Introduction
Colon cancer is one of the most common malignant
tumors, with a high incident rate in the 40–50 age group. Colon
cancer affects ~150,000 patients in the USA annually (1). Due to the changing of diets, colon
cancer has become the 4th cause for malignant tumor mortality in
China and there are ~140,000 diagnosed cases annually (2). Surgery is the primary therapy for
colon cancer and patients exhibit 5-year survival rate of 50%
following surgery (3).
However, 15–20% patients experience recurrence
following treatment. Tumor recurrence following curative surgery is
a major hindrance for the improvement of overall survival (4). Therefore, it is important to identify
the molecular changes in patients and to determine the underlying
reason for colon cancer recurrence. Biomarkers have been used as
tools in the detection and management of the disease in patients
with colon cancer (5). For
instance, the CpG island methylator phenotype is independently
associated with an unfavorable prognosis in patients with colon
cancer (6). Epithelial cell
adhesion molecule, cluster of differentiation (CD)26, musashi RNA
binding protein 1, CD29, CD24, leucine rich repeat containing G
protein-coupled receptor 5 and aldehyde dehydrogenase 1 family
member A1 have been identified as potential putative markers for
colon cancer (7,8). DNA methylation may also predict
recurrence of resected stage III proximal colon cancer (9). MicroRNA-93 inhibited the early
relapse of colon cancer by targeting cell cycle-associated genes
(10).
Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit a
mutation in colorectal cancer may act as a predictive molecular
biomarker for adjuvant aspirin therapy in colon cancer (11).
An improved understanding of the biology of
recurrence may improve the development of novel recurrence
prevention or treatment methods in colon cancer. In order to
investigate the recurrence-associated genes in colon cancer for
future therapy, a co-expression network of differentially expressed
genes (DEGs) in colon cancer was constructed in the present study
and the most significant modules in the network were used to reveal
the recurrence-associated genes. Subsequently, the functions of
recurrence-associated genes were enriched to determine the
importance of these genes in the relapse of patients with colon
cancer.
Materials and methods
Microarray profiles
Two microarray profiles of colon cancer samples
including recurrence information (E-GEOD-39,582 and E-GEOD-33,113)
were downloaded from ArrayExpress (http://www.ebi.ac.uk/arrayexpress/). E-GEOD-39,582
included 566 samples, based on the platform of AFFY HG-U133_Plus_2,
which were divided into the training dataset and the validation
dataset for the weighted correlation network analysis (WGCNA)
network construction. There were a total of 90 colon cancer samples
in E-GEOD-33,113, with the recurrence status and clinical
information of the samples, which was used as the validation
dataset for mining vital module associated with recurrence.
Sample classification
Samples in the profiles were classified using
Consensus Clustering in R (http://www.bioconductor.org/packages/release/bioc/html/ConsensusClusterPlus.html)
to identify their sub-types. The parameters set in this analysis
are presented in Table I.
Following the classification of samples, the association between
colon cancer samples and corresponding clinical performance
[survival, sex, cghdata, chemotherapy, mismatch repair (Mmr)
status, tumor node metastasis (TNM) stage, and Tumor location] were
determiend using the χ2 test. P<0.05 was considered
to indicate a statistically significant difference.
 | Table I.Parameters used in sample
classification using consensus clustering. |
Table I.
Parameters used in sample
classification using consensus clustering.
| Parameter | Value |
|---|
| Max cluster
group | 20 |
| Subsample
number | 5,000 |
| Proportion
sample | 0.9 |
| Proportion
feature | 1 |
| Distance
method | Pearson |
| Clustering
method | K-means dist |
Pretreatment of genes expression
data
Data in the profiles were initially normalized using
the robust multi-array analysis (RMA) method in Affy package
(http://www.bioconductor.org/packages/release/bioc/html/affy.html)
and were transformed using a log2 transformation. Probes
were converted into gene symbols and the average expression value
was used as the only value of the gene with multiple corresponding
probes. Following that, the cv method in the genefilter package was
applied to filter the genes with significant variation. Genes with
coefficient of variation >0.4 were recognized as the candidate
genes.
Screening of DEGs
The limma package in R (http://www.bioconductor.org/packages/release/bioc/html/limma.html)
was used to identify the DEGs from the candidate genes. P<0.01
and log fold change (FC) >1.5 were set as the cut-off
criteria.
Construction of the WGCNA
co-expression network
The WGCNA method (12) was used to construct the
co-expression network of the genes in the test samples of colon
cancer. The interaction coefficient between genes was calculated
using the following formula:
aij=Sijβ,Sij=|cor(xi,xj)|(1)
Where xi and xj are the
expression vectors of gene i and j, respectively. The Pearson
coefficient of these two vectors was cor, which was transited into
the interaction coefficient using Sij. This transition
intended to give more weight to the strong connections and reduce
the importance of the weak connections in the predicted
co-expression network in order to improve the reliability of the
co-expression network.
The connection coefficient will be transformed into
a weighted coefficient Wij using the following
formula:
Wij=lij+aijmin{ki,kj}+1–aij(2)lij=∑uaiuauj,ki=∑uaiu
Subsequently, the co-expression network would be
constructed based on the W matrix, followed by module mining. The
reliability of the minded modules was verified using the verify
sample set E-GEOD-33113. The topological properties were also
confirmed. Modules with module significance <0.05 were
identified to be recurrence-associated modules.
Verification of recurrence-associated
modules
The average expression value of genes in the
significant modules in each sample was calculated. Subsequently,
the samples were ranked based on the expression level of modules.
According to 1/4 and 3/4 value of expression level, the modules
were divided into high expression level, median expression level,
and low expression level. Finally, the Kaplan-Meier (KM) curves of
recurrence-associated modules expression level and recurrence
status were drawn. The significance of the different expression
level modules was compared, and P<0.05 was considered to
indicate a statistically significant difference. The identified
vital module was verified using the independent dataset
E-GEOD-33113.
Function enrichment of recurrence
associated modules
In order to determine the functions of the genes in
the recurrence-associated modules, genes were subjected to Database
for Annotation, Visualization and Integrated Discovery (13) for function and pathway enrichment.
P<0.05 was the threshold used for the significant terms.
Survival analysis of genes in
recurrence-associated modules
Survival analysis was performed on the genes in the
recurrence-associated modules. The degree (k) of a gene and the
significance (P) between each gene and sample survival time were
also calculated using a Cox regression. Subsequently, the
interaction coefficient (coef) between k and -log10(P)
was computed to identify the hub genes associated with survival.
Finally, selected genes were determined in the verifed samples for
an association with survival. Recurrence associated genes were also
subject to sample classification.
Results
Data preprocessing and sample
classification
From the 19,846 irredundant genes in E-GEOD-39582,
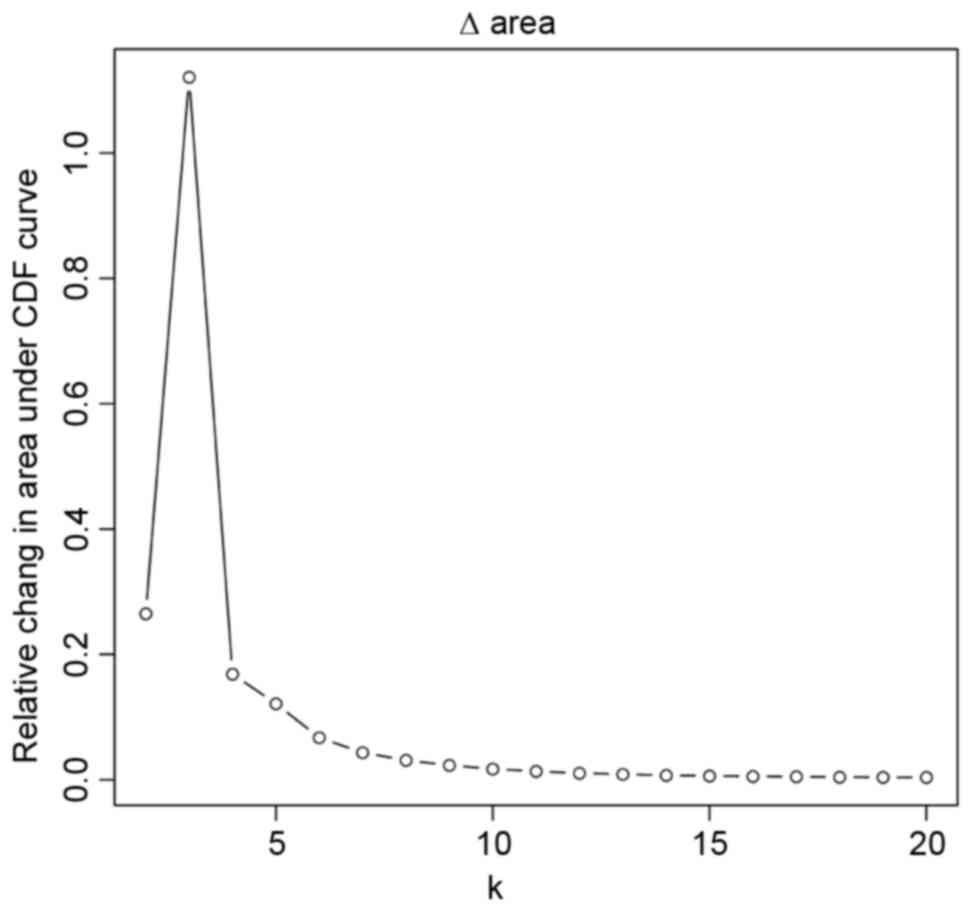
6,600 were the variation genes. Consensus Clustering analysis
revealed that cumulative distribution function (CDF) was at a high
level when there were 3 sub-types, accompanied with a satisfied
classification effect (Fig. 1).
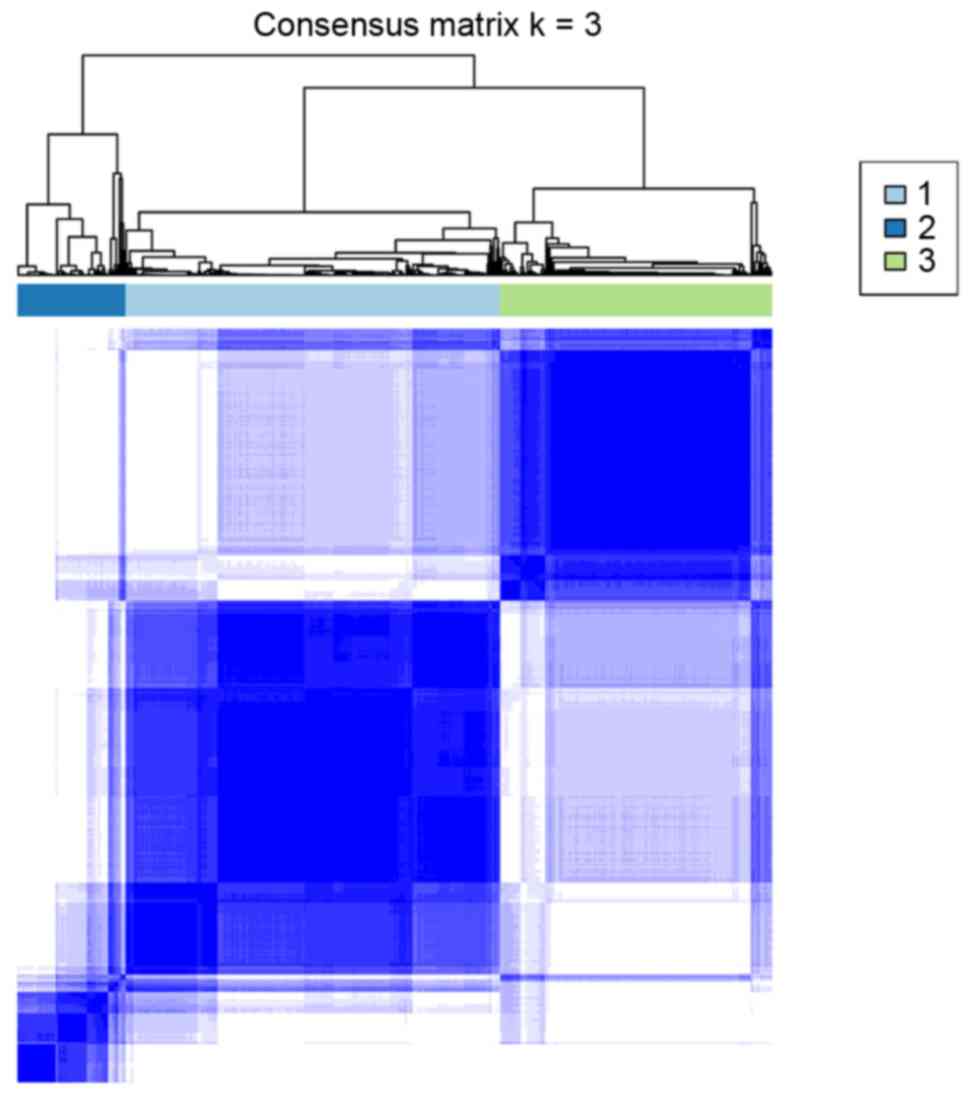
Therefore, the 556 samples in E-GEOD-39582 were divided into 3
subgroups: G1, G2, and G3 (Fig.
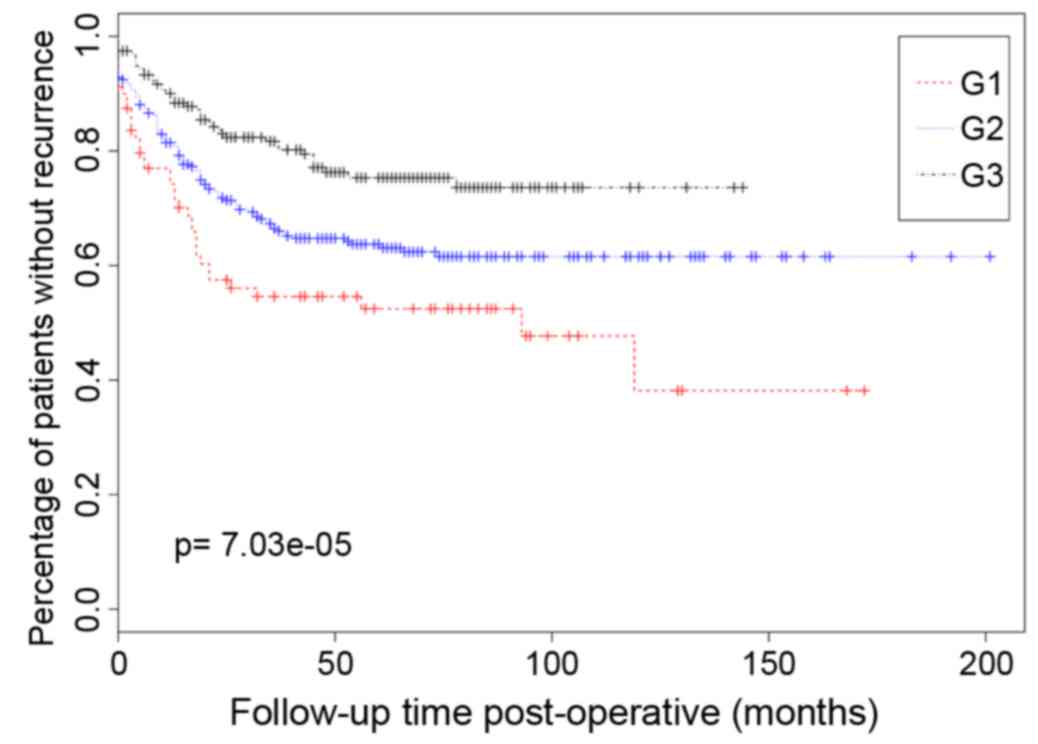
2). There were significant survival differences among these 3
types of samples, and as depicted in Fig. 3, G1 exhibited the highest survival
status, whereas G3 exhibited the lowest survival status.
Additionally, the c2 test determined that all clinical
data, with the exception of sex, were significantly different among
these 3 groups (Table II).
| Table II.Association analysis on clinical data
with sub-samples groups. |
Table II.
Association analysis on clinical data
with sub-samples groups.
| Characteristic | χ2 | df | P-value |
|---|
| Sex | 1.1653 | 2 |
5.58×10−1 |
| CGH data | 17.0540 | 2 |
1.98×10−4 |
| Chemotherapy | 19.7370 | 4 |
5.63×10−4 |
| MMR status | 23.4140 | 4 |
1.05×10−4 |
| TNM stage | 26.2990 | 8 |
9.34×10−4 |
| Tumor location | 14.7690 | 2 |
6.21×10−4 |
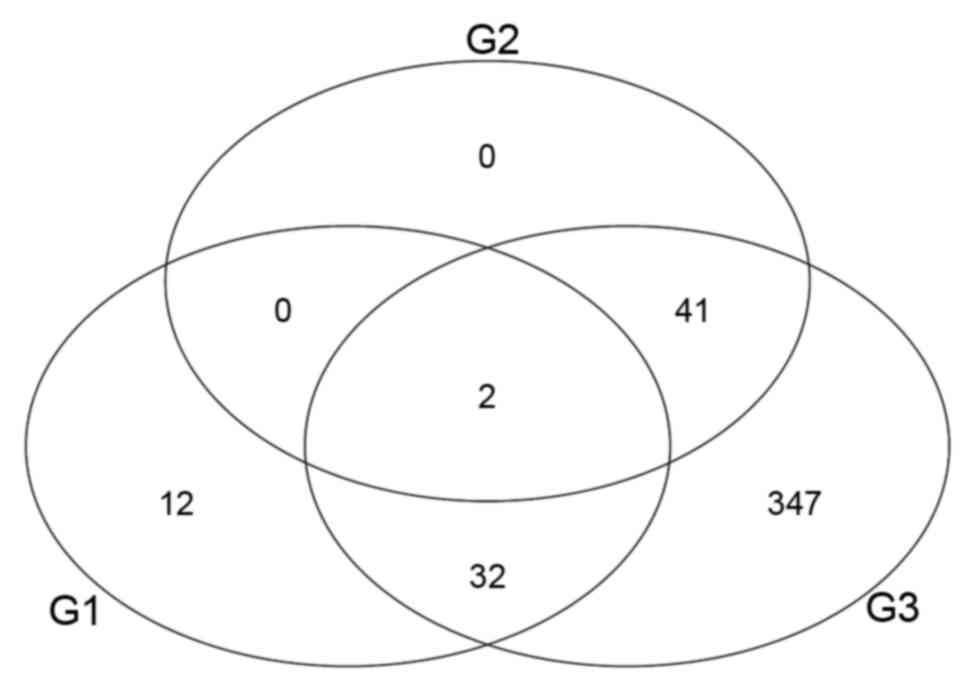
DEGs among the 3 sub-groups
From the 434 DEGs, 76 were the DEGs between G1 and
G2, 390 were the DEGs between G1 and G3 and 63 were DEGs between G2
and G3. A total of 2 DEGs were identified in all 3 groups (Fig. 4). These two common DEGs were
stress-associated endoplasmic reticulum protein family member 2
(SERP2) and long non-coding RNA-0219 (LINC0219).
Co-expression network construction and
module mining
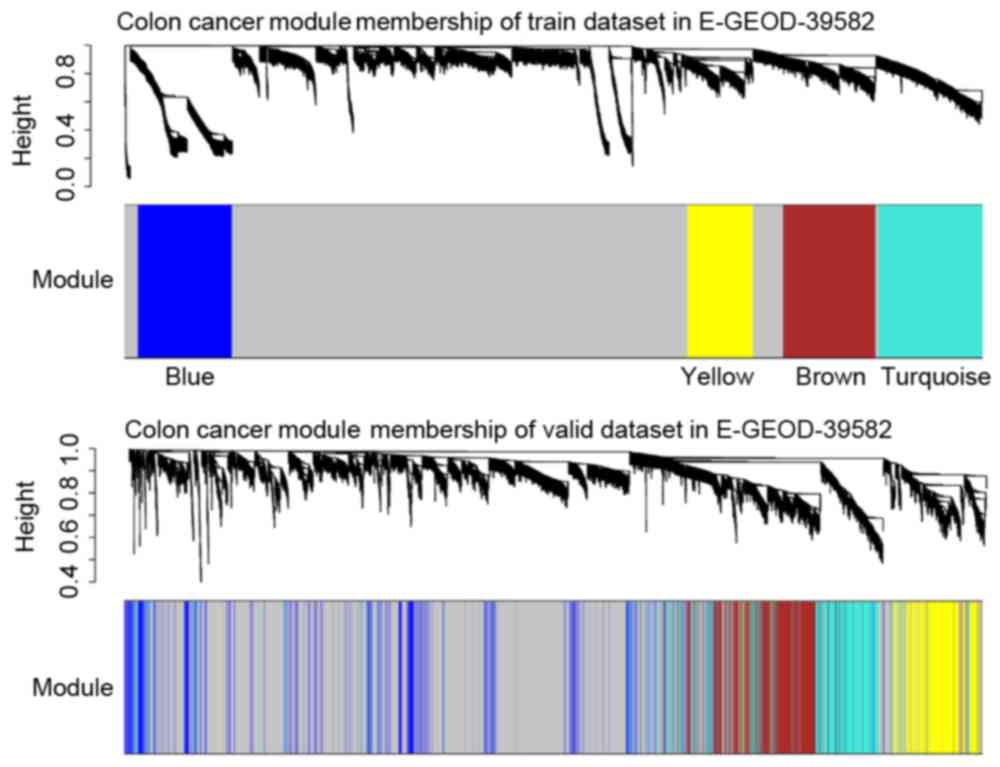
The co-expression network of the training dataset
was divided into 4 modules (Fig.
5), which were verified with the validation dataset. By
calculating the correlation coefficient, the connections between
the genes in each module and survival status of the samples were
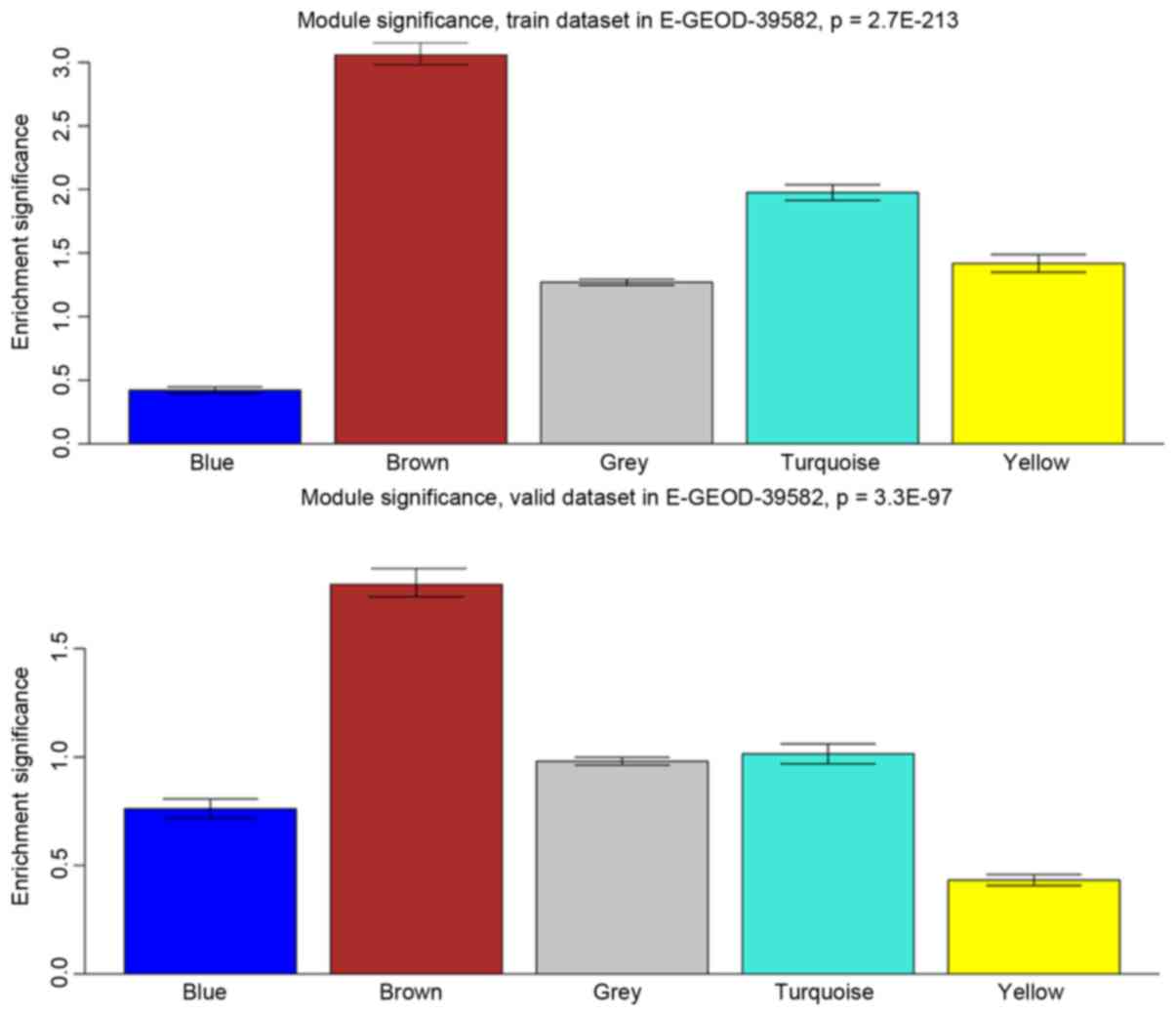
identified. It was determined that the Brown module had the highest
module significance and there were various survival-associated
genes (hub genes) in this module (Fig.
6). These connections were also verified in the validation
dataset.
Recurrence-associated module
analysis
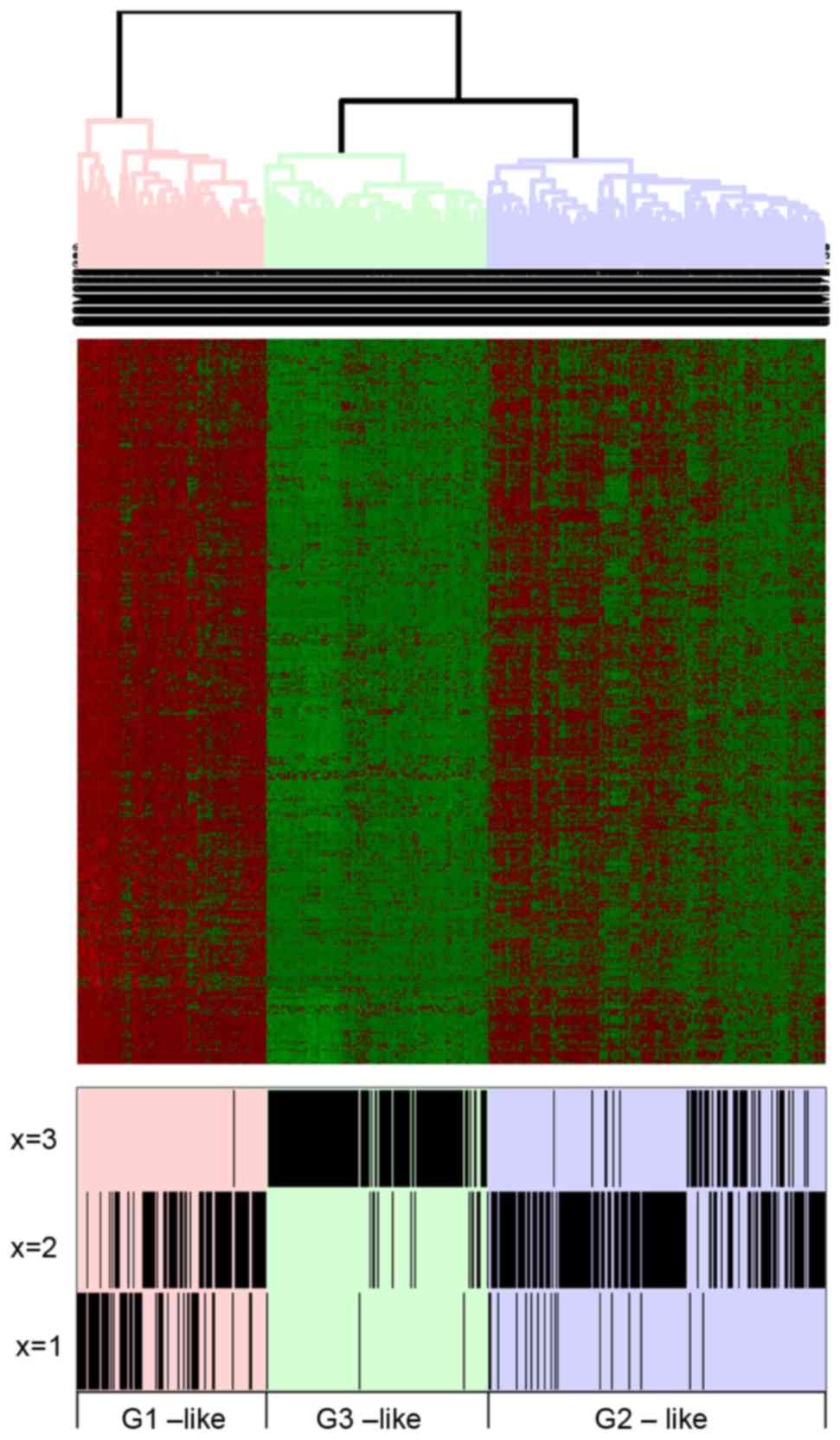
All 431 genes in the Brown module were subject to a
clustering analysis and it was revealed that there were 3 types of
samples (Fig. 7). As presented in
Fig. 7, certain G2 type samples
were also observed in with G1 and G2 type samples, suggesting it
may have a medium role for the connection between the G1 and G3
status. Additionally, the recurrence interval of G3 samples was
short, accompanied with downregulated genes.
The top 10 genes were collagen type VI α 3 chain
(COL6A3), EGF containing fibulin like extracellular matrix protein
2 (EFEMP2), fibrillin 1 (FBN1), follistatin-like 1 (FSTL1),
glycosyltransferase 8 domain containing 2 (GLT8D2), heart
development protein with EGF like domains 1 (HEG1), RAB31, member
RAS oncogene family (RAB31), secreted protein acidic and cysteine
rich (SPARC), SPARC/osteonectin, cwcv and kazal like domains
proteoglycan 1 (SPOCK1) and TIMP metallopeptidase inhibitor 2
(TIMP2) in the Brown module with higher degrees were associated
with survival (Table III). Genes
in the Brown module were primarily enriched in tumor
recurrence-associated functions and pathways, including cell
adhesion, biological adhesion, ECM organism, the ECM-receptor
interaction and the focal adhesion pathways (Table IV).
| Table III.Top 10 genes with high degrees in
Brown module. |
Table III.
Top 10 genes with high degrees in
Brown module.
| Gene name | Coefficient | P-value | k total | k within |
|---|
| COL6A3 |
2.17×10−4 |
1.9×10−6 | 136.82 | 91.86 |
| EFEMP2 |
2.10×10−3 |
9.2×10−7 | 172.24 | 110.00 |
| FBN1 |
1.42×10−3 |
5.7×10−7 | 137.03 | 92.14 |
| FSTL1 |
1.50×10−3 |
5.4×10−7 | 143.42 | 91.08 |
| GLT8D2 |
2.91×10−3 |
7.5×10−7 | 137.54 | 91.21 |
| HEG1 |
2.17×10−3 |
2.3×10−5 | 168.71 | 94.84 |
| RAB31 |
5.27×10−4 |
5.9×10−5 | 138.09 | 93.74 |
| SPARC |
3.06×10−4 |
9.8×10−8 | 132.03 | 95.57 |
| SPOCK1 |
1.49×10−3 |
9.7×10−7 | 128.75 | 88.57 |
| TIMP2 |
5.34×10−4 |
4.2×10−7 | 166.30 | 105.73 |
| Table IV.Significantly enriched functional
terms of genes in Brown module. |
Table IV.
Significantly enriched functional
terms of genes in Brown module.
| Term ID | Function | Count | P-value |
|---|
| GO:0007155 | Cell adhesion | 84 |
7.29×10−36 |
| GO:0022610 | Biological
adhesion | 84 |
8.12×10−36 |
| GO:0030198 | Extracellular
matrix organization | 30 |
2.31×10−23 |
| GO:0043062 | Extracellular
structure organization | 32 |
1.46×10−19 |
| GO:0001501 | Skeletal system
development | 40 |
2.50×10−17 |
| GO:0001944 | Vasculature
development | 35 |
1.35×10−16 |
| GO:0001568 | Blood vessel
development | 34 |
5.71×10−16 |
| GO:0009611 | Response to
wounding | 45 |
3.90×10−13 |
| GO:0006928 | Cell motion | 40 |
1.36×10−11 |
| GO:0042060 | Wound healing | 25 |
2.72×10−11 |
| GO:0016477 | Cell migration | 28 |
5.08×10−10 |
| GO:0048870 | Cell motility | 29 |
1.19×10−9 |
| GO:0051674 | Localization of
cell | 29 |
1.19×10−9 |
| GO:0048514 | Blood vessel
morphogenesis | 24 |
1.25×10−9 |
| GO:0051270 | Regulation of cell
motion | 22 |
6.93×10−9 |
| GO:0035295 | Tube
development | 23 |
1.44×10−8 |
| GO:0030334 | Regulation of cell
migration | 20 |
2.15×10−8 |
| GO:0040012 | Regulation of
locomotion | 21 |
3.35×10−8 |
| GO:0042127 | Regulation of cell
proliferation | 45 |
1.12×10−7 |
| GO:0016337 | Cell-cell
adhesion | 22 |
2.97×10−6 |
| GO:0007167 | Enzyme linked
receptor protein signaling pathway | 23 |
2.49×10−5 |
| GO:0010033 | Response to organic
substance | 36 |
5.15×10−5 |
| GO:0008285 | Negative regulation
of cell proliferation | 23 |
5.67×10−5 |
| GO:0006954 | Inflammatory
response | 21 |
1.07×10−4 |
| GO:0000902 | Cell
morphogenesis | 22 |
1.32×10−4 |
| GO:0032989 | Cellular component
morphogenesis | 23 |
2.24×10−4 |
| GO:0008284 | Positive regulation
of cell proliferation | 23 |
3.97×10−4 |
| GO:0030182 | Neuron
differentiation | 23 |
8.49×10−4 |
| hsa04512 | ECM-receptor
interaction | 26 |
1.80×10−21 |
| hsa04510 | Focal adhesion | 35 |
1.85×10−20 |
| hsa05200 | Pathways in
cancer | 22 |
3.45×10−5 |
| hsa04060 | Cytokine-cytokine
receptor interaction | 15 |
4.34×10−3 |
| hsa04810 | Regulation of actin
cytoskeleton | 13 |
5.83×10−3 |
Validation of recurrence related
modules
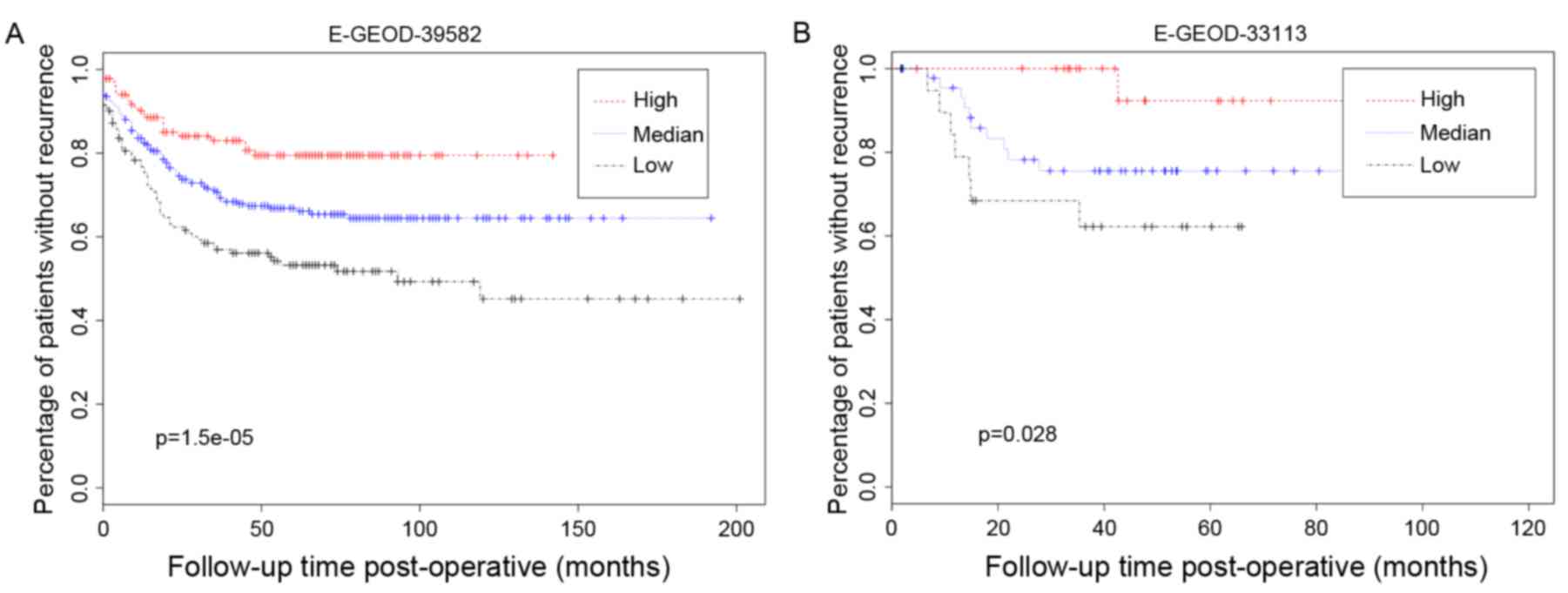
As presented in the KM curves (Fig. 8), there was a significant
difference in terms of recurrence status (P=1.5×10−6)
among the samples with high, median, and low gene expression level
of the Brown module. High expression levels inidcated higher
incidence of recurrence, whereas low expression levels show lower
recurrence incidence. This association between gene expression
level and recurrence incidence was validated in the dataset of
E-GEOD-33113 (P=2.8×10−2).
Discussion
Classification of the colon samples was downloaded
from ArrayExpress revealed that there were significant differences
of survival status and clinical data among the 3 subtype samples.
From the 434 DEGs, SERP2 and LINC0129 were the common DEGs of the 3
subgroups, suggesting they may have an important role in the
recurrence of colon cancer. The Brown module was the
recurrence-associated module in the co-expression network of DEGs
and the top 10 genes (COL6A3, EFEMP2, FBN1, FSTL1, GLT8D2, HEG1,
RAB31, SPARC, SPOCK1, and TIMP2) in this module with higher degrees
were demonstrated to be significantly associated with survival.
Enrichment analysis revealed that genes in the Brown module were
primarily enriched in tumor recurrence-associated functions and
pathways, including cell and biological adhesion, ECM organization,
the ECM-receptor interaction, and the focal adhesion pathways.
Additionally, the association between the module and tumor
recurrence were verified in another dataset E-GEOD-33113.
SERP2 belongs to the serine proteinase inhibitor
family, which are key regulators for the biological pathways that
initiate coagulation, inflammation, angiogenesis, apoptosis,
complement activation response and ECM composition (14). SERP2 methylation was identified to
be a marker for the detection and diagnosis of colon cancer
(15). Expression of SERP2 is
reported to be an early event in colon cancer, and is associated
with carcinogenesis and its development (16). LINC0129 is a long non-coding RNA
(lincRNA) and lincRNAs are RNAs >200 nt, which are not
translated into proteins. Dysfunctions of lincRNAs have been
associated with cancer. A previous study revealed that the
downregulation of lincRNA BRAF-activated non-protein coding RNA may
promote the proliferation of colorectal cancer cells (17). Another previous study reported that
lincRNA HOX transcript antisense RNA expression may be a poor
prognosis indicator in colon cancer (18). Additionally, overexpression of
lincRNA prostate cancer associated transcript 1 was identified to
be a novel biomarker of poor prognosis in patients with colon
cancer (19). However, there are
currently no direct findings that have determined the connection
between LINC0129 and colon cancer, all aforementioned findings may
have suggested that it may have an important role in colorectal
cancer by contributing to the process of relapse.
EFEMP2 is a serum biomarker for the early detection
of colon cancer (20) and a
superior biomarker compared with carcinoembryonic-antigen, which is
the sole biomarker currently used for the diagnosis and treatment
monitor in colon cancer (21).
FBN1 is a component of the extracellular microfibril and the
hypermethylation status of its promoter is a specific and sensitive
biomarker for colon cancer (22).
SPARC is a matricellular protein involved in cell migration,
angiogenesis and tissue remodeling. High SPARC expression may be
associated with an improved clinical outcome in stage II colon
cancer (23). A previous study
determined that the absence of stromal SPARC is an independent
prognostic predicator for poor prognosis of colon cancer (24). The high degrees of these genes in
the recurrence-associated modules indicated that they have
important roles in colon cancer relapse. Among the significantly
enriched pathways, the ECM-receptor interaction and focal adhesion
pathways were functionally clearly associated with the progression
and prognosis of colon cancer (25).
By constructing the co-expression network of genes
and identifying the recurrence related modules in the network, the
present study identified several survival and recurrence-associated
genes in colon cancer. These genes, including SERP2, EFEMP2, FBN1,
SPARC, and LINC0219 were identified as recurrence-associated
molecular and prognosis indicators in colon cancer.
Acknowledgements
The present study was supported by the Shanghai
Municipal Commission of Health and Family Planning (grant no.
ZYSNXD-CC-ZDYJ032), the Fund from Shanghai Science and Technology
Committee (grant no. 12401907800) and the Shanghai Science and
Technology Committee (grant no 15401931700).
References
|
1
|
American Cancer Society, . Colorectal
cancer facts & figures, 2011–2013. Atlanta, GA: American Cancer
Society; 2011
|
|
2
|
Liang Z, Baige L, Shen G, Lina S, Lingxia
L and Chengyan H: Study on the Serum Periostin Concentrations of
colon cancer patients. Chin J Lab Dia. 4:422015.
|
|
3
|
Yangming G, Chunxiao W, Minlu Z, Minlu Z,
Peng P, Kai GU, Pingping B, Zhezhou H, Yongmei X and Ying Z:
Colorectal cancer survival analysis in major areas in Shanghai.
China Oncol. 25:497–504. 2015.
|
|
4
|
Gerger A, Zhang W, Yang D, Bohanes P, Ning
Y, Winder T, LaBonte MJ, Wilson PM, Benhaim L, Paez D, et al:
Common cancer stem cell gene variants predict colon cancer
recurrence. Clin Cancer Res. 17:6934–6943. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Duffy M, Lamerz R, Haglund C, Nicolini A,
Kalousová M, Holubec L and Sturgeon C: Tumor markers in colorectal
cancer, gastric cancer and gastrointestinal stromal cancers:
European group on tumor markers 2014 guidelines update. Int J
Cancer. 134:2513–2522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Juo YY, Johnston FM, Zhang DY, Juo HH,
Wang H, Pappou EP, Yu T, Easwaran H, Baylin S, van Engeland M and
Ahuja N: Prognostic value of CpG island methylator phenotype among
colorectal cancer patients: A systematic review and meta-analysis.
Ann Oncol. 25:2314–2327. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sanders MA and Majumdar AP: Colon cancer
stem cells: Implications in carcinogenesis. Front Biosci (Landmark
Ed). 16:1651–1662. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vermeulen L, Todaro M, de Sousa Mello F,
Sprick MR, Kemper K, Alea M Perez, Richel DJ, Stassi G and Medema
JP: Single-cell cloning of colon cancer stem cells reveals a
multi-lineage differentiation capacity. Proc Natl Acad Sci USA.
105:13427–13432. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ahn JB, Chung WB, Maeda O, Shin SJ, Kim
HS, Chung HC, Kim NK and Issa JP: DNA methylation predicts
recurrence from resected stage III proximal colon cancer. Cancer.
117:1847–1854. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang IP, Tsai HL, Hou MF, Chen KC, Tsai
PC, Huang SW, Chou WW, Wang JY and Juo SH: MicroRNA-93 inhibits
tumor growth and early relapse of human colorectal cancer by
affecting genes involved in the cell cycle. Carcinogenesis.
33:1522–1530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liao X, Lochhead P, Nishihara R, Morikawa
T, Kuchiba A, Yamauchi M, Imamura Y, Qian ZR, Baba Y, Shima K, et
al: Aspirin use, tumor PIK3CA mutation, and colorectal-cancer
survival. N Engl J Med. 367:1596–1606. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
da W Huang, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.PubMed/NCBI
|
|
14
|
Richardson J, Viswanathan K and Lucas A:
Serpins, the vasculature, and viral therapeutics. Front Biosci.
11:1042–1056. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang X, Song YF, Lu HN, Wang DP, Zhang
XS, Huang SL, Sun BL and Huang ZG: Combined detection of plasma
GATA5 and SFRP2 methylation is a valid noninvasive biomarker for
colorectal cancer and adenomas. World J Gastroenterol.
21:2629–2637. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ning L, Haixing J, Zhenyu H, Shanyu Q and
Xin L: Expression of SFRP2, β-catenin and their roles in colorectal
carcinoma. J Dig Oncol (Electronic Version). 2008.
|
|
17
|
Shi Y, Liu Y, Wang J, Jie D, Yun T, Li W,
Yan L, Wang K and Feng J: Downregulated long noncoding RNA BANCR
promotes the proliferation of colorectal cancer cells via
downregualtion of p21 expression. PLoS One. 10:e01226792015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ge X, Chen Y, Liao X, Liu D, Li F, Ruan H
and Jia W: Overexpression of long noncoding RNA PCAT-1 is a novel
biomarker of poor prognosis in patients with colorectal cancer. Med
Oncol. 30:5882013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yao L, Lao W, Zhang Y, Tang X, Hu X, He C,
Hu X and Xu LX: Identification of EFEMP2 as a serum biomarker for
the early detection of colorectal cancer with lectin affinity
capture assisted secretome analysis of cultured fresh tissues. J
Proteome Res. 11:3281–3294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McPherson RA and Pincus MR: Henry's
Clinical Diagnosis and Management by Laboratory Methods. 22nd
edition. Elsevier Saunders; Philadelphia, PA: 2011, View Article : Google Scholar
|
|
22
|
Guo Q, Song Y, Zhang H, Wu X, Xia P and
Dang C: Detection of hypermethylated fibrillin-1 in the stool
samples of colorectal cancer patients. Med Oncol. 30:6952013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chew A, Salama P, Robbshaw A, Klopcic B,
Zeps N, Platell C and Lawrance IC: SPARC, FOXP3, CD8 and CD45
correlation with disease recurrence and long-term disease-free
survival in colorectal cancer. PLoS One. 6:e220472011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liang JF, Wang HK, Xiao H, Li N, Cheng CX,
Zhao YZ, Ma YB, Gao JZ, Bai RB and Zheng HX: Relationship and
prognostic significance of SPARC and VEGF protein expression in
colon cancer. J Exp Clin Cancer Res. 29:712010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lascorz J, Chen B, Hemminki K and Försti
A: Consensus pathways implicated in prognosis of colorectal cancer
identified through systematic enrichment analysis of gene
expression profiling studies. PLoS One. 6:e188672011. View Article : Google Scholar : PubMed/NCBI
|