Introduction
High oxygen mechanical ventilation is widely used as
a clinical therapy to treat various diseases (1), including conditions associated with
serious respiratory failure, such as hypoxemia and acute
respiratory distress syndrome (ARDS) (2). However, as suggested in a previous
report (3), exposure to high
levels of oxygen for prolonged periods of time may lead to
inflammation and lung injury (4).
Reactive oxygen species (ROS) are generated under hyperoxic
conditions and result in oxidative damage and inflammatory response
in the injured lung tissue (5).
Inflammatory cells accumulating in the lung also generate ROS
(6). Currently, antioxidants and
antiallergic drugs are used to protect the lung against hyperoxic
lung injury (7,8). However, these drugs often bring side
effects. Therefore, the development of a therapeutic strategy that
alleviates hyperoxia-induced lung injury without side effects is
required.
Interest in the biological activities of marine
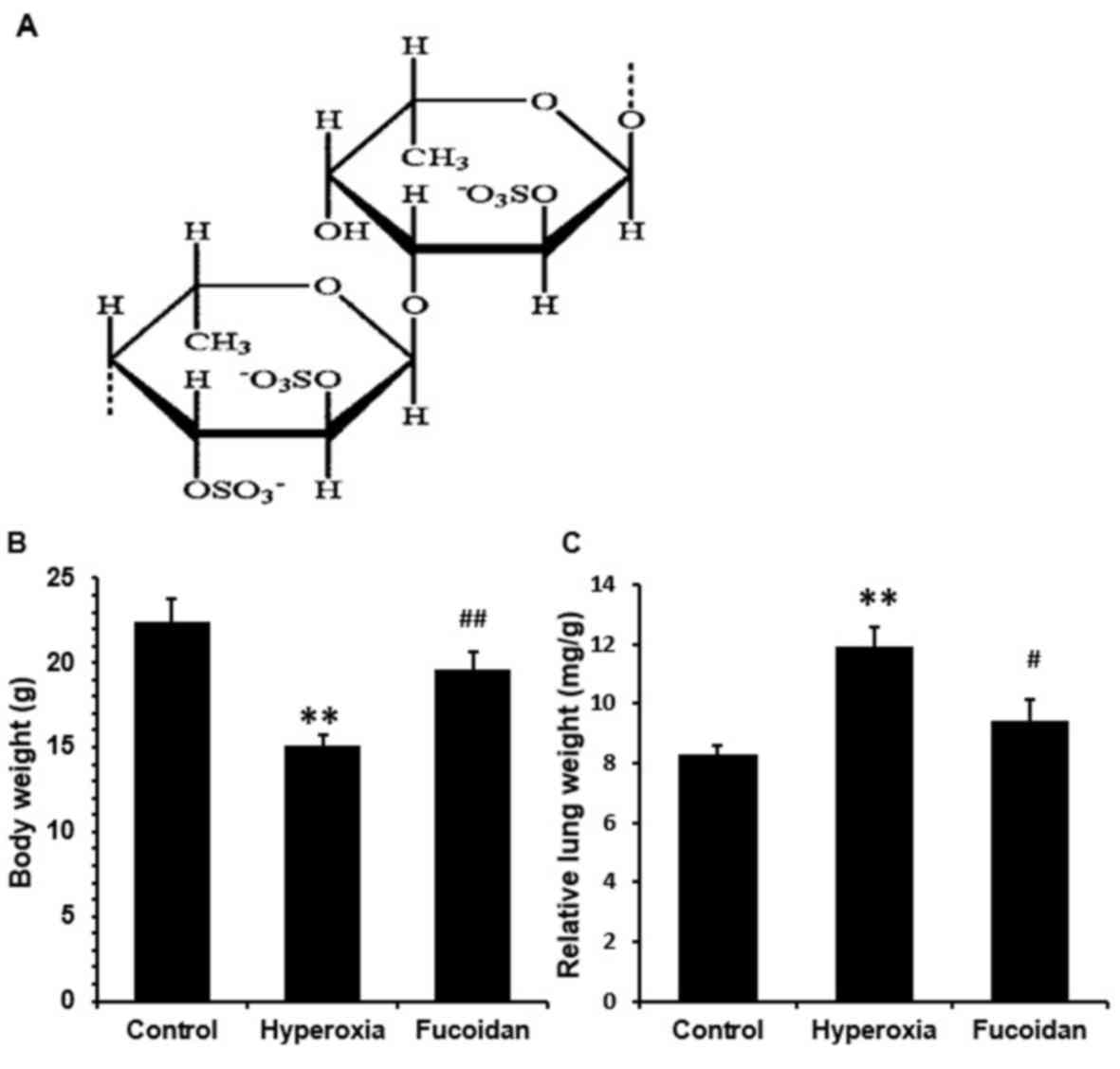
organisms has intensified in the last few years (9). Fucoidan, the chemical structure of
which is presented in Fig. 1A, is
an extract of the seaweed Fucus vesiculosus, which has been
widely investigated and has been reported to serve an important
role in cancer (10) and
inflammation (11). Fucoidan
exhibits antioxidant, anticancer and anti-inflammatory activities
(12), and our previous study
demonstrated its effects against diabetic nephropathy (13). The present study aimed to determine
whether fucoidan may attenuate or prevent hyperoxia-induced lung
injury.
Activation of extracellular signal-regulated kinase
(ERK) 1/2 has been reported to be associated with defense signaling
against injury (14). High oxygen
mechanical ventilation may induce the phosphorylation of ERK1/2,
and the secretion of chemokines and cytokines, such as interleukin
(IL)-1, IL-6 and tumor necrosis factor (TNF)-α (15), finally leading to inflammatory cell
infiltration (16) and lung
injury. The present study indicated that hyperoxia induced
inflammation and morphological alterations in mouse lung tissue.
Conversely, atomization inhalation of fucoidan reduced
hyperoxia-induced inflammation, morphological alterations, the
expression levels of IL-1, IL-6 and TNF-α, and the phosphorylation
of ERK1/2. These findings suggest that fucoidan may attenuate
hyperoxic lung injury via the ERK1/2 signaling pathway, thus
indicating a novel therapeutic strategy for the alleviation of
hyperoxia-induced lung injury.
Materials and methods
Animals
The present study was approved by the Ethics
Committee of Harbin Medical University (Harbin, China). The
experiments were conducted at the Animal Experimental Center of
Harbin Medical University in accordance with the Animal Experiment
Guidelines from Harbin Medical University. Balb/c mice, not limited
to gender (male/female ratio was 1:1), were housed in a controlled
environment, with a temperature of 24±1°C, and a 12-h light/dark
cycle. Briefly, Balb/c mice (age, 6–8 weeks; Institute of
Laboratory Animal Sciences, Shanghai, China) were fed normal chow
and water ad libitum. The mice (n=5-6 per group) were
ventilated with 100% oxygen, with or without atomization inhalation
of fucoidan (100 µg/ml; Kanehide Bio Co., Ltd., Okinawa, Japan),
for 36 h, according to previous studies (17,18).
Untreated mice were used as control. Following exposure to 100%
oxygen, the body weight and relative lung weight of Balb/c mice
were measured before and after sacrifice respectively, and lung
tissue and bronchoalveolar lavage fluid (BALF) samples were
collected for further experimentation.
Preparation of BALF
The lungs were washed with 0.5 ml sterile saline
four times through a 21G flat syringe needle cannulated 0.7 cm into
the trachea. The BALF recovered from each mouse was subsequently
used for quantitative cell counting. Cell quantity and
differentiation were determined using a hemocytometer
(Countess® II Automated Cell Counter; AMQAX1000; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) for >500 cells placed
on cytocentrifuge slides and stained with Wright-Giemsa (19). The total cell number was counted by
the Automated Cell Counter, and the macrophages, neutrophils and
lymphocytes with Giemsa staining were confirmed and counted under a
microscope (Olympus Corporation, Tokyo, Japan) at a magnification
of ×100. A 100 ml aliquot of BALF was collected, after total cell
counting, the remainder was immediately centrifuged at 1,000 × g
for 10 min. Macrophages were isolated from the BALF. The BALF
supernatants were stored at −80°C for cytokine and chemokine
analyses.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the lung tissue by
TRIzol (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and the
relative mRNA expression levels of cluster of differentiation
(CD)68, F4/80, CD64 and CD19 were normalized to GAPDH. The
following primers were used: CD68, forward
5′-CATCAGAGCCCGAGTACAGTCTACC-3′, reverse
5′-AATTCTGCGCCATGAATGTCC-3′; F4/80, forward
5′-GAGATTGTGGAAGCATCCGAGAC-3′, reverse,
5′-GATGACTGTACCCACATGGCTGA-3′; CD64, forward
5′-CTTCTCCTTCTATGTGGGCAGT-3′, reverse 5′-GCTACCTCGCACCAGTATGAT-3′;
CD19, forward 5′-CCACAAAGTCCCAGCTGAAT-3′, reverse,
5′-GGGGTCCCAGATTTCAAAGT-3′; and GAPDH, forward
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse 5′-TGGTGAAGACGCCAGTGGA-3′. A
Transcriptor First Strand cDNA Synthesis kit (Roche Applied
Science, Madison, WI, USA) was used for the reverse transcription.
qPCR was performed using the ABI 7300 Fast Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and the
thermocycling conditions were: Holding stage 95°C for 30 sec, 1
cycle and cycling stage at 95°C for 3 sec and 40 cycles at 60°C for
31 sec. The 30 min electrophoresis with agarose gel with ethidium
bromide (Sigma-Aldrich, Merck KGaA). Every experiment was repeated
3 times. As described in reference (20), the image analysis was performed
using the Image Lab™ software (version 2.1; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). PCR results were analyzed
by 2−ΔΔCq method (21).
Morphometric analysis
As previously described, (22), lung tissues were fixed in formalin
(Sigma-Aldrich; Merck KGaA), cleared in xylene (Sigma-Aldrich;
Merck KGaA) and embedded in paraffin. The paraffin-embedded lung
tissue samples were examined by conventional light microscopic
examination: 5 µm sections were stained with hematoxylin and eosin,
and were assessed in a blinded manner. An automatic microscope
(Provis AX-70) with a camera (Olympus Corporation, Tokyo, Japan)
was used to capture the microscopic images of the lung samples. The
morphometric analysis was performed using ImageJ software version
1.60 (National Institutes of Health, Bethesda, MD, USA) (23).
Enzyme-linked immunosorbent assay
(ELISA)
BALF and homogenized lung tissue were collected and
underwent IL-1, IL-6 and TNF-α assays using respective mouse ELISA
kits (RAB0275, RAB0309 and GERPN2718; Merck KGaA). The ELISA plates
were incubated with 100 µl capture antibody per well at 4°C
overnight. After appropriate washing by the washing buffer from the
kits, 200 µl assay dilution buffer was added per well for blocking
at room temperature for 1 h. Serial dilutions of standards and
samples were added to each well and were incubated at 4°C
overnight. Avidin-horseradish peroxidase (HRP) was added following
incubation with the detection antibody for 30 min at room
temperature, and the samples were further incubated at room
temperature for another 30 min. Subsequently, the
3,3′,5,5′-tetramethylbenzidine substrate was added, and following
incubation for 15 min at room temperature,
NH2SO4 was added to terminate the reaction
and absorbance was measured at 450 nm using an ELISA reader
(MTP-800 microplate reader; Corona Electric Co., Ltd., Tokyo,
Japan).
Western blotting
According to method described by Laemmli (24) for western blotting, electrophoresis
was performed using a vertical slab gel containing 12%
polyacrylamide. Lung tissues were ground and lysed with
radio-immunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Shanghai, China). The concentration of proteins
extracted from lung tissues was determined by Gene Spec III from
Hitachi Genetic Systems (MiraiBio Group of Hitachi Solutions
America, Ltd., South San Francisco, CA, USA) and 20 µg of protein
was loaded per gel lane. Proteins were separated by SDS-PAGE and
were then electrophoretically transferred to a supported
nitrocellulose membrane (Bio-Rad Laboratories, Inc.) using a
Semi-Dry Electroblotter for 90 min at 15 V (Sartorius AG,
Göttingen, Germany) (25). The
membrane was then treated with Block Ace™ (4%) at 22°C for 30 min.
The membrane was then incubated with rabbit immunoglobulin (Ig)G
anti-ERK1/2 (SAB1305560; 1:1,000), anti-p-ERK1/2 (SAB4301578;
1:1,000) and anti-β-actin (SAB2100037; 1:1, Sigma-Aldrich; Merck
KGaA) in phosphate-buffered saline containing 0.03% Tween 20 at
22°C for 1 h. Following washing in the same buffer, the membrane
was incubated with HRP-conjugated goat anti-rabbit IgG (A0545; 20
ng/ml; Sigma-Aldrich; Merck KGaA) at 22°C for 30 min. Following
further washing, an Enhanced Chemiluminescence Plus Western
Blotting Detection system (GE Healthcare Life Sciences, Tokyo,
Japan) was used to visualize the blot. ImageJ version 1.48 u
(National Institutes of Health) was used for the quantification of
western blots.
Statistical analysis
Analyses were performed using SPSS version 19.0 (IBM
Corp., Armonk, NY, USA). Results are presented as the mean ±
standard deviation. Each experiment was repeated at least three
times. Multiple comparisons between groups were performed using
one-way analysis of variance and Dunnett's test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Hyperoxia-induced decreases in body
weight and increases in lung weight are attenuated by fucoidan
Balb/c mice were ventilated with 100% oxygen, with
or without atomization inhalation of fucoidan (100 µg/ml), for 36
h. Hyperoxia significantly reduced mouse body weight (Fig. 1B) and increased relative lung
weight (Fig. 1C; P<0.01).
Conversely, fucoidan significantly increased body weight
(P<0.01) and reduced relative lung weight (P<0.05) compared
with the hyperoxia group.
Hyperoxia-induced inflammatory cell
infiltration into the BALF is attenuated by fucoidan
Following ventilation with 100% oxygen, with or
without atomization inhalation of fucoidan (100 µg/ml), for 36 h,
BALF samples of Balb/c mice were collected. Total cell counts, and
the number of macrophages, neutrophils and lymphocytes, in the BALF
were determined on cytocentrifuge slides using a hemocytometer.
Hyperoxia significantly increased total cell count (Fig. 2A), and the number of macrophages
(Fig. 2B), neutrophils (Fig. 2C) and lymphocytes (Fig. 2D) compared with the control mice
(P<0.01). The hyperoxia-induced increases in the total cell
count, and the number of macrophages, neutrophils and lymphocytes,
in the BALF were significantly reduced in mice receiving fucoidan
(P<0.05).
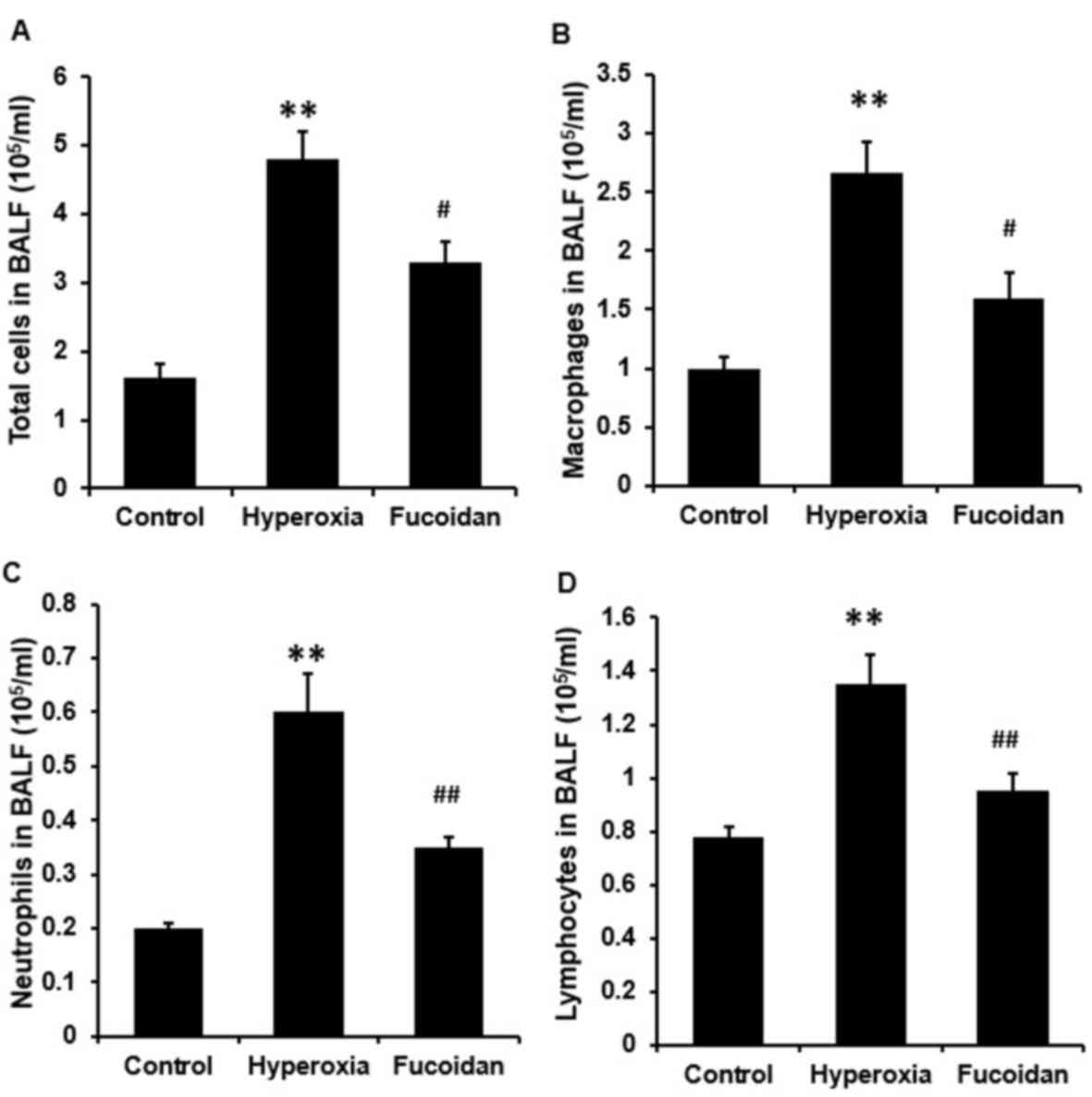 | Figure 2.Hyperoxia-induced inflammatory cell
infiltration into the BALF is attenuated by fucoidan. Following
ventilation with 100% oxygen, with or without atomization
inhalation of fucoidan (100 µg/ml), for 36 h, BALF samples of
Balb/c mice were collected. (A) Total cell counts and the number of
(B) macrophages, (C) neutrophils and (D) lymphocytes in the BALF
were determined on cytocentrifuge slides using a hemocytometer.
Hyperoxia significantly increased total cell count, and the number
of macrophages, neutrophils and lymphocytes, compared with control
mice. The hyperoxia-induced increases in total cell count, and the
number of macrophages, neutrophils and lymphocytes, in the BALF
were significantly reduced in mice receiving fucoidan. Data are
presented as the mean ± standard deviation (n=6 per group).
**P<0.01, vs. control mice; #P<0.05,
##P<0.01, vs. hyperoxia-treated mice. BALF,
bronchoalveolar lavage fluid. |
Hyperoxia-induced inflammatory cell
infiltration into lung tissue is attenuated by fucoidan
Following ventilation with 100% oxygen, with or
without atomization inhalation of fucoidan (100 µg/ml), for 36 h,
lung tissue of Balb/c mice was collected. The mRNA expression
levels of CD68, F4/80 (macrophage markers) (Fig. 3A), CD64 (neutrophil marker) and
CD19 (lymphocyte marker) (Fig. 3B)
were detected in the lung tissue samples. The mRNA expression
levels of CD68, F4/80, CD64 and CD19 were significantly increased
in lung tissue by hyperoxia. Treatment with fucoidan significantly
suppressed hyperoxia-induced mRNA expression (P<0.01).
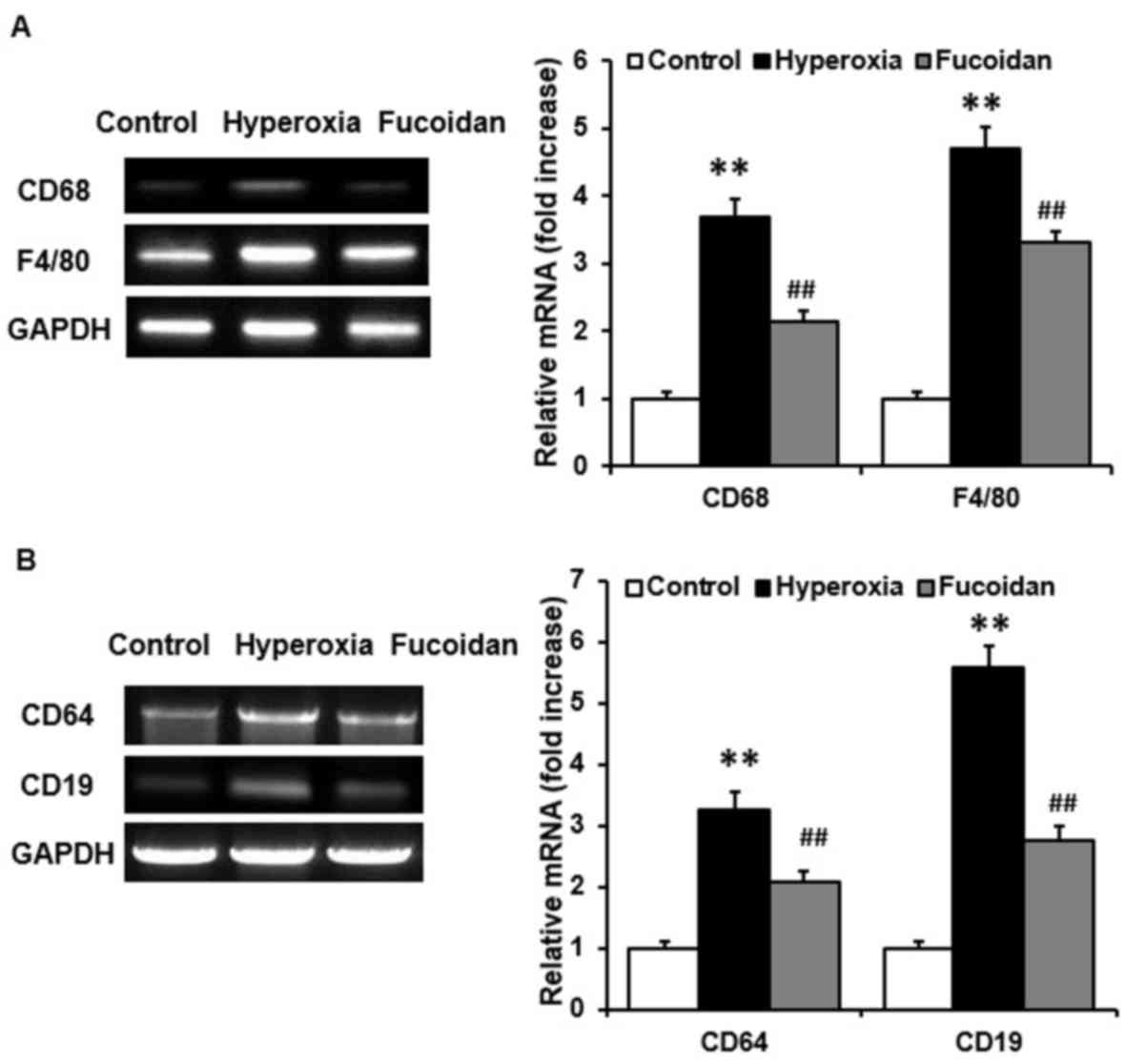 | Figure 3.Hyperoxia-induced inflammatory cell
infiltration in lung tissue is attenuated by fucoidan. Following
ventilation with 100% oxygen, with or without atomization
inhalation of fucoidan (100 µg/ml), for 36 h, lung tissue was
collected from Balb/c mice. The mRNA expression levels of (A) CD68,
F4/80 (macrophage markers), (B) CD64 (neutrophil marker) and CD19
(lymphocyte marker) were detected in the lung tissue samples. The
mRNA expression levels of CD68, F4/80, CD64 and CD19 were
significantly increased in lung tissue by hyperoxia. Treatment with
fucoidan significantly suppressed hyperoxia-induced mRNA
expression. Data are presented as the mean ± standard deviation
(n=6 per group). **P<0.01, vs. control mice;
##P<0.01, vs. hyperoxia-treated mice. CD, cluster of
differentiation. |
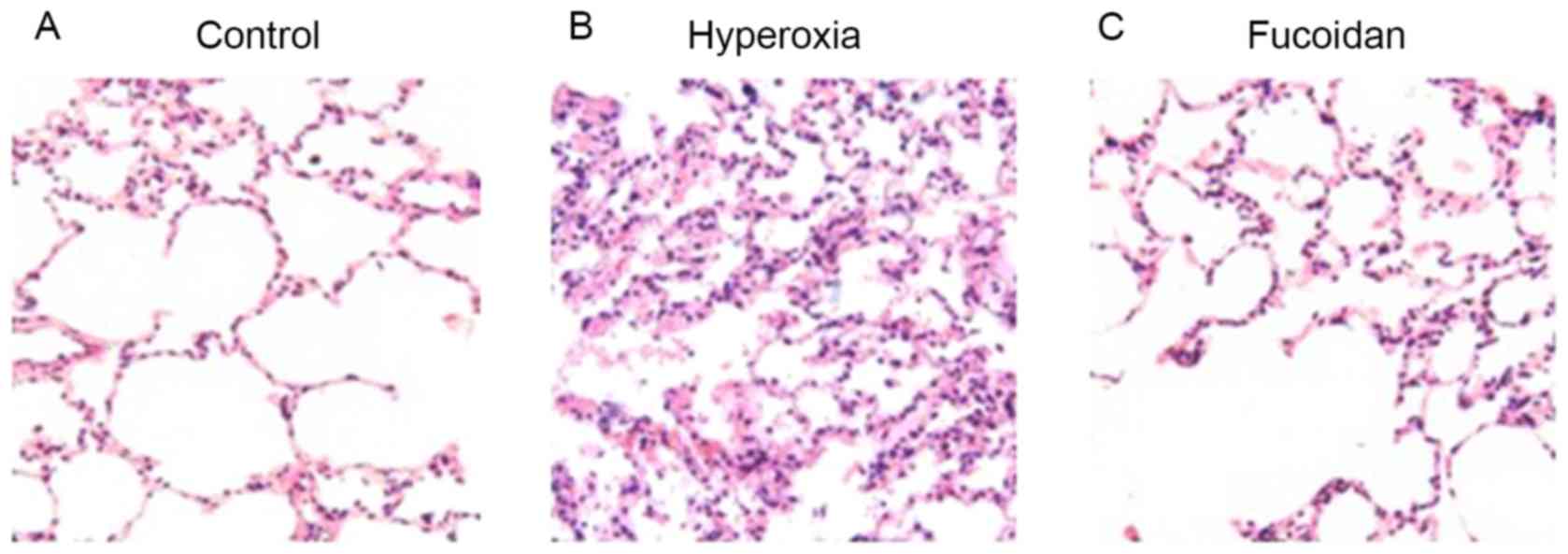
Hyperoxia-induced lung morphological
alterations are attenuated by fucoidan
Balb/c mice were ventilated with 100% oxygen, with
or without atomization inhalation of fucoidan (100 µg/ml), for 36
h. Lung morphological alterations were detected. Hyperoxia
increased the thickness of the alveolar walls and induced
inflammatory cell infiltration into the lung tissue. Conversely,
the hyperoxia-induced morphological alterations were attenuated by
fucoidan treatment (Fig. 4;
magnification, ×200).
Hyperoxia-induced increases in the
expression of inflammatory factors in the BALF and lung tissue are
attenuated by fucoidan
Balb/c mice were ventilated with 100% oxygen, with
or without atomization inhalation of fucoidan (100 µg/ml), for 36
h. BALF and lung tissue samples were then harvested. The expression
levels of IL-1, IL-6 and TNF-α in the BALF (Fig. 5A) and lung tissue (Fig. 5B) were significantly increased by
hyperoxia (P<0.01). Hyperoxia-induced increases in the
expression levels of IL-1, IL-6 and TNF-α in the BALF and lung
tissue were significantly reduced following treatment with fucoidan
(P<0.05).
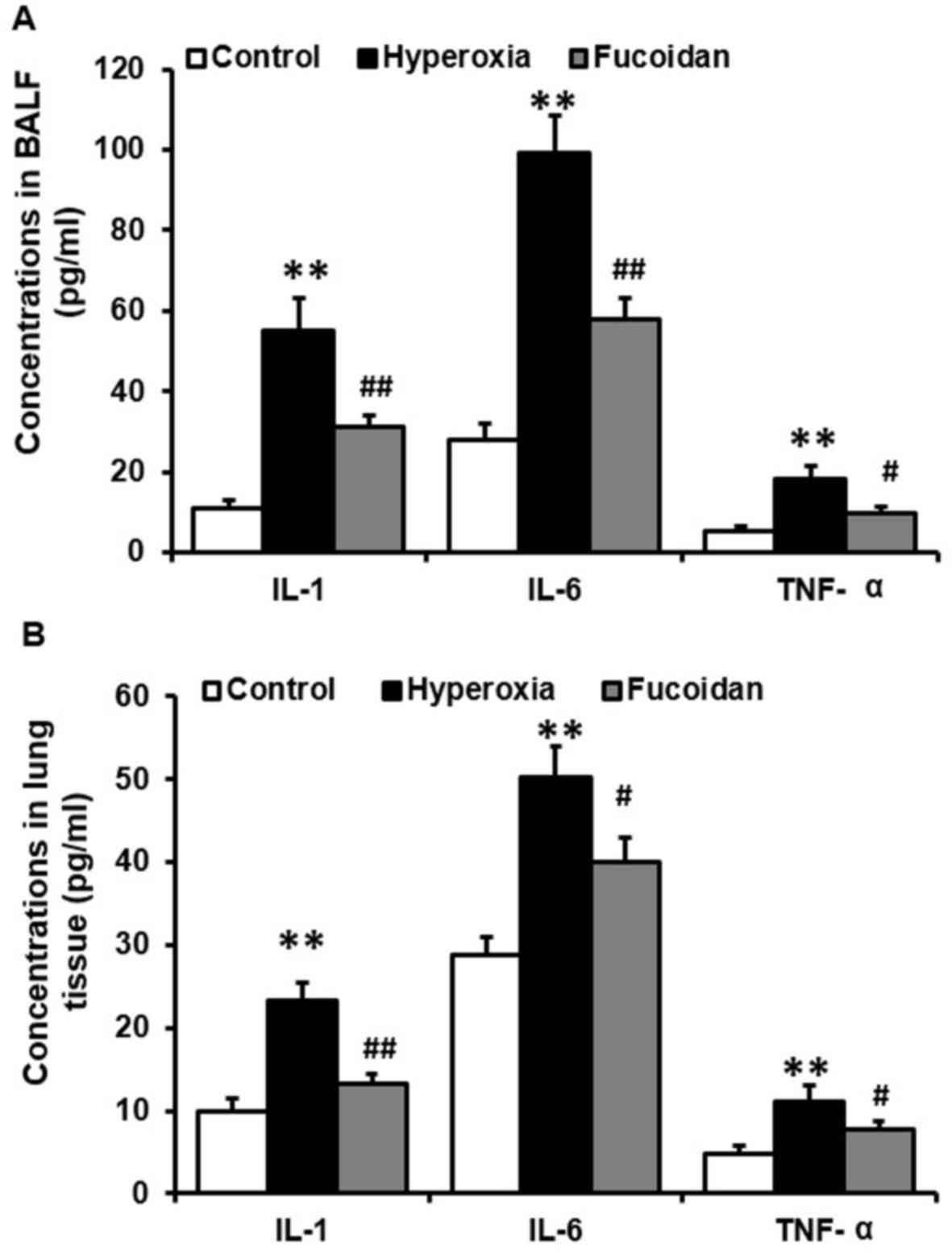 | Figure 5.Hyperoxia-induced increases in the
expression levels of inflammatory factors in the (A) BALF and (B)
lung tissue are attenuated by fucoidan. Balb/c mice were ventilated
with 100% oxygen, with or without atomization inhalation of
fucoidan (100 µg/ml), for 36 h. BALF and lung tissue samples were
then harvested. The expression levels of IL-1, IL-6 and TNF-α in
the BALF and lung tissue samples were significantly increased by
hyperoxia, whereas the expression levels of IL-1, IL-6 and TNF-α
were significantly reduced in the BALF and lung tissue following
treatment with Fucoidan. Data are presented as the mean ± standard
deviation (n=6 per group). **P<0.01 vs. control mice;
#P<0.05, ##P<0.01, vs.
hyperoxia-treated mice. BALF, bronchoalveolar lavage fluid; IL,
interleukin; TNF-α, tumor necrosis factor-α. |
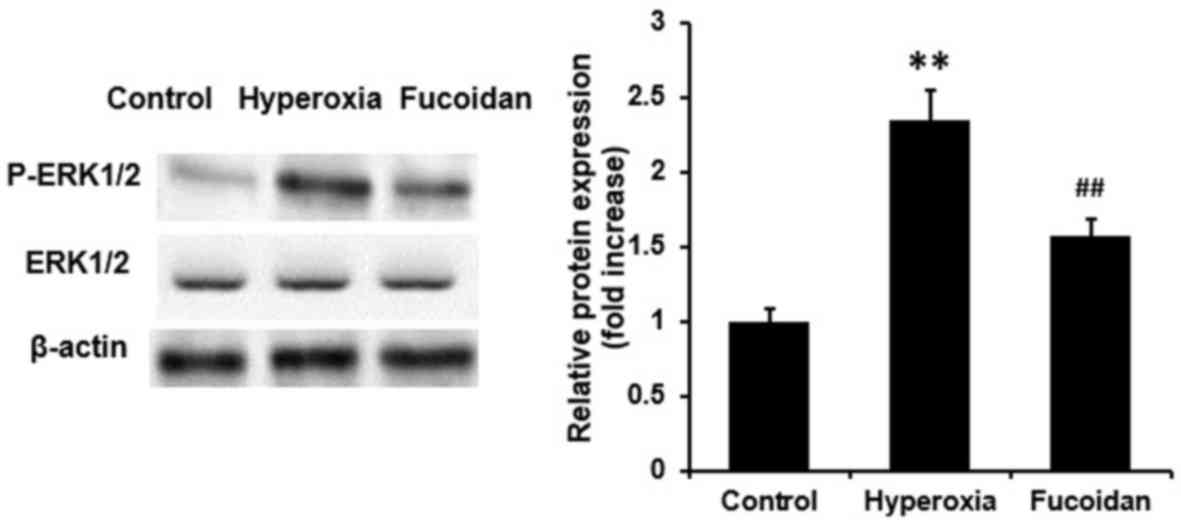
Hyperoxia-induced phosphorylation of
ERK1/2 is attenuated by fucoidan
Balb/c mice were ventilated with 100% oxygen, with
or without atomization inhalation of fucoidan (100 µg/ml), for 36
h. Lung tissue samples were then harvested. Hyperoxia significantly
increased phosphorylation of ERK1/2 in lung tissue (Fig. 6). However, the hyperoxia-induced
phosphorylation of ERK1/2 in the lung tissue was significantly
reduced following treatment with fucoidan (P<0.01).
Discussion
In recent years, high oxygen mechanical ventilation
has been widely used as a clinical therapy to treat various
diseases (1), including conditions
associated with serious respiratory failure, such as hypoxemia and
ARDS (2). However, as suggested in
a previous study (3), and
clinically (26), exposure to high
levels of oxygen for prolonged periods of time may lead to
inflammation and lung injury (4).
Therefore, a therapeutic strategy that alleviates hyperoxia-induced
lung injury is required.
Interest in the biological activities of marine
organisms has intensified (9).
Fucoidan, which is an extract of the seaweed Fucus
vesiculosus, has been widely investigated and has been reported
to serve an important role in cancer (10) and inflammation (11). Fucoidan is able to suppress various
inflammatory cytokines, including IL-1β, TNF-α, interferon-γ and
cyclooxygenase-2 (11). Fucoidan
is an antioxidant, anticancer and anti-inflammatory agent (12), which also exerts effects against
diabetic nephropathy (13). The
present study investigated the protective effects of fucoidan
against hyperoxia-induced lung injury. Prior to the present study,
we performed a dose-dependent study using 1, 10, 100 and 1,000
µg/ml fucoidan; significant alterations occurred when ≥100 µg/ml
fucoidan was used; therefore, 100 µg/ml was used as the standard
concentration in the present study.
The present study demonstrated that hyperoxia
significantly increased total cell count, and the number of
macrophages, neutrophils and lymphocytes in the BALF. In addition,
hyperoxia significantly increased the mRNA expression levels of
CD68 and F4/80 (macrophage markers), CD64 (neutrophil marker) and
CD19 (lymphocyte marker), and induced increases in alveolar wall
thickness and inflammatory cell infiltration into the lung tissue.
These results are consistent with those of a previous study, which
suggested that hyperoxia may recruit inflammatory cells, such as
macrophages, lymphocytes and neutrophils (27). Fucoidan significantly reduced
hyperoxia-induced inflammatory cell infiltration into the BALF and
lung tissue, and attenuated the morphological alterations.
Furthermore, normal mice were treated with fucoidan only; no
significant differences were detected between the control and
fucoidan-treated normal mice (data not shown), strongly indicating
that fucoidan did not exhibit cytotoxicity.
High oxygen mechanical ventilation may lead to lung
injury, and secretion of chemokines and cytokines, such as IL-1
(28), IL-6 (29) and TNF-α (30), thus resulting in inflammatory cell
infiltration (15,16). It has previously been reported that
IL-1 and IL-6 are associated with anti- and proinflammatory
responses, through the regulation of leukocyte apoptosis and
function, and may beneficially regulate neutrophil migration and
adhesion (31). Elevated levels of
IL-1, IL-6 and TNF-α have been demonstrated as biological markers
in numerous models of lung injury (32,33).
To improve understanding regarding hyperoxia-induced lung injury,
the expression levels of IL-1, IL-6 and TNF-α in the BALF and lung
tissue were examined in the present study. Hyperoxia significantly
increased the expression levels of IL-1, IL-6 and TNF-α in the BALF
and lung tissue, whereas hyperoxia-induced IL-1, IL-6 and TNF-α
expression was significantly attenuated in the BALF and lung tissue
following treatment with fucoidan.
It has also been demonstrated that activation of the
ERK1/2 pathway may be involved in defense signaling against
oxidative injury (14). The ERK1/2
pathway is a signaling cascade that is involved in protection from
damage, the activation of which is generally hypothesized to
mediate cell survival (34).
Secretion of chemokines and cytokines, such as IL-1, IL-6 and
TNF-α, are associated with the phosphorylation of ERK1/2 (15). To better understand the underlying
mechanism, the present study examined the phosphorylation of
ERK1/2. Hyperoxia significantly induced phosphorylation of ERK1/2
in lung tissue; however, hyperoxia-induced phosphorylation of
ERK1/2 was significantly reduced following treatment with
fucoidan.
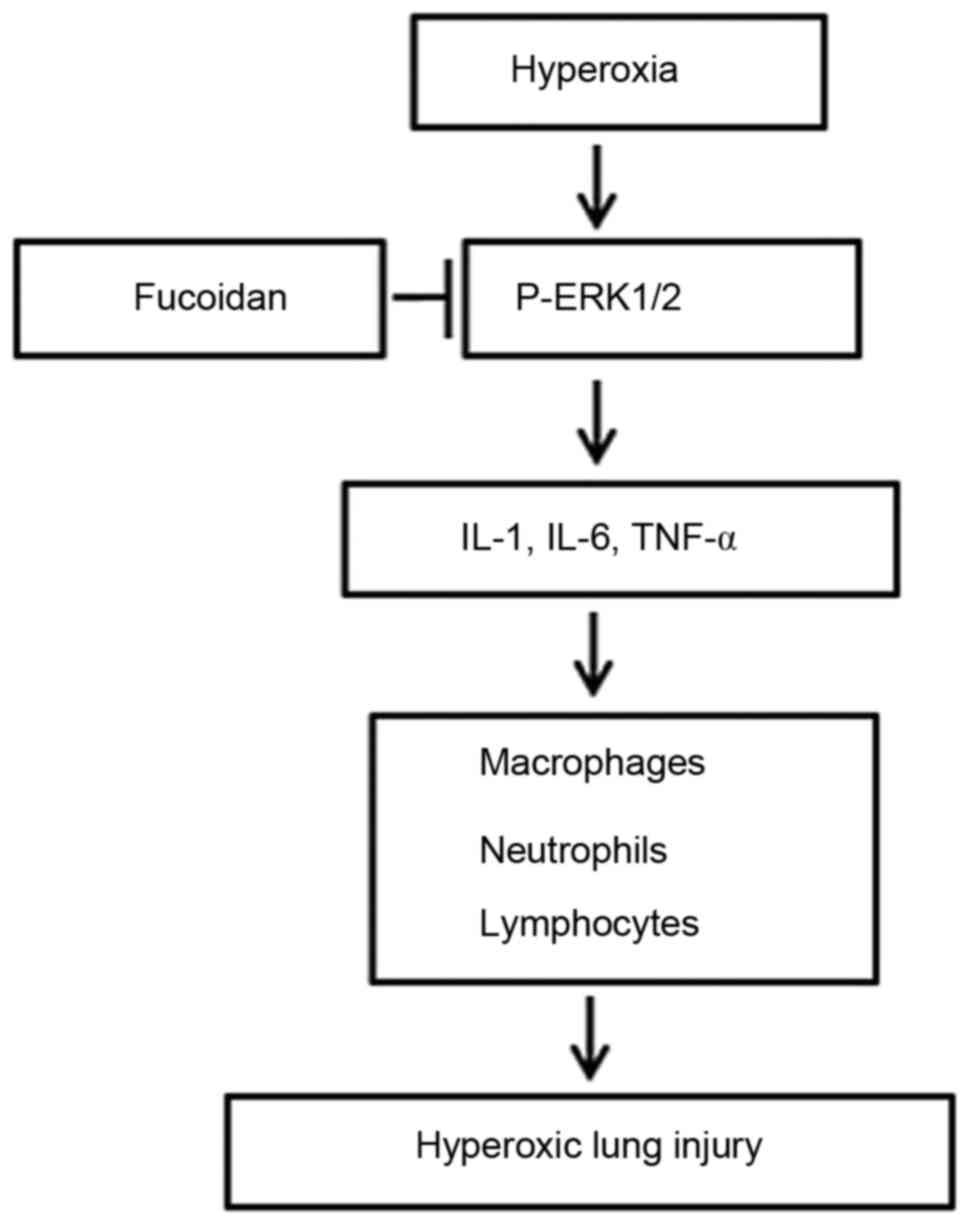
The protective mechanism of fucoidan in
hyperoxia-induced lung injury is presented in Fig. 7. Briefly, hyperoxia induced
phosphorylation of ERK1/2, increased the expression levels of IL-1,
IL-6 and TNF-α, and induced lung morphological alterations and
inflammation. However, these effects were attenuated following
treatment with fucoidan.
In conclusion, to the best of our knowledge, the
present study is the first to demonstrate the crucial role of
fucoidan against hyperoxia-induced lung injury via the ERK1/2
signaling pathway. These results suggested a novel therapeutic
strategy for the alleviation of hyperoxia-induced lung injury.
References
|
1
|
Xu Y, Tian Z and Xie P: Targeting
complement anaphylatoxin C5a receptor in hyperoxic lung injury in
mice. Mol Med Rep. 10:1786–1792. 2014. View Article : Google Scholar :
|
|
2
|
Vitacca M, Bianchi L, Bazza A and Clini
EM: Advanced COPD patients under home mechanical ventilation and/or
long term oxygen therapy: Italian healthcare costs. Monaldi Arch
Chest Dis. 75:207–214. 2011.
|
|
3
|
Panikkanvalappil SR, James M, Hira SM,
Mobley J, Jilling T, Ambalavanan N and El-Sayed MA: Hyperoxia
induces intracellular acidification in neonatal mouse lung
fibroblasts: Real-time investigation using plasmonically enhanced
Raman spectroscopy. J Am Chem Soc. 138:3779–3788. 2016. View Article : Google Scholar
|
|
4
|
Vosdoganes P, Lim R, Koulaeva E, Chan ST,
Acharya R, Moss TJ and Wallace EM: Human amnion epithelial cells
modulate hyperoxia-induced neonatal lung injury in mice.
Cytotherapy. 15:1021–1029. 2013. View Article : Google Scholar
|
|
5
|
George CL, Fantuzzi G, Bursten S, Leer L
and Abraham E: Effects of lisofylline on hyperoxia-induced lung
injury. Am J Physiol. 276:L776–L785. 1999.
|
|
6
|
Kallet RH and Matthay MA: Hyperoxic acute
lung injury. Respir Care. 58:123–141. 2013. View Article : Google Scholar :
|
|
7
|
Johnson-Varghese L, Brodsky N and Bhandari
V: Effect of antioxidants on apoptosis and cytokine release in
fetal rat Type II pneumocytes exposed to hyperoxia and nitric
oxide. Cytokine. 28:10–16. 2004. View Article : Google Scholar
|
|
8
|
Fukuhara K, Nakashima T, Abe M, Masuda T,
Hamada H, Iwamoto H, Fujitaka K, Kohno N and Hattori N: Suplatast
tosilate protects the lung against hyperoxic lung injury by
scavenging hydroxyl radicals. Free Radic Biol Med. 106:1–9. 2017.
View Article : Google Scholar
|
|
9
|
Xie P, Fujii I, Zhao J, Shinohara M and
Matsukura M: A novel polysaccharide compound derived from algae
extracts protects retinal pigment epithelial cells from high
glucose-induced oxidative damage in vitro. Biol Pharm Bull.
35:1447–1453. 2012. View Article : Google Scholar
|
|
10
|
Kasai A, Arafuka S, Koshiba N, Takahashi D
and Toshima K: Systematic synthesis of low-molecular weight
fucoidan derivatives and their effect on cancer cells. Org Biomol
Chem. 13:10556–10568. 2015. View Article : Google Scholar
|
|
11
|
Fukahori S, Yano H, Akiba J, Ogasawara S,
Momosaki S, Sanada S, Kuratomi K, Ishizaki Y, Moriya F, Yagi M and
Kojiro M: Fucoidan, a major component of brown seaweed, prohibits
the growth of human cancer cell lines in vitro. Mol Med Rep.
1:537–542. 2008.
|
|
12
|
Li X, Zhao H, Wang Q, Liang H and Jiang X:
Fucoidan protects ARPE-19 cells from oxidative stress via
normalization of reactive oxygen species generation through the
Ca2+-dependent ERK signaling pathway. Mol Med Rep.
11:3746–3752. 2015. View Article : Google Scholar
|
|
13
|
Wang Y, Nie M, Lu Y, Wang R, Li J, Yang B,
Xia M, Zhang H and Li X: Fucoidan exerts protective effects against
diabetic nephropathy related to spontaneous diabetes through the
NF-κB signaling pathway in vivo and in vitro. Int J Mol Med.
35:1067–1073. 2015. View Article : Google Scholar
|
|
14
|
Glotin AL, Calipel A, Brossas JY, Faussat
AM, Tréton J and Mascarelli F: Sustained versus transient ERK1/2
signaling underlies the anti- and proapoptotic effects of oxidative
stress in human RPE cells. Invest Ophthalmol Vis Sci. 47:4614–4623.
2006. View Article : Google Scholar
|
|
15
|
Pushparaj PN, Tay HK, Wang CC, Hong W and
Melendez AJ: VAMP8 is essential in anaphylatoxin-induced
degranulation, TNF-alpha secretion, peritonitis, and systemic
inflammation. J Immunol. 183:1413–1418. 2009. View Article : Google Scholar
|
|
16
|
Davies J, Karmouty-Quintana H, Le TT, Chen
NY, Weng T, Luo F, Molina J, Moorthy B and Blackburn MR: Adenosine
promotes vascular barrier function in hyperoxic lung injury.
Physiol Rep. 2:pii: e12155. 2014. View Article : Google Scholar :
|
|
17
|
Mikawa K, Nishina K, Maekawa N and Obara
H: Attenuation of hyperoxic lung injury in rabbits with superoxide
dismutase: Effects on inflammatory mediators. Acta Anaesthesiol
Scand. 39:317–322. 1995. View Article : Google Scholar
|
|
18
|
Takao Y, Mikawa K, Nishina K, Maekawa N
and Obara H: Lidocaine attenuates hyperoxic lung injury in rabbits.
Acta Anaesthesiol Scand. 40:318–325. 1996. View Article : Google Scholar
|
|
19
|
Dolinay T, Wu W, Kaminski N, Ifedigbo E,
Kaynar AM, Szilasi M, Watkins SC, Ryter SW, Hoetzel A and Choi AM:
Mitogen-activated protein kinases regulate susceptibility to
ventilator-induced lung injury. PLoS One. 3:e16012008. View Article : Google Scholar :
|
|
20
|
Camacho Villarreal JL, Torres Mendoza E,
Cadena C, Prieto J, Varela Prieto LL and DA Torregroza Villanueva:
The use of factorial design, image analysis, and an efficiency
calculation for multiplex PCR optimization. J Clin Lab Anal.
27:249–252. 2013. View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Jiang X, Ma Y, Yu J, Li H and Xie F:
Protective effect of C4a against hyperoxic lung injury via a
macrophage-dependent but not a neutrophil/lymphocyte-dependent
signaling pathway. Mol Med Rep. 13:1250–1256. 2016. View Article : Google Scholar
|
|
23
|
Runzi M, Raptopoulos V, Saluja AK, Kaiser
AM, Nishino H, Gerdes D and Steer ML: Evaluation of necrotizing
pancreatitis in the opossum by dynamic contrast-enhanced computed
tomography: Correlation between radiographic and morphologic
changes. J Am Coll Surg. 180:673–682. 1995.
|
|
24
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar
|
|
25
|
Tsuruta T, Yamamoto T, Matsubara S,
Nagasawa S, Tanase S, Tanaka J, Takagi K and Kambara T: Novel
function of C4a anaphylatoxin. Release from monocytes of protein
which inhibits monocyte chemotaxis. Am J Pathol. 142:1848–1857.
1993.
|
|
26
|
Matsubara S, Yamamoto T, Tsuruta T, Takagi
K and Kambara T: Complement C4-derived monocyte-directed chemotaxis
inhibitory factor. A molecular mechanism to cause polymorphonuclear
leukocyte-predominant infiltration in rheumatoid arthritis synovial
cavities. Am J Pathol. 138:1279–1291. 1991.
|
|
27
|
Janssen WJ, Barthel L, Muldrow A,
Oberley-Deegan RE, Kearns MT, Jakubzick C and Henson PM: Fas
determines differential fates of resident and recruited macrophages
during resolution of acute lung injury. Am J Respir Crit Care Med.
184:547–560. 2011. View Article : Google Scholar :
|
|
28
|
Zhao Y, Xu H, Yu W and Xie BD: Complement
anaphylatoxin C4a inhibits C5a-induced neointima formation
following arterial injury. Mol Med Rep. 10:45–52. 2014. View Article : Google Scholar :
|
|
29
|
Jones SA: Directing transition from innate
to acquired immunity: Defining a role for IL-6. J Immunol.
175:3463–3468. 2005. View Article : Google Scholar
|
|
30
|
Bao JP, Jiang LF, Li J, Chen WP, Hu PF and
Wu LD: Visceral adipose tissue-derived serine protease inhibitor
inhibits interleukin-1β-induced catabolic and inflammatory
responses in murine chondrocytes. Mol Med Rep. 10:2191–2197. 2014.
View Article : Google Scholar
|
|
31
|
Wolters PJ, Wray C, Sutherland RE, Kim SS,
Koff J, Mao Y and Frank JA: Neutrophil-derived IL-6 limits alveolar
barrier disruption in experimental ventilator-induced lung injury.
J Immunol. 182:8056–8062. 2009. View Article : Google Scholar :
|
|
32
|
Frank JA, Parsons PE and Matthay MA:
Pathogenetic significance of biological markers of
ventilator-associated lung injury in experimental and clinical
studies. Chest. 130:1906–1914. 2006. View Article : Google Scholar :
|
|
33
|
Piguet PF, Collart MA, Grau GE, Sappino AP
and Vassalli P: Requirement of tumour necrosis factor for
development of silica-induced pulmonary fibrosis. Nature.
344:245–247. 1990. View
Article : Google Scholar
|
|
34
|
Guillonneau X, Régnier-Ricard F, Dupuis C,
Courtois Y and Mascarelli F: Paracrine effects of phosphorylated
and excreted FGF1 by retinal pigmented epithelial cells. Growth
Factors. 15:95–112. 1998. View Article : Google Scholar
|