Introduction
Due to the rapid development of stem cell
transplantation in preclinical research and clinical trials over
the last decade, stem cell replacement therapies have become modern
therapeutic approaches (1).
Similar to bone-marrow stromal cells, adipose-derived stem cells
(ADSCs) have the ability to differentiate into various types of
cell lineage, including adipogenic, osteogenic, myogenic and
chondrogenic cells (2,3). In addition, the yield of mesenchymal
stem cells (MSCs) from adipose tissue is 100–500 fold that from
bone marrow (4). The unique
biology of autologous ADSCs, including being easily expanded,
immune-privileged and capable of long-term transgene expression
following multiple stages of differentiation, demonstrates their
potential as a gene delivery vehicle (5–7).
The endothelial differentiation of ADSCs serves an
essential role in vascular development, function and disease
(8). Numerous peptide growth
factors promote angiogenesis by promoting differentiation, and
enhancing endothelial cell proliferation, migration and capillary
network stability (9).
The knockdown of microRNA-126 (miR-126) in zebrafish
in vivo resulted in the loss of vascular integrity and
hemorrhage during embryonic development, and targeted deletion of
miR-126 in mice caused delayed angiogenic sprouting, widespread
hemorrhaging and partial embryonic lethality (10–12).
These vascular abnormalities may be attributed to diminished
angiogenic growth factor signaling, resulting in reduced
endothelial cell differentiation, growth, sprouting and adhesion
(13–16). This evidence indicates that miR-126
may be involved in multiple aspects of fundamental biological
processes, including angiogenesis. However, the role of miR-126 in
mediating the differentiation of ADSCs to endothelial cells has not
been demonstrated.
The present study focused on the association between
miR-126 and the endothelial differentiation of ADSCs.
Materials and methods
Cell growth curve and endothelial
differentiation of ADSCs
ADSCs were purchased from the American Type Culture
Collection (Manassas, VA, USA) and cultured in α-Minimum Essential
Medium (α-MEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), supplemented with 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.) at 37°C in 5% CO2. Equal
numbers (2×104) of passage (p)1, 3, 6 and 8 cells were
plated and cultured. Cells images were obtained at ×100
magnification using a phase-contrast inverted light microscope.
Cells were harvested each day for 8 days. The total number of cells
was counted in each plate using a hemocytometer. The cell growth
curve was drawn by plotting the mean cell number of each plate
against the culture time. For endothelial differentiation, ADSCs
were cultured in Endothelial Cell Growth Medium-2 (EGM-2; Lonza,
Basel, Switzerland). Human umbilical vein endothelial cells
(HUVECs; CRL-1730; American Type Culture Collection) were cultured
in F-12K medium (30–2004; American Type Culture Collection) with
10% fetal bovine serum (30–2020; American Type Culture Collection),
0.05 mg/ml ECGS (no. 354006, BD Biosciences, Franklin Lakes, NJ,
USA) and 0.1 mg/ml heparin (#H3393; Sigma-Aldrich, Merck KGaA,
Darmstadt, Germany).
Transfection
The hsa-miR-126-3p inhibitor (CGC AUU AUU ACU CAC
GGU ACGA), hsa-miR-126-3p mimic (5–3′, UCG UAC CGU GAG UAA UAA
UGCG; 3–5′, AGC AUG GCA CUC AUU AUU ACGC;), and their negative
controls (NCs) (mimic NC, 5–3′, UCA CAA CCU CCU AGA AAG AGU AGA;
3–5′, AGU GUU GGA GGA UCU UUC UCA UCU; inhibitor NC, UCU ACU CUU
UCU AGG AGG UUG UGA; all Biomics Biotechnologies, Nantong, China)
were diluted to a final concentration of 50 nM with RNase-free
H2O. ADSCs were transfected respectively with these
oligonucleotides using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.), following the manufacturer's protocol. At
each time point (days 1–8), cells were harvested for further
analysis. The efficiency of transfection was determined using
aCy3-short interfering RNA (Cy3-siRNA) transfection control
(Biomics Biotechnologies). The inhibition and overexpression
efficiencies were determined by comparing with the NCs.
Immunofluorescence staining
Cells were uninduced in α-MEM or induced in EGM-2
for 7–14 days, fixed with 4% paraformaldehyde (PFA), blocked with
10% normal goat serum (50062Z; Thermo Fisher Scientific, Inc.) for
10 min at room temperature and incubated with one of three primary
antibodies: Rabbit anti-human cluster of differentiation (CD)31
(1:50; 11265-AP), von Willebrand factor (vWF) (1:50; 11778-1-AP) or
endothelial nitric oxide synthase (eNOS) (1:50; 20116-1-AP) (all
from Proteintech Group, Inc., Chicago, IL, USA) at 4°C overnight.
Following incubation with fluorescein isothiocyanate
(FITC)-conjugated goat anti-rabbit IgG (CW0114S, 1:100; CWBiotech,
Beijing, China) for 3 h at room temperature, the cells were
counterstained by incubation with DAPI (Sigma-Aldrich, Merck KGaA).
Images were obtained at magnification ×200 using a phase contrast
fluorescence microscope (Olympus Corporation, Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA and miRNA were isolated using a miRNeasy
Mini kit (Qiagen, Inc., Valencia, CA, USA). Synthesis of
complementary DNA from mRNA and miRNA was performed using a
PrimeScript RT reagent kit (Takara Bio, Inc., Otsu, Japan) or a
miRcute miRNA First-Strand cDNA Synthesis kit (Tiangen Biotech Co.,
Ltd., Beijing, China), respectively. RT-qPCR analysis of mRNA and
miRNA was performed using SYBR Premix Ex Taq™ (Takara
Bio, Inc.) or a miRcute miRNA qPCR Detection kit (SYBR Green)
(Tiangen Biotech Co., Ltd.), respectively. Amplification data was
recorded using an ABI 7500 Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The PCR analysis of
miR-126 consisted of 38 cycles of 95°C for 15 sec and 60°C for 1
min, following an initial denaturation step of 95°C for 10 min. The
expression level of miR-126 was normalized to U6 small nuclear RNA,
and the expression levels of CD31, vWF and eNOS mRNA were
normalized to GAPDH. The primers used are presented in Table I. The results were subjected to
melting curve analysis, and the data were analyzed using the
2−ΔΔCq method (17).
 | Table I.Primers used in the present study. |
Table I.
Primers used in the present study.
| Name | Sequence (5′-3′) |
|---|
| CD31 | F,
TGTATTTCAAGACCTCTGTGCACTT |
|
| R,
TTAGCCTGAGGAATTGCTGTGTT |
| vWF | F,
TAAGTCTGAAGTAGAGGTGG |
|
| R,
AGAGCAGCAGGAGCACTGGT |
| eNOS | F,
CAGTGTCCAACATGCTGCTGGAAATTG |
|
| R,
TAAAGGAGGTCTTCTTCCTGGTGATGCC |
| GAPDH | F,
ACCACAGTCCATGCCATCAC |
|
| R,
TCCACCACCCTGTTGCTGTA |
| miR-126-3p |
RT,CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCGCATTAT |
|
| F,
ACACTCCAGCTGGGTCGTACCGTGAGTA |
|
| R,
CTCAACTGGTGTCGTGGAGT |
| U6 | RT,
CAAAATATGGAACGCTTC |
|
| F,
GTGCTCGCTTCGGCAGC |
|
| R,
CAAAATATGGAACGCTTC |
Western blot analysis
Confluent cells were removed by scraping, lysed in
mammalian protein extraction reagent (Thermo Fisher Scientific,
Inc.) supplemented with the protease inhibitor phenylmethylsulfonyl
fluoride, and quantified using a bicinchoninic acid assay. Proteins
(20 µg/lane) were resolved on a 10% SDS-PAGE gel and transferred
onto a polyvinylidene fluoride membrane. Following blocking in 5%
non-fat dried milk and TBS with 0.1% Tween-20 (TBST) for 2 h, the
blots were incubated with a rabbit anti-human CD31 (1:500;
11265-AP), vWF (1:500; 11778-1-AP) or eNOS (1:500; 20116-1-AP) (all
from ABclonal Biotech Co., Ltd., Cambridge, MA, USA) overnight at
4°C. GAPDH (1:3,000; sc-47724; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) was used as an internal control. Subsequent to
washing with TBST, the blots were incubated with HRP-conjugated
goat anti-rabbit IgG (CW0103S, 1:2,000; CWBiotech) at room
temperature for 1 h and visualized by enhanced chemiluminescence
(GE Health care Biosciences, Pittsburgh, PA, USA). Image band area
and density were estimated using Quantity One 4.6.2 (Bio-Rad
Laboratories Inc., Hercules, CA, USA).
Statistical analysis
Each experiment was repeated three times
independently. All data were expressed as the mean ± standard
deviation. Statistical analysis was performed using a one-way
analysis of variance test and SPSS software (version 19.0; IBM
SPSS, Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Morphological features of cultured
ADSCs
The ADSCs attached to the culture dish surface
exhibited a typical fibroblast-like morphology. The cells were
maintained in culture with no sign of senescence or differentiation
following repeated subculturing to p6 (Fig. 1A-C). In order to investigate
whether passages affect cell growth, the number of cells was
recorded every day for 8 days and compared with the cell growth
curve of p1, 3, 6 and 8. Following an initial lag or stationary
period (day 1), the cells expanded rapidly in a logarithmic manner
until a plateau was reached (day 6). However, the ADSCs exhibited
markedly more rapid growth at p3 and 6 (Fig. 1D).
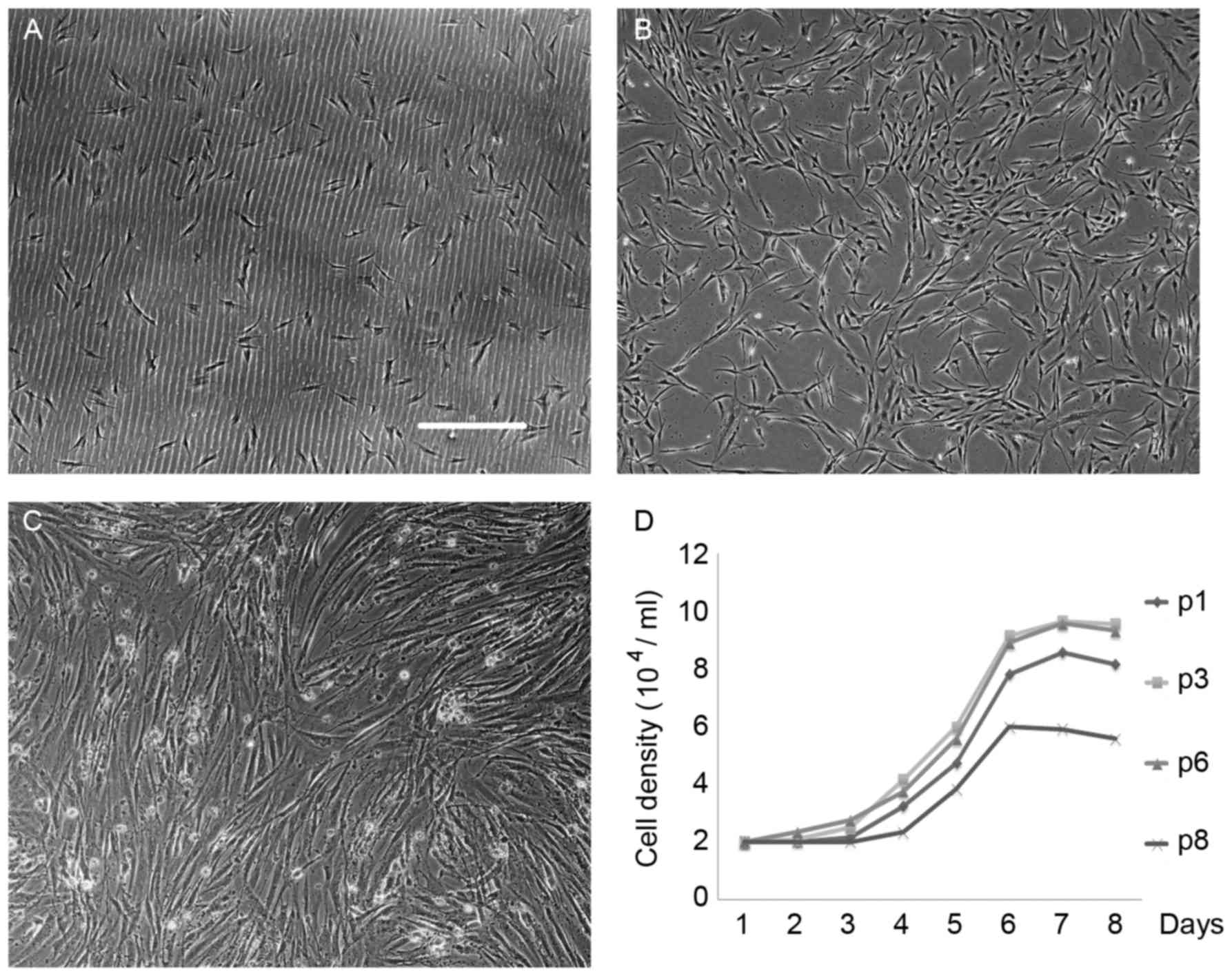 | Figure 1.Biological characteristics of ADSCs.
(A) The majority of cells were adherent and appeared round in
morphology 24 h following passage. Magnification, ×100; scale bar,
200 µm. (B) After 3 days, the adherent cells in complete medium
became active, proliferated quickly, formed processes, and expanded
to generate small and large colonies. (C) 6 days later, the cells
reached 80% confluence. Cells that attached to the culture dish
surface exhibited a typical fibroblast-like morphology. (D) The
growth of ADSCs was slow and limited to the first 24 h.
Representative growth curves of ADSCs in their logarithmic phase
after day 3 are presented. A marked inhibition of cell growth was
observed in both p1and 8 compared with p3 and 6. Growth curves of
p3 were similar to the p6. Cells grown at p3 and 6 exhibited
increased growth rates, shorter lag-phase periods and higher cell
concentrations, compared with p1 and 8 (each point represents the
number of cells against time). ADSCs, adipose-derived stem cells;
p, passage. |
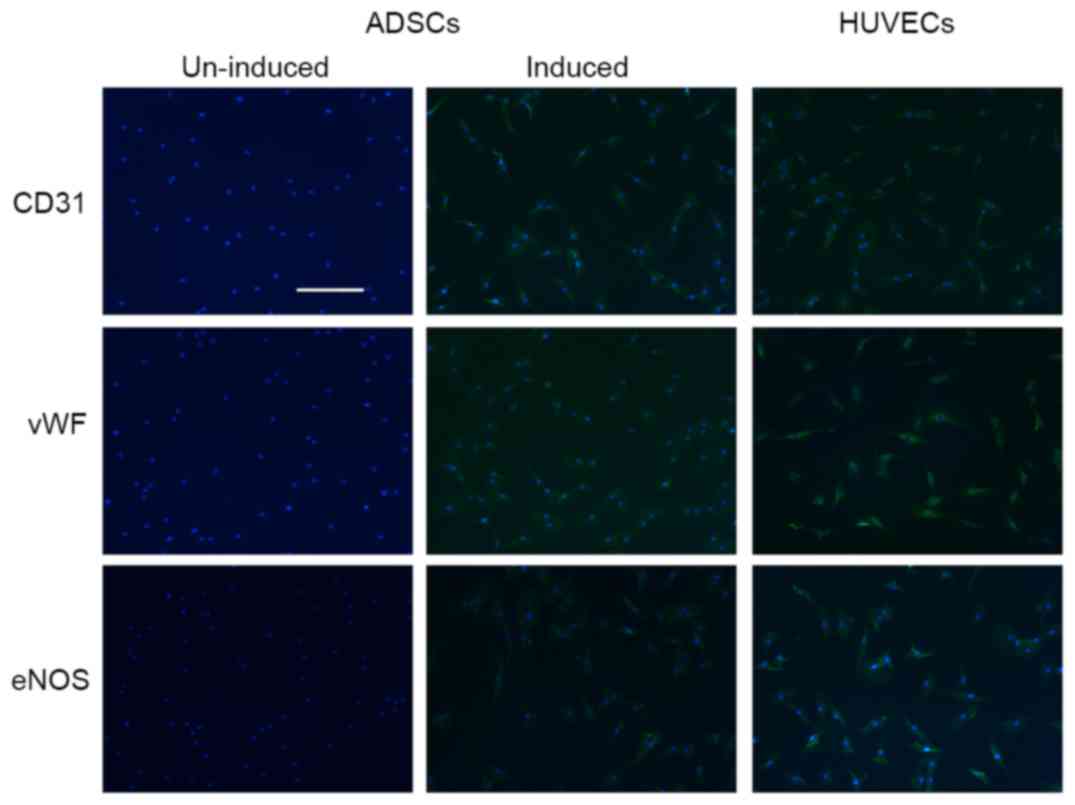
Endothelial phenotype during
endothelial differentiation in ADSCs
ADSCs were subjected to incubation in EGM-2 to
stimulate endothelial differentiation. When cultured in medium with
endothelial cell growth supplement, ADSCs expressed
endothelial-specific markers, including CD31, vWF and eNOS
(Fig. 2). These endothelial
markers were rapidly upregulated during differentiation and
remained elevated 14 days post-differentiation (data not
shown).
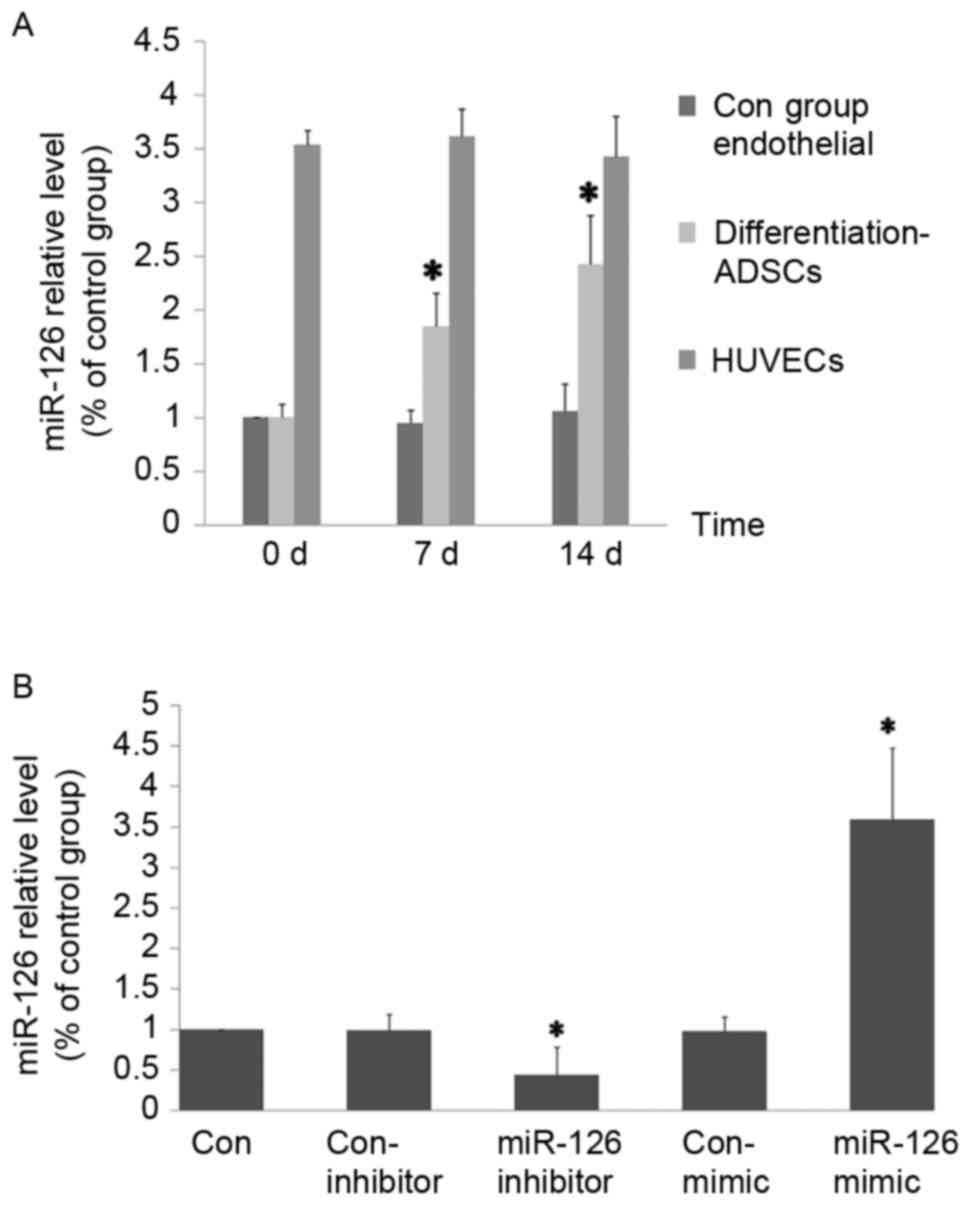
Differential expression of miR-126
during endothelial differentiation of ADSCs
The non-induced and endothelial-induced ADSCs were
harvested on days 0, 7 and 14, and analyzed for miR-126 level using
RT-qPCR analysis, to determine the expression of miR-126 at the
different stages of endothelial differentiation. It was identified
that the expression of miR-126 was enriched in HUVECs and markedly
increased during the endothelial differentiation of ADSCs in a
time-dependent manner (Fig.
3A).
Transfection of ADSCs
In order to study the effect of miR-126 in the
endothelial differentiation of ADSCs, a miR-126 overexpression and
inhibition model was established. After 24 h, transfection
efficiency was detected by Cy3-siRNA transfection control
(siR-Rib). Following transfection with miR-126 mimic or inhibitor
for 3 days, the expression of miR-126 was detected to assess the
efficiency of overexpression and inhibition (Fig. 3B).
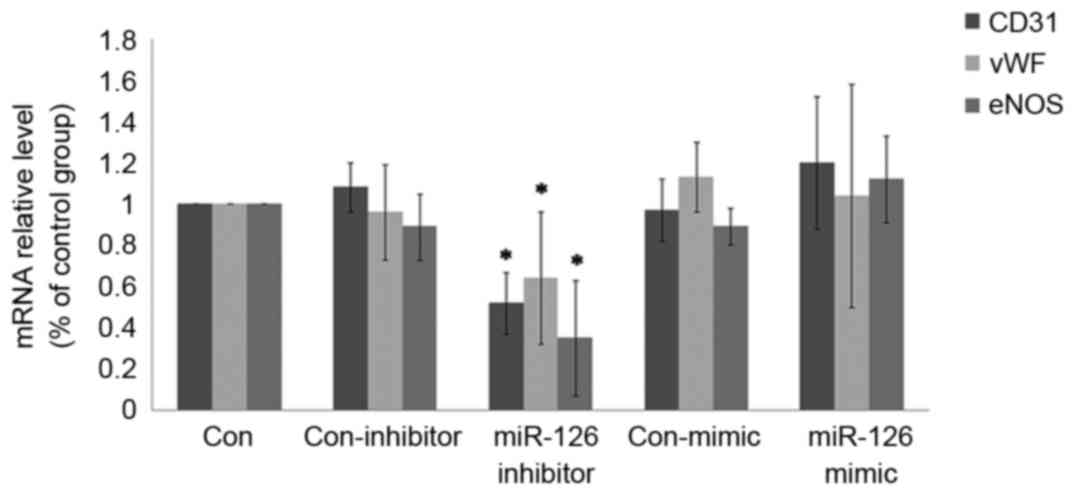
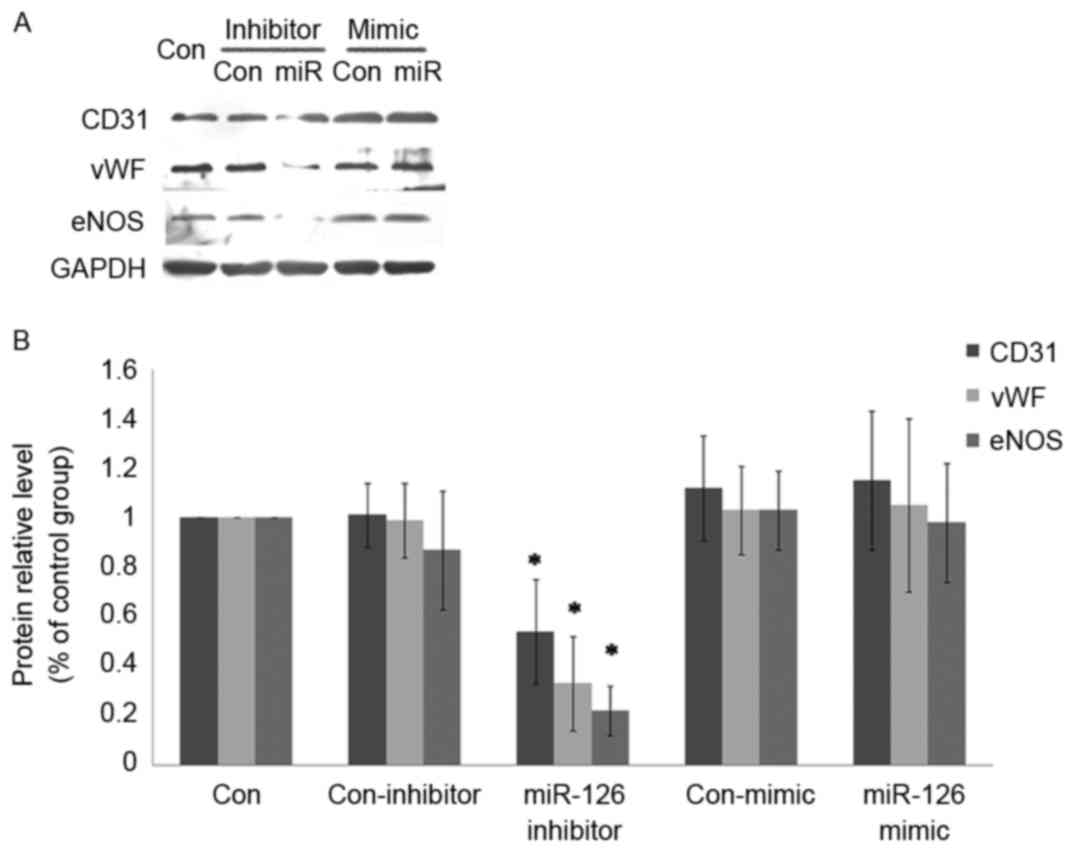
miR-126 modulates the endothelial
phenotype of ADSCs
The expression of CD31, vWF and eNOS largely
mirrored that of endothelial markers during endothelialization. The
expression of these markers was decreased by miR-126 inhibitor
transfection during endothelial differentiation at the mRNA
(Fig. 4) and protein level
(Fig. 5). In addition, ADSCs with
miR-126 overexpression were generated through transfection of a
miR-126 mimic. It was identified that the expression of endothelial
markers was not notably affected by miR-126 overexpression.
Discussion
ADSCs may be easily harvested from lipoaspirate and
the extraction maybe less invasive and less expensive compared with
extraction from bone marrow. Additionally, ADSCs have a
significantly shorter doubling time when expanded in vitro
(18). In the present study, ADSCs
of p3-6were used for the experiments and it was observed that the
majority of the cells exhibited typical fibroblastoid morphology,
expressed MSC surface markers, and demonstrated the capability for
multipotency (data not shown). These intrinsic characteristics and
advantages make ADSCs an ideal stem cell source for the present
study and future cell-based tissue engineering and therapies
(19,20).
In the present study, expression analyses revealed
an abundant level of miR-126 in HUVECs and identified miR-126 as
one of the miRs known to be specifically expressed in the
endothelial cell lineage. Expression of miR-126 gradually increased
during ADSC endothelial differentiation, and expression was marked
when endothelial markers were expressed. Endothelial
differentiation markers (CD31, vWF and eNOS) were used to evaluate
the differentiation capability of ADSCs. The results of the present
study suggest that miR-126 may be involved in the directional
differentiation of ADSCs into vascular endothelial cells, and may
promote the progress of the differentiation.
Due to the high expression of miR-126 in HUVECs, and
the increased expression during the endothelial differentiation of
ADSCs, the potential role of this miR in the regulation of
differentiation towards the endothelial lineage was investigated.
In order to confirm the role of miR-126 in the endothelial
differentiation of ADSCs, down- and up-regulation of miR-126
expression was achieved through transfection withmiR-126 inhibitor
and mimic, respectively. As a result of miR-126 inhibition in ADSCs
undergoing the endothelial differentiation, diminished mRNA
expression of endothelial cell markers was observed. These results
demonstrated that the downregulation of miR-126 may exhibit a
negative effect on endothelial differentiation. By contrast, the
overexpression of miR-126 did not improve the expression levels of
endothelial markers. The results of the present study suggest that
while miR-126 is enriched in vascular endothelial cells, and is
essential to endothelial differentiation, it is not sufficient to
promote the differentiation of ADSCs towards an endothelial
phenotype.
Previous research has demonstrated that miR-126
serves an essential role in stem cell differentiation (21) and angiogenesis. However, the
mechanism remains unknown and there is a lack of consistency
between studies. It is difficult to connect the physiological
functions of miR-126 in vasculature to its functions in tumor. It
has been established that angiogenesis represents one of the key
features in the pathogenesis of cancer (22), but little is known about the role
of miR-126 in tumor neoangiogenesis. The present study suggests
that miR-126 may have a supportive role in the progression of
cancer, which maybe mediated by the promotion of blood vessel
growth. By contrast, it was reported that the downregulation of
miR-126 increased the activity of vascular endothelial growth
factor-A in lung and breast cancer (23–25).
miR-126 is regarded as a putative tumor suppressor due to its
potential role in anti-angiogenesis in cancer (26). Consequently, miR-126 may function
differently in stromal cells compared with tumor cells. The effect
and mechanism of miR-126 on angiogenesis in different cells or
tissue requires further research, to compare the effects of miR-126
in tumor cells with the functions of miR-126 in tumor vasculature
and the surrounding vessels of the tumor periphery.
Acknowledgements
The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81200443).
References
|
1
|
Trounson A and McDonald C: Stem Cell
Therapies in Clinical Trials: Progress and Challenges. Cell Stem
Cell. 17:11–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang Y, Chen XH, Li FG, Chen YX, Gu LQ,
Zhu JK and Li P: In vitro induction of human adipose-derived stem
cells into lymphatic endothelial-like cells. Cell Reprogram.
17:69–76. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu F, Wang X, Liang G, Lv L, Zhu Y, Sun B
and Xiao Z: Effects of epidermal growth factor and basic fibroblast
growth factor on the proliferation and osteogenic and neural
differentiation of adipose-derived stem cells. Cell Reprogram.
15:224–232. 2013.PubMed/NCBI
|
|
4
|
De Ugarte DA, Morizono K, Elbarbary A,
Alfonso Z, Zuk PA, Zhu M, Dragoo JL, Ashjian P, Thomas B, Benhaim
P, et al: Comparison of multi-lineage cells from human adipose
tissue and bone marrow. Cells Tissues Organs. 174:101–109. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ikegame Y, Yamashita K, Hayashi S, Mizuno
H, Tawada M, You F, Yamada K, Tanaka Y, Egashira Y, Nakashima S, et
al: Comparison of mesenchymal stem cells from adipose tissue and
bone marrow for ischemic stroke therapy. Cytotherapy. 13:675–685.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Locke M, Feisst V and Dunbar PR: Concise
review: Human adipose-derived stem cells: Separating promise from
clinical need. Stem Cells. 29:404–411. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Khan WS, Adesida AB, Tew SR, Longo UG and
Hardingham TE: Fat pad-derived mesenchymal stem cells as a
potential source for cell-based adipose tissue repair strategies.
Cell Prolif. 45:111–120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fraser JK, Wulur I, Alfonso Z and Hedrick
MH: Fat tissue: An underappreciated source of stem cells for
biotechnology. Trends Biotechnol. 24:150–154. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park IS, Rhie JW and Kim SH: A novel
three-dimensional adipose-derived stem cell cluster for vascular
regeneration in ischemic tissue. Cytotherapy. 16:508–522. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fish JE, Santoro MM, Morton SU, Yu S, Yeh
RF, Wythe JD, Ivey KN, Bruneau BG, Stainier DY and Srivastava D:
miR-126 regulates angiogenic signaling and vascular integrity. Dev
Cell. 15:272–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang S, Aurora AB, Johnson BA, Qi X,
McAnally J, Hill JA, Richardson JA, Bassel-Duby R and Olson EN: The
endothelial-specific microRNA miR-126 governs vascular integrity
and angiogenesis. Dev Cell. 15:261–271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuhnert F, Mancuso MR, Hampton J,
Stankunas K, Asano T, Chen CZ and Kuo CJ: Attribution of vascular
phenotypes of the murine Egfl7 locus to the microRNA miR-126.
Development. 135:3989–3993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sasahira T, Kurihara M, Bhawal UK, Ueda N,
Shimomoto T, Yamamoto K, Kirita T and Kuniyasu H: Downregulation of
miR-126 induces angiogenesis and lymphangiogenesis by activation of
VEGF-A in oral cancer. Br J Cancer. 107:700–706. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nowak WN, Florczyk U, Józkowicz A and
Dulak J: Role of microRNA in endothelial cells-regulation of
differentiation and angiogenesis. Postepy Biochem. 59:405–414.
2013.(In Polish). PubMed/NCBI
|
|
15
|
Yan T, Liu Y, Cui K, Hu B, Wang F and Zou
L: MicroRNA-126 regulates EPCs function: Implications for a role of
miR-126 in preeclampsia. J Cell Biochem. 114:2148–2159. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meister J and Schmidt MH: miR-126 and
miR-126*: New players in cancer. ScientificWorldJournal.
10:2090–2100. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ikegame Y, Yamashita K, Hayashi S, Mizuno
H, Tawada M, You F, Yamada K, Tanaka Y, Egashira Y, Nakashima S, et
al: Comparison of mesenchymal stem cells from adiposetissue and
bonemarrow for ischemicstroke therapy. Cytotherapy. 13:675–685.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nakagami H, Maeda K, Morishita R, Iguchi
S, Nishikawa T, Takami Y, Kikuchi Y, Saito Y, Tamai K, Ogihara T
and Kaneda Y: Novel autologous cell therapy in ischemic limb
disease through growth factor secretion by cultured adipose
tissue-derived stromal cells. Arterioscler Thromb Vasc Biol.
25:2542–2547. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Su SJ, Yeh YT, Su SH, Chang KL, Shyu HW,
Chen KM and Yeh H: Biochanin a promotes osteogenic but inhibits
adipogenic differentiation: Evidence with primary adipose-derived
stem cells. Evid Based Complement Alternat Med. 2013:8460392013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu Z, Yin H, Liu T, Yan W, Li Z, Chen J,
Chen H, Wang T, Jiang Z, Zhou W and Xiao J: MiR-126-5p regulates
osteoclast differentiation and bone resorption in giant cell tumor
through inhibition of MMP-13. Biochem Biophys Res Commun.
443:944–949. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
El-Kenawi AE and El-Remessy AB:
Angiogenesis inhibitors in cancer therapy: Mechanistic perspective
on classification and treatment rationales. Br J Pharmacol.
170:712–729. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu B, Peng XC, Zheng XL, Wang J and Qin
YW: MiR-126 restoration down-regulate VEGF and inhibit the growth
of lung cancer cell lines in vitro and in vivo. Lung Cancer.
66:169–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu X, Li H, Long L, Hui L, Chen H, Wang
X, Shen H and Xu W: miR-126 enhances the sensitivity of non-small
cell lung cancer cells to anticancer agents by targeting vascular
endothelial growth factor A. Acta Biochim Biophys Sin (Shanghai).
44:519–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu N, Zhang D, Xie H, Zhou Z, Chen H, Hu
T, Bai Y, Shen Y, Yuan W, Jing Q and Qin Y: Endothelial-specific
intron-derived miR-126 is down-regulated in human breast cancer and
targets both VEGFA and PIK3R2. Mol Cell Biochem. 351:157–164. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng R, Chen X, Yu Y, Su L, Yu B, Li J,
Cai Q, Yan M, Liu B and Zhu Z: miR-126 functions as a tumour
suppressor in human gastric cancer. Cancer Lett. 298:50–63. 2010.
View Article : Google Scholar : PubMed/NCBI
|