a p-mTOR antibody. As demonstrated in Fig. 2A and B, when cells were exposed to
HG alone, mTOR phosphorylation was increased compared to the
control group, whereas p-mTOR expression was significantly
decreased when cells were co-treated with
1,25(OH)2D3. These results suggested that
1,25(OH)2D3 regulates autophagy via the mTOR
pathway in HPMCs.
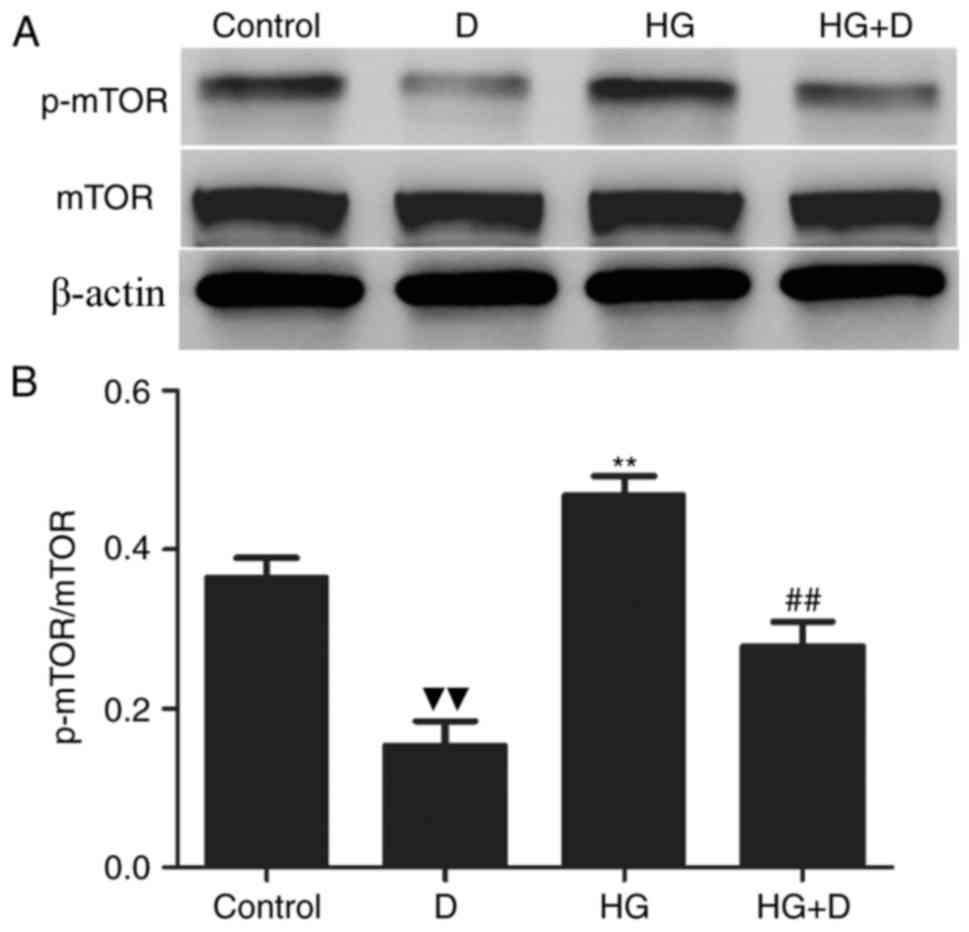 | Figure 2.Effects of
1,25(OH)2D3 on the mTOR signaling pathway in
HG-treated HPMCs. HPMCs were exposed to 126 mM HG in the presence
or absence of 10−7 mol/l
1,25(OH)2D3 pretreatment. (A) Western
blotting was performed with mTOR and p-mTOR antibodies. (B) Protein
expression levels were assessed using densitometry and were
expressed as relative intensities. Each value represents the mean ±
standard error (n=3). ▼▼P<0.01 vs. control;
**P<0.01 vs. control; ##P<0.01 vs. HG group. mTOR,
mechanistic target of rapamycin; HG, high glucose; HPMCs, human
peritoneal mesothelial cells; p-mTOR, phosphorylated mechanistic
target of rapamycin, D, 1,25(OH)2D3; HG + D,
high glucose and 1,25(OH)2D3. |
Influence of
1,25(OH)2D3 on HG-induced autophagy
inhibition in peritoneal mesothelium
A previously published report by the authors of the
present study revealed that 1,25(OH)2D3
protects HPMCs from inflammation and apoptosis (1). The present study used a mouse model
to further assess the underlying mechanism by which
1,25(OH)2D3 influences HG induced peritoneal
injury. Compared to the control group, HG PDF significantly
downregulated the expression levels of Beclin-1 and LC3-II, and
upregulated the expression levels of p62 (Fig. 3A-D). However,
1,25(OH)2D3 induced autophagy, and HG-induced
autophagy inhibition may be attenuated by co-treatment with 1 and 5
µg/kg 1,25(OH)2D3.
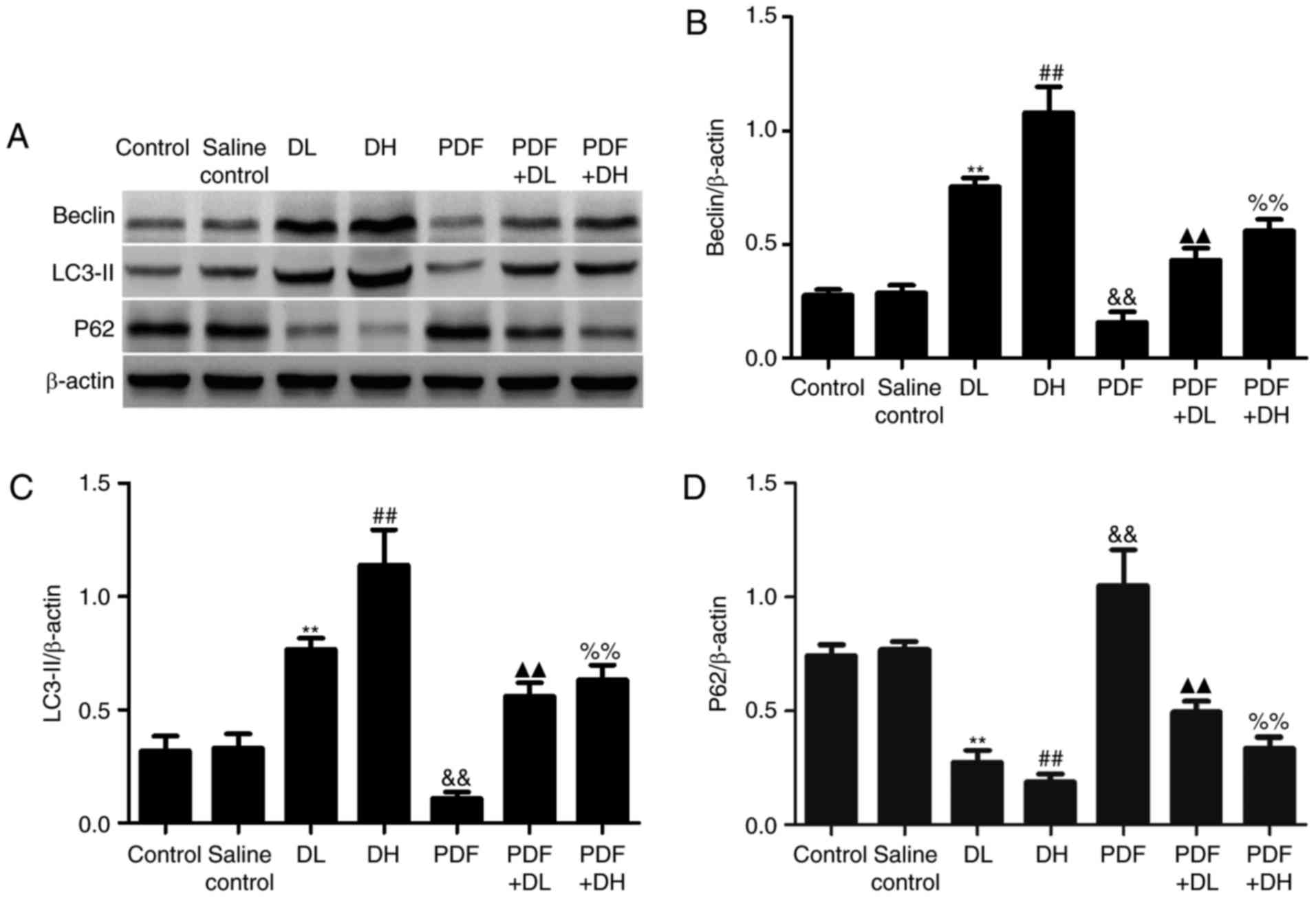 | Figure 3.Effects of
1,25(OH)2D3 on autophagy in HG PDF-treated
peritoneal mesothelium. DL, vitamin D low-dose group [1 µg/kg
1,25(OH)2D3]; DH, vitamin D high-dose group
[5 µg/kg 1,25(OH)2D3]; PDF + DL, PDF + low
dose vitamin D [1 µg/kg 1,25(OH)2D3] group;
PDF + DH, PDF + high-dose vitamin D [5 µg/kg
1,25(OH)2D3] group. (A) Western blotting was
performed using the antibodies of Beclin-1, LC3-II and p62. (B-D)
Relative expression levels of Beclin, LC3-II and p62 were
calculated and normalized to the loading control by densitometric
analysis. Each value represents the mean ± standard error (n=5).
**P<0.01 vs. control; ##P<0.01 vs. control;
&&P<0.01 vs. control; ▲▲P<0.01
vs. PDF; %%P<0.01 vs. PDF. HG, high glucose; PDF,
peritoneal dialysis fluid; LC3-II, microtubule-associated proteins
1A/1B light chain 3B; p62, ubiquitin-binding protein. |
Effects of
1,25(OH)2D3 on the mTOR pathway in HG-treated
peritoneal mesothelium
A mouse model was used to further assess the
association of the mTOR signaling pathway and autophagy. When mice
were exposed to HG PDF, mTOR phosphorylation was significantly
increased compared with the control group; however, p-mTOR was
decreased when co-treated with 1,25(OH)2D3.
These results suggested that 1,25(OH)2D3
regulates autophagy via the mTOR signaling pathway in peritoneal
mesothelium.
Discussion
PD is generally accepted as an important renal
replacement therapy for the treatment of ESRD. However, long-term
PD may lead to peritoneal membrane failure. A previous study
published by the authors of the present study indicated that HG
induced peritoneal inflammation, apoptosis, oxidative stress and
EMT, which were involved in the development of peritoneal
dysfunction (1,2). Studies have demonstrated that HG may
also affect autophagy in certain diseases (8–10).
Autophagy is emerging as a key factor in various
physiological and pathological events. A previous study showed that
autophagy serves a protective role in diseases such as renal
ischemia/reperfusion, cancer, fibrosis and inflammation (14–16,26).
It is understood that high glucose levels may affect autophagy in
numerous cell types (9,27,28).
Accumulating evidence suggests that
1,25(OH)2D3 affects organ fibrosis, exhibits
antioxidant properties and induces autophagic capabilities
(17,29,30).
Research has also demonstrated that
1,25(OH)2D3 may induce autophagy in human
monocytes and macrophages (17).
Previous research has focused on the autophagic induction of
vitamin D3 primarily in cancer cells (31–33),
suggesting the potential use of vitamin D3 as an anticancer drug.
However, the pathophysiological role of autophagy in peritoneal
injury and whether 1,25(OH)2D3 regulates
autophagy in peritoneum remains unknown.
In the present study, in vitro and in
vivo experiments revealed that high glucose induced autophagic
inhibition, due to increased expression of Beclin and LC3-II, and
decreased expression of p62. However,
1,25(OH)2D3 induced autophagy and attenuated
HG-induced autophagy inhibition in HPMCs and peritoneal
mesothelium. Earlier research by the authors of the present study
demonstrated that HG induced peritoneal injury includes apoptosis,
oxidative stress, inflammation and EMT, and
1,25(OH)2D3 attenuated HG induced peritoneal
injury (1,2). In the present study, HG decreased
autophagy and 1,25(OH)2D3 induced autophagy.
Therefore, HG may induce peritoneal injury by decreasing autophagy
and 1,25(OH)2D3 exhibits a protective effect
by increasing autophagy.
The mTOR signaling pathway is a classic pathway in
autophagy that has been extensively studied (18,34).
A previous study demonstrated that
1,25(OH)2D3 protected β cells in the pancreas
against high glucose-induced apoptosis via the suppression of the
mTOR signaling pathway (35). Jang
et al (36) found that
vitamin D protects against rotenone-induced neurotoxicity by
enhancing autophagy via the mTOR pathway. In the present study, the
association between the mTOR signaling pathway and peritoneal
autophagy after HG treatment was investigated. Results demonstrated
that treatment with HG activated the mTOR pathway, induced
peritoneal autophagy inhibition, and these alterations may be
attenuated by 1,25(OH)2D3 pretreatment.
In conclusion, it was found that HG induced
autophagy inhibition in peritoneum,
1,25(OH)2D3 induced autophagy and attenuated
the HG-induced autophagy inhibition in peritoneum, possibly via the
mTOR signaling pathway. Further investigation in this area will
generate novel insights into the critical role of
1,25(OH)2D3, and will provide an experimental
basis for its clinical use in the treatment of PD. One limitation
of the present study is that we only measured mTOR and p-mTOR,
which is only one protein involved in the mTOR signaling pathway.
Measuring the expression levels of other proteins in this pathway
will be considered for future studies.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81300636 and
81370865).
References
|
1
|
Yang L, Wu L, Du S, Hu Y, Fan Y and Ma J:
1,25(OH)2D3 inhibit high glucose-induced apoptosis and ROS
production in human peritoneal mesothelial cells via the MAPK/P38
pathway. Mol Med Rep. 14:839–844. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang X, Wang J, Fan Y, Yang L, Wang L and
Ma J: Zinc supplementation attenuates high glucose-induced
epithelial-to-mesenchymal transition of peritoneal mesothelial
cells. Biol Trace Elem Res. 150:229–235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kanasaki K, Taduri G and Koya D: Diabetic
nephropathy: The role of inflammation in fibroblast activation and
kidney fibrosis. Front Endocrinol (Lausanne). 4:72013.PubMed/NCBI
|
|
4
|
Zhang C, Hou B, Yu S, Chen Q, Zhang N and
Li H: HGF alleviates high glucose-induced injury in podocytes by
GSK3β inhibition and autophagy restoration. Biochim Biophys Acta.
1863:2690–2699. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang PW, Tian C, Xu FY, Chen Z, Burnside
R, Yi WJ, Xiang SY, Xie X, Wu NN, Yang H, et al: Green tea
polyphenols alleviate autophagy inhibition induced by high glucose
in endothelial cells. Biomed Environ Sci. 29:524–528.
2016.PubMed/NCBI
|
|
6
|
de Faria JM Lopes, Duarte DA, Montemurro
C, Papadimitriou A, Consonni SR and de Faria JB Lopes: Defective
autophagy in diabetic retinopathy. Invest Ophthalmol Vis Sci.
57:4356–4366. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang TC, Hsu MF and Wu KK: High glucose
induces bone marrow-derived mesenchymal stem cell senescence by
upregulating autophagy. PLoS One. 10:e01265372015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu X, Fan Q, Xu L, Li L, Yue Y, Xu Y, Su
Y, Zhang D and Wang L: Ursolic acid attenuates diabetic mesangial
cell injury through the up-regulation of autophagy via
miRNA-21/PTEN/Akt/mTOR suppression. PLoS One. 10:e01174002015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wei M, Li Z and Yang Z: Crosstalk between
protective autophagy and NF-κB signal in high glucose-induced
podocytes. Mol Cell Biochem. 394:261–273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xie Z and Klionsky DJ: Autophagosome
formation: Core machinery and adaptations. Nat Cell Biol.
9:1102–1109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ohsumi Y: Molecular dissection of
autophagy: Two ubiquitin-like systems. Nat Rev Mol Cell Biol.
2:211–216. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Suzuki K, Kirisako T, Kamada Y, Mizushima
N, Noda T and Ohsumi Y: The pre-autophagosomal structure organized
by concerted functions of APG genes is essential for autophagosome
formation. EMBO J. 20:5971–5981. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang M, Liu K, Luo J and Dong Z:
Autophagy is a renoprotective mechanism during in vitro hypoxia and
in vivo ischemia-reperfusion injury. Am J Pathol. 176:1181–1192.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang PM, Liu YL, Lin YC, Shun CT, Wu MS
and Chen CC: Inhibition of autophagy enhances anticancer effects of
atorvastatin in digestive malignancies. Cancer Res. 70:7699–7709.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Degenhardt K, Mathew R, Beaudoin B, Bray
K, Anderson D, Chen G, Mukherjee C, Shi Y, Gélinas C, Fan Y, et al:
Autophagy promotes tumor cell survival and restricts necrosis,
inflammation, and tumorigenesis. Cancer Cell. 10:51–64. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Catalano M, D'Alessandro G, Lepore F,
Corazzari M, Caldarola S, Valacca C, Faienza F, Esposito V,
Limatola C, Cecconi F and Di Bartolomeo S: Autophagy induction
impairs migration and invasion by reversing EMT in glioblastoma
cells. Mol Oncol. 9:1612–1625. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ding YH, Zhou ZW, Ha CF, Zhang XY, Pan ST,
He ZX, Edelman JL, Wang D, Yang YX, Zhang X, et al: Alisertib, an
Aurora kinase A inhibitor, induces apoptosis and autophagy but
inhibits epithelial to mesenchymal transition in human epithelial
ovarian cancer cells. Drug Des Devel Ther. 9:425–464.
2015.PubMed/NCBI
|
|
19
|
Gocek E, Kiełbiński M, Wyłób P, Kutner A
and Marcinkowska E: Side-chain modified vitamin D analogs induce
rapid accumulation of VDR in the cell nuclei proportionately to
their differentiation-inducing potential. Steroids. 73:1359–1366.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Foroozanfard F, Jamilian M, Bahmani F,
Talaee R, Talaee N, Hashemi T, Nasri K, Asemi Z and Esmaillzadeh A:
Calcium plus vitamin D supplementation influences biomarkers of
inflammation and oxidative stress in overweight and vitamin
D-deficient women with polycystic ovary syndrome: A randomized
double-blind placebo-controlled clinical trial. Clin Endocrinol
(Oxf). 83:888–894. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fischer KD and Agrawal DK: Vitamin D
regulating TGF-β induced epithelial-mesenchymal transition. Respir
Res. 15:1462014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yuk JM, Shin DM, Lee HM, Yang CS, Jin HS,
Kim KK, Lee ZW, Lee SH, Kim JM and Jo EK: Vitamin D3 induces
autophagy in human monocytes/macrophages via cathelicidin. Cell
Host Microbe. 6:231–243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Uberti F, Morsanuto V, Bardelli C and
Molinari C: Protective effects of 1α,25-dihydroxyvitamin D3 on
cultured neural cells exposed to catalytic iron. Physiol Rep.
4:e127692016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lawrence DW and Sharma B: A review of the
neuroprotective role of vitamin D in traumatic brain injury with
implications for supplementation post-concussion. Brain Inj.
30:960–968. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang L, Ma J, Zhang X, Fan Y and Wang L:
Protective role of the vitamin D receptor. Cell Immunol.
279:160–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lodder J, Denaës T, Chobert MN, Wan J,
El-Benna J, Pawlotsky JM, Lotersztajn S and Teixeira-Clerc F:
Macrophage autophagy protects against liver fibrosis in mice.
Autophagy. 11:1280–1292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xie Z, Lau K, Eby B, Lozano P, He C,
Pennington B, Li H, Rathi S, Dong Y, Tian R, et al: Improvement of
cardiac functions by chronic metformin treatment is associated with
enhanced cardiac autophagy in diabetic OVE26 mice. Diabetes.
60:1770–1778. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen F, Chen B, Xiao FQ, Wu YT, Wang RH,
Sun ZW, Fu GS, Mou Y, Tao W, Hu XS and Hu SJ: Autophagy protects
against senescence and apoptosis via the RAS-mitochondria in
high-glucose-induced endothelial cells. Cell Physiol Biochem.
33:1058–1074. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu L, Kong M, Han YP, Bai L, Zhang X,
Chen Y, Zheng S, Yuan H and Duan Z: Spontaneous liver fibrosis
induced by long term dietary vitamin D deficiency in adult mice is
related to chronic inflammation and enhanced apoptosis. Can J
Physiol Pharmacol. 93:385–394. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Greń A: Effects of vitamin EC and D
supplementation on inflammation and oxidative stress in
streptozotocin-induced diabetic mice. Int J Vitam Nutr Res.
83:168–175. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Høyer-Hansen M, Bastholm L, Mathiasen IS,
Elling F and Jäättelä M: Vitamin D analog EB1089 triggers dramatic
lysosomal changes and Beclin 1-mediated autophagic cell death. Cell
Death Differ. 12:1297–1309. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Demasters G, Di X, Newsham I, Shiu R and
Gewirtz DA: Potentiation of radiation sensitivity in breast tumor
cells by the vitamin D3 analogue, EB 1089, through promotion of
autophagy and interference with proliferative recovery. Mol Cancer
Ther. 5:2786–2797. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang J, Lian H, Zhao Y, Kauss MA and
Spindel S: Vitamin D3 induces autophagy of human myeloid leukemia
cells. J Biol Chem. 283:25596–25605. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang X, Howell GM, Guo L, Collage RD,
Loughran PA, Zuckerbraun BS and Rosengart MR: CaMKIV-dependent
preservation of mTOR expression is required for autophagy during
lipopolysaccharide-induced inflammation and acute kidney injury. J
Immunol. 193:2405–2415. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang Z, Liu F, Qu H, Wang H, Xiao X and
Deng H: 1, 25(OH)2D3 protects β cell against high glucose-induced
apoptosis through mTOR suppressing. Mol Cell Endocrinol.
414:111–119. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jang W, Kim HJ, Li H, Jo KD, Lee MK, Song
SH and Yang HO: 1,25-Dyhydroxyvitamin D3 attenuates
rotenone-induced neurotoxicity in SH-SY5Y cells through induction
of autophagy. Biochem Biophys Res Commun. 451:142–147. 2014.
View Article : Google Scholar : PubMed/NCBI
|














