Introduction
Gliomas are the most common malignant neoplasm of
the central nervous system in adults, and accounts for 78% of
intracranial primary tumors (1).
Based on the degree of malignancy, according to the World Health
Organization (WHO) classification, glioma are classified into four
histopathologic grades: WHO grades I and II, which are considered
low-grade astrocytomas, and WHO grades III and IV, which are
considered high-grade astrocytomas (2). Glioblastoma multiforme (GBM), a WHO
grade IV glioma, is the most common and aggressive type of primary
brain tumor in humans (3). GBM
tumors are characterized by an increased degree of invasiveness and
rapid growth (4–7). Despite a combination of therapeutic
strategies, including surgery, radiotherapy and chemotherapy, the
average 5-year survival rate of patients with GBM is <5%
(8); the median survival time of
patients with GBM is ~1 year (9).
Therefore, the molecular mechanisms underlying the tumorigenicity
and development of GBM require further study, and novel therapeutic
treatments for patients with this disease are necessary.
MicroRNAs (miRNAs) belong to a family of small
(19–25 nucleotides long) noncoding RNA molecules (10), that negatively regulate gene
expression at the post-transcriptional level by imperfectly base
pairing with complementary sites within the 3′-untranslated regions
(3′-UTRs) of their target genes, thereby leading to degradation or
translational repression of target mRNAs (11). miRNAs have been hypothesized to
participate in modulating the expression of more than one-third of
human genes (12). Previous
studies have reported that a number of miRNAs are aberrantly
expressed in various types of human cancer, including GBM (13–15).
miRNAs are involved in tumorigenesis and tumor progression through
the regulation of a variety of biological processes, including cell
proliferation, cell cycle, apoptosis, differentiation, motility and
angiogenesis (16–18). Overexpression of miRNAs in human
cancers may act as oncogenes by negatively regulating the
expression of tumor-suppressor genes. By contrast, downregulated
miRNA expression levels may serve roles of tumor suppressors by
targeting oncogenes (19,20). Therefore, identifying misregulated
miRNAs and investigating their biological roles may provide novel
effective therapeutic targets for cancer treatment.
miR-376a, which has been mapped to the 14q32.31
locus, is abnormally expressed in multiple human cancers (21–23).
However, the expression pattern and role of miR-376a in GBM, as
well as the underlying molecular mechanisms remain to be
elucidated. SP1 was previously reported to be upregulated in GBM
and associated with the WHO grading and survival status (24). Additionally, SP1 may be involved in
initiation and progression of GBM by regulating proliferation, cell
cycle and invasion (24–27). The present study demonstrated that
miR-376a is expressed at low levels in GBM tissues and cell lines.
miR-376a upregulation inhibits GBM cell proliferation and invasion
by directly targeting specificity protein 1 (SP1).
Materials and methods
Clinical specimens
The present study was approved by the Ethics
Committee of Central Hospital of Linyi (Linyi, China), and written
informed consent was obtained from patients with GBM for use of
tissues. A total of 26 paired GBM tissues and adjacent normal brain
tissues were obtained from the patients with GBM (17 males and 9
females; age range, 42–77 years) who underwent surgical resection
at the Central Hospital of Linyi between August 2012 and February
2016. None of the patients were treated with radiotherapy or
chemotherapy prior to surgery. Collected specimens were immediately
frozen following surgical resection and stored in liquid
nitrogen.
Cell lines and transfection
Human GBM cell lines (U138, U251, LN229, T98) were
purchased from Shanghai Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). GBM cell lines were cultured in
Dulbecco's modified Eagle medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.), 100
U/ml penicillin and 100 mg/ml streptomycin (Invitrogen; Thermo
Fisher Scientific, Inc.). Normal human astrocytes (NHAs) were
obtained from ScienCell Research Laboratories, Inc. (San Diego, CA,
USA) and maintained in Astrocyte Medium (ScienCell Research
Laboratories, Inc.). All cell lines were cultured at 37°C in a
humidified chamber containing 5% CO2.
miR-376a mimics and miRNA negative control (miR-NC)
were obtained from Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
The miR-376a mimics sequence was 5′-AUCAUAGAGGAAAAUCCACGU-3′ and
the miR-NC sequence was 5′-UUCUCCGAACGUGUCACGUTT-3′. The
SP1-overexpression plasmid pcDNA3.1-SP1 and bland pcDNA3.1 plasmid
were purchased from Shanghai GenePharma Co., Ltd. (Shanghai,
China). For transfection, cells were seeded in a 6-well plate at a
density of 5×105 cells/well. When 70% confluence was
achieved, cells were transfected with miRNA mimics (100 pmol),
miR-NC (100 pmol) or plasmids (2 µg) at room temperature using
Lipofectamine 2000 Transfection Reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Following transfection for 48 h, reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) was performed to detect
miR-376a or SP1 mRNA level. CCK-8 and Matrigel chamber invasion
assays were conducted at 24 and 48 h post transfection. Western
blot analysis was carried out at 72 h following transfection.
RNA extraction and RT-qPCR
Total RNA form tissue specimens (100 mg) or cells
(1.5×106) was isolated using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. For the determination of miR-376a expression levels, cDNA
was synthesized from 1 µg total RNA using TaqMan MicroRNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.), following the manufacturer's protocol. Relative miR-376a
expression was quantified using the TaqMan MicroRNA PCR kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.); U6 small
nuclear RNA was used as an endogenous control. The following
thermocycling conditions were used for the PCR: 50°C for 2 min,
95°C for 10 min; 40 cycles of 95°C for 15 sec and 60°C for 60 sec.
For SP1 mRNA expression, total RNA was reverse transcribed into
cDNA using PrimeScript RT Reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China). A SYBR Premix Ex Taq™ kit (Takara
Biotechnology Co., Ltd.) was used to detect SP1 mRNA expression;
GAPDH was used as an internal control. The following thermocycling
conditions were used for the PCR: 5 min at 95°C, followed by 40
cycles of 95°C for 30 sec and 65°C for 45 sec. qPCR was performed
using an ABI-7500 Fast Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.), and data was analyzed using the
2−ΔΔCq method (28).
The following primers were used in the present study: miR-376a,
forward 5′-GTGCAGGGTCCGAGGT-3′, reverse 5′-ATCATAGAGGAAAATCCACG-3′;
U6, forward 5′-ATTGGAACGATACAGAGAAGATT-3′, reverse
5′-GGAACGCTTCACGAATTTG-3′; SP1, forward 5′-TGGTGGGCAGTATGTTGT-3′,
reverse 5′-GCTATTGGCATTGGTGAA-3′; and GAPDH, forward
5′-ATGTCGTGGAGTCTACTGGC-3′, reverse 5′-TGACCTTGCCCACAGCCTTG-3′.
Cell Counting Kit-8 (CCK-8)
proliferation assay
The proliferative abilities of GBM U251 cells were
determined using CCK-8 proliferation assays (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan). Transfected cells were seeded
in 96-well plates at 3,000 cells/well and cultured for 0, 24, 48
and 72 h at 37°C. At the indicated times, 10 µl CCK-8 Regent was
added into each well and, following 2 h incubation at 37°C, the
absorbance was measured at a wavelength of 450 nm using an
automatic multi-well spectrophotometer (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Matrigel chamber invasion assay
Transwell filters (BD Biosciences, Franklin Lakes,
NJ, USA) coated with Matrigel (BD Biosciences) on the upper surface
of the 8 µm pore polycarbonate membrane were used to assess cell
invasion, in accordance with the manufacturer's protocol.
Transfected cells (1×105 cells/well) were seeded in the
upper well of the Transwell filters in FBS-free DMEM. The lower
Transwell chamber were filled with DMEM medium containing 10% FBS
as a chemoattractant. Following incubation for 24 h at 37°C, cells
on the top membranes were removed using cotton swabs. Invasive
cells on the lower membrane were fixed with 100% methanol at room
temperature for 15 min, stained with 0.5% crystal violet solution
at room temperature for 15 min and washed with PBS. Images of
invasive cells were captured and cells were counted under an
Olympus IX53 inverted fluorescent microscope (Olympus Corporation,
Tokyo, Japan) in five independent fields for each well to analyze
the invasive ability of cells.
Bioinformatics analysis and luciferase
reporter assay
Potential targets of miR-376a were predicted using
miRanda (http://www.microrna.org/microrna/home.do) and
TargetScan (http://www.targetscan.org). For
luciferase reporter assay, reporter plasmids, including
pGL3-SP1-3′UTR-wild-type (Wt) and pGL3-SP1-3′UTR-mutant (Mut), were
synthesized and verified by Shanghai GenePharma Co., Ltd. Cells
were seeded into a 24-well plate at a density of 1.5×105
cells/well and cultured overnight at 37°C prior to co-transfection
with pGL3-SP1-3′UTR-Wt or pGL3-SP1-3′UTR-Mut with miR-376a mimics
or miR-NC using Lipofectamine® 2000 transfection reagent
at 37°C. A total of 48 h following transfection, Firefly and
Renilla luciferase activities were determined using the
Dual-Luciferase Reporter Assay System (Promega Corporation,
Madison, WI, USA), according to the manufacturer's protocol.
Renilla luciferase was selected for normalization. Experiments were
repeated three times independently.
Protein extraction and western blot
analysis
Total protein was extracted from tissue samples (200
mg) and U251 cells (1.5×106) using Total Protein
Extraction kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China)
supplemented with protease inhibitors (EMD Millipore, Billerica,
MA, USA). Protein concentrations were measured using a
Bicinchoninic Acid Assay kit (Beyotime Institute of Biotechnology,
Haimen, China). Equal quantities of protein (20 µg/lane) were
separated by 10% SDS-PAGE and transferred to polyvinylidene
difluoride membranes (EMD Millipore). The membranes were
subsequently blocked with Tris-buffered saline and 0.05% Tween-20
(TBST) containing 5% non-fat milk at room temperature for 1 h,
followed by incubation with primary antibodies at 4°C overnight.
Primary antibodies used in the present study include mouse anti-SP1
monoclonal antibody (1:1,000; cat. no. sc-420) and mouse anti-GAPDH
monoclonal antibody (1:1,000; cat. no. sc-32233), both from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). Following three washes
with TBST, the membranes were incubated with a goat anti-mouse
horseradish peroxidase-conjugated secondary antibody (1:5,000
dilution; cat. no. sc-2005; Santa Cruz Biotechnology) at room
temperature for 2 h. Protein bands were visualized using an
Enhanced Chemiluminescence kit (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol, and
quantification of band intensities was conducted using ImageJ 1.49
software (National Institutes of Health, Bethesda, MD, USA). GAPDH
was used as a loading control.
Statistical analysis
Data are presented as the mean ± standard deviation.
Each assay was repeated three times independently. Data were
analyzed with two-tailed Student's t-test or one-way analysis of
variance using SPSS statistical software (version 19.0; IBM Corp.,
Armonk, NY, USA). Student-Newman-Keuls method was used to compare
differences between two groups in multiple comparison analyses
following one-way analysis of variance. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-376a is downregulated in GBM
tissue samples and cell lines
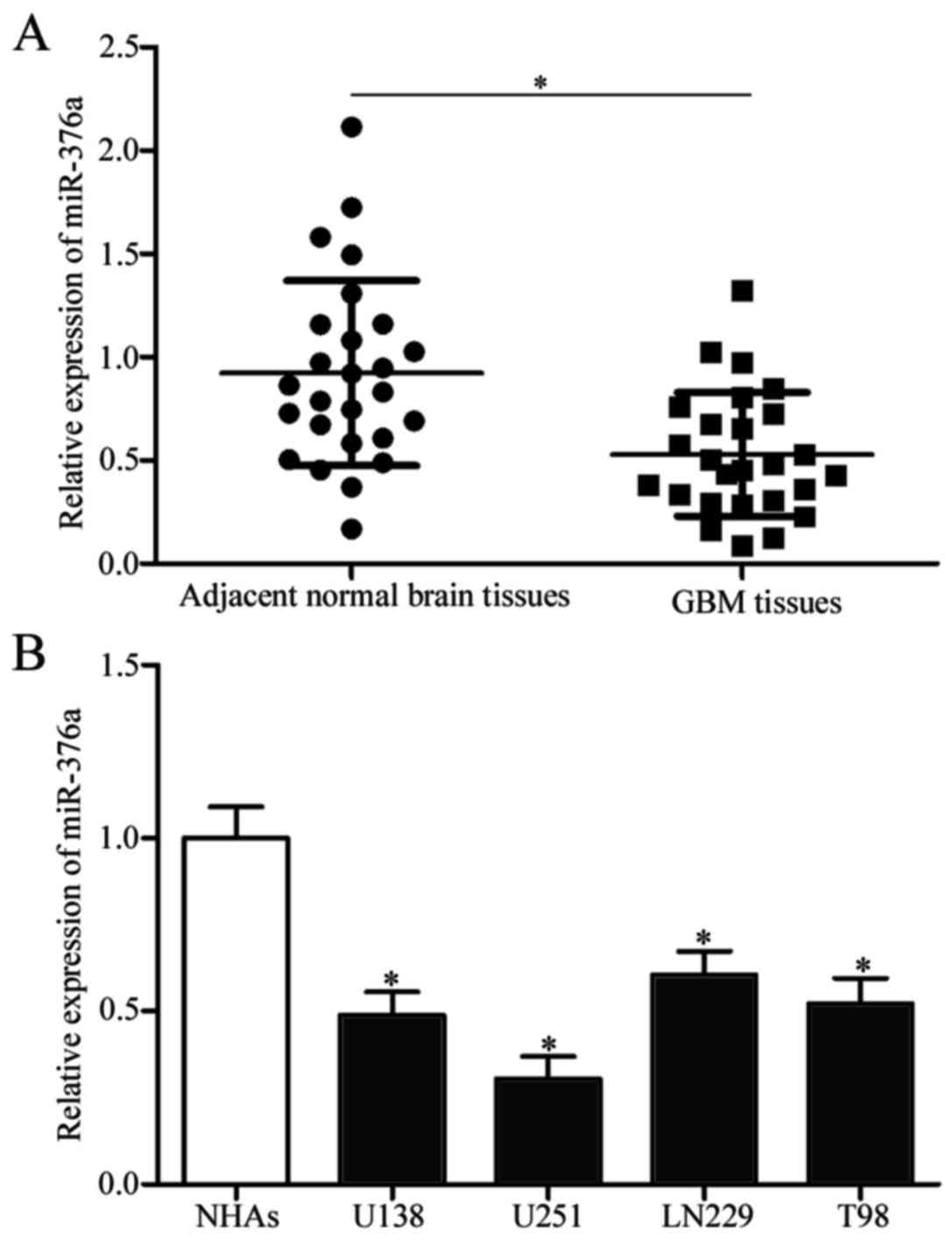
Expression levels of miR-376a were detected in GBM
tissues and adjacent normal brain tissues to investigate the
potential roles of miR-376a in GBM. miR376a expression levels were
significantly reduced in GBM tissues compared with adjacent normal
brain tissues (P<0.05; Fig.
1A). Subsequently, miR-376a expression was determined in GBM
cell lines (U138, U251, LN229 and T98) and normal human astrocytes
(NHAs). miR-376a expression levels were significantly lower in all
GBM cell lines compared with NHA cells (P<0.05; Fig. 1B). These results suggested that
miR-376a may serve a role in GBM progression.
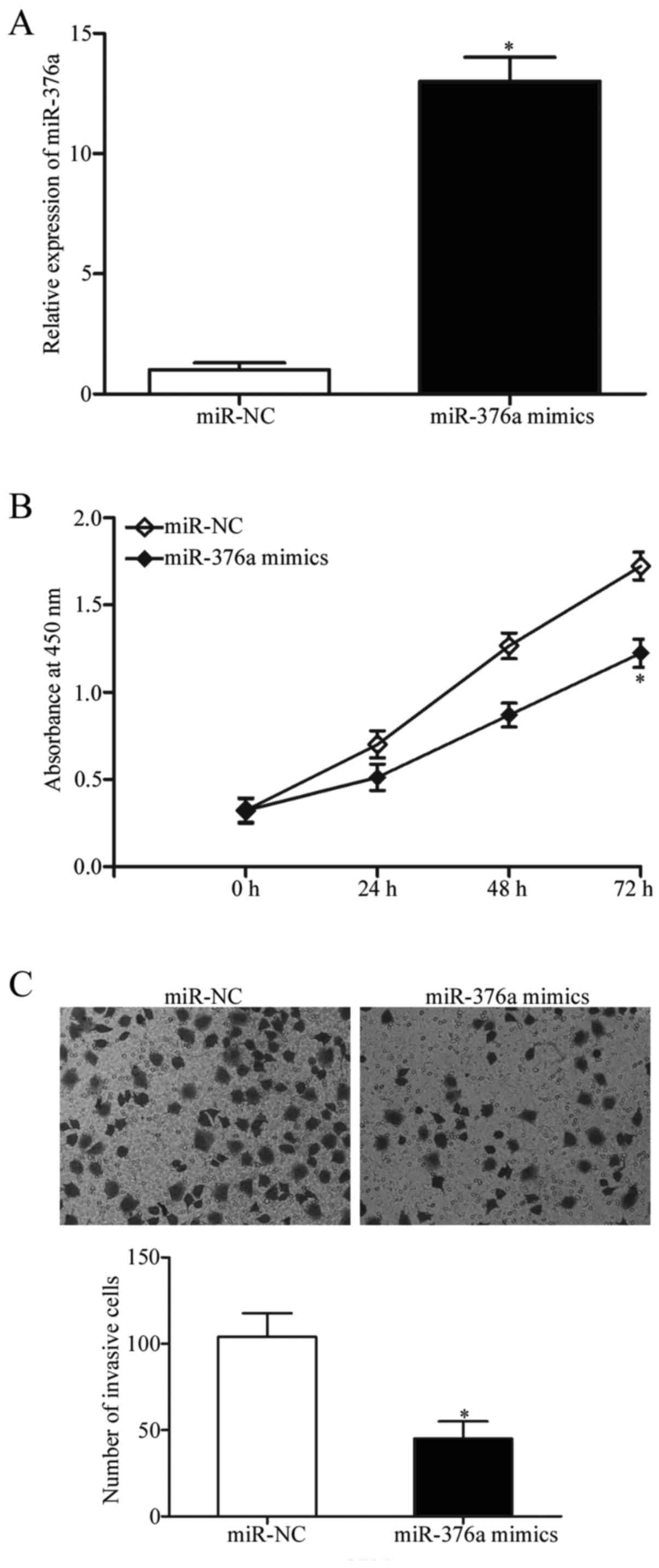
miR-376a overexpression inhibits cell
proliferation and invasion in GBM cell lines
As aforementioned, miR-376a expression was reduced
in GBM tissues and cell lines; therefore, it was hypothesized that
miR-376a may be a tumor suppressor in GBM. miR-376a mimics or
miR-NCs were transfected into U251 cells, and the transfection
efficiency of miR-376a was confirmed using RT-qPCR, which
demonstrated that miR-376a expression was significantly upregulated
in cells transfected with miR-376a mimics compared with cells
transfected with miR-NC (P<0.05; Fig. 2A). The effects of miR-376a
overexpression on the proliferation of GBM cells were determined by
CCK-8 proliferation assay. Upregulation of miR-376a expression
levels significantly suppressed U251 cell proliferation at 72 h
(P<0.05; Fig. 2B). Matrigel
chamber invasion assays were performed to determine whether
miR-376a influenced invasive abilities of GBM cells. The results
demonstrated that increased miR-376a expression significantly
decreased the invasive capacities of U251 cells compared with
miR-NC-transfected cells (P<0.05; Fig. 2C). These results suggested that
miR-376a may serve a role as a tumor suppressor in GBM
progression.
SP1 is a target of miR-376a in
GBM
Bioinformatics analysis was used to predict
potential targets of miR-376a and to investigate the mechanisms
underlying miR-376a-mediated regulation of GBM progression. A
number of candidate genes were predicted, including SP1 (Fig. 3A). SP1 was selected for further
verification as it was previously demonstrated to function as a
proto-oncogene in GBM (24–27).
Luciferase reporter assays were performed to confirm this
hypothesis. The SP1-3′-UTR-Wt or SP1-3′-UTR-Mut reporter plasmid
was co-transfected into U251 cells with miR-376a mimics or miR-NC.
The results demonstrated that miR-376a mimics reduced the
luciferase activity level of SP1-3′-UTR-Wt compared with miR-NC
(P<0.05; Fig. 3B), whereas no
significant difference was identified for SP1-3′-UTR-Mut. RT-qPCR
and western blot analyses were used to confirm the effects of
miR-376a on SP1 expression in GBM cells. The results indicated that
both mRNA and protein levels of SP1 were significantly reduced by
miR-376a overexpression in U251 cells (P<0.05; Fig. 3C and D, respectively).
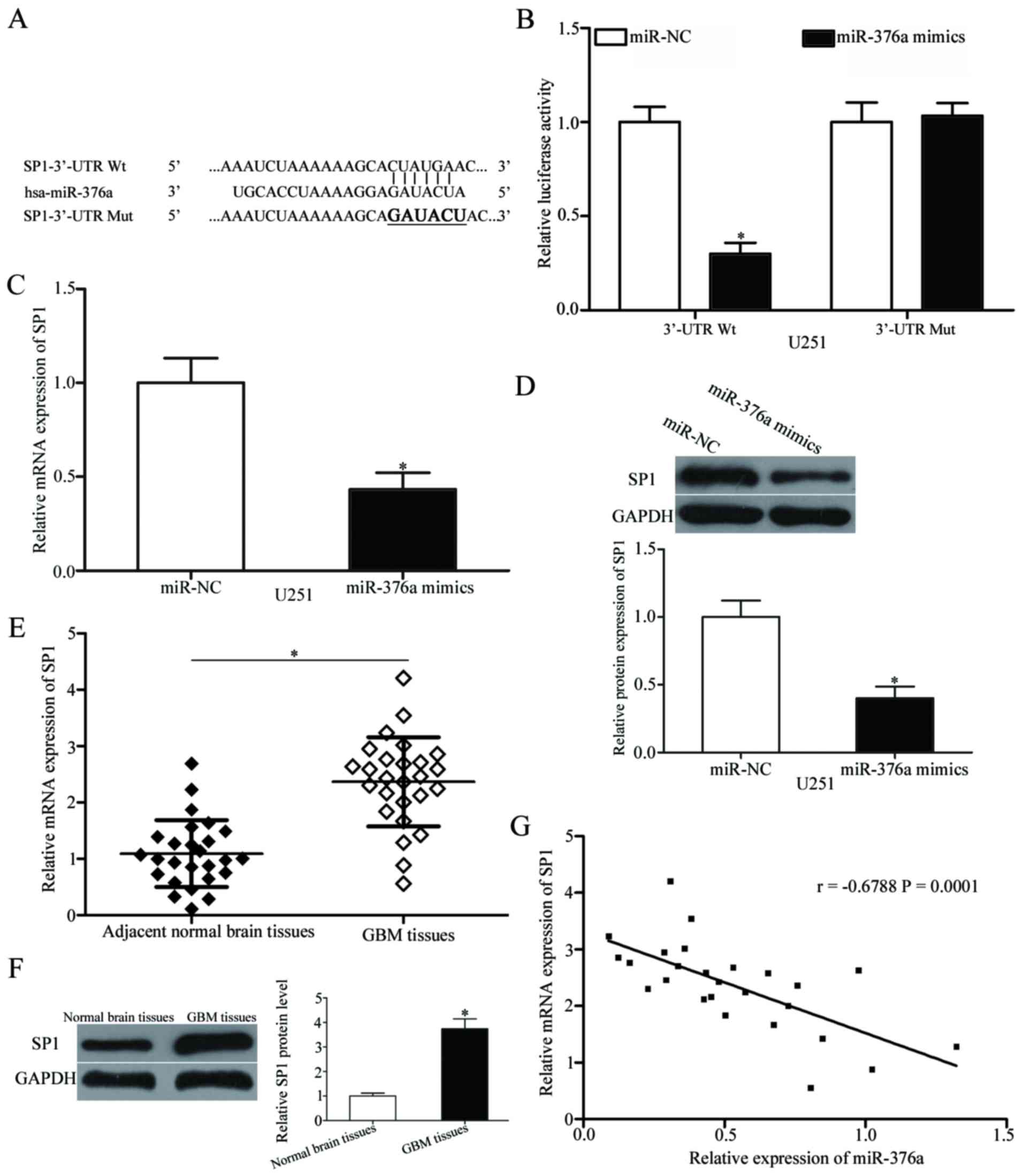 | Figure 3.SP1 is a direct target gene of
miR-376a in GBM. (A) Predicted miR-376a target sequence in the
3′-UTR of SP1 Wt and Mut containing six mutated nucleotides in the
3′-UTR of SP1. (B) Luciferase reporter assays were performed in
U251 cells following co-transfection with pGL3-SP1-3′UTR-Wt or
pGL3-SP1-3′UTR-Mut, together with miR-376a mimics or miR-NC. Data
are presented as the mean ± standard deviation; *P<0.05 vs.
miR-NC. (C) RT-qPCR and (D) western blot analysis of the SP1 mRNA
and protein expression levels, respectively, in U251 cells
following transfection with miR-376a mimics or miR-NC; *P<0.05
vs. miR-NC. Expression levels of SP1 (E) mRNA and (F) protein in
GBM tissues and adjacent normal brain tissues detected using
RT-qPCR and western blot analysis, respectively. Data are presented
as the mean ± standard deviation; *P<0.05 vs. adjacent normal
brain tissues. (G) Spearman's correlation analysis of the
association between miR-376a and SP1 mRNA level in GBM tissues.
r=−0.6788; P=0.0001. GBM, glioblastoma multiforme; hsa, Homo
sapiens RNA miR, microRNA; mut, mutant; NC, negative control;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; SP1, specificity protein 1; UTR, untranslated region; wt,
wild-type. |
SP1 mRNA and protein levels were also measured in
GBM tissues and adjacent normal brain tissues. SP1 mRNA and protein
levels were elevated in GBM tissues compared with adjacent normal
brain tissues (P<0.05; Fig. 3E and
F). Spearman's correlation analysis indicated a significant
negative correlation between miR-376a expression and SP1 mRNA
levels in GBM tissues (Fig. 3G;
r=−0.6788, P=0.0001). These data indicated that SP1 was a direct
target of miR-376a in GBM.
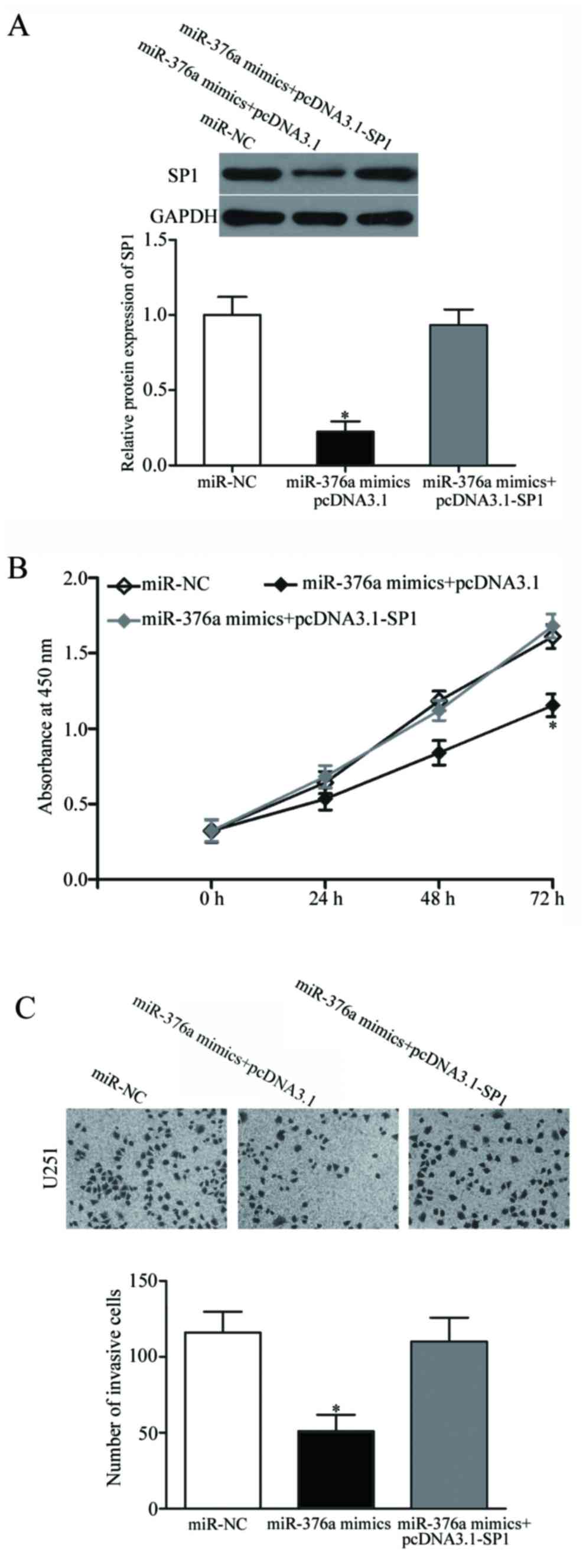
SP1 overexpression reduces the
suppressive effects of miR-376a on GBM cells
Rescue experiments were performed to confirm whether
SP1 mediates the biological role of miR-376a in GBM cells. U251
cells were transfected with miR-376a mimics, with or without the
pcDNA3.1-SP1 overexpression vector. Western blot analysis revealed
that the miR376a mimics-induced reduction of SP1 protein expression
was restored by co-transfection with pcDNA3.1-SP1 (P<0.05;
Fig. 4A). Subsequently, CCK-8
proliferation and Matrigel invasion assays demonstrated that
overexpression of SP1 in U251 cells reversed the suppressive
effects of miR-376a mimics on cell proliferation and invasion
(P<0.05; Fig. 4B and C). These
results suggested that miR-376a may inhibit GBM progression, at
least in part, through negative regulation of SP1.
Discussion
A number of previous studies have demonstrated that
miRNAs are aberrantly expressed in almost all types of cancers,
including GBM (29–31). Abnormally expressed miRNAs are
commonly associated with malignant biological behaviors of GBM,
including malignant growth, proliferation, apoptosis, invasion,
metastasis and chemotherapy resistance (32). In the present study, miR-376a was
downregulated in GBM tissues and cell lines. Functional assays
revealed that miR-376a overexpression led to decreased GBM cell
proliferation and invasion in vitro. Additionally, SP1 was
confirmed as a direct target gene of miR-376a in GBM. mRNA and
protein expression levels of SP1 in GBM tissues were markedly
upregulated compared with the adjacent normal brain tissues.
Furthermore, expression of SP1 mRNA was inversely correlated with
the expression of miR-376a in GBM tissues. The restored expression
of SP1 partially attenuated miR-376a-inhibited cell proliferation
and invasion. The results of the present study suggested that
miR-376a may be a therapeutic target for treatment of patients with
GBM.
miR-376a is abnormally expressed in several human
cancers. For example, miR-376a expression is upregulated in ovarian
cancer and associated with International Federation of Gynecology
and Obstetrics stages of ovarian cancer (21,33);
however, miR-376a is downregulated in hepatocellular carcinoma
(22). Low miR-376a expression
levels are associated with low serum α-fetoprotein levels in
hepatocellular carcinoma (22). In
prostate cancer, miR-376a was reported to be downregulated in tumor
tissues and cell lines, it was associated with an increased
incidence of metastatic events and elevated prostate specific
antigen levels, and similar trends were observed for lymph node
invasion and Gleason score (34).
miR-376a downregulation was identified in giant cell tumors of bone
(35). These data suggested that
miR-376a may be a prognostic marker of certain cancers.
miR-376a may serves roles in carcinogenesis and
cancer progression; for example, Yang et al reported that
miR-376a overexpression promoted ovarian cancer cell growth and
metastasis (21). However, Zheng
et al demonstrated that miR-376a serves a role as a tumor
suppressor in hepatocellular carcinoma by inhibiting cell
proliferation and increasing apoptosis (22). Formosa et al demonstrated
that miR-376a overexpression decreases cell growth and motility,
and increases apoptosis in prostate cancer (34). A study by Fellenberg et al
indicated that restoration of miR-376a expression attenuates cell
proliferation, migration, colony formation and spheroid formation
of giant cell tumors of bone (35). The aforementioned data appears
contradictory as miR-376a acted as an oncogene in certain cancers
and as a tumor suppressor in others. This contradiction may be
explained by imperfect complementarity of interactions between
miRNAs and target genes (36).
Several miR-376a target genes have been previously
identified, including Krüppel-like factor 15 and caspase-8 in
ovarian cancer, p85α and histone deacetylase in hepatocellular
carcinoma, insulin-like growth factor 1 receptor in melanoma,
cytochrome c oxidase assembly factor 1 homolog, and nucleoporin
GLE1 and protein disulfide-isomerase A6 in giant cell tumors of
bone (21–23,35,37).
In the present study, SP1 was identified as a novel target of
miR-376a. SP1 is a member of the Sp/Krüppel-like factor
transcription factor family and is upregulated in several types of
human cancer, including gastric, colorectal, prostate, thyroid and
liver (38–42). SP1 serves a role in a number of
pathophysiological processes, including cell proliferation, cell
cycle, apoptosis, angiogenesis and metastasis (43–45).
Therefore, SP1 may be a therapeutic target for suppression of
tumorigenesis and tumor development.
SP1 was upregulated in glioma tissues and cell
lines. Elevated SP1 expression was associated with the WHO grading
and survival status of patients with glioma. Patients with lower
SP1 expression demonstrated better overall survival compared with
patients with increased SP1 expression (24); suggesting that SP1 expression may
be a prognostic indicator of survival of patients with glioma
(24). Functional experiments
demonstrated that SP1 serves a role in glioma progression by
regulating proliferation, cell cycle and invasion (24–27).
In the present study, miR-376a was demonstrated to target SP1 mRNA
to inhibit GBM cell proliferation and invasion, which suggested
that miR-376a/SP1-based targeted therapy may be a novel therapeutic
strategy for treatment of patients with GBM.
In conclusion, the present study demonstrated that
miR-376a expression was reduced in GBM tissues and cell lines.
miR-376a overexpression exhibited a suppressive effect on
proliferation and invasion of GBM cells, which may occur by
directly targeting SP1. miR-376a may be a novel therapeutic target
for the treatment of GBM.
References
|
1
|
Miller CR and Perry A: Glioblastoma. Arch
Pathol Lab Med. 131:397–406. 2007.
|
|
2
|
Komori T, Sasaki H and Yoshida K: Revised
WHO classification of tumours of the central nervous system:
Summary of the revision and perspective. No Shinkei Geka.
44:625–635. 2016.(In Japanese).
|
|
3
|
Easaw JC, Mason WP, Perry J, Laperrière N,
Eisenstat DD, Del Maestro R, Bélanger K, Fulton D and Macdonald D:
Canadian Glioblastoma Recommendations Committee: Canadian
recommendations for the treatment of recurrent or progressive
glioblastoma multiforme. Curr Oncol. 18:e126–e136. 2011. View Article : Google Scholar :
|
|
4
|
Smith AW, Mehta MP and Wernicke AG: Neural
stem cells, the subventricular zone and radiotherapy: Implications
for treating glioblastoma. J Neurooncol. 128:207–216. 2016.
View Article : Google Scholar
|
|
5
|
Binder DC, Davis AA and Wainwright DA:
Immunotherapy for cancer in the central nervous system: Current and
future directions. Oncoimmunology. 5:e10820272016. View Article : Google Scholar
|
|
6
|
Katakowski M and Chopp M: Exosomes as
tools to suppress primary brain tumor. Cell Mol Neurobiol.
36:343–352. 2016. View Article : Google Scholar
|
|
7
|
Khosla D: Concurrent therapy to enhance
radiotherapeutic outcomes in glioblastoma. Ann Transl Med.
4:542016.
|
|
8
|
Kegelman TP, Hu B, Emdad L, Das SK, Sarkar
D and Fisher PB: In vivo modeling of malignant glioma: The road to
effective therapy. Adv Cancer Res. 121:261–330. 2014. View Article : Google Scholar
|
|
9
|
Van Meir EG, Hadjipanayis CG, Norden AD,
Shu HK, Wen PY and Olson JJ: Exciting new advances in
neuro-oncology: The avenue to a cure for malignant glioma. CA
Cancer J Clin. 60:166–193. 2010. View Article : Google Scholar :
|
|
10
|
Alvarez-Garcia I and Miska EA: MicroRNA
functions in animal development and human disease. Development.
132:4653–4662. 2005. View Article : Google Scholar
|
|
11
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar
|
|
12
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar
|
|
13
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar
|
|
14
|
Wang Z, Yang J, Xu G, Wang W, Liu C, Yang
H, Yu Z, Lei Q, Xiao L, Xiong J, et al: Targeting miR-381-NEFL axis
sensitizes glioblastoma cells to temozolomide by regulating
stemness factors and multidrug resistance factors. Oncotarget.
6:3147–3164. 2015. View Article : Google Scholar
|
|
15
|
Chang JH, Hwang YH, Lee DJ, Kim DH, Park
JM, Wu HG and Kim IA: MicroRNA-203 modulates the radiation
sensitivity of human malignant glioma cells. Int J Radiat Oncol
Biol Phys. 94:412–420. 2016. View Article : Google Scholar
|
|
16
|
Sampson VB, Yoo S, Kumar A, Vetter NS and
Kolb EA: MicroRNAs and potential targets in osteosarcoma: Review.
Front Pediatr. 3:692015. View Article : Google Scholar :
|
|
17
|
Luna-Aguirre CM, de la Luz Martinez-Fierro
M, Mar-Aguilar F, Garza-Veloz I, Treviño-Alvarado V, Rojas-Martinez
A, Jaime-Perez JC, Malagon-Santiago GI, Gutierrez-Aguirre CH,
Gonzalez-Llano O, et al: Circulating microRNA expression profile in
B-cell acute lymphoblastic leukemia. Cancer Biomark. 15:299–310.
2015. View Article : Google Scholar
|
|
18
|
Kagiya T: MicroRNAs and osteolytic bone
metastasis: The roles of MicroRNAs in tumor-induced osteoclast
differentiation. J Clin Med. 4:1741–1752. 2015. View Article : Google Scholar :
|
|
19
|
Ventura A and Jacks T: MicroRNAs and
cancer: Short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar :
|
|
20
|
Gao B, Gao K, Li L, Huang Z and Lin L:
miR-184 functions as an oncogenic regulator in hepatocellular
carcinoma (HCC). Biomed Pharmacother. 68:143–148. 2014. View Article : Google Scholar
|
|
21
|
Yang L, Wei QM, Zhang XW, Sheng Q and Yan
XT: MiR-376a promotion of proliferation and metastases in ovarian
cancer: Potential role as a biomarker. Life Sci. 173:62–67. 2017.
View Article : Google Scholar
|
|
22
|
Zheng Y, Yin L, Chen H, Yang S, Pan C, Lu
S, Miao M and Jiao B: miR-376a suppresses proliferation and induces
apoptosis in hepatocellular carcinoma. FEBS Lett. 586:2396–2403.
2012. View Article : Google Scholar
|
|
23
|
Zheng Y, Chen H, Yin M, Ye X, Chen G, Zhou
X, Yin L, Zhang C and Ding B: MiR-376a and histone deacetylation 9
form a regulatory circuitry in hepatocellular carcinoma. Cell
Physiol Biochem. 35:729–739. 2015. View Article : Google Scholar
|
|
24
|
Guan H, Cai J, Zhang N, Wu J, Yuan J, Li J
and Li M: Sp1 is upregulated in human glioma, promotes
MMP-2-mediated cell invasion and predicts poor clinical outcome.
Int J Cancer. 130:593–601. 2012. View Article : Google Scholar
|
|
25
|
Luo J, Wang X, Xia Z, Yang L, Ding Z, Chen
S, Lai B and Zhang N: Transcriptional factor specificity protein 1
(SP1) promotes the proliferation of glioma cells by up-regulating
midkine (MDK). Mol Biol Cell. 26:430–439. 2015. View Article : Google Scholar :
|
|
26
|
Dong Q, Cai N, Tao T, Zhang R, Yan W, Li
R, Zhang J, Luo H, Shi Y, Luan W, et al: An axis involving SNAI1,
microRNA-128 and SP1 modulates glioma progression. PLoS One.
9:e986512014. View Article : Google Scholar :
|
|
27
|
Lee WS, Kwon J, Yun DH, Lee YN, Woo EY,
Park MJ, Lee JS, Han YH and Bae IH: Specificity protein 1
expression contributes to Bcl-w-induced aggressiveness in
glioblastoma multiforme. Mol Cells. 37:17–23. 2014. View Article : Google Scholar :
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Zhou X, Ren Y, Moore L, Mei M, You Y, Xu
P, Wang B, Wang G, Jia Z, Pu P, et al: Downregulation of miR-21
inhibits EGFR pathway and suppresses the growth of human
glioblastoma cells independent of PTEN status. Lab Invest.
90:144–155. 2010. View Article : Google Scholar
|
|
30
|
Ma R, Yan W, Zhang G, Lv H, Liu Z, Fang F,
Zhang W, Zhang J, Tao T, You Y, et al: Upregulation of miR-196b
confers a poor prognosis in glioblastoma patients via inducing a
proliferative phenotype. PLoS One. 7:e380962012. View Article : Google Scholar :
|
|
31
|
Qu S, Yao Y, Shang C, Xue Y, Ma J, Li Z
and Liu Y: MicroRNA-330 is an oncogenic factor in glioblastoma
cells by regulating SH3GL2 gene. PLoS One. 7:e460102012. View Article : Google Scholar :
|
|
32
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar
|
|
33
|
Meng X, Joosse SA, Müler V, Trillsch F,
Milde-Langosch K, Mahner S, Geffken M, Pantel K and Schwarzenbach
H: Diagnostic and prognostic potential of serum miR-7, miR-16,
miR-25, miR-93, miR-182, miR-376a and miR-429 in ovarian cancer
patients. Br J Cancer. 113:1358–1366. 2015. View Article : Google Scholar :
|
|
34
|
Formosa A, Markert EK, Lena AM, Italiano
D, Finazzi-Agro' E, Levine AJ, Bernardini S, Garabadgiu AV, Melino
G and Candi E: MicroRNAs, miR-154, miR-299-5p, miR-376a, miR-376c,
miR-377, miR-381, miR-487b, miR-485-3p, miR-495 and miR-654-3p,
mapped to the 14q32.31 locus, regulate proliferation, apoptosis,
migration and invasion in metastatic prostate cancer cells.
Oncogene. 33:5173–5182. 2014. View Article : Google Scholar
|
|
35
|
Fellenberg J, Sähr H, Kunz P, Zhao Z, Liu
L, Tichy D and Herr I: Restoration of miR-127-3p and miR-376a-3p
counteracts the neoplastic phenotype of giant cell tumor of bone
derived stromal cells by targeting COA1, GLE1 and PDIA6. Cancer
Lett. 371:134–141. 2016. View Article : Google Scholar
|
|
36
|
Yu Z, Ni L, Chen D, Zhang Q, Su Z, Wang Y,
Yu W, Wu X, Ye J, Yang S, et al: Identification of miR-7 as an
oncogene in renal cell carcinoma. J Mol Histol. 44:669–677. 2013.
View Article : Google Scholar
|
|
37
|
Zehavi L, Avraham R, Barzilai A, Bar-Ilan
D, Navon R, Sidi Y, Avni D and Leibowitz-Amit R: Silencing of a
large microRNA cluster on human chromosome 14q32 in melanoma:
Biological effects of mir-376a and mir-376c on insulin growth
factor 1 receptor. Mol Cancer. 11:442012. View Article : Google Scholar :
|
|
38
|
Kanai M, Wei D, Li Q, Jia Z, Ajani J, Le
X, Yao J and Xie K: Loss of Krüppel-like factor 4 expression
contributes to Sp1 overexpression and human gastric cancer
development and progression. Clin Cancer Res. 12:6395–6402. 2006.
View Article : Google Scholar
|
|
39
|
Hosoi Y, Watanabe T, Nakagawa K, Matsumoto
Y, Enomoto A, Morita A, Nagawa H and Suzuki N: Up-regulation of
DNA-dependent protein kinase activity and Sp1 in colorectal cancer.
Int J Oncol. 25:461–468. 2004.
|
|
40
|
Yin P, Zhao C, Li Z, Mei C, Yao W, Liu Y,
Li N, Qi J, Wang L, Shi Y, et al: Sp1 is involved in regulation of
cystathionine γ-lyase gene expression and biological function by
PI3K/Akt pathway in human hepatocellular carcinoma cell lines. Cell
Signal. 24:1229–1240. 2012. View Article : Google Scholar
|
|
41
|
Sankpal UT, Goodison S, Abdelrahim M and
Basha R: Targeting Sp1 transcription factors in prostate cancer
therapy. Med Chem. 7:518–525. 2011. View Article : Google Scholar
|
|
42
|
Chiefari E, Brunetti A, Arturi F, Bidart
JM, Russo D, Schlumberger M and Filetti S: Increased expression of
AP2 and Sp1 transcription factors in human thyroid tumors: A role
in NIS expression regulation? BMC Cancer. 2:352002. View Article : Google Scholar :
|
|
43
|
Beishline K and Azizkhan-Clifford J: Sp1
and the ‘hallmarks of cancer’. FEBS J. 282:224–258. 2015.
View Article : Google Scholar
|
|
44
|
Wang YT, Yang WB, Chang WC and Hung JJ:
Interplay of posttranslational modifications in Sp1 mediates Sp1
stability during cell cycle progression. J Mol Biol. 414:1–14.
2011. View Article : Google Scholar
|
|
45
|
Yen WH, Ke WS, Hung JJ, Chen TM, Chen JS
and Sun HS: Sp1-mediated ectopic expression of T-cell lymphoma
invasion and metastasis 2 in hepatocellular carcinoma. Cancer Med.
5:465–477. 2016. View
Article : Google Scholar :
|