Introduction
Ankylosing spondylitis (AS) is a chronic
inflammatory arthritis which primarily affects the sacroiliac
joints and spine. It is characterized by the formation of bony
spurs leading to ankyloses, and the associated functional
impairment ultimately leads to disability (1). AS is a frequently occurring condition
in young men ~26 years of age, with a prevalence of 0.15–0.8% in
the ordinary population (2).
Conventional medical treatment, including disease-modifying
anti-rheumatic drugs, non-steroidal anti-inflammatory drugs and
corticosteroids do not adequately treat or control the symptoms
associated with AS in the majority of patients (3). The etiology and pathogenesis of AS
remains to be fully elucidated, therefore, control of the excessive
bone formation remains the primary aim in the treatment of AS
(4).
The perennial vine Tripterygium wilfordii
Hook. f. is a traditional Chinese herb with verified
anti-inflammatory, immunosuppressive and anti-proliferative
properties (5,6). The primary active component is
triptolide, which is clinically used in the treatment of rheumatoid
arthritis, glomerulonephritis and various other autoimmune diseases
(7–9). It has previously been demonstrated
that triptolide additionally exhibits anti-tumor activities due to
the observed inhibition of cell growth and induction of apoptosis
(10–12). However, to the best of our
knowledge, there is no available information demonstrating if
triptolide exhibits a role in the regulation of osteoblast
biological activity.
In the present study, a mouse osteoblast cell line
was employed to investigate the effect of triptolide on the
proliferation, differentiation and apoptosis of osteoblasts. The
present study determined that treatment with triptolide may inhibit
proliferation, induce cell cycle arrest and apoptosis of the
MC3T3-E1 mouse osteoblast cells in a dose-dependent manner.
Additionally, the osteogenic differentiation of these osteoblast
cells was suppressed by triptolide. These results may be of
important clinical significance in the control of AS.
Materials and methods
Cell culture and osteogenic
induction
The MC3T3-E1 mouse osteoblast cell line was
purchased from Type Culture Collection Center of Chinese Academy of
Sciences (Shanghai, China). Cells were cultured in Minimum
Essential Medium (MEM)-α (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Hyclone; GE Healthcare, Logan, UT, USA) at 37°C in an environment
containing 5% CO2. For osteogenic induction, the culture
medium was altered to MEM-α containing 5 mM β-glycerophosphate
(BioSharp, Hefei, China) and 50 µg/ml vitamin C (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany).
Bromodeoxyuridine (BrdU) assay
Cell viability was detected by BrdU ELISA kit (cat.
no. CEL-BRDU, Mai Biological Technology Co., Ltd., Changsha, China)
according to the manufacturer's protocol. Briefly, cells were
seeded in 96-well plates with MEM-α for 24 h, then supplemented
with 0, 2.8, 7 or 14 nM triptolide (Sigma-Aldrich; Merck KGaA).
Following incubation for 24, 48, or 72 h, 10 µl BrdU labeling
buffer was added, and incubated for a further 2 h at 37°C. Next,
the cells were fixed and denaturalized with 200 µl FixDenat
solution and incubated with 100 µl peroxidase conjugated
anti-BrdU-monoclonal antibody at room temperature for 90 min. Then,
100 µl substrate solution was added and the color was developed for
20 min. Finally, the reaction was stopped with the addition of 25
µl stop solution and absorbance was subsequently measured at a
wavelength of 450 nm.
Flow cytometry
Cells were seeded in 6-well plates at a density of
1×105 cells/well, and allowed to grow to 90% confluence.
Then, the cells were treated with 0, 2.8, 7 or 14 nM triptolide for
72 h. For cell cycle analysis, the cells were fixed with 70%
alcohol at 4°C for 2 h, then incubated with 25 µl propidium iodide
(PI) staining solution (Beyotime Institute of Biotechnology,
Haimen, China) and 10 µl RNase A for 30 min at 37°C in the dark.
Following this, the cell cycle progression was quantified using a
flow cytometer (BD Accuri C6; BD Biosciences, Franklin Lakes, NJ,
USA). Cell apoptosis was detected using the Annexin-V fluorescein
isothiocyanate (FITC) and PI double staining kit (Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China) as per the manufacturer's
protocol. The cells were stained with 5 µl Annexin V-FITC and 5 µl
PI for 15 min in dark, then subjected to BD Accuri C6 flow
cytometer. Cell cycle and apoptosis status were analyzed by Accuri
CFlow Plus version 1.0.1727 (BD Biosciences).
Hoechst staining
Nuclear apoptosis was detected using a Hoechst
staining kit (Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. Following treatment with different
concentrations of triptolide, cells were fixed with 500 µl fixative
at 4°C overnight. The cells were stained with 2 µg/ml Hoechst 33258
staining solution at 37°C for 30 min. The stained cells were
observed under a fluorescent microscope (BX53; Olympus Corporation,
Tokyo, Japan).
Western blotting
Cells were lysed using a radioimmunoprecipitation
buffer (Beyotime Institute of Biotechnology) supplemented with 10%
phenylmethane sulfonyl fluoride (Beyotime Institute of
Biotechnology). The concentration of extracted proteins was
determined by a conventional bicinchoninic acid method (Beyotime
Institute of Biotechnology). An equal amount of 40 µg protein was
electrophoresed on 10 or 13% SDS-PAGE. The proteins were
electrotransferred to a polyvinylidene fluoride membrane (EMD
Millipore, Billerica, MA, USA). Following blocking with 5% (M/V)
skimmed milk, the membrane was immunoblotted with primary
antibodies against proliferating cell nuclear antigen (PCNA; 1:500;
cat. no. bs-2006R; BIOSS, Beijing, China); cyclin D1, (1:400; cat.
no. BM0771); cyclin E, (1:400; cat. no BA0774); B cell lymphoma
(Bcl)-2, (1:400; cat. no. BA0412); Bcl-2 associated X protein
(Bax), (cat. no. BA0315; 1:400) all purchased from Boster
Biological Technology, Pleasanton, CA, USA) and cleaved-caspase 3,
(cat. no. ab2302; 1:1,000; Abcam, Cambridge, MA, USA) at 4°C
overnight. Following this, the membrane was incubated with
horseradish peroxidase-conjugated goat anti-rabbit (1:5,000; cat.
no. WLA023; Wanleibio, Shenyang, China) or goat anti-mice antibody
(1:5,000, cat. no. WLA024; Wanleibio) at 37°C for 45 min. The
specific protein bands were detected by an enhanced
chemiluminescence reagent (7Sea biotech Co., Ltd., Shanghai, China)
and quantified by Gel-Pro Analyzer Version 3.0 (Media Cybernetics,
Inc., Rockville, MD, USA). β-actin served as an internal
control.
Picrosirius staining
Cells were cultured in osteogenic induction medium
with various concentrations of triptolide at 37°C in an environment
containing 5% CO2 for 7 days. Following fixing with 1 ml
Bouin's solution (Sigma-Aldrich; Merck KGaA) at room temperature
for 1 h, the cells were stained with 1% Sirius red in saturated
picric acid for 1 h. The cells were then washed and observed under
an optical microscope (DP73; Olympus Corporation).
Measurement of alkaline phosphatase
activity and calcium deposition
MC3T3-E1 cells were cultured in osteogenic induction
medium supplemented with various concentrations of triptolide for
12 days. The cells were then collected and lysed by repeated
freezing and thawing with liquid nitrogen. Proteins were quantified
using a conventional bicinchoninic acid method and diluted to 1
mg/ml. Alkaline phosphatase activity and calcium deposition were
measured using the commercial kits following the manufacturer's
instructions. The AKP/ALP detection kit was purchased from Nanjing
Jiancheng Bioengineering Institute (cat. no. A059-1, Nanjing,
China) and the calcium colorimetric assay kit was purchased from
Sigma-Aldrich (cat. no. MAK022, Merck KGaA, Darmstadt, Germany).
Absorbance value was detected at 520 nm for ALP and at 575 nm for
calcium ions were measured using a microplate spectrophotometer and
its assorted software Gen5 version 2.0 (ELx-800, Biotek
Instruments, Winooski, VT, USA).
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical analysis was performed using SPSS software,
version 16.0 (SPSS, Inc., Chicago, IL, USA) and data were evaluated
by analysis of variance, followed by the Bonferroni test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Triptolide inhibits MC3T3-E1 cell
proliferation
MC3T3-E1 cells were treated with increasing
concentrations of triptolide (0, 2.8, 7 or 14 nM) for 24, 48 or 72
h and the effect of triptolide on the cells was tested using the
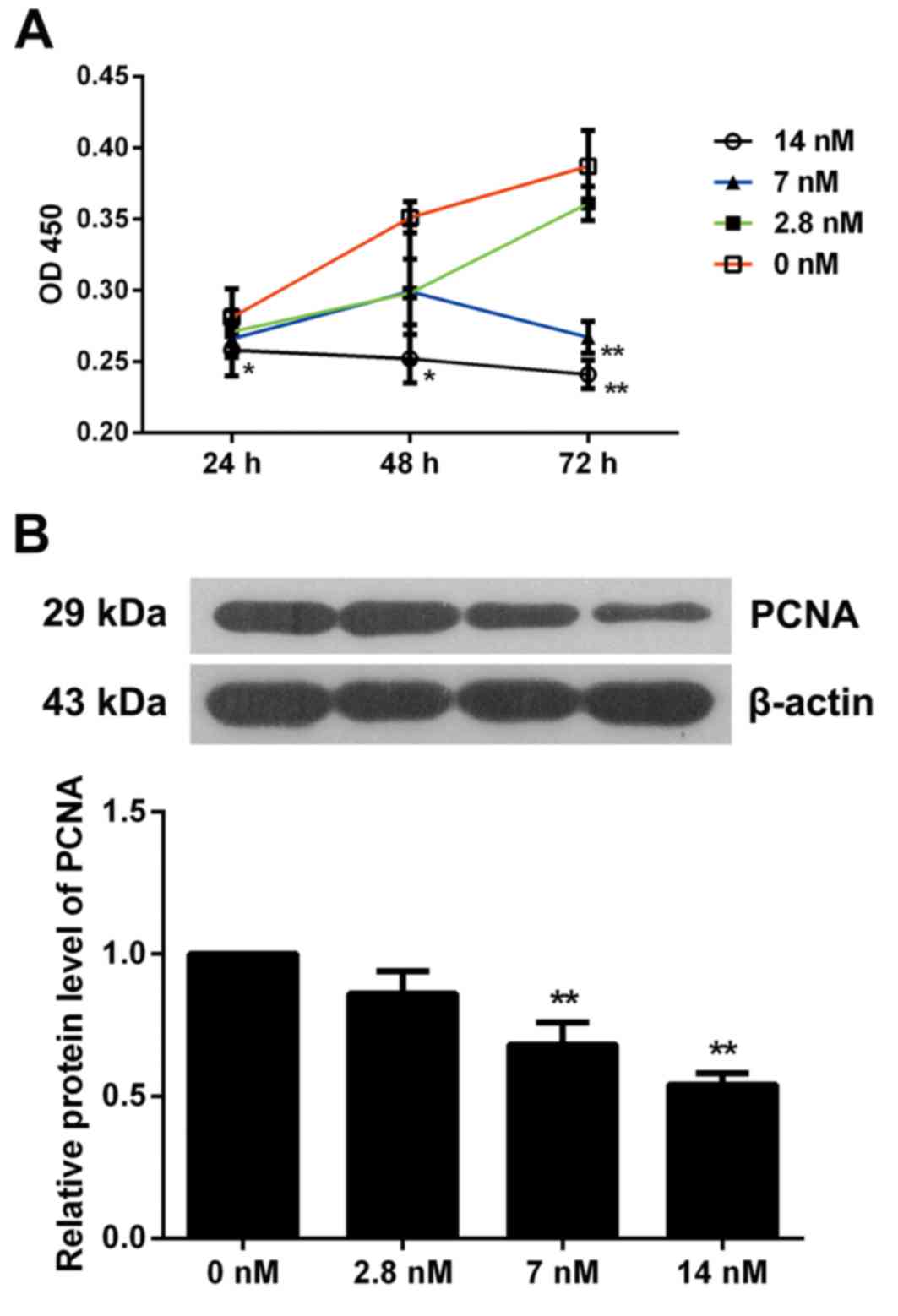
BrdU-based viability assay. As presented in Fig. 1A, 2.8 nM triptolide did not affect
the cell viability, 7 nM triptolide significantly inhibited cell
viability at 72 h (P<0.01), and cells exposed to 14 nM
triptolide exhibited a marked decrease in proliferation at 24, 48
and 72 h when compared with non-treated cells (P<0.05).
Therefore, triptolide inhibited MC3T3-E1 cell viability in a dose
and time-dependent manner. Furthermore, the expression of PCNA was
detected by western blotting and it was observed that MC3T3-E1
cells exposed to 7 or 14 nM triptolide for 72 h exhibited
significantly inhibited PCNA levels (P<0.01). These results
further indicated the suppressive effect of triptolide on
osteoblast proliferation (Fig.
1B).
Triptolide induces cell cycle arrest
of MC3T3-E1 cells
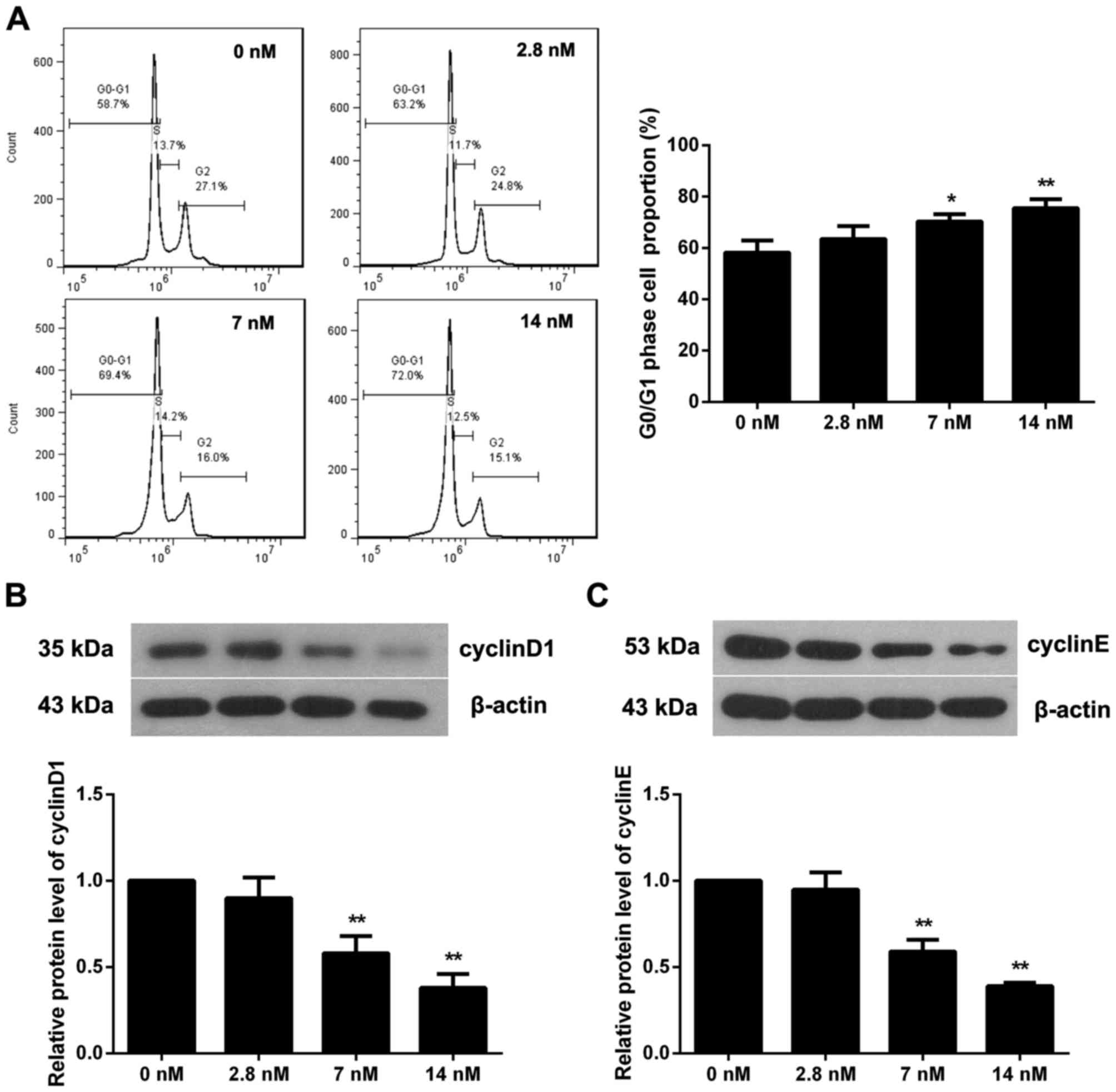
To investigate if triptolide affects the MC3T3-E1
cell cycle, flow cytometry was performed when cells were incubated
with triptolide for 72 h. As presented in Fig. 2A, the proportion of cells in the
G0/G1 phase in the 7 or 14 nM triptolide treatment groups was
significantly increased (P<0.05), indicating a cell cycle arrest
induced by triptolide. The expression of two pivotal proteins in
the regulation of the cell cycle, cyclin D1 and cyclin E, was
additionally determined (Fig. 2B and
C). The results demonstrated that 7 or 14 nM triptolide
significantly reduced the level of cyclin D1 and cyclin E
(P<0.01). Therefore, tripolide may induce cell cycle arrest at
the G0/G1 phase via inhibition of cyclin D1 and cyclin E
expression.
Triptolide induces apoptosis of
MC3T3-E1 cells
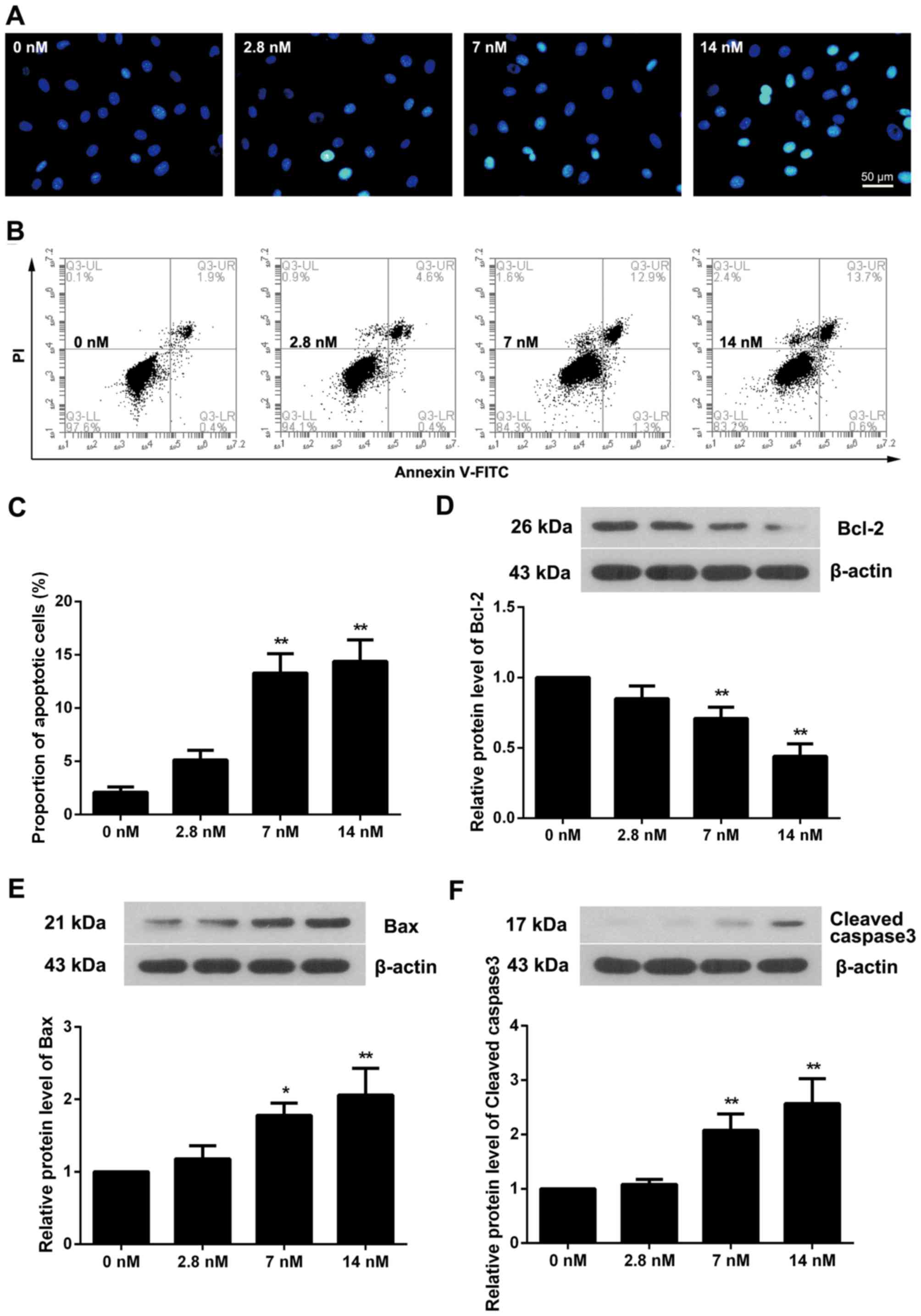
Hoechst staining was used to evaluate the apoptotic
status of triptolide treated cells. As presented in Fig. 3A, blue apoptotic nuclei were
observed in 2.8 nM triptolide treated cells, and this was elevated
with the increased triptolide concentration. Flow cytometry
additionally demonstrated that the proportion of apoptotic cells
was increased in triptolide treated cells, particularly in the 7
and 14 nM triptolide treated groups (Fig. 3B and C; P<0.01). Measurement of
apoptosis-associated proteins demonstrated that the level of Bcl-2
was decreased as the triptolide concentration increased (Fig. 3D). Conversely, the protein levels
of Bax and Cleaved caspase3 were upregulated (Fig. 3E and F).
Triptolide suppresses differentiation
and mineralization of MC3T3-E1 cells
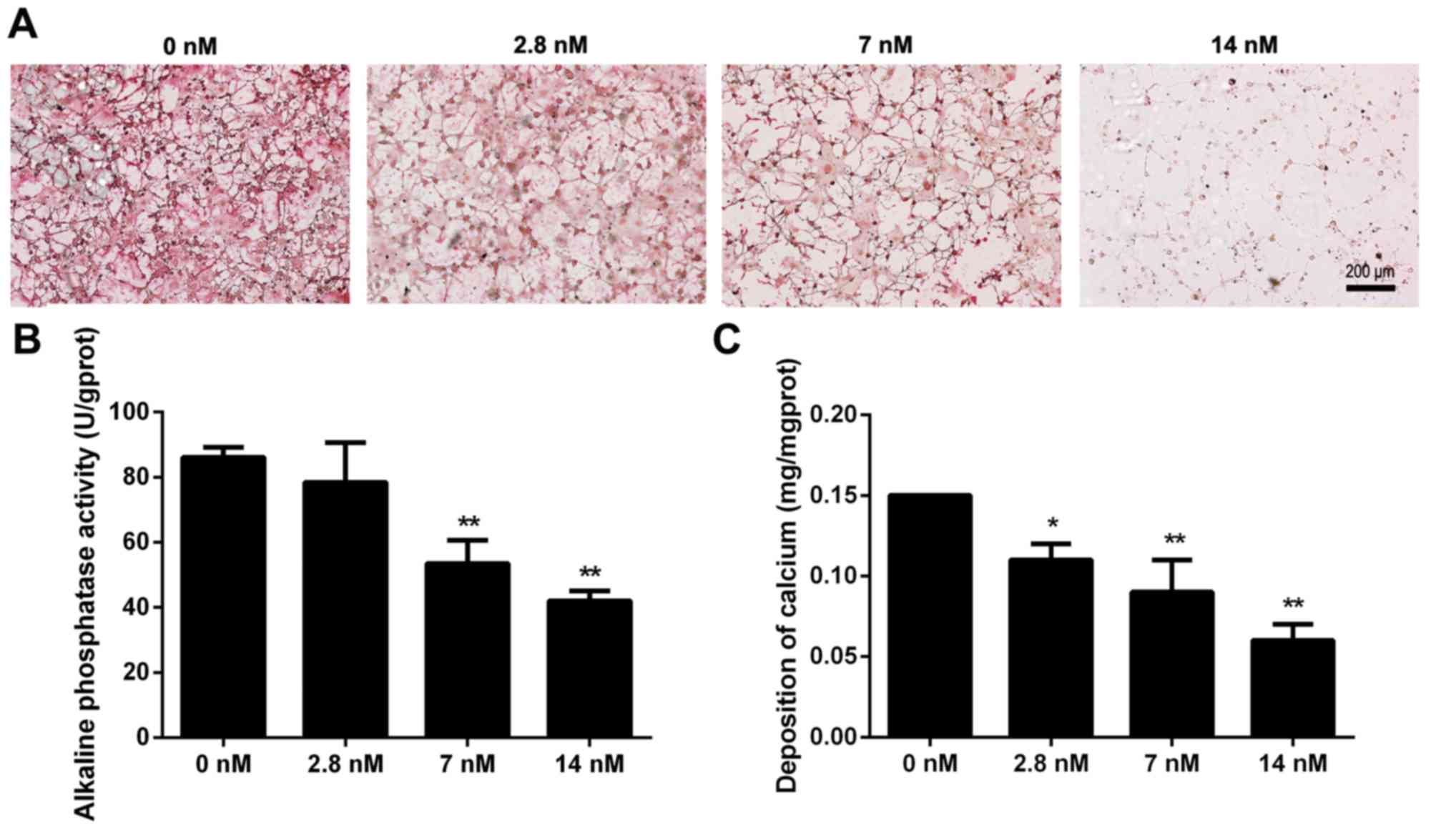
To examine the effect of triptolide on osteoblast
differentiation, MC3T3-E1 cells were cultured in osteogenic
induction medium with different concentrations of triptolide. The
collagen formation was observed by picrosirius staining at day 7
and the results demonstrated that the collagen fibers were
decreased with the increased triptolide concentrations (Fig. 4A). The activity of alkaline
phosphatase is a marker of osteoblastic differentiation, and the
level of activity was subsequently determined at day 12. It was
demonstrated that 7 and 14 nM triptolide significantly reduced the
alkaline phosphatase activity (Fig.
4B; P<0.01). Furthermore, calcium deposition was measured to
investigate the mineralized matrix formation in triptolide treated
cells, as demonstrated in Fig. 4C.
The cells exposed to 2.8, 7 or 14 nM triptolide exhibited a
significantly decreased level of calcium deposition, when compared
with the control group (P<0.05). Triptolide therefore suppressed
differentiation and mineralization of MC3T3-E1 cells.
Discussion
The anti-inflammatory and anti-neoplastic properties
of triptolide have been extensively reported (13–16),
however, the effect on the regulation of biological activities in
osteoblasts remains to be elucidated. In the present study, mouse
osteoblast MC3T3-E1 cells were treated with increasing
concentrations of triptolide at different time intervals, and it
was demonstrated that triptolide inhibited cell proliferation in a
dose and time-dependent manner. Triptolide additionally induced
cell cycle arrest and apoptosis of MC3T3-E1 cells. Treatment with
triptolide resulted in delayed collagen formation, decreased
alkaline phosphatase activity and reduced calcium deposition. The
findings therefore demonstrated the suppressive effect of
triptolide on osteoblast proliferation and bone formation.
The growth inhibition effect of triptolide has been
demonstrated in a variety of solid tumor types, in vivo and
in vitro (17,18). It was revealed that triptolide
inhibited cell proliferation in a dose and time-dependent manner in
mouse osteoblast MC3T3-E1 cells. PCNA is a highly conservative
nuclear protein of DNA polymerase-∆, and is considered to be a
useful marker to assess cell proliferation and progression
(19,20). In the present study, 7 and 14 nM
triptolide significantly reduced the expression of PCNA in MC3T3-E1
cells and further proved the inhibitory effect of triptolide on
cell proliferation. These results are in accordance with previous
studies on tumor (21) and
T-lymphocyte cells (22).
The cell cycle is regulated by a range of components
including cyclin-proteins, cyclin-dependent kinases (CDK) and
various other factors. The loss of function of these regulators may
result in cell cycle disorder and uncontrolled cell proliferation.
During the cell cycle, cyclin D1 combines with CDK4 or CDK6 and
cyclin E combines with CDK2. The cyclin and CDK complexes
phosphorylate Rb, release E2F and thereby activate the transition
from the G1 to the S phase (23–25).
It has previously been demonstrated that triptolide decreases
proliferation, induces G0/G1 cycle arrest and suppresses the
expression of cyclin-proteins in multiple myeloma cells (26), malignant glioma cells (27) and colon-rectal cancer cells
(28,29). Consistent with previous studies,
the present study demonstrated that triptolide triggered G0/G1
arrest and inhibited the expression of cyclin D1 and cyclin E in
osteoblasts.
Cell cycle arrest and apoptosis are highly
associated events and disruption of the cell cycle induces
apoptosis (30). Bcl-2 family
proteins, including anti-apoptotic Bcl-2, Bcl-XL and
pro-apoptotic Bax, are key regulators in mitochondria-dependent
apoptosis (31), and caspase 3 is
the final executor in this pathway (32). Li et al (33) reported in human renal cell
carcinoma cells that triptolide increases Bax expression, reduces
Bcl-2 level, increased caspase 3 activity, and induces cell
apoptosis in a dose-dependent manner. The results of the present
study are in accordance with the reports on human renal cell
carcinoma cells, and triptolide induced the apoptosis of MC3T3-E1
cells via suppression of Bcl-2 and promotion of Bax.
Osteogenesis is a well-coordinated physiological
process including extracellular matrix production, secretion, and
matrix mineralization (34). To
verify the effect of triptolide on osteogenesis, MC3T3-E1 cells
were incubated in osteogenesis induction medium with different
concentrations of triptolide. The results demonstrated that
triptolide blocked collagen formation, suppressed alkaline
phosphatase activity and reduced calcium deposition, suggesting an
inhibitory effect of triptolide on osteoblast differentiation.
However, Huang et al (35)
and Park (36) demonstrated that
triptolide inhibits osteoclastogenesis and bone resorption,
respectively. Therefore, triptolide may exhibit a bidirectional
role in bone regulation, and further studies are required in order
to fully elucidate the underlying mechanisms.
In conclusion, the data from the present study
demonstrated that triptolide inhibited proliferation and induced
cell cycle arrest and apoptosis of osteoblasts. In addition,
triptolide was identified as a potent suppressor of osteoblast
differentiation and mineralization. These results verify the
inhibitory effect of triptolide in bone formation and suggest a
potential treatment strategy for AS in the future.
Acknowledgements
The present study was supported by the High-Level
Talent Project of Jiangsu Province Hospital of Traditional Chinese
Medicine (grant no. y2014rc08), the Natural Science Foundation of
Jiangsu Province (grant no. BK20151602) and the National Natural
Science Foundation of China (grant no. 81403174).
References
|
1
|
Braun J and Sieper J: The sacroiliac joint
in the spondyloarthropathies. Curr Opin Rheumatol. 8:275–287. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reveille JD and Brown MA: Epidemiology of
ankylosing spondylitis: IGAS 2009. J Rheumatol. 37:2624–2625. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Toussirot E and Wendling D: Current
guidelines for the drug treatment of ankylosing spondylitis. Drugs.
56:225–240. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schett G and Rudwaleit M: Can we stop
progression of ankylosing spondylitis? Best Pract Res Clin
Rheumatol. 24:363–371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang J, Wang A, Zeng H, Liu L, Jiang W,
Zhu Y and Xu Y: Effect of triptolide on T-cell receptor beta
variable gene mRNA expression in rats with collagen-induced
arthritis. Anat Rec (Hoboken). 295:922–927. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xue M, Jiang ZZ, Liu JP, Zhang LY, Wang T,
Wang H, Liu L and Zhou ZX: Comparative study on the
anti-inflammatory and immune suppressive effect of Wilforlide A.
Fitoterapia. 81:1109–1112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shui G, Wan Y, Jiang C, Zhang H, Chen P,
Wang C and Yao J: Progress in Tripterygium wilfordiiand its
bioactive components in the field of pharmacodynamics and
pharmacology. Zhongguo Zhong Yao Za Zhi. 35:515–520.
2010.PubMed/NCBI
|
|
8
|
Brinker AM, Ma J, Lipsky PE and Raskin I:
Medicinal chemistry and pharmacology of genus Tripterygium
(Celastraceae). Phytochemistry. 68:732–766. 2007.(In Chinese).
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu C, Zhang Y, Kong X, Zhu L, Pang J, Xu
Y, Chen W, Zhan H, Lu A and Lin N: Triptolide prevents bone
destruction in the collagen-induced arthritis model of rheumatoid
arthritis by targeting RANKL/RANK/OPG signal pathway. Evid Based
Complement Alternat Med. 2013:6260382013.PubMed/NCBI
|
|
10
|
Jiang QW, Cheng KJ, Mei XL, Qiu JG, Zhang
WJ, Xue YQ, Qin WM, Yang Y, Zheng DW, Chen Y, et al: Synergistic
anticancer effects of triptolide and celastrol, two main compounds
from thunder god vine. Oncotarget. 6:32790–32804. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jao HY, Yu FS, Yu CS, Chang SJ, Liu KC,
Liao CL, Ji BC, Bau DT and Chung JG: Suppression of the migration
and invasion is mediated by triptolide in B16F10 mouse melanoma
cells through the NF-kappaB-dependent pathway. Environ Toxicol. Sep
29–2015.(Epub ahead of print). PubMed/NCBI
|
|
12
|
Jiang N, Dong XP, Zhang SL, You QY, Jiang
XT and Zhao XG: Triptolide reverses the Taxol resistance of lung
adenocarcinoma by inhibiting the NF-κB signaling pathway and the
expression of NF-κB-regulated drug-resistant genes. Mol Med Rep.
13:153–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou GX, Ding XL, Huang JF, Zhang H, Wu
SB, Cheng JP and Wei Q: Apoptosis of human pancreatic cancer cells
induced by Triptolide. World J Gastroenterol. 14:1504–1509. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pan J: RNA polymerase-an important
molecular target of triptolide in cancer cells. Cancer Lett.
292:149–152. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qiu D and Kao PN: Immunosuppressive and
anti-inflammatory mechanisms of triptolide, the principal active
diterpenoid from the Chinese medicinal herb Tripterygium wilfordii
Hook. f. Drugs R D. 4:1–18. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Matta R, Wang X, Ge H, Ray W, Nelin LD and
Liu Y: Triptolide induces anti-inflammatory cellular responses. Am
J Transl Res. 1:267–282. 2009.PubMed/NCBI
|
|
17
|
Chen L, Liu Q, Huang Z, Wu F, Li Z, Chen X
and Lin T: Tripchlorolide induces cell death in lung cancer cells
by autophagy. Int J Oncol. 40:1066–1070. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Westfall SD, Nilsson EE and Skinner MK:
Role of triptolide as an adjunct chemotherapy for ovarian cancer.
Chemotherapy. 54:67–76. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bodduluru LN, Kasala ER, Madhana RM, Barua
CC, Hussain MI, Haloi P and Borah P: Naringenin ameliorates
inflammation and cell proliferation in benzo(a)pyrene induced
pulmonary carcinogenesis by modulating CYP1A1, NFκB and PCNA
expression. Int Immunopharmacol. 30:102–110. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ben-Izhak O, Bar-Chana M, Sussman L,
Dobiner V, Sandbank J, Cagnano M, Cohen H and Sabo E: Ki67 antigen
and PCNA proliferation markers predict survival in anorectal
malignant melanoma. Histopathology. 41:519–525. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang S, Chen J, Guo Z, Xu XM, Wang L, Pei
XF, Yang J, Underhill CB and Zhang L: Triptolide inhibits the
growth and metastasis of solid tumors. Mol Cancer Ther. 2:65–72.
2003.PubMed/NCBI
|
|
22
|
Yang SX, Xie SS, Gao HL, Ma DL and Long
ZZ: Triptolide suppresses T-lymphocyte proliferation by inhibiting
interleukin-2 receptor expression, but spares interleukin-2
production and mRNA expression. Int J Immunopharmacol. 16:895–904.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sherr CJ: G1 phase progression: Cycling on
cue. Cell. 79:551–555. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sherr CJ: Growth factor-regulated G1
cyclins. Stem Cells. 12 Suppl 1:S47–S57. 1994.
|
|
25
|
Sherr CJ, Kato J, Quelle DE, Matsuoka M
and Roussel MF: D-type cyclins and their cyclin-dependent kinases:
G1 phase integrators of the mitogenic response. Cold Spring Harb
Symp Quant Biol. 59:11–19. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao F, Chen Y, Li R, Liu Y, Wen L and
Zhang C: Triptolide alters histone H3K9 and H3K27 methylation state
and induces G0/G1 arrest and caspase-dependent apoptosis in
multiple myeloma in vitro. Toxicology. 267:70–79. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang H, Zhu W, Su X, Wu S, Lin Y, Li J,
Wang Y, Chen J, Zhou Y, Qiu P, et al: Triptolide inhibits
proliferation and invasion of malignant glioma cells. J Neurooncol.
109:53–62. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu J, Shen M, Yue Z, Yang Z, Wang M, Li
C, Xin C, Wang Y, Mei Q and Wang Z: Triptolide inhibits
colon-rectal cancer cells proliferation by induction of G1 phase
arrest through upregulation of p21. Phytomedicine. 19:756–762.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oliveira AR, Beyer G, Chugh R, Skube SJ,
Majumder K, Banerjee S, Sangwan V, Li L, Dawra RK, Subramanian S,
et al: Triptolide abrogates growth of colon cancer and induces cell
cycle arrest by inhibiting transcriptional activation of E2F. Lab
Invest. 95:648–659. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pucci B, Kasten M and Giordano A: Cell
cycle and apoptosis. Neoplasia. 2:291–299. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martinou JC and Youle RJ: Mitochondria in
apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev
cell. 21:92–101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Donovan M and Cotter TG: Control of
mitochondrial integrity by Bcl-2 family members and
caspase-independent cell death. Biochim Biophys Acta. 1644:133–147.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li J, Zhu W, Leng T, Shu M, Huang Y, Xu D,
Qiu P, Su X and Yan G: Triptolide-induced cell cycle arrest and
apoptosis in human renal cell carcinoma cells. Oncol Rep.
25:979–987. 2011.PubMed/NCBI
|
|
34
|
Owen TA, Aronow M, Shalhoub V, Barone LM,
Wilming L, Tassinari MS, Kennedy MB, Pockwinse S, Lian JB and Stein
GS: Progressive development of the rat osteoblast phenotype in
vitro: Reciprocal relationships in expression of genes associated
with osteoblast proliferation and differentiation during formation
of the bone extracellular matrix. J Cell Physiol. 143:420–430.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang J, Zhou L, Wu H, Pavlos N, Chim SM,
Liu Q, Zhao J, Xue W, Tan RX, Ye J, et al: Triptolide inhibits
osteoclast formation, bone resorption, RANKL-mediated NF-κB
activation and titanium particle-induced osteolysis in a mouse
model. Mol Cell Endocrinol. 399:346–353. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Park B: Triptolide, a diterpene, inhibits
osteoclastogenesis, induced by RANKL signaling and human cancer
cells. Biochimie. 105:129–136. 2014. View Article : Google Scholar : PubMed/NCBI
|