Introduction
The incidence and mortality of diabetes have been
increasing rapidly in recent decades worldwide (1). The mortality of diabetes is
predominantly due to its severe cardiovascular complications
(2). Diabetic cardiomyopathy
(DbCM) is one of the major clinical manifestations of diabetic
cardiovascular complications, which is characterized by cardiac
dysfunction and arrhythmia (3). As
one of the most noteworthy pathological features of DbCM, cardiac
fibrosis leads to cardiac remodeling, cardiac dilatation and
congestive heart failure (4).
The essential pathological characteristic of cardiac
fibrosis is an excessively produced and accumulated extracellular
matrix (ECM). By synthesizing ECM, cardiac fibroblasts (CFs) serve
a central role in inducing and developing cardiac fibrosis
(5). The transforming growth
factor-β1 (TGF-β1) pathway induces organ fibrosis by mediating its
downstream effectors, which are a family of proteins termed Smads.
Activated Smads form complexes and mediate nuclear translocation,
which leads to initiation of the transcription of genes encoding
the ECM (6). It has also been
reported that the proliferation of the ECM, and ECM synthesis, were
increased in human CFs incubated under high-glucose conditions
(7). However, the exact molecular
mechanism has yet to be fully elucidated. Inhibition of CFs may be
an effective strategy for the treatment of cardiac fibrosis of
DbCM.
Matrine, the active molecule of Sophora
alopecuroides L. (formula:
C15H24N2O), has been gathering
interest in the research community due to its potent and various
biological activities. Results from a previous study published by
the present authors indicated that matrine improved the left
ventricular (LV) function of animals in an experimental DbCM model
(8). The anti-fibrotic activity of
matrine on organ fibrosis via inhibition of the TGF-β/Smad pathway
has also been suggested by another previously published study
(9). In the present study, the
anti-fibrotic effect of matrine on cardiac fibrosis was
investigated with an experimental DbCM model. In the in vivo
part of the study, the effects of matrine on cardiac function and
compliance were studied. In the in vitro part, the possible
molecular mechanisms involved in the anti-fibrotic activity of
matrine were investigated. The results of the present study should
add further to our in-depth understanding of the pathological
mechanism of cardiac fibrosis associated with DbCM. Furthermore,
the results from this study should provide a theoretical foundation
for the application of matrine-based compounds in DbCM
treatment.
Materials and methods
Animal experimental protocol
An experimental animal model of DbCM was induced as
described previously in 7-week oldmale and female (1:1)
Sprague-Dawley rats (weight, 180–220 g) provided by Animal
Experimental Center of Xi'an Jiaotong University (10). The rats were raised in independent
cages in a 12-h light/dark cycle, 25±2°C and 50% humidity. Animals
had access to standard chow and clean water ad libitum.
Intraperitoneal injection of streptozotocin (STZ; 65 mg/kg body
weight) was employed to induce diabetes in rats. Prior to the
induction of diabetes, rats were administered with matrine
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) orally at a dose of
300 mg/body weight per day for 10 days. Further details of the
treatments, and the treatment groups, are provided in Table I. In the present study, all animal
experimental procedures were approved by the Experimental Animal
Use Ethics Committee of Xi'an Jiaotong University, and were
performed in accordance with guidelines for the Care and Use of
Laboratory Animals issued by the Chinese Council on Animal
Research.
 | Table I.Treatment of the rats in the different
groups. |
Table I.
Treatment of the rats in the different
groups.
|
| Treatment 1 | Treatment 2 |
|---|
|
|
|
|
|---|
| Group | Reagent | Description | Reagent | Description |
|---|
| Ctrl | Physiological
saline | 10 days prior to
injection; oral administration; equal volume to matrine | Sodium citrate
buffer (pH 4.5) | Equal volume to STZ
injection; single intraperitoneal injection |
| DbCM | Physiological
saline | 10 days before
injection; oral administration; equal volume to matrine | STZ (dissolved in
sodium citrate buffer, pH 4.5) | 65 mg/kg body
weight; single intraperitoneal injection |
| Mat | Matrine | 10 days before
injection; oral administration; 200 mg/kg/day for 10 consecutive
days | Sodium citrate
buffer, pH 4.5 | Equal volume to STZ
injection; single intraperitoneal injection |
| DbCM+Mat | Matrine | 10 days prior to
injection; oral administration; 200 mg/kg/day for 10 consecutive
days | STZ (dissolved in
sodium citrate buffer with pH 4.5) | 65 mg/kg body
weight; single intraperitoneal injection |
Invasive hemodynamic evaluation of
cardiac function
An invasive hemodynamic method described in our
previous study (11) was employed
to evaluate the cardiac functions. Briefly, following
anesthetization with chloral hydrate (10% v/v, 0.85 mg/kg body
weight), a Mikro-Tip catheter transducer (Millar Instruments,
Houston, TX, USA) was intubated into the left ventricle via the
right carotid artery. Signals captured by the transducer were
inputted into the PowerLab 4/25 Biological Information Analysis
system (ADInstruments, Dunedin, New Zealand), from which the
parameters, including LV end-diastolic pressure (LVEDP), LV
systolic pressure (LVSP), positive maximal rate of LV increased
pressure (+LVdP/dtmax) and negative maximal rate of LV
increased pressure (-LVdP/dtmax), were outputted.
Cardiac compliance assessments
By plotting a pressure-volume (P-V) curve from the
measurements, cardiac compliance was evaluated in isolated perfused
hearts. K-H buffering solution (95% O2/5% CO2
at 37°C) was used to perfuse the isolated Langendorff hearts.
Through left atrial appendage and mitral valves, a non-compliant
balloon filled with K-H solution was inserted into the left
ventricle. The non-compliant balloon was connected to a pressure
transducer (Millar Instruments), and the measurements were recorded
using the PowerLab 4/25 Biological Information Analysis system
(ADInstruments). LVDP was recorded when the volume of the balloon
increased from 0.06 to 0.42 ml. Based on these measurements, the
P-V curve was plotted.
Cardiac collagen staining
Sirius red staining was used to visualize collagens
in cardiac tissue. Following harvesting and trimming, cardiac
tissues were fixed in neutral buffered formalin (10%, v/v) at room
temperature for 10 h. Subsequently, the cardiac tissue was embedded
in paraffin and sectioned into 4 µm-thick slices. A Picro Sirius
Red stain kit (cat. no. ab150681; Abcam, Cambridge, UK) was used to
treat the sections, according to the protocol provided by the
manufacturer. Images were captured using a phase-contrast
microscope. The addition of Sirius Red stain specifically enabled
the positive staining of collagens.
Isolation of primary CFs
Primary CFs were isolated from the left ventricles
of neonatal SD rats following the protocol described in a previous
study (12). After having been
harvested from 2-day-old SD rats, the hearts were minced and
digested using type II collagenase (120 U/ml; Beijing Solarbio
Science and Technology Co., Ltd., Beijing, China). The dissociated
cells were collected by centrifugation (200 × g for 8 min at room
temperature) and further cultured in MEM medium (Hyclone™)
supplemented with fetal bovine serum (FBS; 10%, Hyclone™) (both
from GE Healthcare Life Sciences, Shanghai, China) and
penicillin-streptomycin antibiotic mix (Sigma-Aldrich; Merck KGaA).
Cells were incubated in a humidified incubator under conditions of
95% fresh air and 5% CO2 at 37°C. The adherent cells
were cultured to reach over 80% confluence. In excess of 95% of the
collected cells were vimentin-positive/α-actin-negative. Cells from
passages 2–3 were used. Table II
shows the details of grouping, and treatments of the isolated
cells.
| Table II.Treatment of CFs in different
groups. |
Table II.
Treatment of CFs in different
groups.
|
| Treatment 1 | Treatment 2 |
|---|
|
|
|
|
|---|
| Group | Reagent | Description | Reagent | Description |
|---|
| Ctrl | Vehicle
solution | Incubation for 48
h | Normal glucose
medium | Glucose
concentration at 5.5 mmol/l |
| HG | Vehicle
solution | Incubation for 48
h | HG medium | Glucose
concentration at 25 mmol/l |
| Mat | Matrine solution
(0.25, 0.5, 1.0, 1.5, 2.0 and 2.5 mmol/l) | Incubation for 48
h | Normal glucose
medium | Glucose
concentration at 5.5 mmol/l |
| HG+Mat | Matrine solution
(0.25, 0.5, 1.0, 1.5, 2.0 and 2.5 mmol/l) | Incubation for 48
h | HG medium | Glucose
concentration at 25 mmol/l |
MTT assay
MTT assay was used to assess the cell viability of
isolated CFs. After seeding into a 96-well plate, cultured CFs were
subsequently incubated at 37°C with matrine at serial
concentrations of 0.25, 0.5, 1.0, 1.5, 2.0 and 2.5 mmol/l for 48 h.
MTT (5 mg/ml) was then added into each well, and the cells were
incubated for a further 4 h. Dimethyl sulfoxide (DMSO) was added to
dissolve the resultant formazan crystals. Measurements of
absorbance at 540 nm (A540) were recorded using a plate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Cell
viability was defined according to the formula: (A540 of
matrine – treated cells/A540 of control cells) ×
100%.
Western blotting
The RIPA lysis buffer system (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) with
phenylmethylsulfonylfluoride (PMSF; Santa Cruz Biotechnology, Inc.)
was used to obtain an homogenate of harvested cardiac tissue and
cultured CFs, as per the manufacturer's instructions. Following
centrifugation (12,500 × g) at 4°C for 10 min, the supernatants
were collected. The total protein concentration was detected using
a bicinchoninic acid (BCA) protein assay kit (Santa Cruz
Biotechnology, Inc.). Proteins were subjected to 8% SDS-PAGE, and
subsequently separated by vertical electrophoresis. The samples
were then transferred to polyvinylidene fluoride (PVDF) membranes
(EMD Millipore, Billerica, MA, USA) electrically at 10–12 V for 50
min. Specific primary antibodies against TGF-β1 (cat. no. SC7892,
1:1,000; Santa Cruz Biotechnology, Inc.), Smad2 (cat. no. ab40855,
1:2,000), Smad3 (cat. no. ab28379, 1:2,000) (both from Abcam),
Smad7 (cat. no. PA1-41506, 1:2,000; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), phospho-Smad2 (cat. no. ab53100, 1:2,000),
phospho-Smad3 (cat. no. ab52903, 1:2,000) (both from Abcam),
collagen I (cat. no. 5D8-G9, 1:2,000; Thermo Fisher Scientific,
Inc.) and GAPDH (cat. no. ab8245, 1:5,000; Abcam) were used to
incubate the membranes at 4°C for 12 h. After washing,
corresponding horseradish peroxidase (HRP)-conjugated secondary
antibodies (cat. no. ab13168, 1:1,000; Abcam) (cat. nos. sc-2350
and sc-2371, 1:1,000; Santa Cruz Biotechnology, Inc.) were used to
incubate the membranes at room temperature for 30 min. The
membranes were developed using Western Blotting Luminal reagent
(Santa Cruz Biotechnology, Inc.), and the bands were visualized on
X-ray film in a dark room.
Statistical analysis
Data in the present study are presented as the mean
± standard deviation. The differences between groups were analyzed
using one-way analysis of variance, and statistical analysis was
performed with SPSS software, version 17.0 (SPSS, Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Matrine attenuates left ventricular
functions impairment in experimental rat model of DbCM
The experimental rat model of DbCM is characterized
by elevated blood glucose levels and deteriorated LV function.
Matrine treatment did not change the blood glucose levels in
diabetic animals. The LV functions of control animals were not
affected by matrine. However, matrine administration increased
LVSP, +LVdP/dtmax and -LVdP/dtmax, whereas
LVEDP was decreased in the experimental rat model of DbCM. These
results were demonstrated in Table
III.
| Table III.Hemodynamic parameters and blood
glucose concentration comparisons between groups (n=3). |
Table III.
Hemodynamic parameters and blood
glucose concentration comparisons between groups (n=3).
| Group | Blood glucose
concentration (mmol/l) | Left ventricular
systolic pressure (mmHg) | Left ventricular
end-diastolic pressure (mmHg) | +Left ventricular
dP/dtmax (mmHg/sec) | -Left ventricular
dP/dtmax (mmHg/sec) |
|---|
| Ctrl |
9.25±3.32 |
107.82±12.61 |
22.18±5.59 |
1,988.35±221.54 |
2,677.16±345.27 |
| Mat |
10.15±3.10 |
112.37±13.05 |
23.29±5.75 |
1,973.00±224.45 |
2,713.05±332.19 |
| DbCM |
18.27±5.56a,b |
68.54±9.75a,b |
64.78±6.14a,b |
549.43±95.18a,b |
1,389.75±154.11a,b |
| DbCM+Mat |
19.34±4.91a,b |
82.10±7.45a–c |
48.20±5.61a–c |
1,244.75±111.43a–c |
1,799.48±177.42a–c |
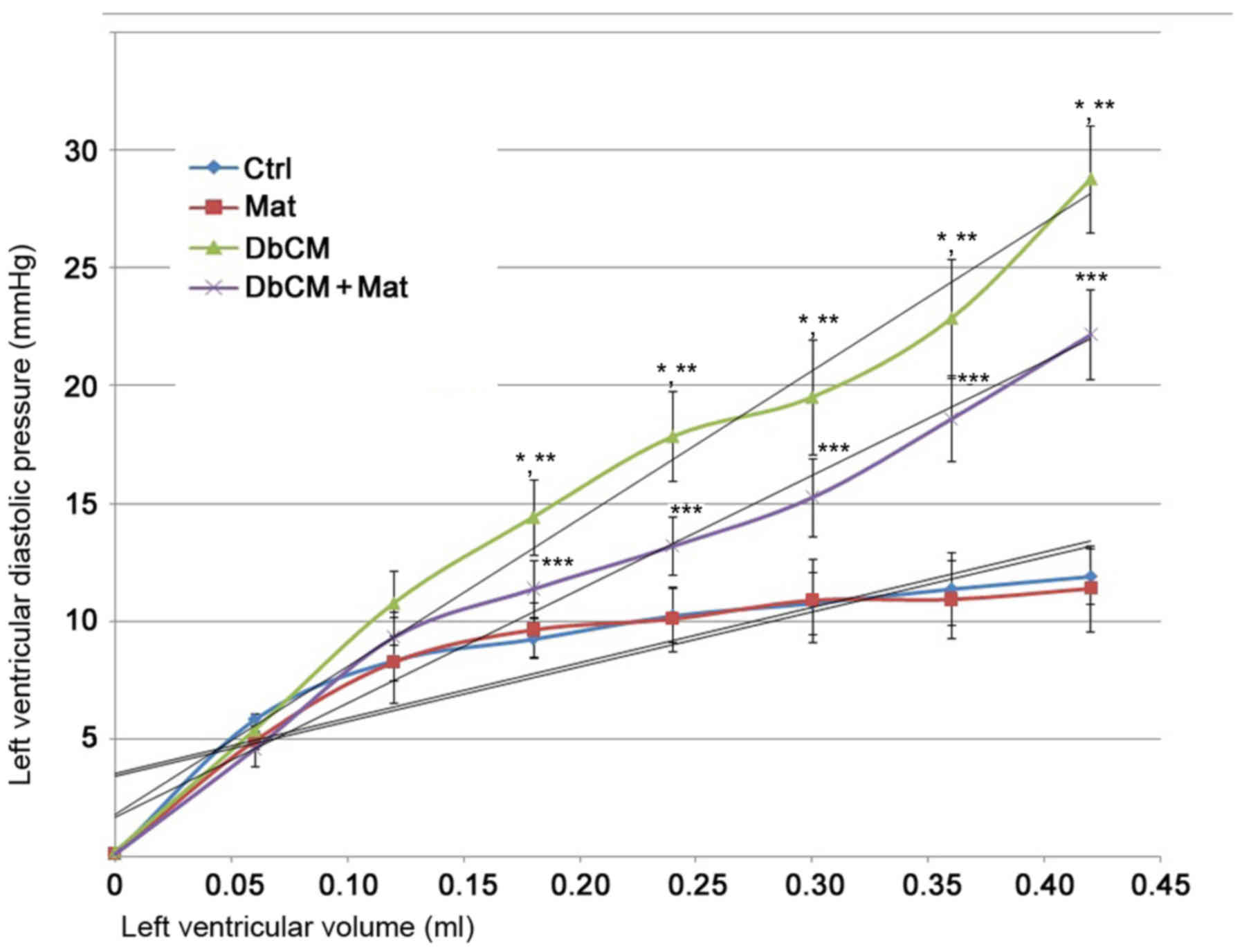
Matrine recovers cardiac compliance
loss in experimental rat model of DbCM
As demonstrated in the P-V curve in Fig. 1, the LV compliance was markedly
impaired in isolated hearts from rats with DbCM. These changes were
demonstrated by the significant P-V curve leftward shift. However,
the P-V curve of isolated hearts from the DbCM+Mat group displayed
a significant rightward shift, indicating markedly recovered LV
compliance.
Matrine inhibits cardiac fibrosis
induced by the activation of the TGF-β1/Smad signaling pathway in
rats with DbCM
Sirius red staining was used to evaluate the extent
of cardiac fibrosis, and captured images are shown in Fig. 2A. Compared with the Ctrl and Mat
groups, the optical density of Sirius Red staining increased
markedly in the DbCM group, suggesting the occurrence of aggravated
cardiac fibrosis. It was also observed that the optical density of
Sirius Red staining decreased markedly in the DbCM+Mat group,
indicating that matrine treatment alleviated cardiac fibrosis.
Fig. 2B demonstrates the effects
of matrine on the TGF-β1/Smad signaling pathway. Compared with Ctrl
and Mat, the protein expression level of TGF-β1 increased in the
DbCM group, but was markedly reduced by matrine administration in
the DbCM group. Correspondingly, compared with Ctrl and Mat, the
phosphorylation levels of Smad2 and 3 were also revealed to be
elevated in DbCM, indicating activation of the TGF-β1/Smad
signaling pathway. By contrast, matrine treatment markedly
decreased the expression level of TGF-β1, and further inhibited
phosphorylation of Smad2 and Smad3 in the DbCM+Mat group. As a
result, the expression level of collagen I increased markedly in
the DbCM group, but decreased in the DbCM+Mat group. The expression
level of inhibitory Smad (I-Smad), namely Smad7, was shown to be
decreased in the DbCM group, although this was not significantly
affected by matrine treatment.
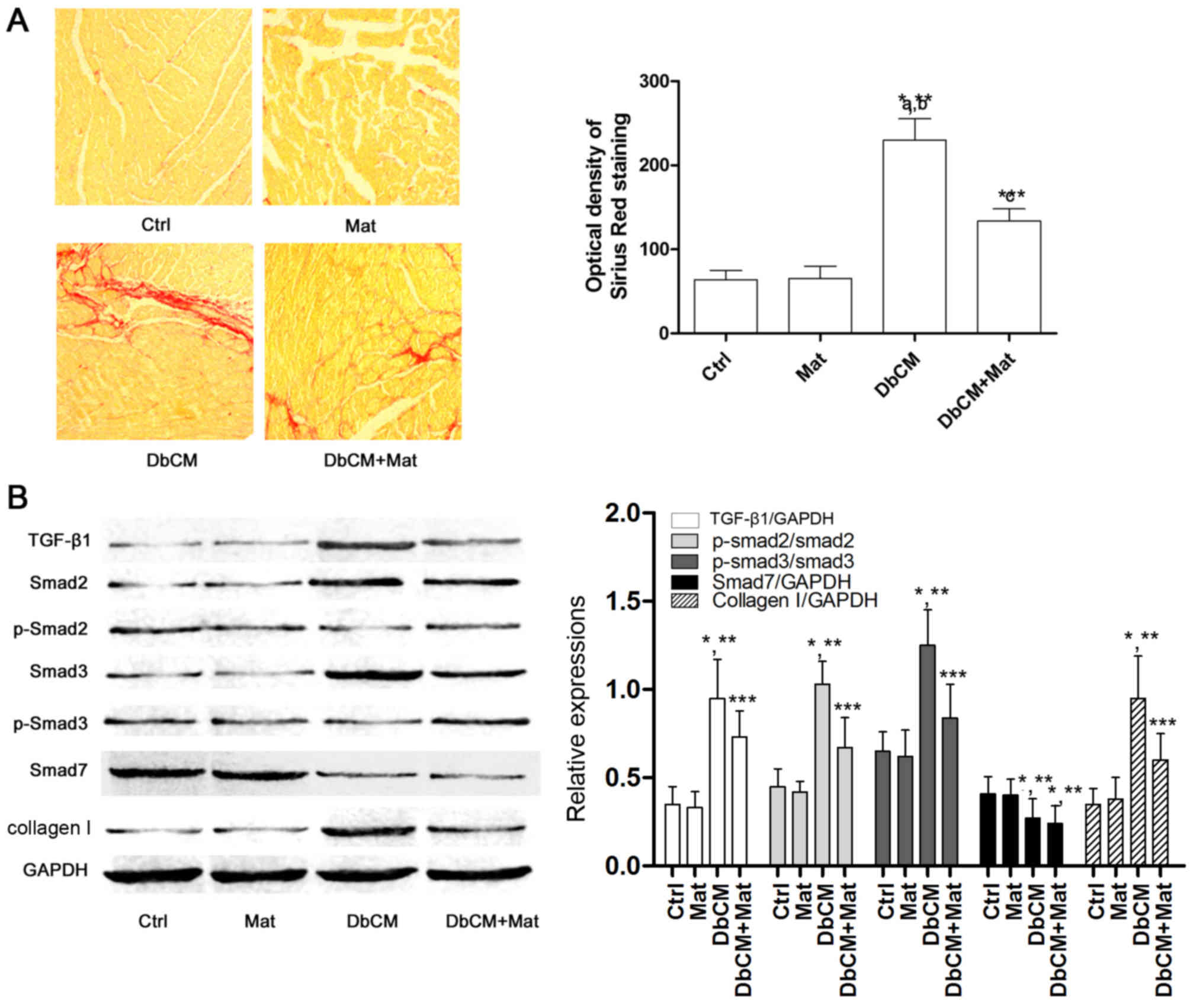 | Figure 2.Collagen deposition and TGF-β1/Smad
signaling pathway activation in cardiac tissue. (A) In the left
part of the figure, captured images of Sirius Red staining of
cardiac tissue harvested from each group (Ctrl, Mat, DbCM and
DbCM+Mat) are shown. The bar chart on the right shows the values of
the optical densities of Sirius Red in each group (results are
presented as the mean ± standard deviation). (B) Immunoblots of
TGF-β1, Smad2, phospho (p)-Smad2, Smad3, p-Smad3, Smad7, collagen I
and the loading control, GAPDH, are shown. The bar chart on the
right shows the relative expression levels of TGF-β1
(TGF-β1/GAPDH), Smad7 (Smad7/GAPDH), phosphorylated Smad2
(phospho-Smad2/Smad2), phosphorylated Smad3 (phospho-Smad3/Smad3)
and collagen I (collagen I/GAPDH). *P<0.05 vs. Ctrl; **P<0.05
vs. Mat; ***P<0.05 vs. DbCM. TGF-β1, transforming growth
factor-β1. Ctrl, control; Mat, matrine; DbCM, diabetic
cardiomyopathy. |
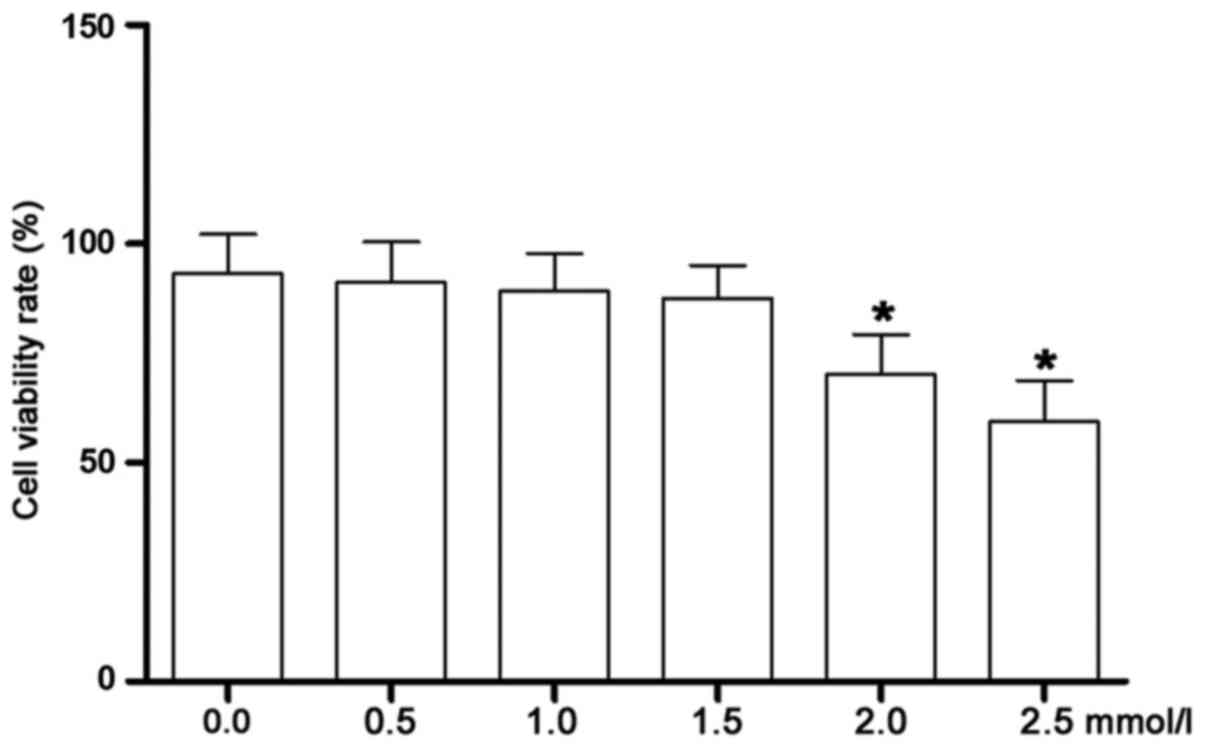
Matrine at low concentrations exerts
no inhibitory effects on cell proliferation of CFs incubated under
high-glucose conditions
As shown in Fig. 3,
the cell viability was assessed by MTT assay. The
high-glucose-incubated CFs were treated with matrine at serial
concentrations. Starting at 2.0 mmol/l, the viability of the CFs
was significantly decreased. Therefore, matrine at concentrations
of 0.25, 0.5 and 1.0 mmol/l were identified as the non-cytotoxic
concentrations that were selected for subsequent experiments.
Matrine treatment inhibits collagen
synthesis by suppressing activation of the TGF-β1/Smad signaling
pathway in high-glucose-incubated CFs
As shown in Fig. 4,
the effects of matrine on the TGF-β1/Smad signaling pathway in
cultured CFs were also investigated in the present study. The
expression of TGF-β1, as well as Smad2 and 3 phosphorylation
levels, were significantly increased in the CFs incubated under
high glucose conditions. Matrine treatment decreased the levels of
TGF-β1 and phosphorylated Smad2/3 in a concentration-dependent
manner. As a result, also in a concentration-dependent manner,
matrine decreased collagen I expression in high-glucose-incubated
CFs. Exposure to a high concentration of glucose suppressed the
expression of Smad7, although this was not affected by matrine.
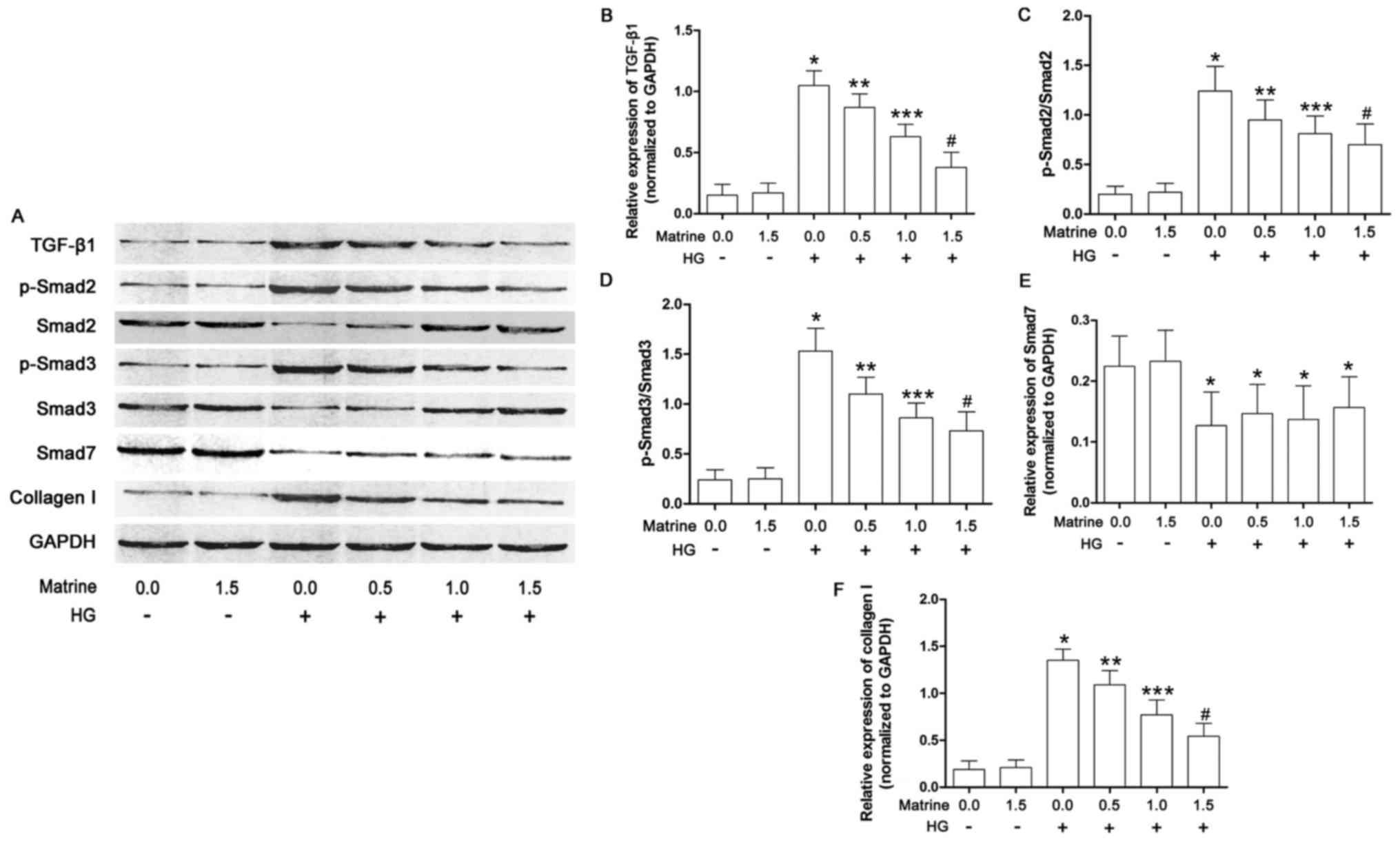 | Figure 4.TGF-β1/Smad signaling pathway
activation in isolated cardiac fibroblasts. (A) Immunoblots of
TGF-β1, Smad2, phospho (p)-Smad2, Smad3, p-Smad3, Smad7 and GAPDH
in isolated cardiac fibroblasts treated with serial concentrations
of matrine (0.0, 0.5, 1.0, 1.5, 2.0 and 2.5 mmol/l) and/or HG
medium are shown. (B) The bar chart shows the relative expression
levels of TGF-β1 (TGF-β1/GAPDH). (C) Bar charts also show the
phosphorylation of Smad2 (p-Smad2/Smad2), (D) the phosphorylation
of Smad3 (p-Smad3/Smad3), (E) the relative expression level of
Smad7 (Smad7/GAPDH), and (F) the relative expression level of
collagen I (collagen I/GAPDH). Data shown in B-F represent the mean
± standard deviation. *P<0.05 vs. first column; **P<0.05 vs.
third column; ***P<0.05 vs. fourth column; #P<0.05
vs. fifth column. TGF-β1, transforming growth factor-β1. |
Discussion
Diabetic macro- and micro-vascular complications are
the predominant causes of mortality due to diabetes. DbCM is one of
the principal clinical manifestations of diabetic cardiovascular
complications. DbCM is classified as a heart disease independent
from other heart diseases, including congenital heart disease,
coronary artery heart disease and valvular heart disease (13). DbCM is characterized by cardiac
remodeling, congestive heart failure and arrhythmias (14).
Cardiac fibrosis is one of the essential
pathological characteristics of cardiac remodeling, leading to
cardiac structural and functional alterations (15). It was considered that cardiac
fibrosis was associated with cardiac dysfunction and cellular
metabolic abnormalities attributable to diabetes (16,17).
Clearly evident cardiac fibrosis has been identified in diabetic
hearts, according to previous studies (18). CFs are activated in diabetic
hearts, and exert a vital role in regulating cardiac extracellular
matrix synthesis and deposition (5), causing cardiac dysfunction and
myocardium stiffening (19). In
the present study, it was revealed that the LV functions were
significantly impaired in rats with DbCM. The chamber stiffness
markedly increased in hearts isolated from rats with DbCM.
Many diabetic pathological products, including
catecholamines, endothelin, insulin- like growth factor-1, advanced
glycation end products and TGF-β1, have been correlated with organ
fibrosis (20–22). The molecular mechanism of cardiac
fibrosis in DbCM has yet to be fully elucidated. Previous studies
indicated that the TGF-β1-induced signaling pathway exerts an
important participatory role in cardiac fibrosis under certain
cardiac pathological conditions (23,24).
As transcription factors, Smads are the downstream effectors of
TGF-β1, which initiate transcription of genes encoding ECM
components (25,26). Studies have demonstrated that
activation of the TGF-β1/Smad signaling pathway is responsible for
hepatic, pulmonary and pancreatic fibrosis (6).
Specifically, the R-Smads (generally Smad2 and 3)
are activated following phosphorylation (27). The I-Smad, also termed Smad7,
suppresses the phosphorylation of Smad2/3 to inhibit the initiation
of downstream target-gene transcription (6,25).
In the present study, in hearts harvested from rats with DbCM, the
expression level of TGF-β1 and phosphorylation levels of Smad2/3
were increased markedly, indicating that the TGF-β1/Smad signaling
pathway was activated. At the same time, expression of Smad7 was
shown to be decreased in diabetic hearts, which also contributed to
the activation of TGF-β1/Smad signaling. In CFs incubated under
high glucose conditions, the TGF-β1/Smad signaling pathway was also
shown to be activated, as revealed by increased TGF-β1 expression,
decreased Smad7 expression and elevated levels of Smad2/3
phosphorylation. As a result, the synthesis of collagen I was shown
to beincreased in vivo and in vitro.
Extracted from the Chinese medical herb Kushen
(Sophora alopecuroides L.), matrine has been demonstrated to
be one of the most important bioactive pharmacological components
(28–30). Matrine was shown to affect the
bioactivities of immune regulation, anti-inflammation and
antioxidation (8,31). Prompted by a study on the
anti-fibrotic effect of matrine in several organs, including liver
and kidneys (29), the present
authors chose to investigate the effects of matrine on cardiac
fibrosis in the present study. It was shown that ECM synthesis and
deposition were suppressed in a rat model of DbCM where cardiac
fibrosis was clearly in evidence. Matrine administration attenuated
cardiac chamber stiffness, and thus the LV systolic and diastolic
functions were improved.
According to previous studies, TGF-β1/Smad signaling
activation in CFs was shown to induce cardiac fibrosis. Several
agents, including efonidipine and tanshinone, inhibited cardiac
fibrosis by inhibiting this pathway (32,33).
In the present study, the potential molecular mechanism of
matrine's anti-fibrosis effect in DbCM was investigated. The in
vivo data revealed that matrine administration inhibited
activation of the TGF-β1/Smad signaling pathway by suppressing the
expression of TGF-β1 and phosphorylation of Smad2/3 in diabetic
hearts. As a result, the accumulation of collagen I in myocytes was
attenuated. However, the expression of the I-Smad, Smad7, was not
shown to be affected by matrine. In the in vitro part of the
present study, matrine at low concentrations (i.e., in order to
avoid its cytotoxicity) was used to treat the
high-glucose-incubated CFs. The results suggested that matrine at
non-toxic concentrations deactivated the TGF-β1/Smad signaling
pathway by suppressing the expression of TGF-β1 and phosphorylation
of Smad2/3 in a concentration-dependent manner, rather than
affecting the expression of Smad7. Hence, the synthesis of collagen
I in CFs was correspondingly reduced.
In conclusion, the present study has shown that a
loss of cardiac compliance and function are the features of DbCM
characterized by cardiac fibrosis. Secondly, the TGF-β1/Smad
signaling pathway was activated to induce fibrosis in diabetic
hearts and CFs incubated under high glucose conditions. Thirdly,
matrine was shown to exert anti-fibrosis effects to improve cardiac
compliance and function by suppressing activation of the
TGF-β1/Smad signaling pathway in CFs in DbCM.
Acknowledgements
This study was supported by the National Scientific
Foundation of China (grant no. 81500308), the China Postdoctoral
Science Foundation (grant no. 2016M590956) and the Sailing
Foundation (grant no. LHJJ20159029).
Glossary
Abbreviations
Abbreviations:
|
DbCM
|
diabetic cardiomyopathy
|
|
ECM
|
extracellular matrix
|
|
CFs
|
cardiac fibroblasts
|
|
TGF-β1
|
transforming growth factor-β1
|
|
STZ
|
streptozotocin
|
|
LV
|
left ventricular
|
|
LVSP
|
left ventricular systolic pressure
|
|
LVEDP
|
left ventricular end-diastolic
pressure
|
|
+LVdP/dtmax
|
positive maximal rate of left
ventricular increased pressure
|
|
-LVdP/dtmax
|
negative maximal rate of left
ventricular increased pressure
|
|
IGF
|
insulin-like growth factor
|
|
FBS
|
fetal bovine serum
|
|
BCA
|
bicinchoninic acid
|
|
PVDF
|
polyvinylidene fluoride
|
|
DMSO
|
dimethyl sulfoxide
|
|
P-V
|
pressure-volume
|
References
|
1
|
Echouffo-Tcheugui JB and Dagogo-Jack S:
Preventing diabetes mellitus in developing countries. Nat Rev
Endocrinol. 8:557–562. 2012. View Article : Google Scholar
|
|
2
|
Lotfy M, Adeghate J, Kalasz H, Singh J and
Adeghate E: Chronic complications of diabetes mellitus: A mini
review. Curr Diabetes Rev. 13:3–10. 2017. View Article : Google Scholar
|
|
3
|
Boudina S and Abel ED: Diabetic
cardiomyopathy revisited. Circulation. 115:3213–3223. 2007.
View Article : Google Scholar
|
|
4
|
Westermeier F, Riquelme JA, Pavez M,
Garrido V, Díaz A, Verdejo HE, Castro PF, García L and Lavandero S:
New molecular insights of insulin in diabetic cardiomyopathy. Front
Physiol. 7:1252016. View Article : Google Scholar :
|
|
5
|
Travers JG, Kamal FA, Robbins J, Yutzey KE
and Blaxall BC: Cardiac fibrosis: The fibroblast awakens. Circ Res.
118:1021–1040. 2016. View Article : Google Scholar :
|
|
6
|
Xu F, Liu C, Zhou D and Zhang L:
TGF-beta/SMAD pathway and its regulation in hepatic fibrosis. J
Histochem Cytochem. 64:157–167. 2016. View Article : Google Scholar :
|
|
7
|
Song SE, Kim YW, Kim JY, Lee DH, Kim JR
and Park SY: IGFBP5 mediates high glucose-induced cardiac
fibroblast activation. J Mol Endocrinol. 50:291–303. 2013.
View Article : Google Scholar
|
|
8
|
Liu ZW, Wang JK, Qiu C, Guan GC, Liu XH,
Li SJ and Deng ZR: Matrine pretreatment improves cardiac function
in rats with diabetic cardiomyopathy via suppressing ROS/TLR-4
signaling pathway. Acta Pharmacol Sin. 36:323–333. 2015. View Article : Google Scholar :
|
|
9
|
Gao HY, Li GY, Lou MM, Li XY, Wei XY and
Wang JH: Hepatoprotective effect of Matrine salvianolic acid B salt
on carbon tetrachloride-induced hepatic fibrosis. J Inflamm (Lond).
9:162012. View Article : Google Scholar :
|
|
10
|
Liu Z, Zhao N, Zhu H, Zhu S, Pan S, Xu J,
Zhang X, Zhang Y and Wang J: Circulating interleukin-1β promotes
endoplasmic reticulum stress-induced myocytes apoptosis in diabetic
cardiomyopathy via interleukin-1 receptor-associated kinase-2.
Cardiovasc Diabetol. 14:1252015. View Article : Google Scholar :
|
|
11
|
Liu Z, Cai H, Zhu H, Toque H, Zhao N, Qiu
C, Guan G, Dang Y and Wang J: Protein kinase RNA-like endoplasmic
reticulum kinase (PERK)/calcineurin signaling is a novel pathway
regulating intracellular calcium accumulation which might be
involved in ventricular arrhythmias in diabetic cardiomyopathy.
Cell Signal. 26:2591–2600. 2014. View Article : Google Scholar
|
|
12
|
Li R, Xiao J, Qing X, Xing J, Xia Y, Qi J,
Liu X, Zhang S, Sheng X, Zhang X and Ji X: Sp1 mediates a
therapeutic role of miR-7a/b in angiotensin II-induced cardiac
fibrosis via mechanism involving the Tgf-β and mapks pathways in
cardiac fibroblasts. PLoS One. 10:e01255132015. View Article : Google Scholar :
|
|
13
|
Adeghate E: Molecular and cellular basis
of the aetiology and management of diabetic cardiomyopathy: A short
review. Mol Cell Biochem. 261:187–191. 2004. View Article : Google Scholar
|
|
14
|
Stratmann B, Worms J and Tschoepe D:
Diabetic cardiomyopathy/heart failure: News regarding etiology,
diagnosis, therapy. Dtsch Med Wochenschr. 139:2006–2009. 2014.(In
German).
|
|
15
|
Fan D, Takawale A, Lee J and Kassiri Z:
Cardiac fibroblasts, fibrosis and extracellular matrix remodeling
in heart disease. Fibrogenesis Tissue Repair. 5:152012. View Article : Google Scholar :
|
|
16
|
Xie Y, Liao J, Li M, Wang X, Yang Y, Ge J,
Chen R and Chen H: Impaired cardiac microvascular endothelial cells
function induced by Coxsackievirus B3 infection and its potential
role in cardiac fibrosis. Virus Res. 169:188–194. 2012. View Article : Google Scholar
|
|
17
|
Cavalera M, Wang J and Frangogiannis NG:
Obesity, metabolic dysfunction, and cardiac fibrosis:
Pathophysiological pathways, molecular mechanisms and therapeutic
opportunities. Transl Res. 164:323–335. 2014. View Article : Google Scholar :
|
|
18
|
Russo I and Frangogiannis NG:
Diabetes-associated cardiac fibrosis: Cellular effectors, molecular
mechanisms and therapeutic opportunities. J Mol Cell Cardiol.
90:84–93. 2016. View Article : Google Scholar
|
|
19
|
Hutchinson KR, Lord CK, West TA and
Stewart JA Jr: Cardiac fibroblast-dependent extracellular matrix
accumulation is associated with diastolic stiffness in type 2
diabetes. PLoS One. 8:e720802013. View Article : Google Scholar :
|
|
20
|
Chen M, Li H, Wang G, Shen X, Zhao S and
Su W: Atorvastatin prevents advanced glycation end products
(AGEs)-induced cardiac fibrosis via activating peroxisome
proliferator-activated receptor gamma (PPAR-γ). Metabolism.
65:441–453. 2016. View Article : Google Scholar
|
|
21
|
Bos R, Mougenot N, Findji L, Médiani O,
Vanhoutte PM and Lechat P: Inhibition of catecholamine-induced
cardiac fibrosis by an aldosterone antagonist. J Cardiovasc
Pharmacol. 45:8–13. 2005. View Article : Google Scholar
|
|
22
|
Liang M, Woodard LE, Liang A, Luo J,
Wilson MH, Mitch WE and Cheng J: Protective role of insulin-like
growth factor-1 receptor in endothelial cells against unilateral
ureteral obstruction-induced renal fibrosis. Am J Pathol.
185:1234–1250. 2015. View Article : Google Scholar :
|
|
23
|
Li FZ, Cai PC, Song LJ, Zhou LL, Zhang Q,
Rao SS, Xia Y, Xiang F, Xin JB, Greer PA, et al: Crosstalk between
calpain activation and TGF-β1 augments collagen-I synthesis in
pulmonary fibrosis. Biochim Biophys Acta. 1852:1796–1804. 2015.
View Article : Google Scholar
|
|
24
|
Cutroneo KR, White SL, Phan SH and Ehrlich
HP: Therapies for bleomycin induced lung fibrosis through
regulation of TGF-beta1 induced collagen gene expression. J Cell
Physiol. 211:585–589. 2007. View Article : Google Scholar
|
|
25
|
Meng XM, Tang PM, Li J and Lan HY:
TGF-β/Smad signaling in renal fibrosis. Front Physiol. 6:822015.
View Article : Google Scholar :
|
|
26
|
Zeglinski MR, Hnatowich M, Jassal DS and
Dixon IM: SnoN as a novel negative regulator of TGF-β/Smad
signaling: A target for tailoring organ fibrosis. Am J Physiol
Heart Circ Physiol. 308:H75–H82. 2015. View Article : Google Scholar
|
|
27
|
Tasanarong A, Kongkham S, Duangchana S,
Thitiarchakul S and Eiam-Ong S: Vitamin E ameliorates renal
fibrosis by inhibition of TGF-beta/Smad2/3 signaling pathway in UUO
mice. J Med Assoc Thai. 94 Suppl 7:S1–S9. 2011.
|
|
28
|
Huang J and Xu H: Matrine: Bioactivities
and structural modifications. Curr Top Med Chem. 16:3365–3378.
2016. View Article : Google Scholar
|
|
29
|
Yu JL, Li JH, Chengz RG, Ma YM, Wang XJ
and Liu JC: Effect of matrine on transforming growth factor β1 and
hepatocyte growth factor in rat liver fibrosis model. Asian Pac J
Trop Med. 7:390–393. 2014. View Article : Google Scholar
|
|
30
|
Yu J, Yang S, Wang X and Gan R: Matrine
improved the function of heart failure in rats via inhibiting
apoptosis and blocking β3adrenoreceptor/endothelial nitric oxide
synthase pathway. Mol Med Rep. 10:3199–3204. 2014. View Article : Google Scholar
|
|
31
|
Zhang L, Zhang H, Zhu Z, Lu X, Zhou M, Sun
X, He L, Bai Y and Ma L: Matrine regulates immune functions to
inhibit the proliferation of leukemic cells. Int J Clin Exp Med.
8:5591–5600. 2015.
|
|
32
|
Lei B, Hitomi H, Mori T, Nagai Y, Deguchi
K, Mori H, Masaki T, Nakano D, Kobori H, Kitaura Y and Nishiyama A:
Effect of efonidipine on TGF-β1-induced cardiac fibrosis through
Smad2-dependent pathway in rat cardiac fibroblasts. J Pharmacol
Sci. 117:98–105. 2011. View Article : Google Scholar :
|
|
33
|
Zhan CY, Tang JH, Zhou DX and Li ZH:
Effects of tanshinone IIA on the transforming growth factor
β1/Smadsignaling pathway in rat cardiac fibroblasts. Indian J
Pharmacol. 46:633–638. 2014. View Article : Google Scholar :
|