Introduction
Liver injury is a common side effect of
antituberculosis drug treatment, and often leads to the cessation
of tuberculosis treatment and the failure of chemotherapy, it is an
important clinical problem, but its pathogenesis has not been fully
elucidated (1,2). Isoniazid is the preferred drug for
the treatment of tuberculosis; it is mainly metabolized through the
cytochrome P450 (CYP450) enzyme system, and its main side effect is
liver injury (1), which often
leads to discontinuation of medication during the treatment of
tuberculosis.
CYP450 is the main metabolic enzyme that is involved
in phase I reactions and in the metabolism of endogenous and
exogenous compounds (3). Other
studies have reported on correlations between the subtypes of P450
and diseases (45), and previous molecular epidemiological studies
indicated that the genetic polymorphism of CYP450 metabolic enzymes
may be related to the occurrence of anti-tuberculosis drug-induced
liver injury (ADLI) (6). It was
also suggested that ADLI may be related to the decrease of drug
metabolism enzyme activity and the accumulation of metabolites.
Alteration or loss of the activity of the metabolic enzyme may
result in an imbalance of cellular detoxification functions,
resulting in liver disease (7).
In epigenetics, promoter methylation is an important
mechanism for the downregulation or inactivation of gene
expression, except gene mutation and deletion (8), and a study on mammalian found that
CpG dinucleotide methylation is associated with the inhibition of
gene expression. CpG islands in the promoter region are rich in CpG
sites, and under normal circumstances are non-methylated, whereas
CpG sites outside of the CpG islands are usually methylated.
Abnormal changes have taken place in the methylation patterns in
cancers or other diseases (9).
High levels of methylation in CpG islands were reported to result
in the downregulation of gene expression, which may be an
underpinning cause of disease (10–12).
Downregulation of genome-wide methylation levels may be observed in
hepatic inflammatory disease, hysteric apepsia, immune deficiency
and tumors. Genome-wide hypomethylation may result in chromosome
conformation opening up and transcriptional activation, which may
lead to chromosome instability and abnormal gene expression.
Alterations in whole-genome DNA methylation level have been
considered as a biomarker of tumor development (13). In a case-control study, CpG island
methylation in the CYP1A1 promoter region served an important role
in ADLI (14). Animal experiments
also confirmed that CYP2E1 promoter hypermethylation and
genome-wide hypomethylation was associated with liver injury
induced by isoniazid (15,16). These results suggested that CYP1A1
and CYP1B1 promoter CpG island methylation and isoniazid-induced
rat liver injury may be related.
Alterations in toll-like receptor (TLR)-4 mRNA
expression and its signaling pathway were observed in liver
injuries induced by acetaminophen, floxacillin, methimazole,
lipopolysaccharide and alcohol (17). TLR4 has been reported to serve an
important role in drug-induced liver injury, and TLR4 signal
transduction cascades may be related to the cause of these diseases
(18). A previous study on
alcoholic fatty liver ischemia-reperfusion injury demonstrated that
the enzyme activity and gene expression of CYP1A1, CYP1A2 and
CYP2B1 were decreased following ischemia-reperfusion, but the
activity and expression of CYP2E1 were increased (19). The results from this previous study
revealed the overexpression of CYP2E1 in Kupffer cells, which
induce TLR4 activation, produced inflammatory responses and may
lead to liver damage (19). Under
exposure to aflatoxin B1 (AFB1), the observed pronounced increase
in TLR4 transcripts in monocytes may be partly attributed to higher
expression of CYP1A1 and CYP1B1 caused by AFB1 peroxidation in
monocytes (20). Extracellular
signal-regulated kinase (ERK) is downstream of TLR4, and may also
be involved in the regulation of DNA methylation patterns by
regulating methyltransferase. Peroxisome proliferator-activated
receptors (PPARs) are a type of nuclear receptor that may be
activated by the peroxisome proliferator, and that regulate the
inflammatory response, lipid metabolism, cell proliferation and
differentiation in the liver. The regulation of PPAR-γ in liver
diseases may serve an important role. Cyclophosphamide may cause
severe liver damage through CYP metabolism transformation; PPAR-γ
was revealed to be significantly reduced in the liver injury
induced by cyclophosphamide (21).
After receiving liver-protective treatment, activated PPAR-γ
downregulated signal transducer and activator of transcription,
nuclear factor-κB and activator protein-1 signaling pathways,
oxidative stress and the inflammatory response were reduced, and
liver injury was lessened (21).
DNA methylation may be modified by promoter
demethylation, which may increase the expression of the silenced
gene and thus restore the expression. This provides a basis for the
prevention and treatment of disease. Currently, there are few
studies on the relationship between drug metabolism-enzyme
methylation and ADLI; therefore the present study aimed to analyze
the relationship between two important drug metabolizing enzymes
CYP1A1 and CYP1B1 promoter methylation and isoniazid-induced liver
injury in rats and the possible mechanism, to provide insight for
the prevention and treatment of liver injury, and to enhance the
effectiveness of the treatment of tuberculosis.
Materials and methods
Animals and treatments
A total of 56 specific-pathogen free Sprague-Dawley
rats (28 male, 28 female; age, 8–9 weeks) were purchased from
Beijing Hua Fukang Biotechnology Co., Ltd. (Beijing, China). The
present study was approved by the Ethics Committee of North China
University of Science and Technology (Tangshen, China). All rats
were maintained at 22–23°C with 65–69% humidity, a 12/12 h
light/dark cycle and with free access to food and water. Following
1 week of adaptive feeding, the rats were randomly divided into 7
groups (8 rats/group); the control group, which received an equal
volume of distilled water and were sacrificed at day 28, and the
remaining 48 rats were subsequently divided into 6 groups (labeled
as day 3, 7, 10, 14, 21 and 28, respectively), which were gavaged
with isoniazid (55 mg·kg−1·d−1; Shenyang
Hongqi Pharmaceutical Co. Ltd, Shenyang, China) and were
subsequently sacrificed at day 3, 7, 10, 14, 21 and 28. Following
sacrifice, blood and liver tissues were collected. Blood was
collected using K2-EDTA as an anticoagulant; serum was separated by
centrifugation at 4,000 × g for 10 min at 4°C. Serum samples and
liver tissues were stored at −80°C.
Histopathological investigation
Liver specimens were fixed in 10% buffered formalin
solution, embedded in paraffin and sections (4 µm) were stained
with hematoxylin and eosin. Images were captured with an Eclipse
80i optical microscope (Nikon Corporation, Tokyo, Japan).
Pathological changes were observed under ×20 magnification.
Serum alanine aminotransferase (ALT)
and aspartate aminotransferase (AST) levels
3 ml serum was collected following sacrifice, and
the levels of ALT and AST were measured with a Hitachi Biochemical
Analyzer (Hitachi, Ltd., Tokyo, Japan).
DNA and RNA extraction
DNA and RNA were extracted from 20 mg liver tissue
of experimental rats using a Tissue Genomic DNA Extraction kit and
an EASYspin Plus RNA kit (Aidlab Biotechnologies Co., Ltd, Beijing,
China), respectively, according to the manufacturer's protocol.
Absorbance values were measured using an ultraviolet
spectrophotometer to determine the concentration and quality of
extracted nucleic acids, 260/280 nm between 1.8 and 2.0 indicated
that the extracted nucleic acid was superior in quality.
Detection of genome-wide methylation
level
Perchloric acid (70%; 25 µl) was added to DNA (50
µl) and incubated in a 95°C water bath for 50 min. KOH (1 mol/l)
was used to adjust the pH to between 3 and 5, and the samples were
centrifuged at 18,500 × g for 10 min at 4°C, and the supernatant
was aliquoted to a clean tube. Methylation levels were determined
using an Agilent 1200 High Performance Liquid Chromatography (HPLC)
System (Agilent Technologies, Inc., Santa Clara, CA, USA).
Separation was achieved with a 150×4.6 mm, 5 µm Agilent Eclipse
XDB-C18 column (Agilent Technologies, Inc.) using methanol:water
(1:9 v:v) as the mobile phase and a flow rate of 0.3 ml/min. The
injection volume was 20 µl, column temperature was 25°C and the
detection wavelength was 284 nm. Pure products of 5C and 5mC were
used as standards to prepare concentration gradients. External
calibration of peak area vs. concentration was used in the
determination of global DNA methylation levels: 5mC (%) = [5mC /
(5mC + 5C)] × 100%; where 5mC is 5-methylcytosine.
Determination of CpG island
methylation level in CYP1A1 and CYP1B1 promoter regions by
polymerase chain reaction (PCR)
5 µl (200 ng/µl) DNA was modified by EZ DNA
Methylation-Gold Kit (Zymo Research Corp., Irvine, CA, USA)
according to the manufacturer's protocol. Methylated and
unmethylated primers for CYP1A1 and CYP1B1 were synthesized by
Shanghai Shengong Biological Engineering Technology Service, Ltd.
(Shanghai, China) (Table I).
Methylation specific PCR (MSP; Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) using the following cycle
parameters: 95°C for 5 min, followed by 40 cycles at 95°C for 30
sec, 55°C for 30 sec and 72°C for 30 sec, and a final extension at
72°C for 7 min. PCR products (2 µl) were subjected to
electrophoresis on 2% agarose gels and stained with 0.5 µg/ml
ethidium bromide. Optical density values were measured using
Quantity One 4.6.2 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The following formula was used to calculate the results of
methylation: Methylation rate (%) = [ODM /
(ODM + ODU)] × 100%; where U indicates
unmethylated and M indicates methylated.
 | Table I.Expected product size and primer
sequences used for methylation polymerase chain reaction. |
Table I.
Expected product size and primer
sequences used for methylation polymerase chain reaction.
| Gene | Primer sequence
(5′→3′) | Product size
(bp) |
|---|
| CYP1A1 |
MF:GGTTTTTGTTTTTAGGTAGAAGTCG | 202 |
|
|
MR:GCGAATCCCAATACTATCACG |
|
|
|
UF:TGTTAGGTTTTTGTTTTTAGGTAGAAGTT | 208 |
|
|
UR:CACAAATCCCAATACTATCACACT |
|
| CYP1B1 | MF:
GCGGCGATTAAGTATTTTTC | 159 |
|
|
MR:ATCTCCCTAAACGCTACTCG |
|
|
|
UF:TTGGTGGTGATTAAGTATTTTTT | 159 |
|
|
UR:ATCTCCCTAAACACTACTCACAA |
|
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA (50 ng) from liver tissue was reverse
transcribed using an M-MLV kit (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. cDNA
synthesis was conducted at 37°C for 15 sec followed by 85°C for 5
min. cDNA products were amplified using the primers presented in
Table II; GAPDH was used for
normalization of the quantity of cDNA. qPCR was performed with an
ABI StepOne Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was carried out in a 20 µl reaction mixture using
the Platinum SYBR Green qPCR kit (Invitrogen; Thermo Fisher
Scientific, Inc.) containing 2 µl cDNA sample, 10 µl 2X SYBR Green,
0.4 µl ROX and 1.6 µl of the primers (0.8 µl upstream primers and
0.8 µl downstream primers), with 6 µl of DEPC water. The
thermocycling program was: 95°C for 10 min, followed by 40 cycles
of 95°C for 15 sec, 63°C for 30 sec and 72°C for 30 sec (the
annealing temperature of each gene is shown in Table II). Relative expression was
calculated using the 2−ΔΔCq method (22).
| Table II.Primer sequences and annealing
temperatures for reverse transcription-quantitative polymerase
chain reaction. |
Table II.
Primer sequences and annealing
temperatures for reverse transcription-quantitative polymerase
chain reaction.
| Gene | Primer sequence
(5′→3′) | Annealing
temperature (°C) |
|---|
| GAPDH |
F:AGCAACTCCCATTCTTCC | 63 |
|
|
R:GTCCAGGGTTTCTTACTCC |
|
| CYP1A1 |
F:CAGACCCAACACTGGCATC | 63 |
|
|
R:GGGAGGTAACGGAGGATAGG |
|
| CYP1B1 |
F:GAGAGTTGGTGGCAGTGTTG | 63 |
|
|
R:CCAGGACGAAGTTGCTGAA |
|
| TLR4 |
F:GCCTCCCTGGTGTTGGATTT | 57 |
|
|
R:AGCACACTGACCACCGATAC |
|
| ERK |
F:GGCACCAACCATTGAGCAGA | 63 |
|
|
R:GATCATTGCTGAGGTGCTGTGTC |
|
| PPAR-γ |
F:CGTCCCCGCCTTATTATTCT | 63 |
|
|
R:GCTTTATCCCCACAGACTCG |
|
| IL-6 |
F:ACAGCGATGATGCACTGTCA | 60 |
|
|
R:AGCACACTAGGTTTGCCGAG |
|
| TNF-α |
F:CTCAAGCCCTGGTATGAGCC | 60 |
|
|
R:GGCTGGGTAGAGAACGGATG |
|
ELISA
Liver tissue (10 g) was used to prepare a 10% (w/v)
tissue homogenate with phosphate-buffered-saline (PBS) (0.02 M
sodium phosphate buffer with 0.15 M sodium chloride, pH 7.4). The
homogenate was centrifuged at 4,500 × g for 10 min at 0°C. Protein
expression levels of CYP1A1 and CYP1B1 were determined using rat
ELISA kits (P4501A1 and P4501B1, respectively) (both from Beijing
Winter Song Boye Biotechnology Co., Ltd, Beijing, China), according
to the manufacturer's protocols. Absorbance was measured at 450 nm
with a microplate reader.
Superoxide dismutase (SOD) and
malondialdehyde (MDA) levels
SOD activity and MDA content was detected using a
Malondialdehyde assay kit and a Superoxide Dismutase assay kit
(Jiancheng Bioengineering Ltd, Nanjing, China) according to the
manufacturer's instructions. Homogenates were prepared as
aforementioned and diluted prior to use as follows: SOD activity,
10% homogenate (w/v); MDA content, 0.25% homogenate (w/v).
Absorbance was measured at 550 and 532 nm with a microplate reader,
respectively.
Statistical analysis
Data are presented as the mean ± standard deviation.
Difference among groups was tested by one-way analysis of variance
with Fisher's least significant difference or Dunnett's T3 (equal
variances not assumed following the variable transformation
justification) post hoc test using SPSS 17.0 software (SPSS, Inc.,
Chicago, IL, USA). Correlation analysis using Pearson's correlation
analysis reflected the true correlation of two variables using
partial correlation analysis. P<0.05 was considered to indicate
a statistically significant difference.
Results
Validation of rat liver injury
model
Tissue pathology is the gold standard for the
diagnosis of liver injury, and ALT and AST levels are important
indicators of liver function and pathological changes in the
laboratory. H&E stained sections of rat liver tissues were
examined under a light microscope to observe the pathological
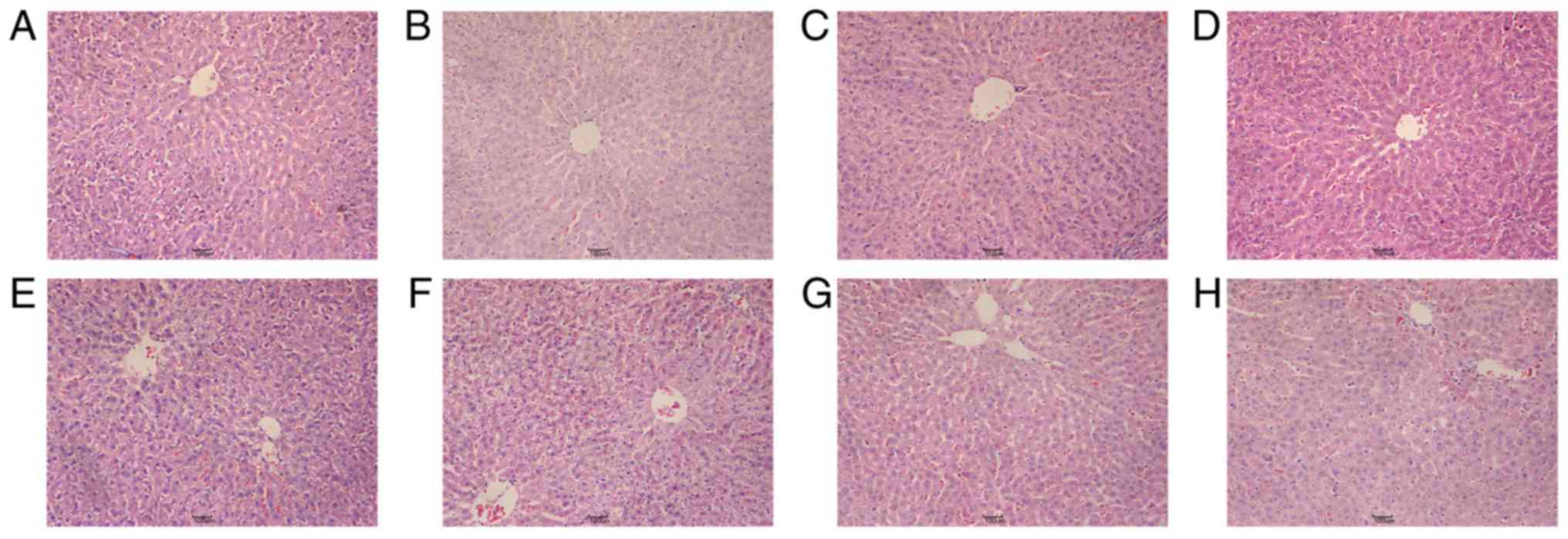
changes (Fig. 1). In the normal
control group, the liver cells appeared normal, the structure of
the liver was complete, liver cells were in radially arranged
cords, with the central vein as the center, arranged neatly and
clearly defined (Fig. 1A). In the
rats groups treated with isoniazid, a large number of inflammatory
cells infiltration and hepatic edema was observed, and the
polygonal arrangement became spherical in shape with irregular
structure at day 10 (Fig. 1D). At
day 14, the cytoplasm was loose and balloon-like degeneration was
diffuse, liver cell structure was disordered and partial liver
cells were degenerated and necrotic (Fig. 1E). At day 28, the structure of
hepatic leaflets was completely destroyed, and most of the hepatic
cells were necrotic, focal necrosis and bridging necrosis were
clearly visible (Fig. 1G).
Alterations in the levels of ALT and AST, as measured by automatic
biochemical analyzer, were also revealed to increase with time of
following isoniazid administration (Table III), and the difference was
statistically significant from 10 day (P<0.05). The pathological
results and liver function indices indicated that isoniazid
treatment successfully induced rat liver injury in the animal
models.
| Table III.Levels of serum alanine
aminotransferase and aspartate aminotransferase in each group
following administration of isoniazid. |
Table III.
Levels of serum alanine
aminotransferase and aspartate aminotransferase in each group
following administration of isoniazid.
| Group | ALT
(U/l−1) | AST
(U/l−1) |
|---|
| Control |
20.23±4.22 |
80.28±26.41 |
| Day 3 |
28.98±10.96 |
89.16±16.92 |
| Day 7 |
34.85±22.72 |
119.95±52.24 |
| Day 10 |
83.85±24.35a |
189.97±35.80a |
| Day 14 |
114.50±33.33a |
243.28±62.16a |
| Day 21 |
118.25±24.60a |
202.36±101.91a |
| Day 28 |
96.10±26.25a |
170.63±77.52a |
Analysis of genome methylation
level
Whole-genome methylation status is related to the
stability of the genome. HPLC analysis provided a good separation
of 5C and 5mC; the standard curves of 5C and 5mC were
Y=75.194X-16.914 (r=0.999; Fig.
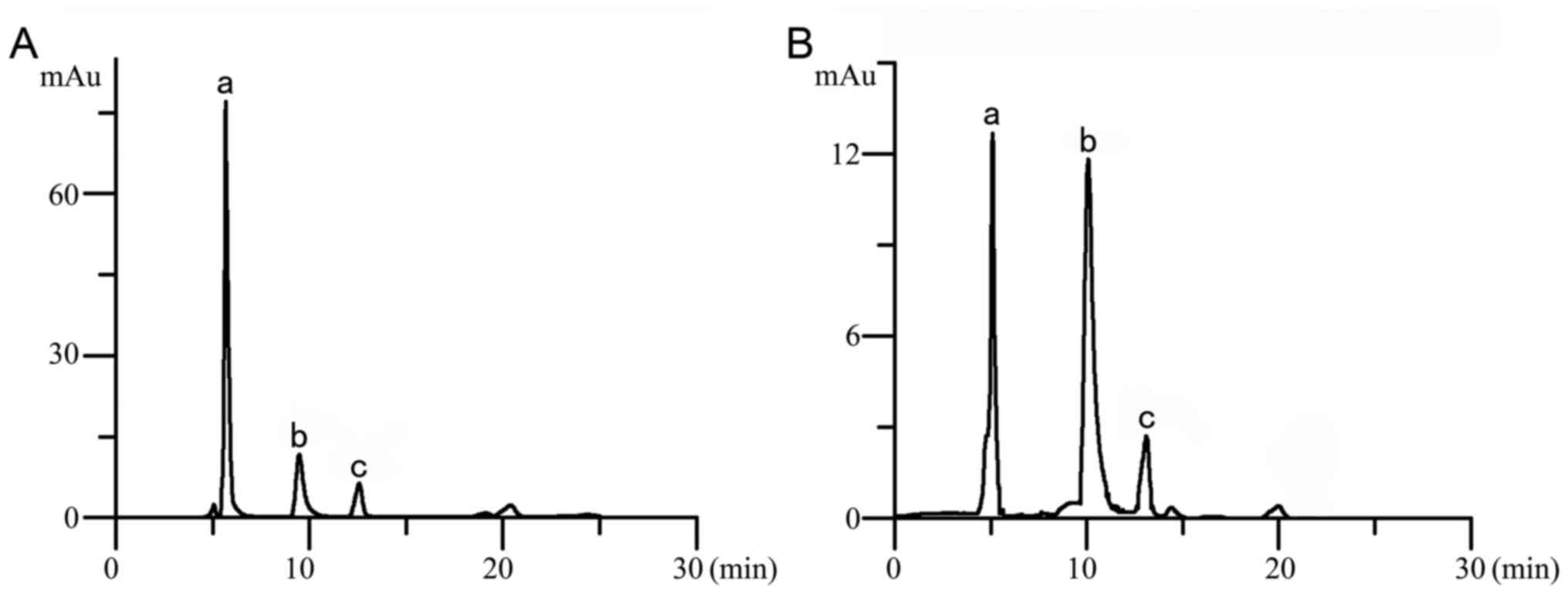
2A), Y=89.829X-6.7363 (r=0.999; Fig. 2B), respectively. Chromatograms of
the control group and a representative experimental group are
presented in Fig. 3A and B,
respectively. Whole-genome methylation levels at days 7, 10, 14, 21
and 28 were significantly decreased compared with the control
group, which indicated that the whole-genome methylation level was
abnormal.
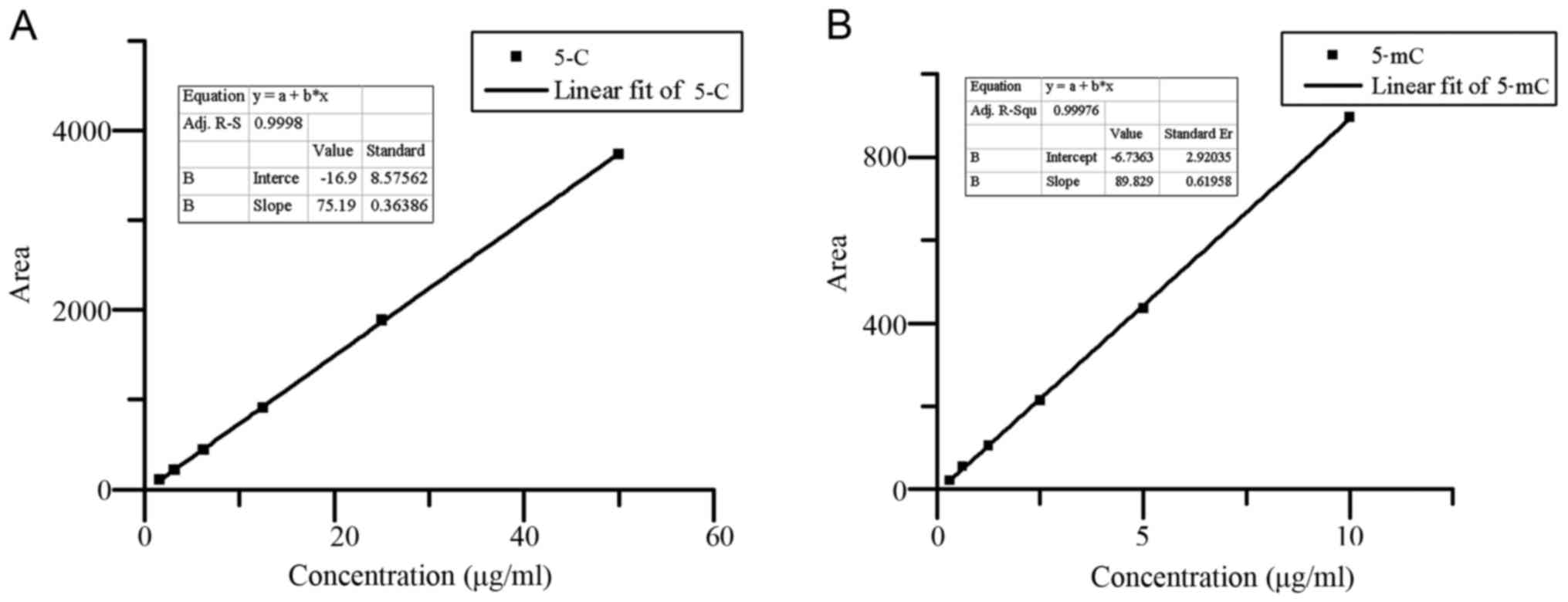 | Figure 2.Standard curves for series
concentrations of 5C and 5mC. (A) 5C as standard; the concentration
series was 50, 25, 12.5, 6.25, 3.175 and 1.5875 µg/ml. (B) 5mC as
standard; the concentration series was 10, 5, 2.5, 1.25, 0.625 and
0.3125 µg/ml. 5C, 5-cytosine; 5mC, 5-methylcytosine. |
Analysis of methylation levels of CpG
islands in CYP1A1 and 1B1 promoter regions and mRNA and protein
expression levels
Gene promoter region-specific hypermethylation and
whole-genome hypomethylation occur side-by-side within the same
individual in certain disease states. The abnormal methylation
pattern of promoter region can lead to the change of gene
expression, and then participate in the occurrence of disease
(23). Isoniazid may also induce
these abnormalities in liver injury. The present study used MSP to
measure the methylation level of CpG islands in the gene promoter
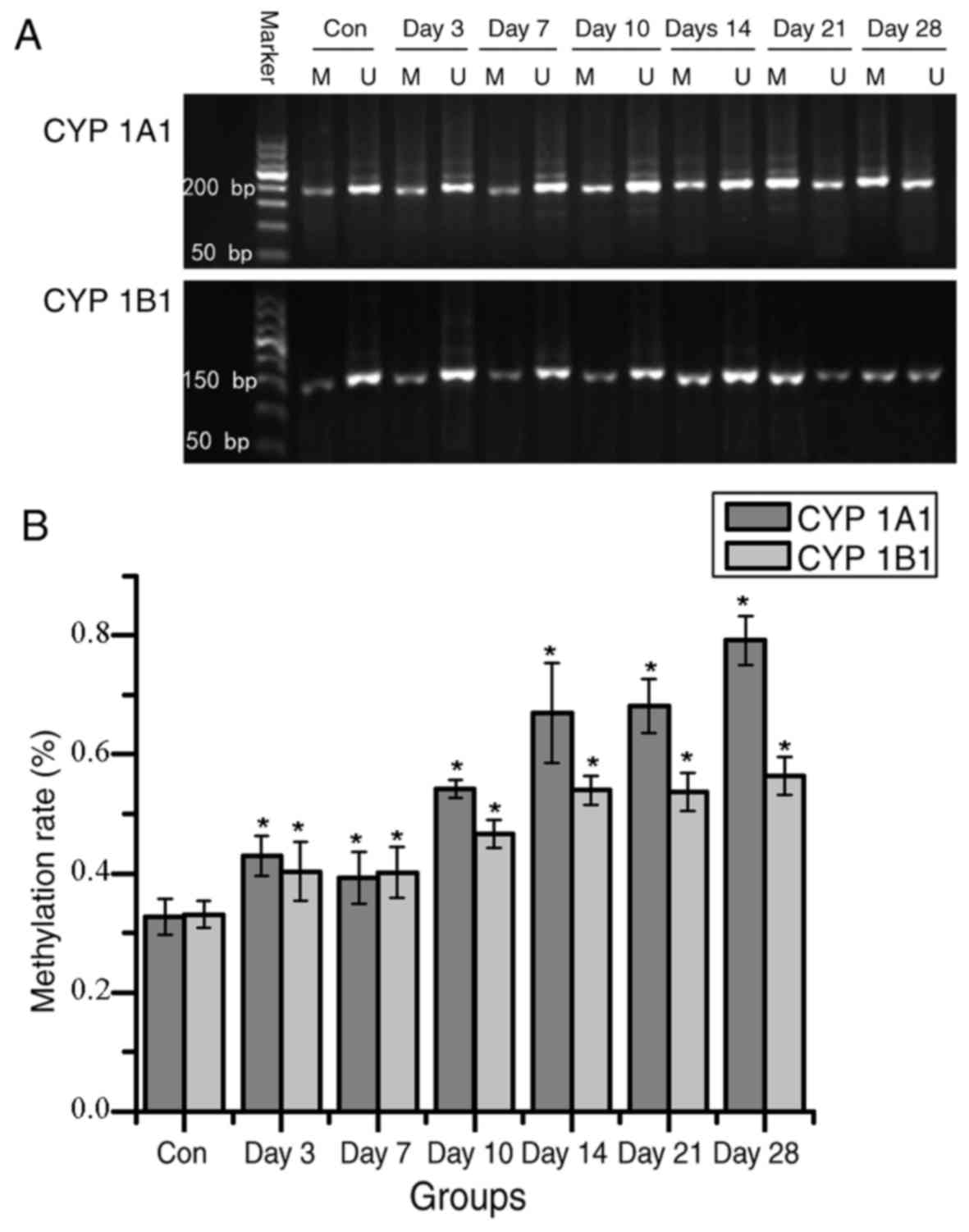
regions. Compared with the control group, CpG island methylation
levels of the CYP1A1 and CYP1B1 promoter regions significantly
increased over time from day 3 following isoniazid administration
(Fig. 4). Alterations in
methylation levels were detected earlier than the changes in liver
function and liver tissue pathology, which indicted that
methylation of the gene promoter region may be able to regulate the
occurrence of liver injury.
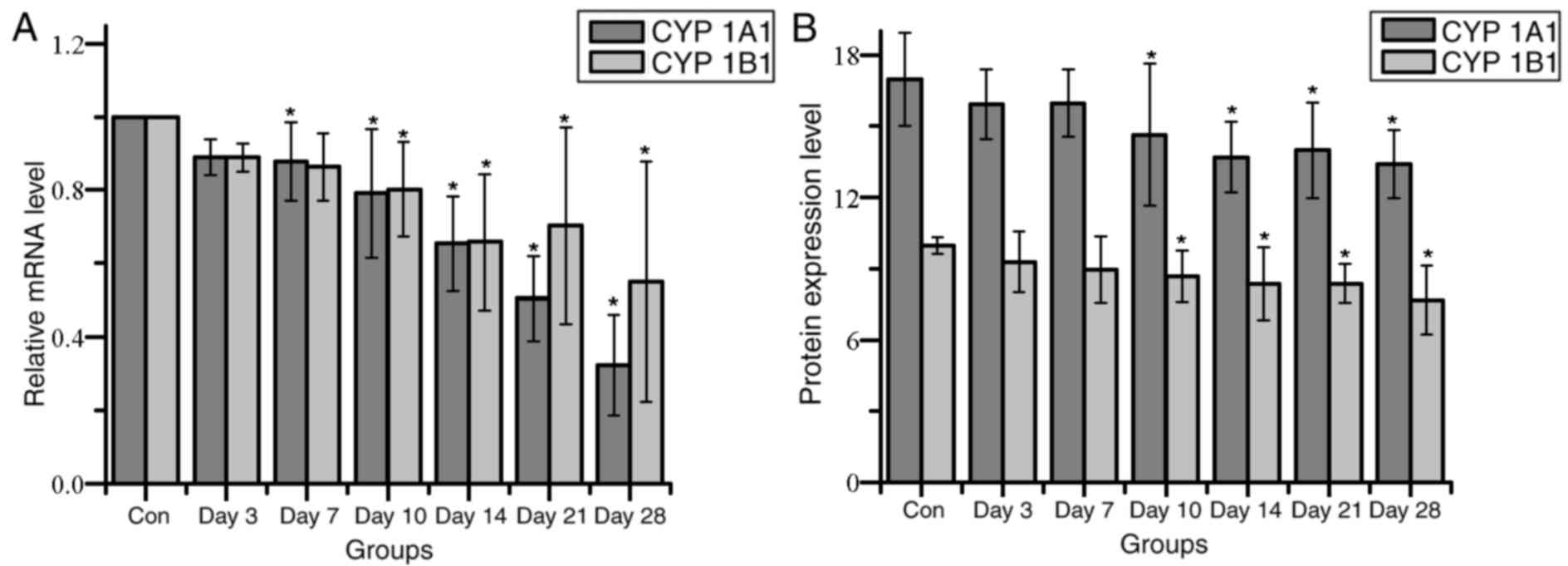
CYP1A1 and CYP1B1 mRNA expression levels exhibited a
downward trend over time (Fig.
5A). CYP1A1 mRNA expression levels were significantly different
from day 7 post-isoniazid treatment, whereas CYP1B1 expression was
significantly lower from 10 day compared with their respective
untreated control group. Protein expression levels of both CYP1A1
and CYP1B1 were significantly decreased from day 10 compared with
their respective controls (Fig.
5B).
With increasing time post-treatment, the methylation
of CpG islands in the CYP1A1 and CYP1B1 gene promoter regions
gradually increased, whereas the gene expression level was
gradually decreased. Correlation analysis indicated that the
methylation levels of the CYP1A1 promoter region were negatively
correlated with its mRNA and protein expression levels (r=−0.824
and −0.518 respectively; P<0.05); similar results were indicated
for CYP1B1 mRNA and protein expression levels compared with
promoter methylation level (r=−0.559 and −0.420, respectively;
P<0.05). The correlation between CpG island methylation level
and gene expression level in CYP1A1 was higher than that for
CYP1B1, which suggested that the expression of CYP1A1 may be more
susceptible to regulation by methylation of CpG islands in the
promoter region.
Correlation analysis also demonstrated that the
CYP1A1 mRNA expression levels were negatively correlated with the
expression of ALT and AST (r=−0.632 and −0.403, respectively;
P<0.05), and similar results were noted for CYP1B1 and ALT and
AST expression levels (r=−0.395 and −0.187, respectively;
P>0.05). Compared with CYP1B1, the correlation coefficient of
CYP1A1 and ALT and AST was greater and indicated a potentially
closer relationship between CYP1A1 and isoniazid-induce liver
injury compared with CYP1B1 and liver injury.
Alterations in mRNA expression level
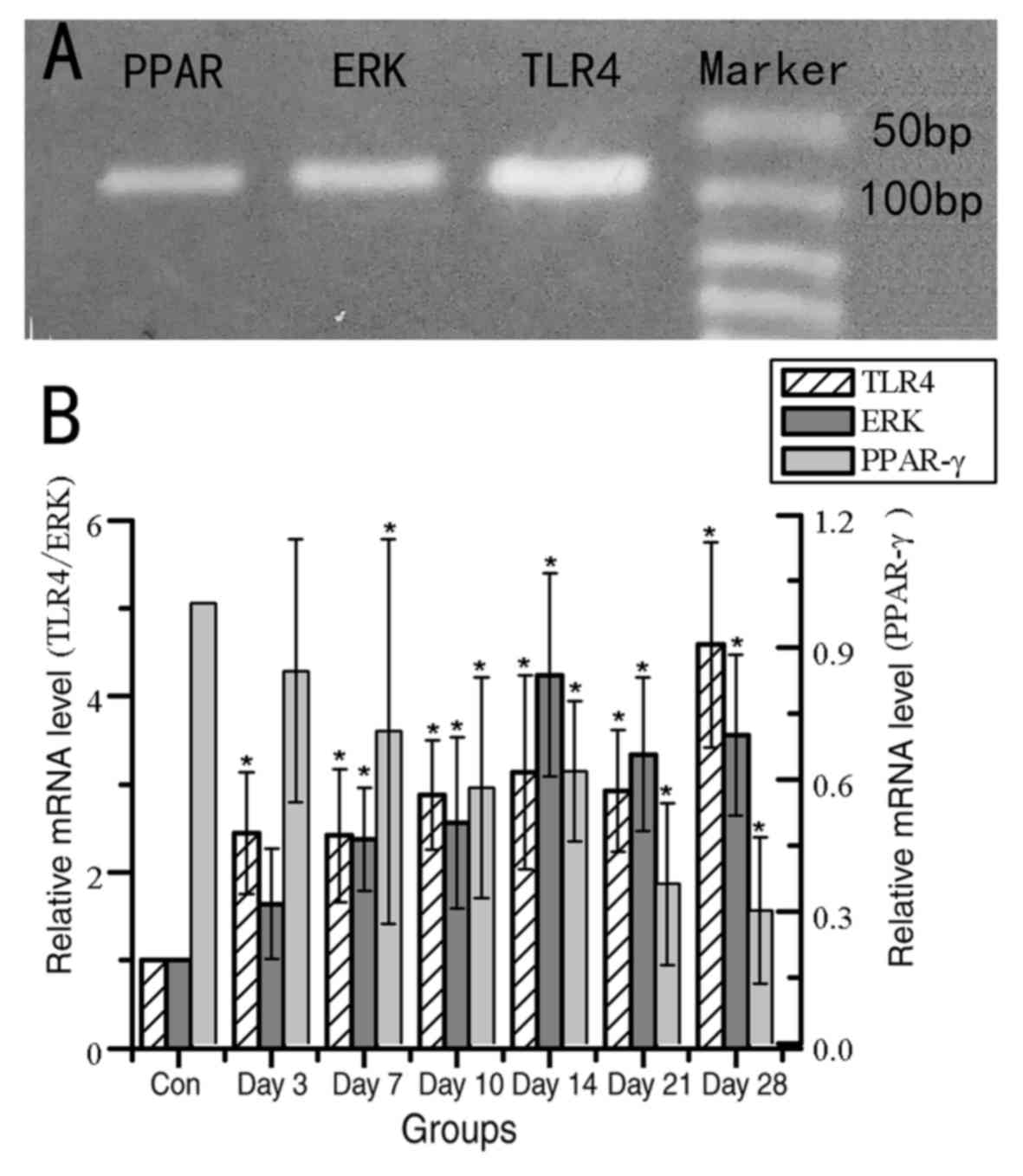
of TLR4, ERK, PPAR-γ
Previous studies have indicated that the changes in
TLR4, ERK, PPAR-γ signaling were regulated by CYPs, and were
related to liver injury (17–21).
Therefore, the relative mRNA expression levels of TLR4, ERK and
PPAR-γ were evaluated by RT-qPCR following isoniazid exposure
(Fig. 6). The expression levels of
TLR4 and ERK exhibited an upward trend from day 3 and 7,
respectively, and these expression levels were significantly
increased compared with their respective controls. The expression
of PPAR-γ exhibited a significant decrease in mRNA expression from
day 7 compared with expression in the untreated control rats. These
results indicated that the TLR4 signaling pathway and PPAR-γ may
participate in liver injury induced by isoniazid. However, it is
unknown whether the changes in these signaling pathway related
factors are related to the expression changes of CYP1A1 and
CYP1B1.
Correlation analysis suggested that TLR4 was
positively correlated with ALT and AST expression (r=0.449 and
0.355, respectively; P<0.05), and negatively correlated with the
mRNA expression levels of CYP1A1 and CYP1B1 (r=−0.627 and −0.647,
respectively; P<0.05). ERK was also positively correlated with
ALT and AST (r=0.634 and 0.499, respectively; P<0.05), and
negatively correlated with the mRNA expression level of CYP1A1 and
CYP1B1 (r=−0.541 and −0.436, respectively; P<0.05). PPAR-γ was
negatively correlated with ALT and AST expression (r=−0.512 and
−0.409, respectively; P<0.05), and positively correlated with
the mRNA expression level of CYP1A1 and CYP1B1 (r=0.599 and 0.387,
respectively; P<0.05).
Partial correlation analysis was performed to
control the mRNA expression level of CYP1A1 and CYP1B1, and the
correlation coefficients of TLR4, ERK and PPAR-γ mRNA, and ALT and
AST were significantly decreased. The partial correlation
coefficients of mRNA expression of TLR4 and ALT and AST were 0.078
and 0.193, respectively (P>0.05). The partial correlation
coefficients of mRNA expression of ERK and ALT and AST were 0.449
and 0.384, respectively (P<0.05). The partial correlation
coefficients of mRNA expression of PPAR-γ and ALT and AST were
−0.213 and −0.233, respectively (P>0.05). These data
demonstrated that the correlation is significantly reduced, which
indicated that the expression level of TLR4, ERK and PPAR-γ in
liver injury may be regulated by the expression of CYP1A1 and
CYP1B1.
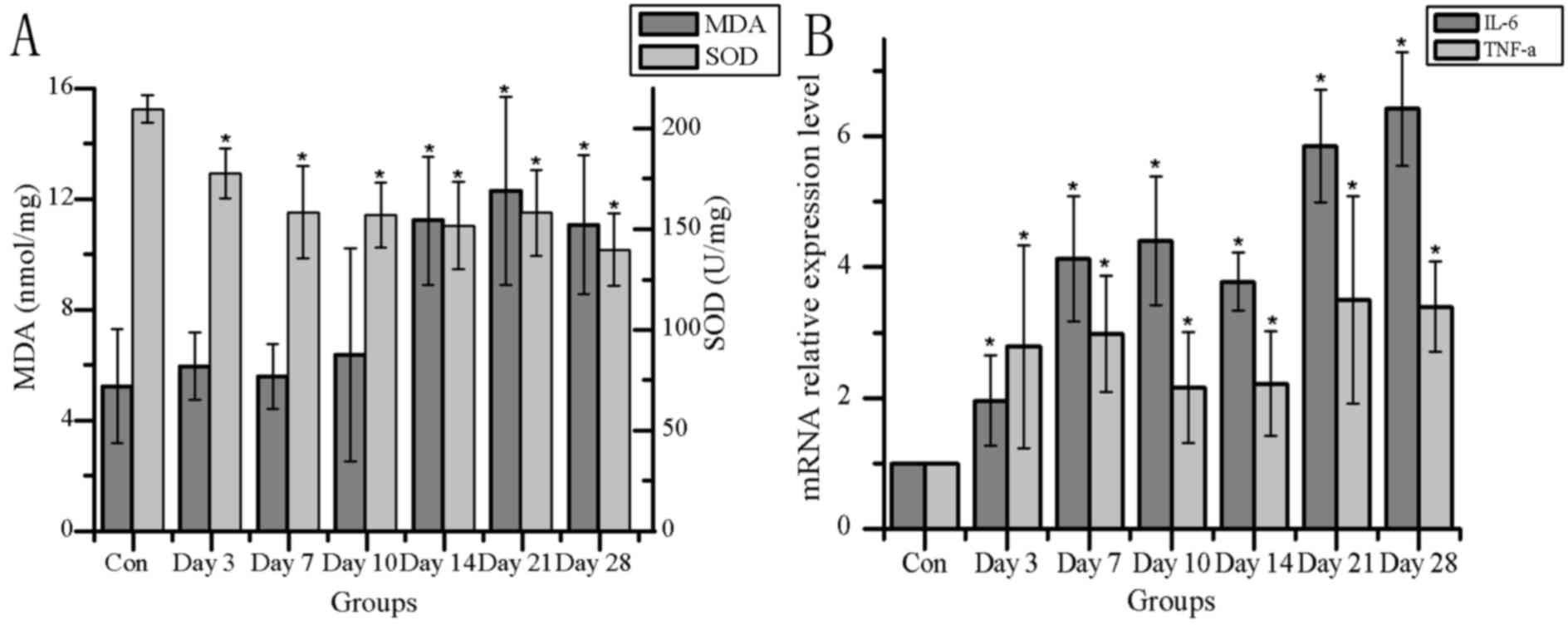
Detection of oxidative stress and
immune-related indices
Oxidative stress and inflammation are involved in
many kinds of liver injury, and TLR4 signaling and PPAR-γ may alter
downstream gene expression or signal pathways to affect changes in
IL-6, TNF-α, MDA and SOD. Alterations in MDA content, SOD activity
and mRNA expression of immune inflammatory factors IL-6 and TNF-α
were examined (Fig. 7). MDA
content increased significantly at 14 days post-isoniazid treatment
compared with the control group, and SOD activity significantly
decreased from 3 days post-treatment (Fig. 7A). Correlation analysis of MDA and
SOD with ALT and AST levels and CYP1A1 and CYP1B1 mRNA expression
indicated that MDA was positively correlated with the degree of
liver injury (r=0.582 and 0.499, respectively; P<0.05), and
negatively correlated with the expression of CYP1A1 and CYP1B1
(r=−0.603 and −0.459, respectively; P<0.05). SOD was negatively
correlated with liver injury (r=−0.582 and −0.453, respectively;
P<0.05), and positively correlated with the expression of CYP1A1
and CYP1B1 (r=0.524 and 0.347, respectively; P<0.05). Through
partial correlation analysis of MDA and SOD with AST and ALT, it
was demonstrated that excluding the effects of CYP1A1 and 1B1
expression alterations, the correlation coefficients between MDA
and SOD with ALT and AST were decreased, indicating that the
relationship became weak.
The mRNA expression of IL-6 and TNF-α exhibited
significant increases from day 3 post-treatment, compared with
their respective controls (Fig.
7B). Correlation analysis indicated that IL-6 were positively
correlated with liver injury (r=0.589 and 0.492, respectively;
P<0.05); the mRNA expression of IL-6 was negatively correlated
with CYP1A1 and CYP1B1 expressions (r=−0.750 and −0.502,
respectively; P<0.05). TNF-α was also positively correlated with
liver injury (r=0.356, P<0.05; r=0.234, P>0.05); the mRNA
expression of TNF-α was negatively correlated with CYP1A1 and
CYP1B1 expressions (r=−0.432, P<0.05; r=−0.243, P>0.05).
Partial correlation analysis was performed to control the
expression of CYP1A1 and CYP1B1; the correlations between IL-6,
TNF-α and ALT, AST were decreased, and the correlation was not
significant. These results indicated that the changes in the
expressions of MDA, SOD, IL-6 and TNF-α in liver injury may be
related to the expression levels of CYP1A1 and CYP1B1.
Discussion
The present study selected epigenetic mechanisms of
drug-induced liver injury as the entry point to study the role of
CpG island methylation in the CYP1A1 and CYP1B1 promoter regions in
the process and mechanism of isoniazid-induced rat liver injury.
Experiments were conducted to confirm that the highly methylated
CpG islands in the CYP1A1 and CYP1B1 promoter regions may induce
low expression of genes, so as to regulate rat liver injury induced
by isoniazid, which may occur through the upregulation of TLR4 and
ERK expression, the downregulation of PPAR-γ expression and by
promoting oxidative stress and inflammatory reaction.
Global DNA methylation is a well-known epigenetic
feature of disease. Medicine may cause changes in the levels of
genome methylation, which may lead to alterations in gene activity
and phenotype. DNA hypomethylation is an important marker of
disease, which can activate the repetitive sequence of the genome,
affect the stability of the genome and increase the risk of cancer,
atherosclerosis, Alzheimer's disease, psychiatric disorders and
other diseases (23). Gene
promoter region-specific hypermethylation and whole-genome
hypomethylation are frequently found in tumor cells (24,25).
In the present study, the methylation level of the whole genome was
detected by HPLC. The results revealed that the methylation level
of whole genome was significantly reduced by day 7 post-isoniazid
treatment, and reached its lowest levels at day 28, which was ~26%
the methylation rate of the control group. The overall methylation
level of DNA was downregulated, which was consistent with previous
studies (16).
Changes in DNA methylation may affect transcription
initiation and lead to the occurrence of disease and cancer. CpG
island methylation is more common than whole-genome methylation,
and CpG island methylation levels are altered as early as the
whole-genome methylation level, is an early event of disease
(24). It has been previously
reported that high methylation of the CYP1A1 gene may cause
transcriptional inactivation, which leads to the decrease of
metabolic activation. DNA methylation may be used to make early
prediction and prognosis evaluations for the occurrence and
development of liver cancer (26,27).
Overexpression of CYP1B1 in the liver may be one of the increased
risks of liver cancer in patients with liver disease (28). In the present study, MSP was used
to detect the methylation level of CYP1A1 and CYP1B1 promoters in
the liver tissue rats treated with intragastric administration of
isoniazid. The results revealed that the CYP1A1 and CYP1B1 promoter
regions were highly methylated; methylation levels at each time
point were higher than that in the control group, and there was a
significant difference from day 3. Alterations in methylation in
the experimental rats were noted to occur in the early stage.
RT-qPCR and ELISA demonstrated that both mRNA and protein
expression of the two genes were decreased. Along with the
upregulation of CpG island methylation in the promoter region of
the gene, the gene expression level was downregulated, which was
consistent with previous studies (29–31).
DNA methylation leads to transcriptional inactivation or reduction
and may by directly interfere with transcription factor binding or
may indirectly attract methylated DNA binding factor, followed by
the recruitment of histone deacetylase to generate invalid
heterochromatin structure (32,33).
The present study revealed that CYP1A1 and CYP1B1
promoter region CpG islands were highly methylated and their
expression levels were reduced, but the changes of CYP1A1 were more
obvious compared with those in CYP1B1. A previous study reported
that estrogen receptor α (ERα) represses CYP1A1 expression by
targeting DNA (cytosine-5)-methyltransferase 3B (Dnmt3B) and the
associated DNA methylation of the promoter, but it had no effect on
CYP1B1 (34). Variations in the
expression levels of Dnmt3B isoforms may partly explain the effects
of ERα on CYP1A1 regulation (34).
Therefore, the differences in CYP1A1 and CYP1B1 methylation levels
in the present study may be due to regulation by different types of
methyltransferases. The expression of CYP1A1 was higher compared
with CYP1B1 in a previous study of liver basal expression (35). Abnormal methylation of CYP1B1 and
changes in expression levels are frequently observed in tumors and
other malignant diseases (36).
These data indicated that CYP1A1 methylation and gene expression
are more evident compared with the changes of CYP1B1 in
isoniazid-induced liver injury, which supports the conclusions made
by the present study.
The TLR4 signaling pathway and PPAR-γ have been
reported to be involved in the occurrence of liver injury in
different degrees (17,21). A number of studies on the
prevention and treatment of liver injury and liver fibrosis have
also demonstrated that drugs may reduce the production of
inflammatory factors and alleviate lipid peroxidation by
downregulating TLR4 and ERK expression and upregulating PPAR-γ
expression, thereby effectively reducing liver disease (37–39).
In the present study, the expression of TLR4 and ERK were
upregulated and the expression of PPAR-γ was downregulated compared
with the control group, which was consistent with the results of
previous studies on liver injury. These changes were significantly
correlated with the mRNA expression level of CYP1A1 and CYP1B1
(P<0.05). Partial correlation analysis indicated that the
expression level of TLR4, ERK and PPAR-γ were affected by the
expression levels of CYP1A1 and CYP1B1 in liver injury. Therefore,
altered expression of CYP1A1 and CYP1B1 through the changes of
TLR4, ERK and PPAR-γ may regulate the occurrence of hepatic injury
induced by isoniazid. However, the processes of isoniazid-induced
liver injury are complex, and involve changes to multiple signaling
pathways and related factors, whereas the present study only
discusses the change of TLR4 signal pathway and the expression of
PPAR-γ.
Oxidative stress and inflammation are closely
related to liver injury; activation of the TLR4 signaling pathway
may lead to the production of inflammatory factors and activation
of immune cells, which may contribute to the occurrence of liver
injury (17). The powerful
anti-inflammatory and anti-oxidative effects of PPAR-γ also serve
an irreplaceable role in the liver. The present study demonstrated
that MDA content increased, SOD activity decreased and the
expression levels of IL-6 and TNF-α mRNA increased. The changes of
these factors were significantly correlated with the expression of
CYP1A1 and CYP1B1, which suggested that the expression of CYP1A1
and CYP1B1, by upregulating the expression of TLR4 signaling
pathway and downregulating PPAR-γ, aggravated the oxidative damage
and immune damage, and further regulated the occurrence of liver
injury.
In conclusion, CpG island hypermethylation of CYP1A1
and CYP1B1 promoter regions regulated the occurrence of rat liver
injury induced by isoniazid, and the mechanism may be through
effecting the expression of TLR4 signaling pathway and PPAR-γ, thus
contributing to oxidative stress and inflammation, leading to liver
damage. However, additional experiments are required to verify the
present results and, as only one aspect of the possible mechanisms
in isoniazid-induced liver injury was discussed, future studies may
identify potential methyltransferase inhibitors and other
mechanisms related to liver injury.
Acknowledgements
Thanks to the strong support of The National Natural
Science Foundation of China (grant no. 81041096) and The Innovation
and Entrepreneurship Training Program for College Students in Hebei
Province (grant no. X2015051).
References
|
1
|
Huang YS: Recent progress in genetic
variation and risk of antituberculosis drug-induced liver injury. J
Chin Med Assoc. 77:169–173. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang S, Pan H, Peng X, Lu H, Fan H, Zheng
X, Xu G, Wang M and Wang J: Preventive use of a hepatoprotectant
against anti-tuberculosis drug-induced liver injury: A randomized
controlled trial. J Gastroenterol Hepatol. 31:409–416. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Corsini A and Bortolini M: Drug-induced
liver injury: The role of drug metabolism and transport. J Clin
Pharmacol. 53:463–474. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hrycay EG and Bandiera SM: Involvement of
cytochrome P450 in reactive oxygen species formation and cancer.
Adv Pharmacol. 74:35–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shahabi P, Siest G, Meyer UA and
Visvikis-Siest S: Human cytochrome P450 epoxygenases: Variability
in expression and role in inflammation-related disorders. Pharmacol
Ther. 144:134–161. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feng FM, Guo M, Chen Y, Li SM, Zhang P,
Sun SF and Zhang GS: Genetic polymorphisms in metabolic enzymes and
susceptibility to anti-tuberculosis drug-induced hepatic injury.
Genet Mol Res. 13:9463–9471. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Turesky RJ and Le Marchand L: Metabolism
and biomarkers of heterocyclic aromatic amines in molecular
epidemiology studies: Lessons learned from aromatic amines. Chem
Res Toxicol. 24:1169–1214. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rodenhiser D and Mann M: Epigenetics and
human disease: Translating basic biology into clinical
applications. CMAJ. 174:341–348. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Picascia A, Grimaldi V, Pignalosa O, De
Pascale MR, Schiano C and Napoli C: Epigenetic control of
autoimmune diseases: From bench to bedside. Clin Immunol. 157:1–15.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Benakanakere M, Abdolhosseini M, Hosur K,
Finoti LS and Kinane DF: TLR2 promoter hypermethylation creates
innate immune dysbiosis. J Dent Res. 94:183–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeybel M, Hardy T, Robinson SM, Fox C,
Anstee QM, Ness T, Masson S, Mathers JC, French J, White S and Mann
J: Differential DNA methylation of genes involved in fibrosis
progression in non-alcoholic fatty liver disease and alcoholic
liver disease. Clin Epigenetics. 7:252015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong Y, Zhao H, Li H, Li X and Yang S: DNA
methylation as an early diagnostic marker of cancer (Review).
Biomed Rep. 2:326–330. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Crary-Dooley FK, Tam ME, Dunaway KW,
Hertz-Picciotto I, Schmidt RJ and LaSalle JM: A comparison of
existing global DNA methylation assays to low-coverage whole-genome
bisulfite sequencing for epidemiological studies. Epigenetics.
12:206–214. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He L, Gao L, Shi Z, Li Y, Zhu L, Li S,
Zhang P, Zheng G, Ren Q, Li Y, et al: Involvement of cytochrome
P450 1A1 and glutathione S-transferase P1 polymorphisms and
promoter hypermethylation in the progression of anti-tuberculosis
drug-induced liver injury: A case-control study. PLoS One.
10:e01194812015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen L, Zhang B, Sun S and Feng F:
Methylation of cytochrome p450 2E1 promoter induced by low dosage
of isoniazid. Environ Toxicol Pharmacol. 36:149–151. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang B, Sun S, Shen L, Zu X, Chen Y, Hao
J, Huang X and Feng F: DNA methylation in the rat livers induced by
low dosage isoniazid treatment. Environ Toxicol Pharmacol.
32:486–490. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akai S, Uematsu Y, Tsuneyama K, Oda S and
Yokoi T: Kupffer cell-mediated exacerbation of methimazole-induced
acute liver injury in rats. J Appl Toxicol. 36:702–715. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takai S, Higuchi S, Yano A, Tsuneyama K,
Fukami T, Nakajima M and Yokoi T: Involvement of immune- and
inflammatory-related factors in flucloxacillin-induced liver injury
in mice. J Appl Toxicol. 35:142–151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Park SW, Kang JW and Lee SM: Role of
Kupffer cells in ischemic injury in alcoholic fatty liver. J Surg
Res. 194:91–100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bahari A, Mehrzad J, Mahmoudi M, Bassami
MR and Dehghani H: Cytochrome P450 isoforms are differently
up-regulated in aflatoxin B1-exposed human lymphocytes
and monocytes. Immunopharmacol Immunotoxicol. 36:1–10. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mahmoud AM and Al Dera HS:
18β-Glycyrrhetinic acid exerts protective effects against
cyclophosphamide-induced hepatotoxicity: Potential role of PPARγ
and Nrf2 upregulation. Genes Nutr. 10:412015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pogribny IP and Beland FA: DNA
hypomethylation in the origin and pathogenesis of human diseases.
Cell Mol Life Sci. 66:2249–2261. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ozden S, Kara Turgut N, Sezerman OU,
Durasi İM, Chen T, Demirel G, Alpertunga B, Chipman JK and Mally A:
Assessment of global and gene-specific DNA methylation in rat liver
and kidney in response to non-genotoxic carcinogen exposure.
Toxicol Appl Pharmacol. 289:203–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meng H, Cao Y, Qin J, Song X, Zhang Q, Shi
Y and Cao L: DNA methylation, its mediators and genome integrity.
Int J Biol Sci. 11:604–617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mah WC and Lee CG: DNA methylation:
Potential biomarker in hepatocellular carcinoma. Biomark Res.
2:52014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Villanueva A, Portela A, Sayols S,
Battiston C, Hoshida Y, Méndez-González J, Imbeaud S, Letouzé E,
Hernandez-Gea V, Cornella H, et al: DNA methylation-based prognosis
and epidrivers in hepatocellular carcinoma. Hepatology.
61:1945–1956. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kurzawski M, Dziedziejko V, Post M,
Wójcicki M, Urasińska E, Miętkiewski J and Droździk M: Expression
of genes involved in xenobiotic metabolism and transport in
end-stage liver disease: Up-regulation of ABCC4 and CYP1B1.
Pharmacol Rep. 64:927–939. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bernstein BE, Meissner A and Lander ES:
The mammalian epigenome. Cell. 128:669–681. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang W, Wang C, Fu F and Chen Q: RhoBTB2
gene in breast cancer is silenced by promoter methylation. Int J
Mol Med. 33:722–728. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ball MP, Li JB, Gao Y, Lee JH, LeProust
EM, Park IH, Xie B, Daley GQ and Church GM: Targeted and
genome-scale strategies reveal gene-body methylation signatures in
human cells. Nat Biotechnol. 27:361–368. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Beedanagari SR, Taylor RT, Bui P, Wang F,
Nickerson DW and Hankinson O: Role of epigenetic mechanisms in
differential regulation of the dioxin-inducible human CYP1A1 and
CYP1B1 genes. Mol Pharmacol. 78:608–616. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ingelman-Sundberg M, Zhong XB, Hankinson
O, Beedanagari S, Yu AM, Peng L and Osawa Y: Potential role of
epigenetic mechanisms in the regulation of drug metabolism and
transport. Drug Metab Dispos. 41:1725–1731. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Marques M, Laflamme L and Gaudreau L:
Estrogen receptor α can selectively repress dioxin
receptor-mediated gene expression by targeting DNA methylation.
Nucleic Acids Res. 41:8094–8106. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao K, Brandt I, Goldstone JV and Jönsson
ME: Cytochrome P450 1A, 1B, and 1C mRNA induction patterns in
three-spined stickleback exposed to a transient and a persistent
inducer. Comp Biochem Physiol C Toxicol Pharmacol. 154:42–55. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tokizane T, Shiina H, Igawa M, Enokida H,
Urakami S, Kawakami T, Ogishima T, Okino ST, Li LC, Tanaka Y, et
al: Cytochrome P450 1B1 is overexpressed and regulated by
hypomethylation in prostate cancer. Clin Cancer Res. 11:5793–5801.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao Y, Ma X, Wang J, He X, Hu Y, Zhang P,
Wang R, Li R, Gong M, Luo S and Xiao X: Curcumin protects against
CCl4-induced liver fibrosis in rats by inhibiting HIF-1α through an
ERK-dependent pathway. Molecules. 19:18767–18780. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qian H, Shi J, Fan TT, Lv J, Chen SW, Song
CY, Zheng ZW, Xie WF and Chen YX: Sophocarpine attenuates liver
fibrosis by inhibiting the TLR4 signaling pathway in rats. World J
Gastroenterol. 20:1822–1832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang F, Wang X, Qiu X, Wang J, Fang H,
Wang Z, Sun Y and Xia Z: The protective effect of esculentoside A
on experimental acute liver injury in mice. PLoS One.
9:e1131072014. View Article : Google Scholar : PubMed/NCBI
|